
Isolation of Yeasts from Some Homemade Fermented Cow-Milk Products of Sikkim and Their Probiotic Characteristics

Abstract
:1. Introduction
2. Materials and Methods
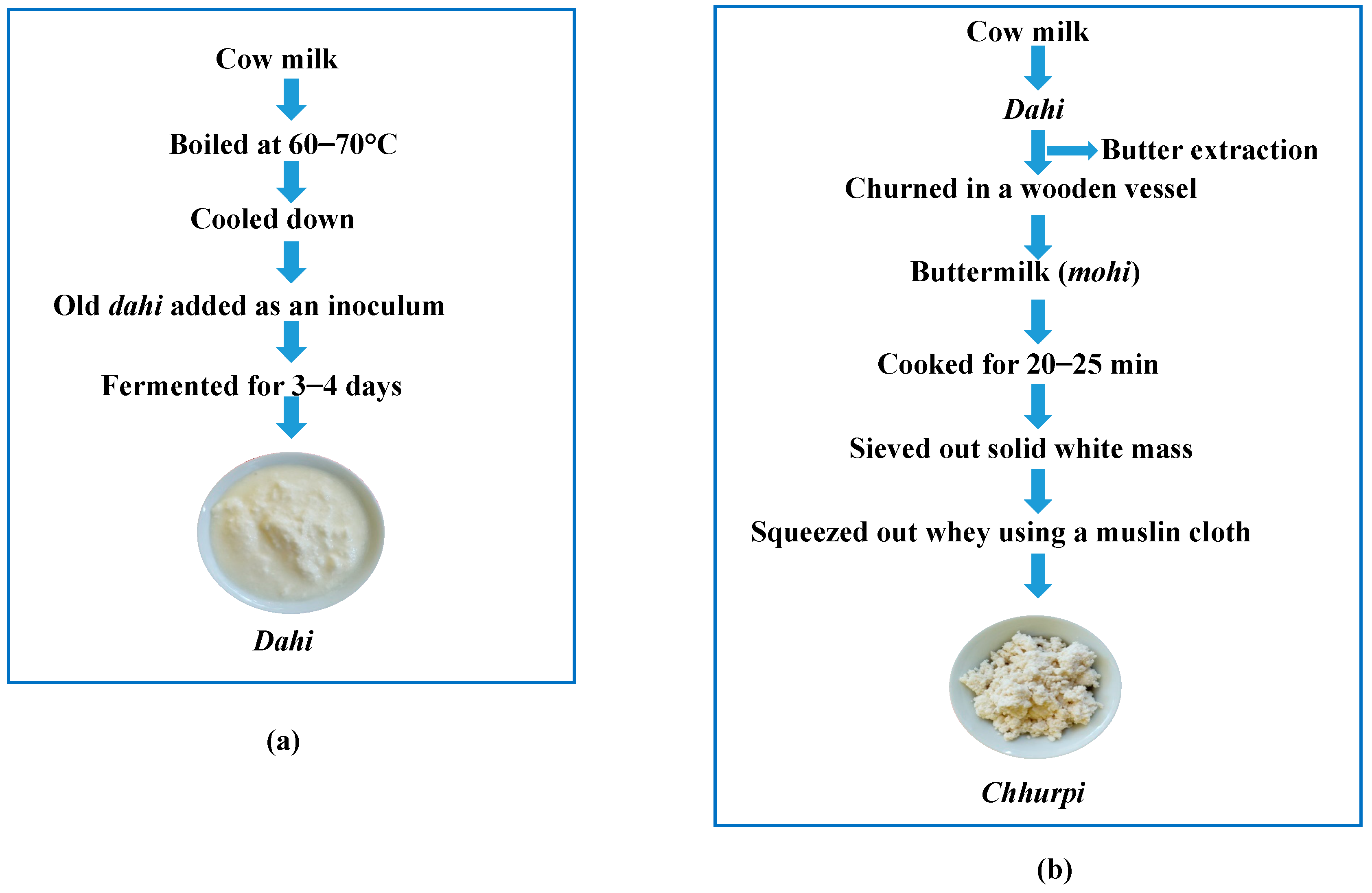
2.1. Collection of Samples
2.2. Analysis of pH
2.3. Titratable Acidity
2.4. Moisture Content
2.5. Viscosity
2.6. Microbiological Analysis
2.7. Preliminary Screening of Probiotic Isolates
2.8. Phenotypic and Biochemical Characterization
2.9. Genotypic Characterization
2.9.1. DNA Extraction
2.9.2. PCR Amplification
2.10. In Vitro Screening of Probiotic Properties
Auto-Aggregation and Co-Aggregation
2.11. Antimicrobial Activity
2.12. Deconjugation of Bile Salts (BSH Activity)
2.13. Lysozyme Tolerance
2.14. Genetic Screening for Probiotic Traits
2.15. Bioinformatics Analysis
3. Results
3.1. Product Characteristics and Preliminary Screening of Probiotic Yeasts
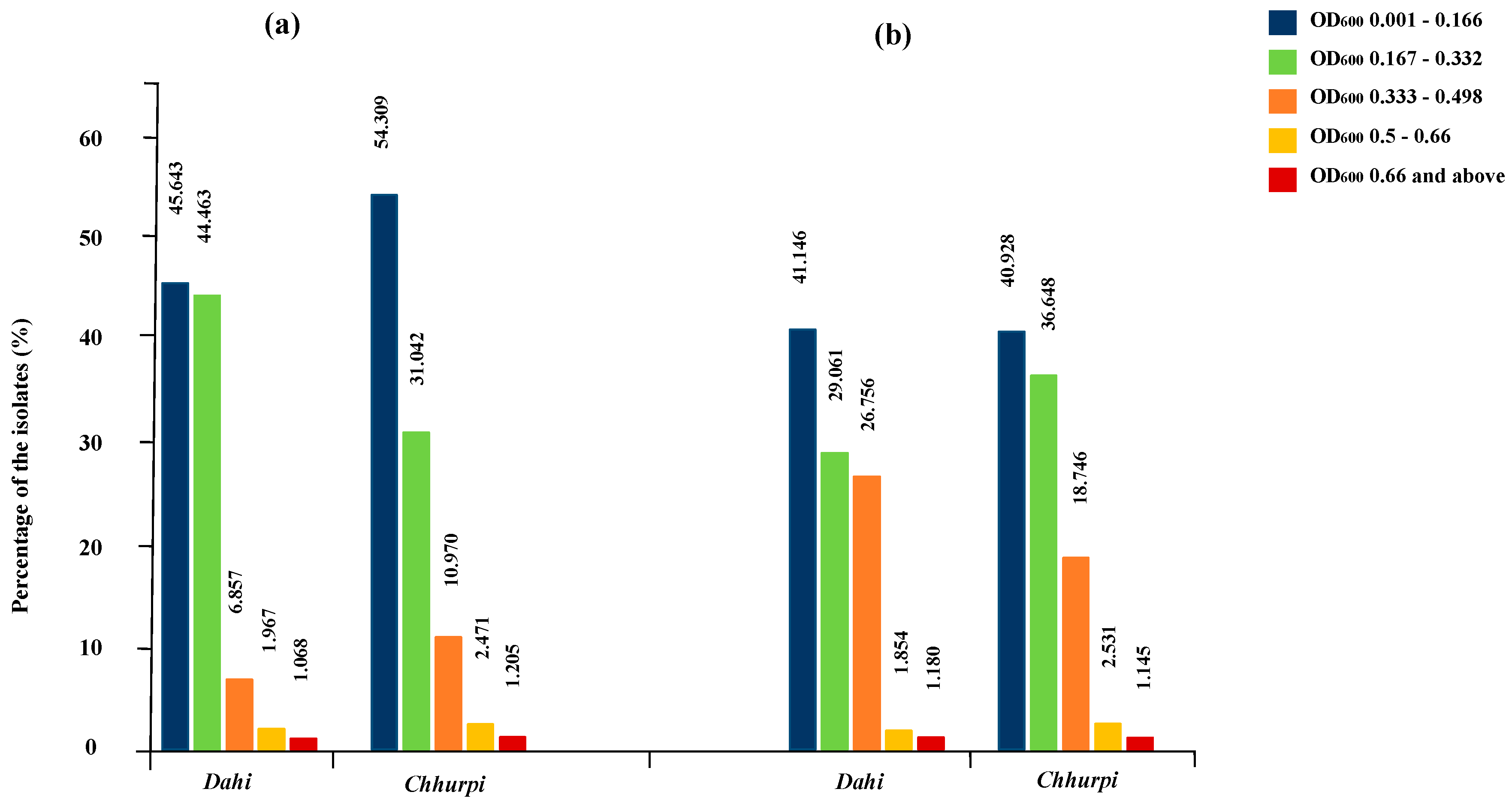
3.2. In Vitro Probiotic Screening
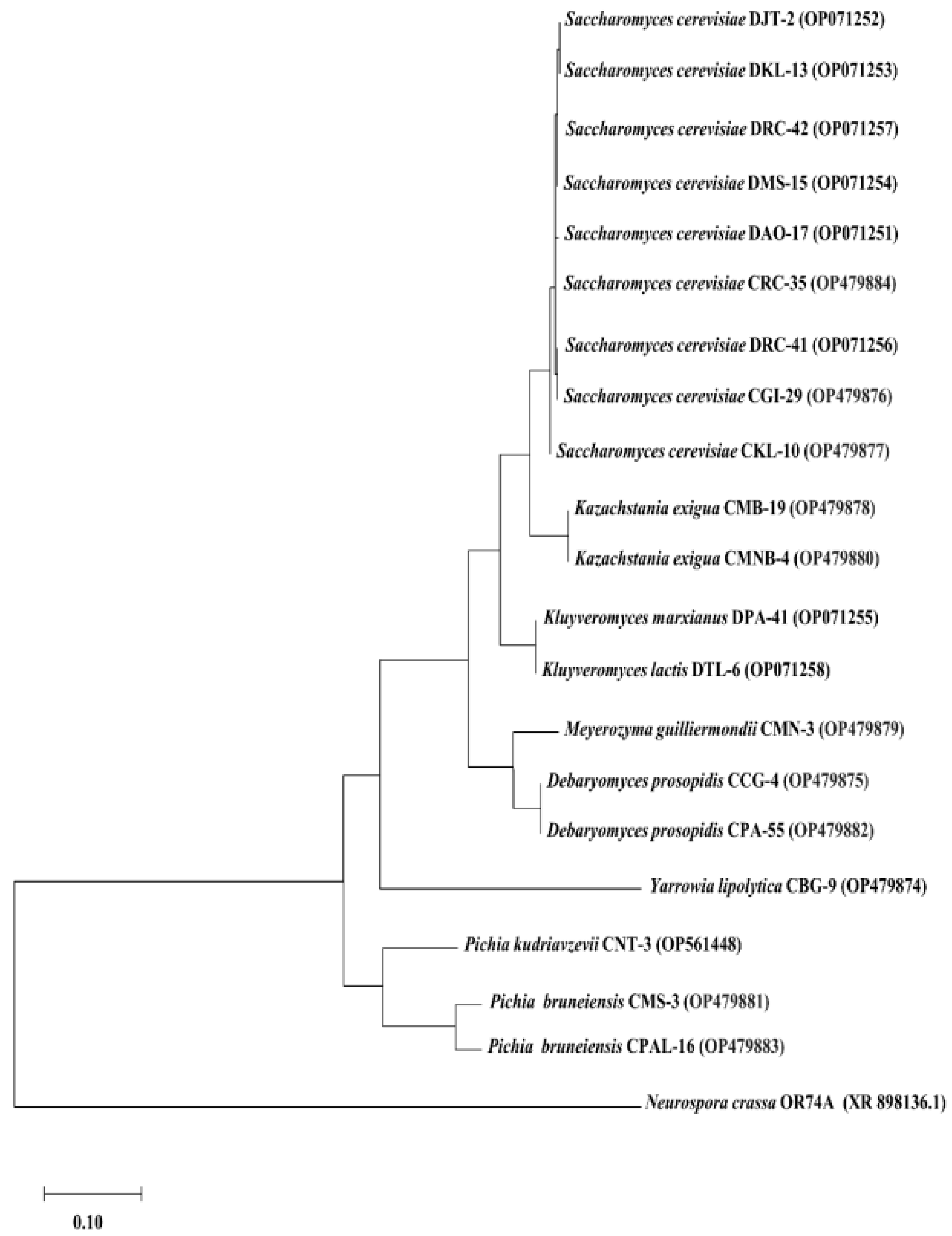
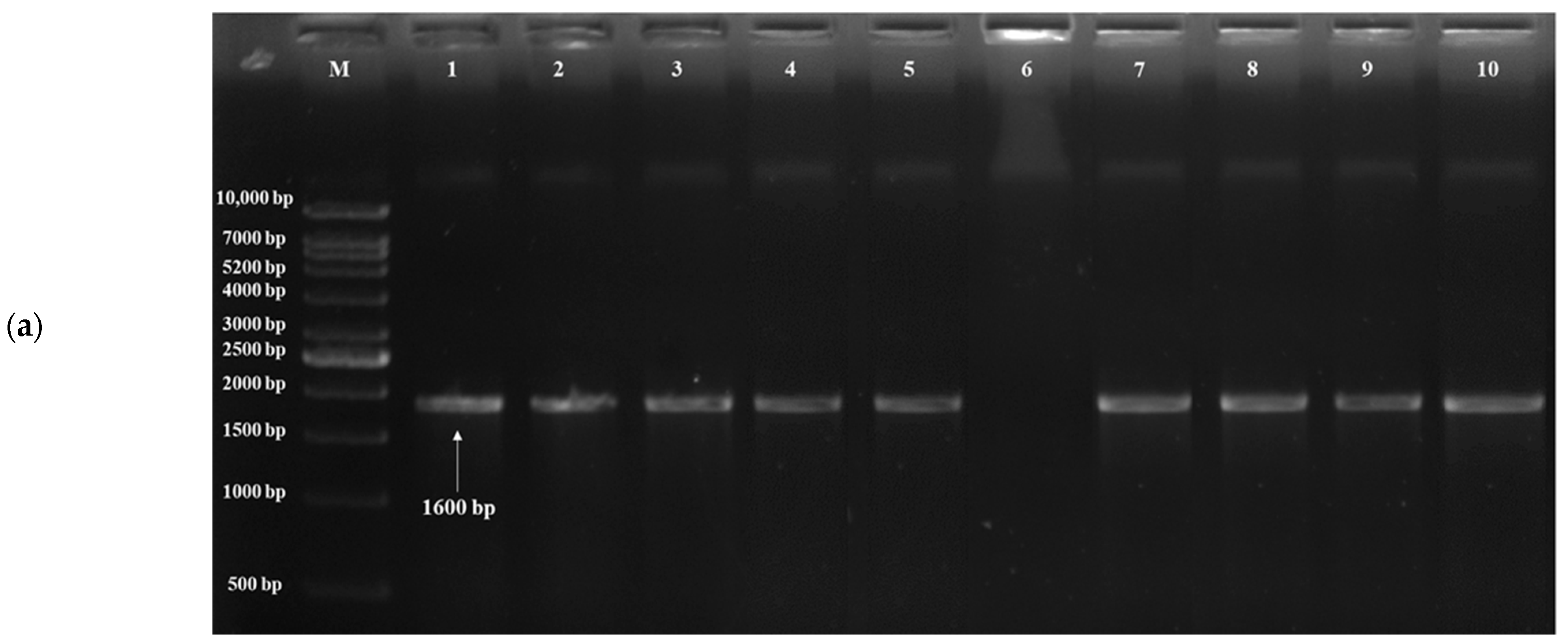
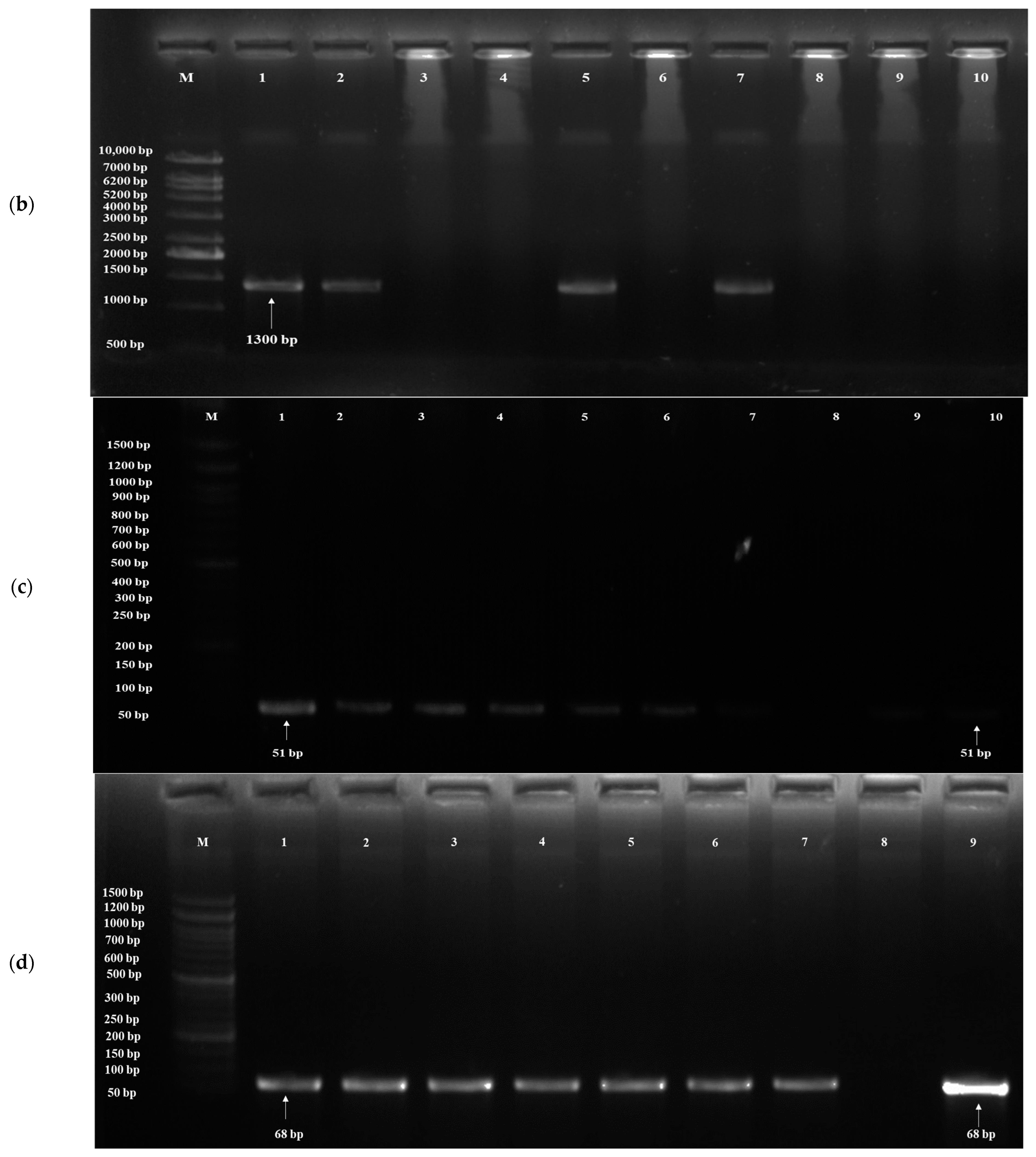
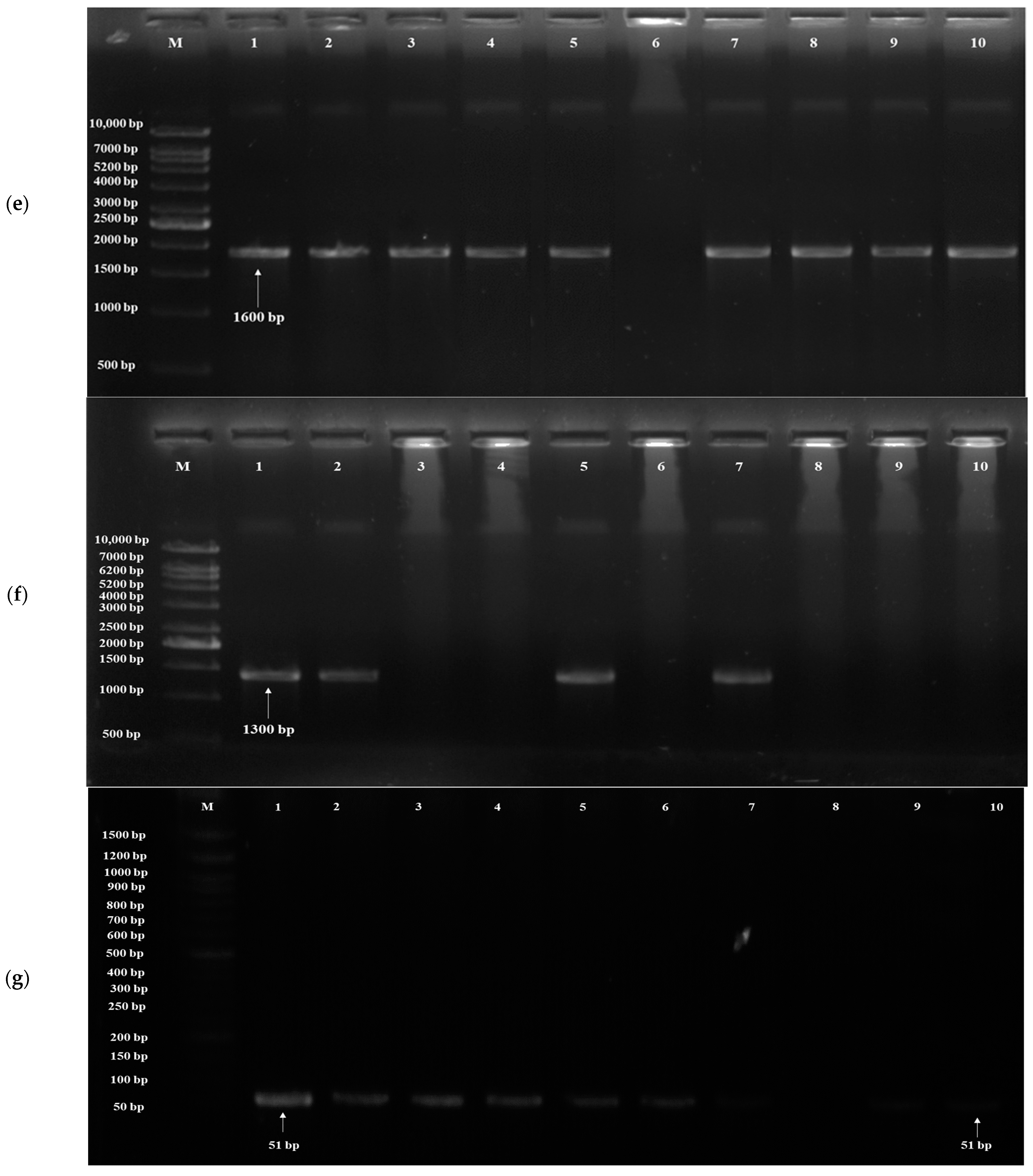
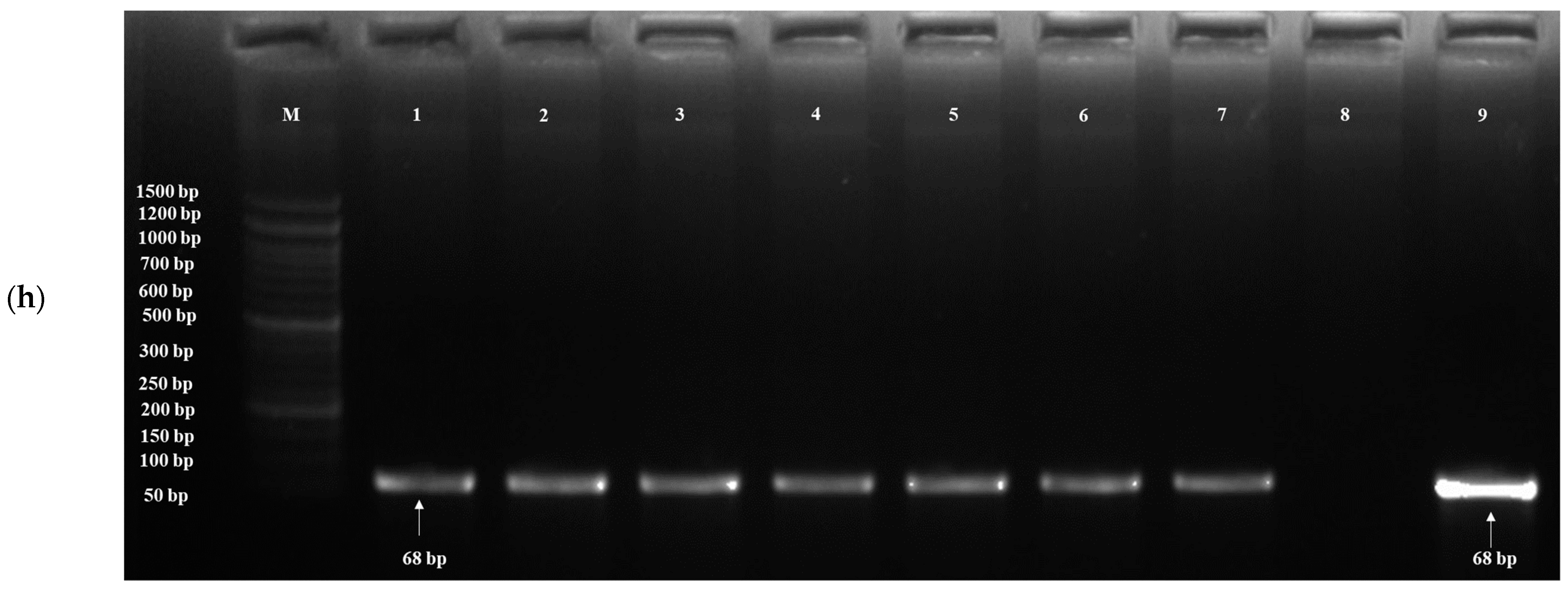
3.3. Detection of Probiotic Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mukhtar, H.; Yaqub, S.; ul Haq, I. Production of probiotic Mozzarella cheese by incorporating locally isolated Lactobacillus acidophilus. Ann. Microbiol. 2020, 70, 56. [Google Scholar] [CrossRef]
