
Galactitol Transport Factor GatA Relieves ATP Supply Restriction to Enhance Acid Tolerance of Escherichia coli in the Two-Stage Fermentation Production of D-Lactate

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Strains, Plasmids, and Culture Conditions
2.2. Growth Curve Assays
2.3. Stress-Tolerance Assays
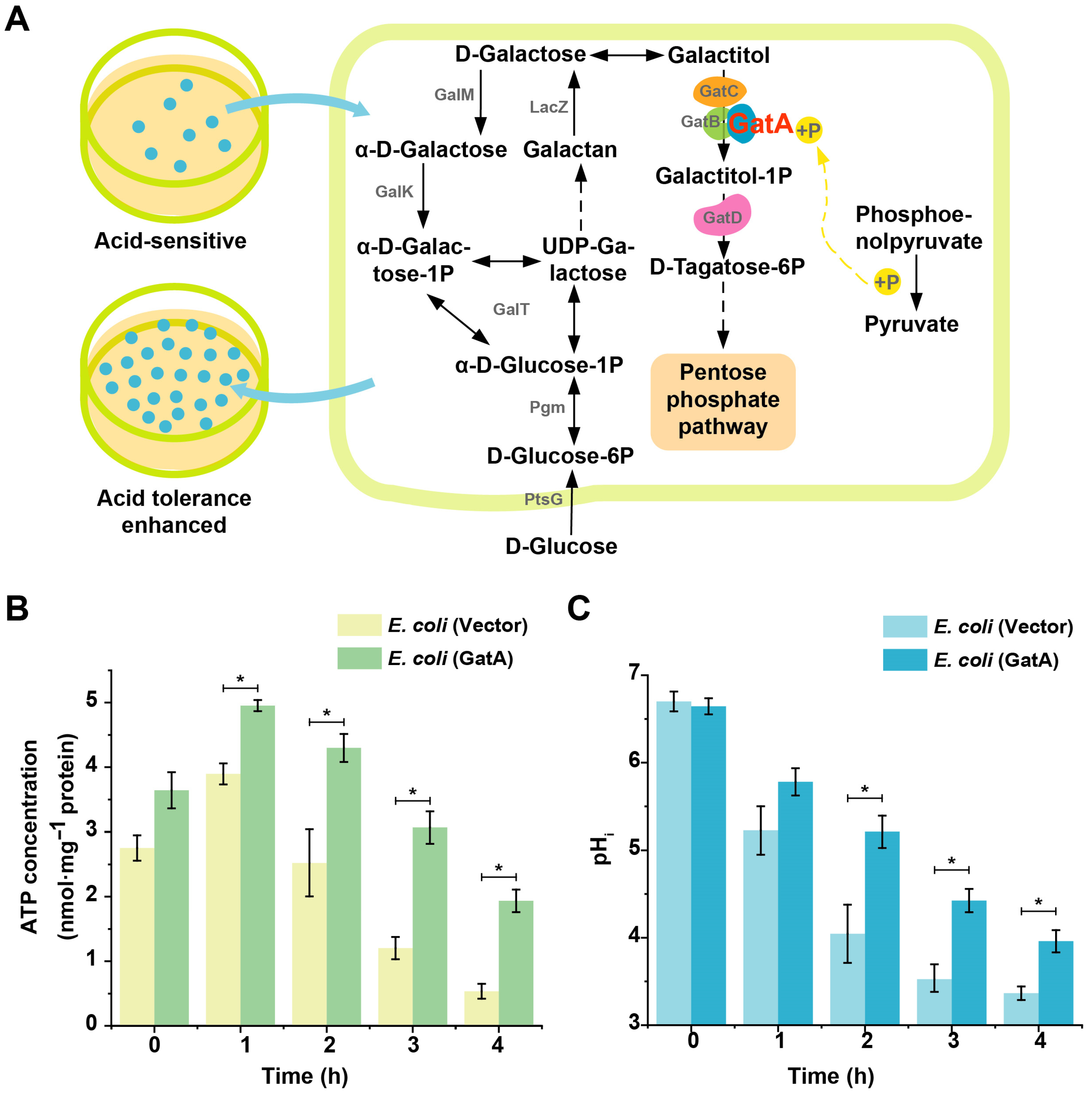
2.4. Intracellular ATP Detection
2.5. Intracellular pH Detection
2.6. Fermentation Analysis
2.7. Metabolite Detection
2.8. Statistical Analysis
3. Results and Discussion
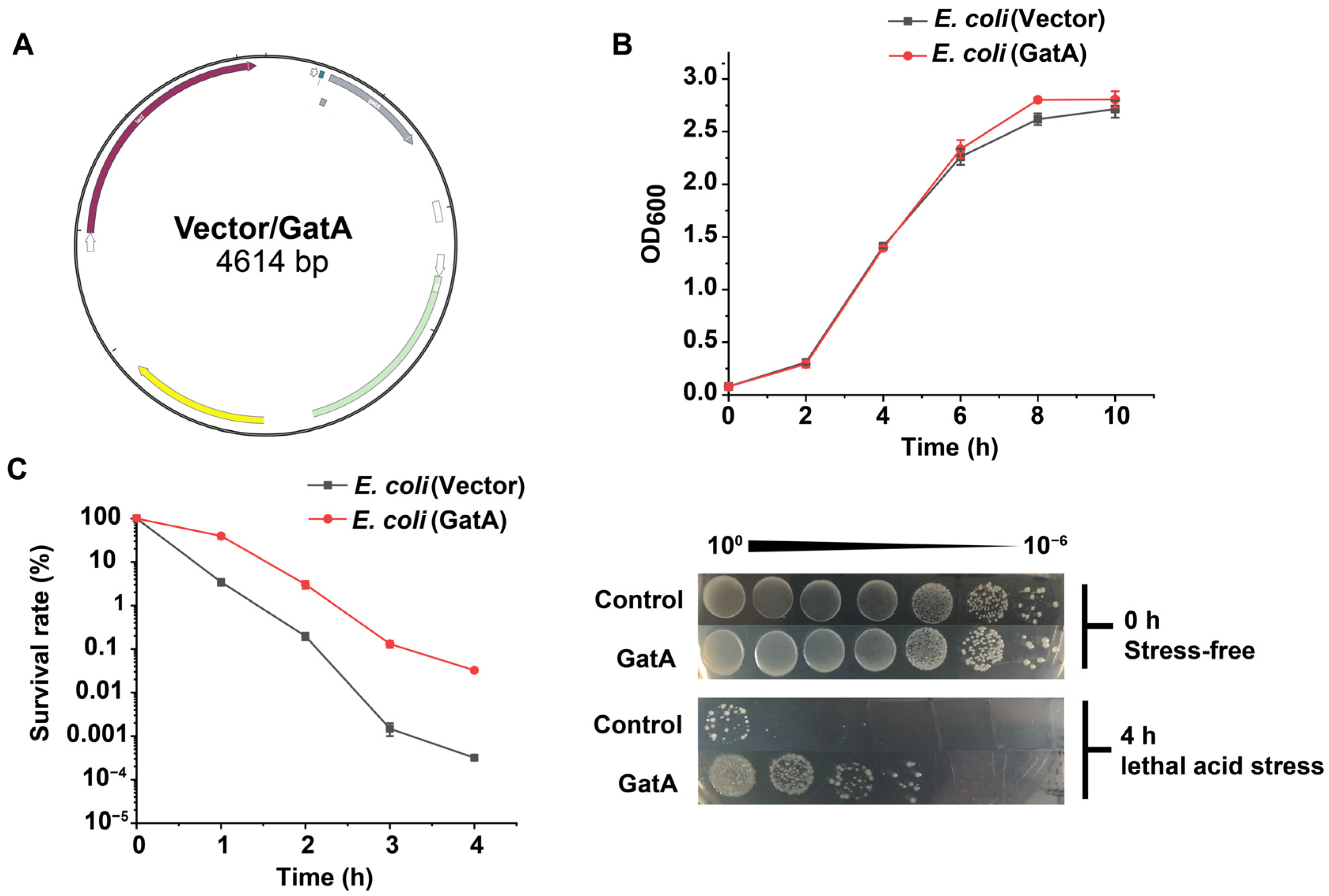
3.1. Identification and Functional Verification of GatA
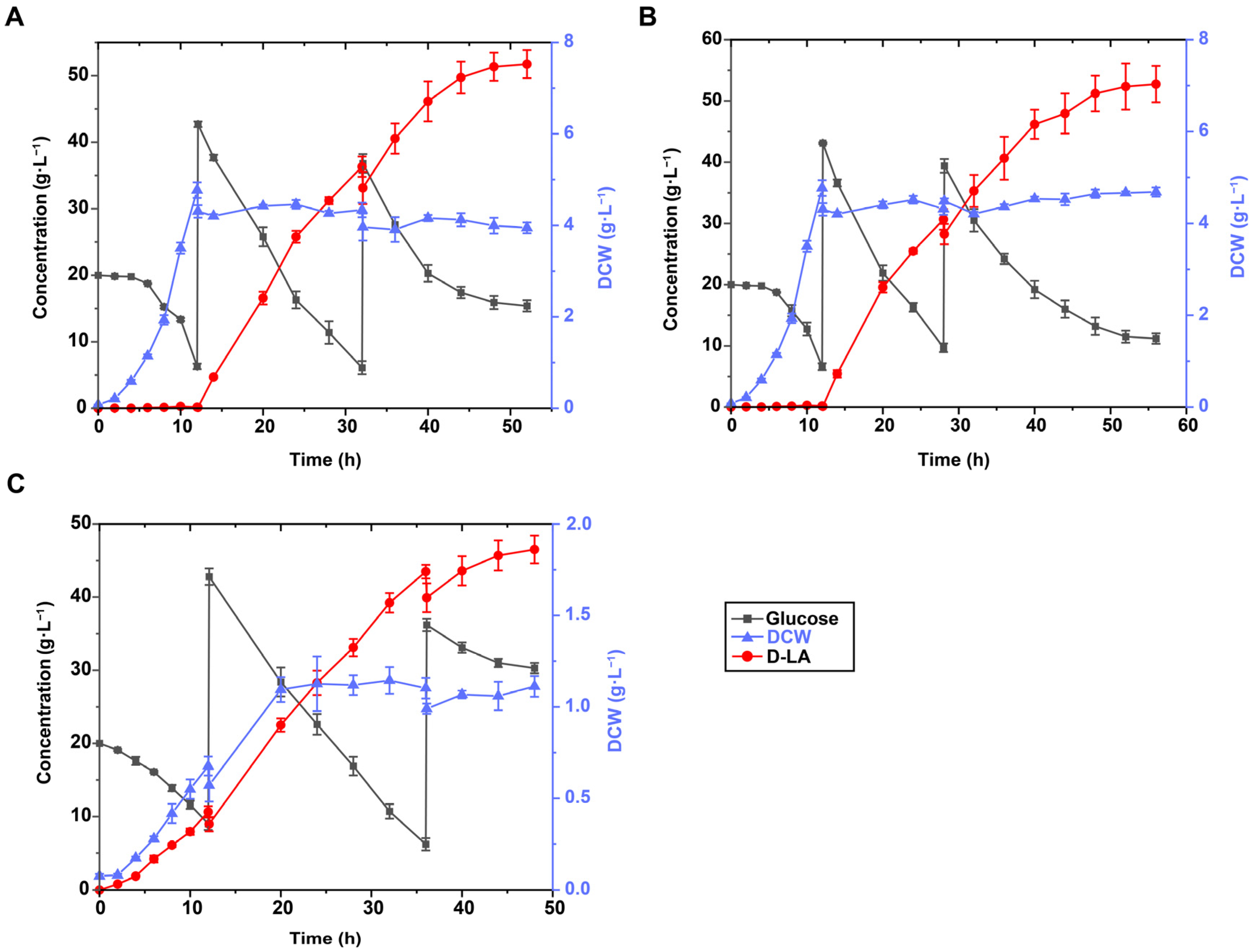
3.2. Aerobic Growth and Oxygen-Limited Fermentation Pattern Facilitate D-LA Production
3.3. Effect of Biomass at the Transition Point between Growth and Acid Production Stages
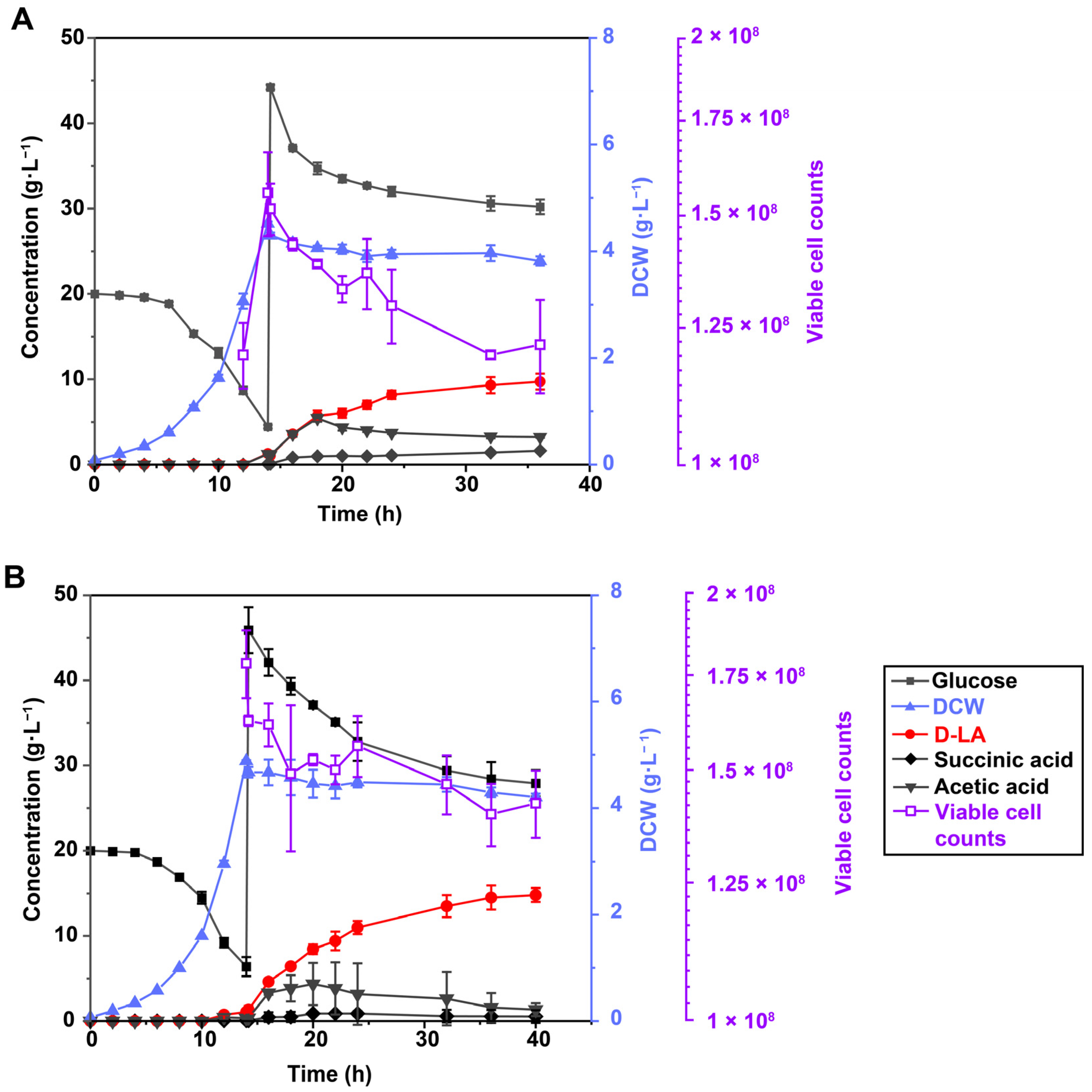
3.4. Inductional Effect on D-LA Synthesis at Different Growth Stages
3.5. LBBE317PGA Enhanced D-LA Production under Low pH Incubation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shi, Y.H.; Sun, H.Y.; Lu, D.M.; Le, Q.; Chen, D.; Zhou, Y. Separation of glycolic acid from glycolonitrile hydrolysate by reactive extraction with tri-n-octylamine. Sep. Purif. Technol. 2006, 49, 20–26. [Google Scholar]
- Ahn, J.H.; Jang, Y.S.; Lee, S.Y. Production of succinic acid by metabolically engineered microorganisms. Curr. Opin. Biotechnol. 2016, 42, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, J.; Zhu, Z.; Du, G. The challenges and prospects of Escherichia coli as an organic acid production host under acid stress. Appl. Microbiol. Biot. 2021, 105, 8091–8107. [Google Scholar] [CrossRef]
- Datta, R.; Henry, M. Lactic acid: Recent advances in products, processes and technologies—A review. J. Chem. Technol. Biot. 2006, 81, 1119–1129. [Google Scholar] [CrossRef]
