
Optimization of Culture Conditions and Wheat Bran Class Selection in the Production of Bacillus thuringiensis-Based Biopesticides

 ,
,  and
and
Abstract
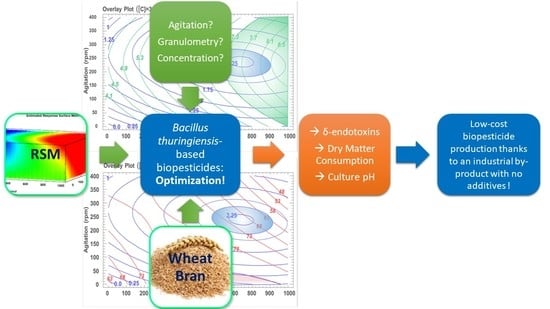
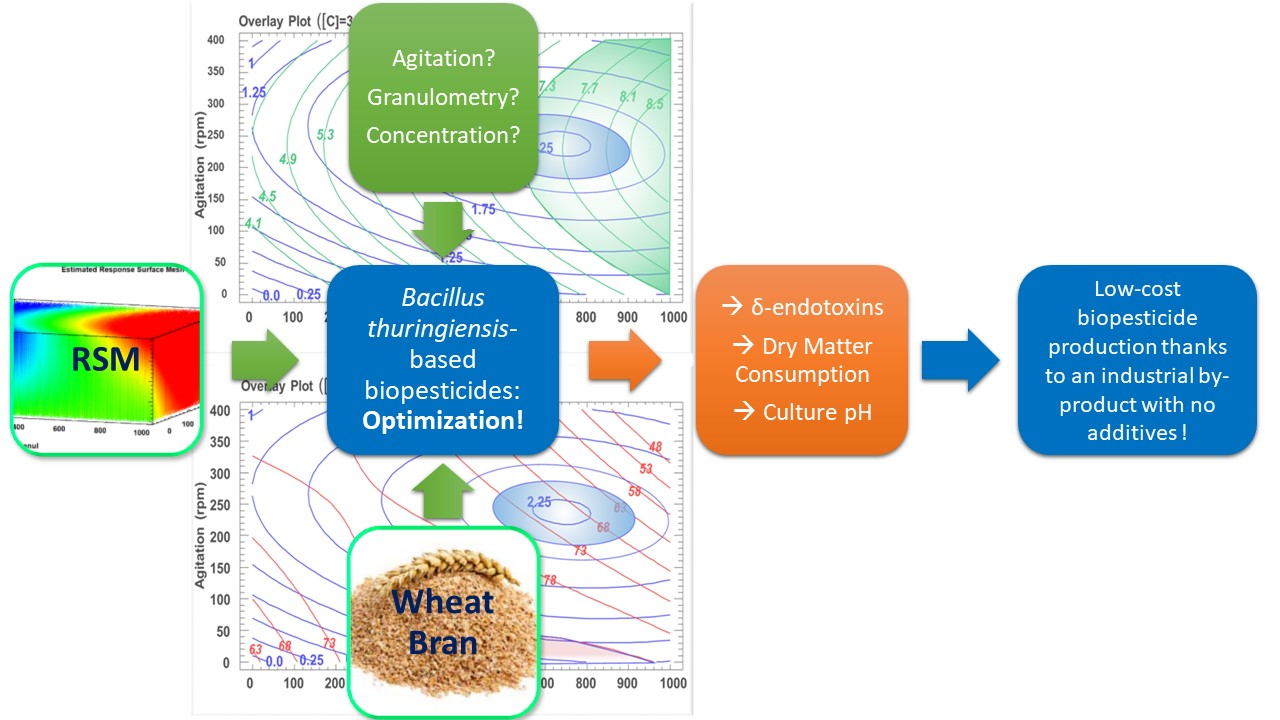
:
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Production Culture Media
2.3. Culture Conditions
2.4. Analytical Methods/Analyzed Parameters
2.4.1. δ-Endotoxins Concentration
2.4.2. pH
2.4.3. Dry-Matter Consumption (DMC) Quantification
2.5. Response Surface Methodology (RSM)
3. Results and Discussion
3.1. RSM Results in WB Media
3.1.1. Response Parameter: δ-Endotoxins Concentration
3.1.2. Response Parameter: Culture pH
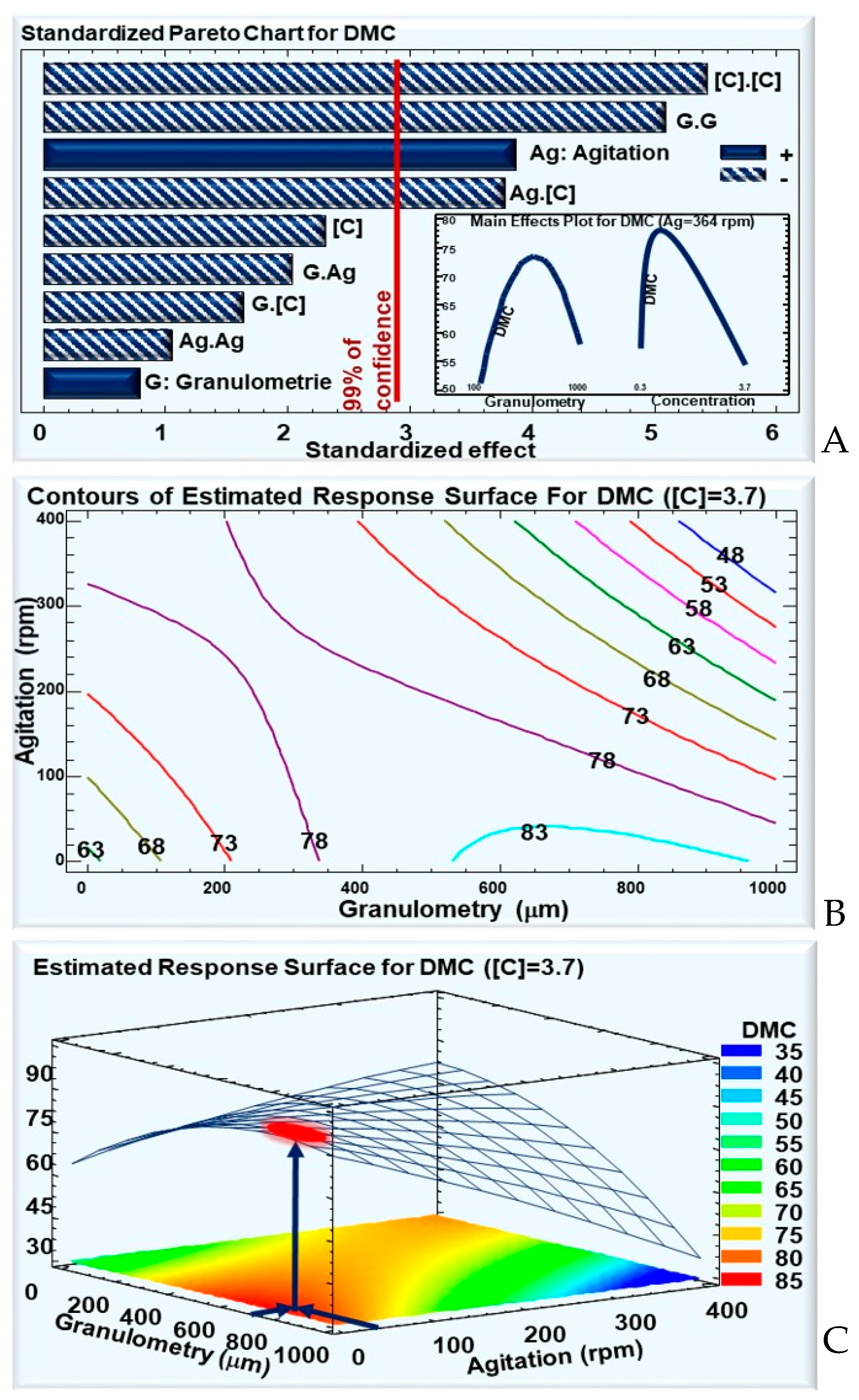
3.1.3. Response Parameter: Dry-Matter Consumption Percentage
3.2. Optimum Parameters Selection
4. Conclusions
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aktar, W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilliom, R.J. Pesticides in U.S. streams and groundwater. Environ. Sci. Technol. 