- Savaiano, D.A.; Hutkins, R.W. Yogurt, cultured fermented milk, and health: A systematic review. Nutr. Rev. 2021, 79, 599–614. [Google Scholar] [CrossRef]
- Zahoor, F.; Sooklim, C.; Songdech, P.; Duangpakdee, O.; Soontorngun, N. Selection of potential yeast probiotics and a cell factory for xylitol or acid production from honeybee samples. Metabolites 2021, 11, 312. [Google Scholar] [CrossRef] [PubMed]
- Hadjimbei, E.; Botsaris, G.; Chrysostomou, S. Beneficial effects of yoghurts and probiotic fermented milks and their functional food potential. Foods 2022, 11, 2691. [Google Scholar] [CrossRef] [PubMed]
- Lynch, K.M.; Wilkinson, S.; Daenen, L.; Arendt, E.K. An update on water kefir: Microbiology, composition and production. Int. J. Food Microbiol. 2021, 345, 109128. [Google Scholar] [CrossRef]
- Castellone, V.; Bancalari, E.; Rubert, J.; Gatti, M.; Neviani, E.; Bottari, B. Eating fermented: Health benefits of LAB-fermented foods. Foods 2021, 10, 2639. [Google Scholar] [CrossRef]
- Yilmaz, B.; Elibol, E.; Shangpliang, H.N.J.; Ozogul, F.; Tamang, J.P. Microbial communities in home-made and commercial kefir and their hypoglycemic properties. Fermentation 2022, 8, 590. [Google Scholar] [CrossRef]