- Tsuji, F. Autocatalytic hydrolysis of amorphous-made polylactides: Effects of L-lactide content, tacticity, and enantiomeric polymer blending. Polymer 2002, 43, 1789–1796. [Google Scholar] [CrossRef]
- Utrilla, J.; Gosset, G.; Martinez, A. ATP limitation in a pyruvate formate lyase mutant of Escherichia coli MG1655 increases glycolytic flux to d-lactate. J. Ind. Microbiol. Biot. 2009, 36, 1057–1062. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, W.; Zhao, X.; Wang, J.H.; Garza, E.; Manow, R.; Zhou, S.D. Pilot scale demonstration of D-lactic acid fermentation facilitated by Ca(OH)(2) using a metabolically engineered Escherichia coli. Bioresour. Technol. 2014, 169, 559–565. [Google Scholar] [CrossRef]
- Zhou, L.; Zuo, Z.R.; Chen, X.Z.; Niu, D.D.; Tian, K.M.; Prior, B.A.; Shen, W.; Shi, G.Y.; Singh, S.; Wang, Z.X. Evaluation of Genetic Manipulation Strategies on d-Lactate Production by Escherichia coli. Curr. Microbiol. 2011, 62, 981–989. [Google Scholar] [CrossRef]
- Zhou, L.; Tian, K.M.; Niu, D.D.; Shen, W.; Shi, G.Y.; Singh, S.; Wang, Z.X. Improvement of d-lactate productivity in recombinant Escherichia coli by coupling production with growth. Biotechnol. Lett. 2012, 34, 1123–1130. [Google Scholar] [CrossRef]
- Olson, E.R. Influence of Ph on Bacterial Gene-Expression. Mol. Microbiol. 1993, 8, 5–14. [Google Scholar] [CrossRef]
- Zhong, W.; Yang, M.H.; Hao, X.M.; Sharshar, M.M.; Wang, Q.H.; Xing, J.M. Improvement of D-lactic acid production at low pH through expressing acid-resistant gene IoGAS1 in engineered Saccharomyces cerevisiae. J. Chem. Technol. Biot. 2021, 96, 732–742. [Google Scholar] [CrossRef]