2007, 41, 3408–3414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topaz, T.; Egozi, R.; Suari, Y.; Ben-Ari, J.; Sade, T.; Chefetz, B.; Yahel, G. Environmental risk dynamics of pesticides toxicity in a Mediterranean micro-estuary. Environ. Pollut. 2020, 265, 114941. [Google Scholar] [CrossRef] [PubMed]
- Corsini, E.; Sokooti, M.; Galli, C.L.; Moretto, A.; Colosio, C. Pesticide induced immunotoxicity in humans: A comprehensive review of the existing evidence. Toxicology 2013, 307, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.G.; Giordano, G.; Guizzetti, M.; Vitalone, A. Neurotoxicity of pesticides: A brief review. Front. Biosci. 2008, 13, 1240–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abou Zeid, M.I.; Jammoul, A.M.; Melki, K.C.; Jawdah, Y.A.; Kallassy Awad, M. Suggested policy and legislation reforms to reduce deleterious effect of pesticides in Lebanon. Heliyon 2020, 6, e05524. [Google Scholar] [CrossRef]
- Intelligence, M. Biopesticides Marke—Growth, Trends, COVID-19 Impact, and Forecasts (2021–2026). 2020. Available online: https://www.mordorintelligence.com/industry-reports/global-biopesticides-market-industry (accessed on 3 August 2022).