- Rai, R.; Shangpliang, H.N.J.; Tamang, J.P. Naturally fermented milk products of the Eastern Himalayas. J. Ethn. Foods 2016, 3, 270–275. [Google Scholar] [CrossRef]
- Teneva-Angelova, T.; Balabanova, T.; Boyanova, P.; Beshkova, D. Traditional Balkan fermented milk products. Eng. Life Sci. 2018, 18, 807–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agyei, D.; Owusu-Kwarteng, J.; Akabanda, F.; Akomea-Frempong, S. Indigenous African fermented dairy products: Processing technology, microbiology and health benefits. Crit. Rev. Food Sci. Nutr. 2020, 60, 991–1006. [Google Scholar] [CrossRef]
- Faccia, M.; D’Alessandro, A.; Summer, A.; Hailu, Y. Milk products from minor dairy species: A review. Animals 2020, 10, 1260. [Google Scholar] [CrossRef] [PubMed]
- Ganzorig, K.; Urashima, T.; Fukuda, K. Exploring potential bioactive peptides in fermented bactrian camel’s milk and mare’s milk made by Mongolian nomads. Foods 2020, 9, 1817. [Google Scholar] [CrossRef]
- Tamang, J.P.; Cotter, P.; Endo, A.; Han, N.S.; Kort, R.; Liu, S.Q.; Mayo, B.; Westerik, N.; Hutkins, R. Fermented foods in a global age: East meets west. Compr. Rev. Food Sci. Food Saf. 2020, 19, 184–217. [Google Scholar] [CrossRef] [Green Version]