- Valli, M.; Sauer, M.; Branduardi, P.; Borth, N.; Porro, D.; Mattanovich, D. Improvement of lactic acid production in Saccharomyces cerevisiae by cell sorting for high intracellular pH. Appl. Environ. Microb. 2006, 72, 5492–5499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piper, P.; Calderon, C.O.; Hatzixanthis, K.; Mollapour, M. Weak acid adaptation: The stress response that confers yeasts with resistance to organic acid food preservatives. Microbiology 2001, 147, 2635–2642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, P.L.; Ma, D.; Chen, Y.L.; Guo, Y.Y.; Chen, G.Q.; Deng, H.T.; Shi, Y.G. L-glutamine provides acid resistance for Escherichia coli through enzymatic release of ammonia. Cell Res. 2013, 23, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Roe, A.J.; O’Byrne, C.; McLaggan, D.; Booth, I.R. Inhibition of Escherichia coli growth by acetic acid: A problem with methionine biosynthesis and homocysteine toxicity. Microbiology 2002, 148, 2215–2222. [Google Scholar] [CrossRef] [Green Version]
- Singhvi, M.; Zendo, T.; Sonomoto, K. Free lactic acid production under acidic conditions by lactic acid bacteria strains: Challenges and future prospects. Appl. Microbiol. Biot. 2018, 102, 5911–5924. [Google Scholar] [CrossRef]
- Fletcher, E.; Feizi, A.; Bisschops, M.M.M.; Hallstrom, B.M.; Khoomrung, S.; Siewers, V.; Nielsen, J. Evolutionary engineering reveals divergent paths when yeast is adapted to different acidic environments. Metab. Eng. 2017, 39, 19–28. [Google Scholar] [CrossRef]
- Fu, X.M.; Wang, Y.X.; Wang, J.H.; Garza, E.; Manow, R.; Zhou, S.D. Semi-industrial scale (30 m(3)) fed-batch fermentation for the production of D-lactate by Escherichia coli strain HBUT-D15. J. Ind. Microbiol. Biot. 2017, 44, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Kim, S.; Kim, W.; Kim, S.; Cha, S.; Moon, H.; Hur, D.H.; Kim, S.Y.; Na, J.G.; Lee, J.W.; et al. Efficient production of d-lactate from methane in a lactate-tolerant strain of Methylomonas sp. DH-1 generated by adaptive laboratory evolution. Biotechnol. Biofuels 2019, 12, 234. [Google Scholar] [CrossRef] [Green Version]