- Kumar, J.; Ramlal, A.; Mallick, D.; Mishra, V. An Overview of Some Biopesticides and Their Importance in Plant Protection for Commercial Acceptance. Plants 2021, 10, 1185. [Google Scholar] [CrossRef] [PubMed]
- Damalas, C.A.; Koutroubas, S.D. Current status and recent developments in biopesticide use. Agriculture 2018, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Singh, A. Biopesticides: Present Status and the Future Prospects. J. Fertil. Pestic. 2015, 6, 2. [Google Scholar] [CrossRef]
- Soberón, M.; Monnerat, R.; Bravo, A. Mode of Action of Cry Toxins from Bacillus thuringiensis and Resistance Mechanisms. In Microbial Toxins—Toxinology; Gopalakrishnakone, P., Stiles, B., Alape-Girón, A., Dubreuil, J., Mandal, M., Eds.; Springer: Dordrecht, The Netherlands, 2018; pp. 15–27. [Google Scholar] [CrossRef]
- Palma, L.; Muñoz, D.; Berry, C.; Murillo, J.; Caballero, P. Bacillus thuringiensis Toxins: An Overview of Their Biocidal Activity. Toxins 2014, 6, 3296–3325. [Google Scholar] [CrossRef] [Green Version]
- Fayad, N.; Patiño-Navarrete, R.; Kambris, Z.; Antoun, M.; Osta, M.; Chopineau, J.; Mahillon, J.; El Chamy, L.; Sanchis, V.; Kallassy Awad, M. Characterization and Whole Genome Sequencing of AR23, a Highly Toxic Bacillus thuringiensis Strain Isolated from Lebanese Soil. Curr. Microbiol. 2019, 76, 1503–1511. [Google Scholar] [CrossRef] [PubMed]
- Fayad, N.; Kambris, Z.; El Chamy, L.; Mahillon, J.; Kallassy Awad, M. A novel anti-dipteran Bacillus thuringiensis strain: Unusual Cry toxin genes in a highly dynamic plasmid environment. Appl. Environ. Microbiol. 2020, 87, e02294-20. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, M.; Azzouz, H.; Chavanieu, A.; Abdelmalak, N.; Chopineau, J.; Kallassy Awad, M. Isolation and characterization of a new Bacillus thuringiensis strain Lip harboring a new cry1Aa gene highly toxic to Ephestia kuehniella (Lepidoptera: Pyralidae) larvae. Arch. Microbiol. 2014, 196, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Day, M.; Ibrahim, M.; Dyer, D.; Bulla, L. Genome sequence of Bacillus thuringiensis subsp. kurstaki strain HD-1. Genome Announc. 2014, 2, 613–627. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Song, L.; Shu, C.; Wang, P.; Deng, C.; Peng, Q.; Lereclus, D.; Wang, X.; Huang, D.; Zhang, J.; et al. Complete Genome Sequence of Bacillus thuringiensis subsp. kurstaki Strain HD73. Genome Announc. 2013, 1, 2–3. [Google Scholar] [CrossRef] [Green Version]
- Qiu, N.; He, J.; Wang, Y.; Cheng, G.; Li, M.; Sun, M.; Yu, Z. Prevalence and diversity of insertion sequences in the genome of Bacillus thuringiensis YBT-1520 and comparison with other Bacillus cereus group members. FEMS Microbiol. Lett. 2010, 310, 9–16. [Google Scholar] [CrossRef]
- Saadaoui, I.; Rouis, S.; Jaoua, S. A new Tunisian strain of Bacillus thuringiensis kurstaki having high insecticidal activity and δ-endotoxin yield. Arch. Microbiol. 2009, 191, 341–348. [Google Scholar] [CrossRef]
- Devi, P.S.V.; Ravinder, T.; Jaidev, C. Cost-effective production of Bacillus thuringiensis by solid-state fermentation. J. Invertebr. Pathol. 2005, 88, 163–168. [Google Scholar] [CrossRef]