- Tamang, J.P. “Ethno-Microbiology” of ethnic Indian fermented foods and alcoholic beverages. J. Appl. Microbiol. 2022, 133, 145–161. [Google Scholar] [CrossRef]
- Parker, M.; Zobrist, S.; Donahue, C.; Edick, C.; Mansen, K.; Hassan, Z.N.M.; Heerikhuisen, M.; Sybesma, W.; Molenaar, D.; Diallo, A.M.; et al. Naturally fermented milk from northern senegal: Bacterial community composition and probiotic enrichment with Lactobacillus rhamnosus. Front. Microbiol. 2018, 9, 2218. [Google Scholar] [CrossRef] [Green Version]
- Takeda, S.; Yamasaki, K.; Takeshita, M.; Kikuchi, Y.; Tsend-Ayush, C.; Dashnyam, B.; Ahhmed, A.M.; Kawahara, S.; Muguruma, M. The investigation of probiotic potential of lactic acid bacteria isolated from traditional Mongolian dairy products. Anim. Sci. J. 2011, 82, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Maleke, M.S.; Adefisoye, M.A.; Doorsamy, W.; Adebo, O.A. Processing, nutritional composition and microbiology of amasi: A Southern African fermented milk product. Sci. Afr. 2021, 12, e00795. [Google Scholar] [CrossRef]
- Venema, K.; Surono, I.S. Microbiota composition of dadih—A traditional fermented buffalo milk of West Sumatra. Lett. Appl. Microbiol. 2019, 68, 234–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balamurugan, R.; Chandragunasekaran, A.S.; Chellappan, G.; Rajaram, K.; Ramamoorthi, G.; Ramakrishna, B.S. Probiotic potential of lactic acid bacteria present in home made curd in southern India. Indian J. Med. Res. 2014, 140, 345–355. [Google Scholar]