- Park, H.J.; Bae, J.H.; Ko, H.J.; Lee, S.H.; Sung, B.H.; Han, J.I.; Sohn, J.H. Low-pH production of d-lactic acid using newly isolated acid tolerant yeast Pichia kudriavzevii NG7. Biotechnol. Bioeng. 2018, 115, 2232–2242. [Google Scholar] [CrossRef]
- Zheng, H.; Gong, J.; Chen, T.; Chen, X.; Zhao, X. Strain improvement of Sporolactobacillus inulinus ATCC 15538 for acid tolerance and production of D-lactic acid by genome shuffling. Appl. Microbiol. Biotechnol. 2010, 85, 1541–1549. [Google Scholar] [CrossRef]
- Ma, J.F.; Wu, M.K.; Zhang, C.Q.; He, A.Y.; Kong, X.P.; Li, G.L.; Wei, C.; Jiang, M. Coupled ARTP and ALE strategy to improve anaerobic cell growth and succinic acid production by Escherichia coli. J. Chem. Technol. Biot. 2016, 91, 711–717. [Google Scholar] [CrossRef]
- Nguyen-Vo, T.P.; Liang, Y.; Sankaranarayanan, M.; Seol, E.; Chun, A.Y.; Ashok, S.; Chauhan, A.S.; Kim, J.R.; Park, S. Development of 3-hydroxypropionic-acid-tolerant strain of Escherichia coli W and role of minor global regulator yieP. Metab. Eng. 2019, 53, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Li, X.; Guo, S.; Lemma, W.D.; Zhang, W.; Ma, J.; Jia, H.; Wu, H.; Jiang, M.; Ouyang, P. Enhanced succinic acid production under acidic conditions by introduction of glutamate decarboxylase system in E. coli AFP111. Bioprocess Biosyst. Eng. 2017, 40, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, J.; Zhu, Z.; Jiang, X.; Zheng, T.; Du, G. Revealing novel synergistic defense and acid tolerant performance of Escherichia coli in response to organic acid stimulation. Appl. Microbiol. Biotechnol. 