- Ndao, A. Mise a L’Echelle de la Production du Biopesticide Bacillus thuringiensis Var. Kurstaki Avec Comme Substrat des Eaux Usées d’Amidon; Université du Québec: Quebec City, QC, Canada, 2014. [Google Scholar]
- Nickerson, K.W.; Bulla, L.A. Physiology of Spore-forming Bacteria Associated with Insects: Minimal Nutritional Requirements for Growth, Sporulation, and Parasporal Crystal Formation of Bacillus thuringiensis. Appl. Microbiol. 1974, 28, 124–128. [Google Scholar] [CrossRef]
- Montiel, M.D.; Tyagi, R.D.; Valero, J.R. Wastewater treatment sludge as a raw material for the production of Bacillus thuringiensis based biopesticides. Water Res. 2001, 35, 3807–3816. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, M. Antioxidant Properties of Wheat Bran against Oxidative Stress. In Wheat and Rice in Disease Prevention and Health; Elsevier Inc.: Amsterdam, The Netherlands, 2014; pp. 181–199. [Google Scholar] [CrossRef]
- Mounsef, J.R.; Salameh, D.; Kallassy Awad, M.; Lteif, R.; Brandam, C. Evaluation of a cereal milling by-product for the low-cost production of Bacillus thuringiensis kurstaki in submerged fermentation. Eur. J. Biotechnol. Biosci. 2014, 1, 10–16. [Google Scholar]
- Anderson, T.B. Effects of Carbon: Nitrogen Ratio and Oxygen on the Growth Kinetics of Bacillus thuringiensis and Yield of Bioinsecticidal Crystal Protein. Ph.D. Thesis, Faculty of Graduate Studies, University of Western Ontario, London, ON, Canada, 1990. [Google Scholar]
- Kallassy, M.I.; Khoury, M.; Louka, N. New Strain of Bacillus Thuringiensis “Bacillus Thuringiensis Lip MKA” Isolated from Lebanese Soil as Biocontrol Agent (biopesticide) and Design, Construction and Installation of Bioreactors for Large Scale Production and Cultivation of This Strain. LEBANON Patent 2017/11-11298L, 15 November 2017. [Google Scholar]
- Loutfi, H.; Fayad, N.; Pellen, F.; Le Jeune, B.; Chakroun, M.; Benfarhat, D.; Lteif, R.; Kallassy, M.; Le Brun, G.; Abboud, M. Morphological Study of Bacillus thuringiensis Crystals and Spores. Appl Sci. 2021, 11, 155. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Abi-Khattar, A.M.; Rajha, H.N.; Abdel-Massih, R.M.; Habchi, R.; Maroun, R.G.; Debs, E.; Louka, N. “Intensification of Vaporization by Decompression to the Vacuum” (IVDV), a novel technology applied as a pretreatment to improve polyphenols extraction from olive leaves. Food Chem. 2021, 342, 128236. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hameed, A. Stirred tank culture of Bacillus thuringiensis H-14 for production of the mosquitocidal δ-endotoxin: Mathematical modelling and scaling-up studies. World J. Microbiol. Biotechnol. 2001, 17, 857–861. [Google Scholar] [CrossRef]
- Avignone-Rossa, C.; Arcas, J.; Mignone, C. Bacillus thuringiensis growth, sporulation and δ-endotoxin production in oxygen limited and non-limited cultures. World J. Microbiol. Biotechnol. 1992, 8, 301–304. [Google Scholar] [CrossRef] [PubMed]
- Jallouli, W.; Driss, F.; Fillaudeau, L.; Rouis, S. Review on biopesticide production by Bacillus thuringiensis subsp. kurstaki since 1990: Focus on bioprocess parameters. Process. Biochem. 2020, 98, 224–232. [Google Scholar] [CrossRef]