- Rai, R.; Tamang, J.P. In vitro and genetic screening of probiotic properties of lactic acid bacteria isolated from naturally fermented cow-milk and yak-milk products of Sikkim, India. World J. Microbiol. Biotechnol. 2022, 38, 25. [Google Scholar] [CrossRef]
- Adegboye, B.; Banwo, K.; Ogunremi, O.; Sanni, A. Probiotic potentials of yeasts isolated from nunu (African fermented milk) and wara (African soft cheese). Adv. Food Sci. 2014, 36, 115–124. [Google Scholar]
- Tamang, J.P.; Watanabe, K.; Holzapfel, W.H. Diversity of microorganisms in global fermented foods and beverages. Front. Microbiol. 2016, 7, 377. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- Ağagündüz, D.; Yılmaz, B.; Şahin, T.Ö.; Güneşliol, B.E.; Ayten, Ş.; Russo, P.; Spano, G.; Rocha, J.M.; Bartkiene, E.; Özogul, F. Dairy lactic acid bacteria and their potential function in dietetics: The food–gut-health axis. Foods 2021, 10, 3099. [Google Scholar] [CrossRef]
- Gaucher, F.; Kponouglo, K.; Rabah, H.; Bonnassie, S.; Ossemond, J.; Pottier, S.; Jardin, J.; Briard-Bion, V.; Marchand, P.; Blanc, P.; et al. Propionibacterium freudenreichii CIRM-BIA 129 osmoadaptation coupled to acid-adaptation increases its viability during freeze-drying. Front. Microbiol. 2019, 10, 2324. [Google Scholar] [CrossRef]