2022, 106, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Peng, Z.; Zhu, Q.; Zhang, J.; Du, G. [NiFe] Hydrogenase Accessory Proteins HypB-HypC Accelerate Proton Conversion to Enhance the Acid Resistance and d-Lactic Acid Production of Escherichia coli. ACS Synth. Biol. 2022, 11, 1521–1530. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.M.; Yang, J.H.; Yang, P.S.; Wu, Z.M.; Zhang, J.; Du, G.C. Enhanced acid-stress tolerance in Lactococcus lactis NZ9000 by overexpression of ABC transporters. Microb. Cell Factories 2019, 18, 1–14. [Google Scholar] [CrossRef]
- Guan, N.Z.; Liu, L.; Shin, H.D.; Chen, R.R.; Zhang, J.; Li, J.H.; Du, G.C.; Shi, Z.P.; Chen, J. Systems-level understanding of how Propionibacterium acidipropionici respond to propionic acid stress at the microenvironment levels: Mechanism and application. J. Biotechnol. 2013, 167, 56–63. [Google Scholar] [CrossRef]
- Zhu, Z.; Ji, X.; Wu, Z.; Zhang, J.; Du, G. Improved acid-stress tolerance of Lactococcus lactis NZ9000 and Escherichia coli BL21 by overexpression of the anti-acid component recT. J. Ind. Microbiol. Biotechnol. 2018, 45, 1091–1101. [Google Scholar] [CrossRef]
- Nobelmann, B.; Lengeler, J.W. Molecular analysis of the gat genes from Escherichia coli and of their roles in galactitol transport and metabolism. J. Bacteriol. 1996, 178, 6790–6795. [Google Scholar] [CrossRef] [Green Version]
- Saier, M.H.; Hvorup, R.N.; Barabote, R.D. Evolution of the bacterial phosphotransferase system: From carriers and enzymes to group translocators. Biochem. Soc. Trans. 2005, 33, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Volpon, L.; Young, C.R.; Matte, A.; Gehring, K. NMR structure of the enzyme GatB of the galactitol-specific phosphoenolpyruvate-dependent phosphotransferase system and its interaction with GatA. Protein Sci. 2006, 15, 