- Içgen, Y.; Içgen, B.; Ozcengiz, G. Regulation of crystal protein biosynthesis by Bacillus thuringiensis: I. Effects of mineral elements and pH. Res. Microbiol. 2002, 153, 599–604. [Google Scholar] [CrossRef]
- Hasanain, A.M. Development of a cheap media for Bacillus thuringiensis growth. Int. J. Biotechnol. Bioeng. 2017, 3, 216–223. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (−α) * | −1 | 0 | +1 | (+α) | |
|---|---|---|---|---|---|
| Granulometry (granule size) (µm) | 97 | 200 | 437 | 675 | 925 |
| Agitation (rpm) | 35.18 | 102 | 200 | 298 | 365 |
| Concentration (g/L) | 6.4 | 20 | 40 | 60 | 73.6 |
| Run | WB Granulometry (G: µm) Real [Coded] Value | Agitation (Ag: rpm) Real [Coded] Value | WB Concentration (C: g/50 mL) Real [Coded] Value | [δ-Endotoxins] (mg/mL) | pH | DMC (%) | |
|---|---|---|---|---|---|---|---|
| Factorial Points | 1 | 200 [−1.0] | 102 [−1.0] | 1 [−1.0] | 0.63 | 6.67 | 62.08 |
| 2 | 675 [1.0] | 102 [−1.0] | 1 [−1.0] | 0.26 | 7.50 | 55.63 | |
| 3 | 200 [−1.0] | 298 [1.0] | 1 [−1.0] | 0.71 | 6.66 | 60.31 | |
| 4 | 675 [1.0] | 298 [1.0] | 1 [−1.0] | 0.45 | 7.32 | 53.54 | |
| 5 | 200 [−1.0] | 102 [−1.0] | 3 [1.0] | 0.43 | 5.98 | 43.82 | |
| 6 | 675 [1.0] | 102 [−1.0] | 3 [1.0] | 1.19 | 7.70 | 53.61 | |
| 7 | 200 [−1.0] | 298 [1.0] | 3 [1.0] | 0.88 | 5.86 | 48.96 | |
| 8 | 675 [1.0] | 298 [1.0] | 3 [1.0] | 1.15 | 7.66 | 53.19 | |
| Axial (star) Points | 9 | 97.5 [−α] | 200 [0.0] | 2 [0.0] | 1.64 | 6.26 | 65.94 |
| 10 | 925 [+α] | 200 [0.0] | 2 [0.0] | 0.41 | 7.98 | 36.82 | |
| 11 | 437 [0] | 35.18 [−α] | 2 [0.0] | 0.07 | 5.54 | 56.20 | |
| 12 | 437 [0] | 365 [+α] | 2 [0.0] | 1.22 | 7.01 | 57.19 | |
| 13 | 437 [0] | 200 [0.0] | 0.32 [−α] | 0.04 | 7.83 | 65.01 | |
| 14 | 437 [0] | 200 [0.0] | 3.68 [+α] | 1.93 | 6.46 | 43.84 | |
| Center Points | 15 | 437 [0] | 200 [0.0] | 2 [0.0] | 1.03 | 7.08 | 57.34 |
| 16 | 437 [0] | 200 [0.0] | 2 [0.0] | 1.14 | 7.06 | 56.20 | |
| 17 | 437 [0] | 200 [0.0] | 2 [0.0] | 1.12 | 7.11 | 57.14 | |
| 18 | 437 [0] | 200 [0.0] | 2 [0.0] | 1.16 | 7.10 | 57.34 | |
| 19 | 437 [0] | 200 [0.0] | 2 [0.0] | 1.22 | 7.39 | 54.11 | |
| 20 | 437 [0] | 200 [0.0] | 2 [0.0] | 1.04 | 6.71 | 49.53 |
| Factor | Low | High | Optimum |
|---|---|---|---|
| Granulometry (µm) | 97.5 | 925.0 | 740 |
| Agitation (rpm) | 35.18 | 365.0 | 240 |
| WB Concentration (g/50 mL) | 0.32 | 3.7 | 3.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fayad, N.; Abboud, J.; Driss, F.; Louka, N.; Kallassy Awad, M. Optimization of Culture Conditions and Wheat Bran Class Selection in the Production of Bacillus thuringiensis-Based Biopesticides. Fermentation 2022, 8, 666. https://doi.org/10.3390/fermentation8120666
Fayad N, Abboud J, Driss F, Louka N, Kallassy Awad M. Optimization of Culture Conditions and Wheat Bran Class Selection in the Production of Bacillus thuringiensis-Based Biopesticides. Fermentation. 2022; 8(12):666. https://doi.org/10.3390/fermentation8120666
Chicago/Turabian StyleFayad, Nancy, Joanna Abboud, Fatma Driss, Nicolas Louka, and Mireille Kallassy Awad. 2022. "Optimization of Culture Conditions and Wheat Bran Class Selection in the Production of Bacillus thuringiensis-Based Biopesticides" Fermentation 8, no. 12: 666. https://doi.org/10.3390/fermentation8120666