- Wong, C.B.; Odamaki, T.; Xiao, J.Z. Beneficial effects of Bifidobacterium longum subsp. longum BB536 on human health: Modulation of gut microbiome as the principal action. J. Funct. Foods 2019, 54, 506–519. [Google Scholar] [CrossRef]
- Gut, A.M.; Vasiljevic, T.; Yeager, T.; Donkor, O.N. Characterization of yeasts isolated from traditional kefir grains for potential probiotic properties. J. Funct. Foods 2019, 58, 56–66. [Google Scholar] [CrossRef]
- Galli, V.; Venturi, M.; Mari, E.; Guerrini, S.; Granchi, L. Selection of yeast and lactic acid bacteria strains, isolated from spontaneous raw milk fermentation, for the production of a potential probiotic fermented milk. Fermentation 2022, 8, 407. [Google Scholar] [CrossRef]
- Tamang, J.P.; Lama, S. Probiotic properties of yeasts in traditional fermented foods and beverages. J. Appl. Microbiol. 2022, 132, 3533–3542. [Google Scholar] [CrossRef]
- Roohvand, F.; Ehsani, P.; Abdollahpour-Alitappeh, M.; Shokri, M.; Kossari, N. Biomedical applications of yeasts—A patent view, part two: Era of humanized yeasts and expanded applications. Expert Opin. Ther. Pat. 2020, 30, 609–631. [Google Scholar] [CrossRef]