2435–2441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Utrilla, J.; Licona-Cassani, C.; Marcellin, E.; Gosset, G.; Nielsen, L.K.; Martinez, A. Engineering and adaptive evolution of Escherichia coli for D-lactate fermentation reveals GatC as a xylose transporter. Metab. Eng. 2012, 14, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.D.; Hill, C. Surviving the acid test: Responses of gram-positive bacteria to low pH. Microbiol. Mol. Biol. R. 2003, 67, 429–453. [Google Scholar] [CrossRef] [Green Version]
- Baek, S.H.; Kwon, E.Y.; Kim, S.Y.; Hahn, J.S. GSF2 deletion increases lactic acid production by alleviating glucose repression in Saccharomyces cerevisiae. Sci. Rep. 2016, 6, 34812. [Google Scholar] [CrossRef] [Green Version]
- Guan, N.Z.; Shin, H.D.; Chen, R.R.; Li, J.H.; Liu, L.; Du, G.C.; Chen, J. Understanding of how Propionibacterium acidipropionici respond to propionic acid stress at the level of proteomics. Sci. Rep. 2014, 4, 6951. [Google Scholar] [CrossRef] [Green Version]
- Luedeking, R.; Piret, E.L. A kinetic study of the lactic acid fermentation. Batch process at controlled pH (Reprinted from Journal of Biochemical and Microbiological Technology and Engineering, vol 1, pg 393, 1959). Biotechnol. Bioeng. 2000, 67, 636–644. [Google Scholar] [CrossRef]
- Yao, P.; You, S.P.; Qi, W.; Su, R.X.; He, Z.M. Investigation of fermentation conditions of biodiesel by-products for high production of beta-farnesene by an engineered Escherichia coli. Environ. Sci. Pollut. R. 2020, 27, 22758–22769. [Google Scholar] [CrossRef]
- Lalwani, M.A.; Ip, S.S.; Carrasco-Lopez, C.; Day, C.; Zhao, E.M.; Kawabe, H.; Avalos, J.L. Optogenetic control of the lac operon for bacterial chemical and protein production. Nat. Chem. Biol. 2021, 17, 71–79. [Google Scholar] [CrossRef]