- Karaolis, C.; Botsaris, G.; Pantelides, I.; Tsaltas, D. Potential application of Saccharomyces boulardii as a probiotic in goat’s yoghurt: Survival and organoleptic effects. Int. J. Food Sci. Technol. 2013, 48, 1445–1452. [Google Scholar] [CrossRef]
- Sarwar, A.; Aziz, T.; Al-Dalali, S.; Zhao, X.; Zhang, J.; ud Din, J.; Chen, C.; Cao, Y.; Yang, Z. Physicochemical and microbiological properties of synbiotic yogurt made with probiotic yeast Saccharomyces boulardii in Combination with inulin. Foods 2019, 8, 468. [Google Scholar] [CrossRef]
- Ansari, F.; Alian Samakkhah, S.; Bahadori, A.; Jafari, S.M.; Ziaee, M.; Khodayari, M.T.; Pourjafar, H. Health-promoting properties of Saccharomyces cerevisiae var. boulardii as a probiotic; characteristics, isolation, and applications in dairy products. Crit. Rev. Food Sci. Nutr. 2021, 13, 1949577. [Google Scholar] [CrossRef] [PubMed]
- Tamang, J.P.; Jeyaram, K.; Rai, A.K.; Mukherjee, P.K. Diversity of beneficial microorganisms and their functionalities in community-specific ethnic fermented foods of the Eastern Himalayas. Food Res. Int. 2021, 148, 110633. [Google Scholar] [CrossRef]
- Tamang, J.P. Dietary culture and antiquity of the Himalayan fermented foods and alcoholic fermented beverages. J. Ethn. Foods 2022, 9, 30. [Google Scholar] [CrossRef]