- Lopes, C.; dos Santos, N.V.; Dupont, J.; Pedrolli, D.B.; Valentini, S.R.; Santos-Ebinuma, V.D.; Pereira, J.F.B. Improving the cost effectiveness of enhanced green fluorescent protein production using recombinant Escherichia coli BL21 (DE3): Decreasing the expression inducer concentration. Biotechnol. Appl. Bioc. 2019, 66, 527–536. [Google Scholar] [CrossRef]
- Omoya, K.; Kato, Z.; Matsukuma, E.; Li, A.L.; Hashimoto, K.; Yamamoto, Y.; Ohnishi, H.; Kondo, N. Systematic optimization of active protein expression using GFP as a folding reporter. Protein Expr. Purif. 2004, 36, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Sandoval, M.T.; Galindez-Mayer, J.; Moss-Acosta, C.L.; Gosset, G.; Martinez, A. Volumetric oxygen transfer coefficient as a means of improving volumetric ethanol productivity and a criterion for scaling up ethanol production with Escherichia coli. J. Chem. Technol. Biot. 2017, 92, 981–989. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Peng, Z.; Ji, X.; Zhang, J.; Du, G. Galactitol Transport Factor GatA Relieves ATP Supply Restriction to Enhance Acid Tolerance of Escherichia coli in the Two-Stage Fermentation Production of D-Lactate. Fermentation 2022, 8, 665. https://doi.org/10.3390/fermentation8120665
Yang J, Peng Z, Ji X, Zhang J, Du G. Galactitol Transport Factor GatA Relieves ATP Supply Restriction to Enhance Acid Tolerance of Escherichia coli in the Two-Stage Fermentation Production of D-Lactate. Fermentation. 2022; 8(12):665. https://doi.org/10.3390/fermentation8120665
Chicago/Turabian StyleYang, Jinhua, Zheng Peng, Xiaomei Ji, Juan Zhang, and Guocheng Du. 2022. "Galactitol Transport Factor GatA Relieves ATP Supply Restriction to Enhance Acid Tolerance of Escherichia coli in the Two-Stage Fermentation Production of D-Lactate" Fermentation 8, no. 12: 665. https://doi.org/10.3390/fermentation8120665