- Shangpliang, H.; Rai, R.; Keisam, S.; Jeyaram, K.; Tamang, J.P. Bacterial community in naturally fermented milk products of Arunachal Pradesh and Sikkim of India analysed by high-throughput amplicon sequencing. Sci. Rep. 2018, 8, 1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shangpliang, H.N.J.; Tamang, J.P. Phenotypic and genotypic characterisation of lactic acid bacteria isolated from exotic naturally fermented milk (cow and yak) products of Arunachal Pradesh, India. Int. Dairy J. 2021, 118, 105038. [Google Scholar] [CrossRef]
- Dewan, S.; Tamang, J.P. Microbial and analytical characterization of Chhu-A traditional fermented milk product of Sikkim Himalayas. J. Sci. Ind. Res. 2006, 65, 747–752. [Google Scholar]
- Dewan, S.; Tamang, J.P. Dominant lactic acid bacteria and their technological properties isolated from the Himalayan ethnic fermented milk products. Antonie Leeuwenhoek 2007, 92, 343–352. [Google Scholar] [CrossRef]
- Tamang, J.P.; Dewan, S.; Thapa, S.; Olasupo, N.A.; Schillinger, U.; Wijaya, A.; Holzapfel, W.H. Identification and enzymatic profiles of the predominant lactic acid bacteria isolated from soft-variety Chhurpi, a traditional cheese typical of the Sikkim Himalayas. Food Biotechnol. 2000, 14, 99–112. [Google Scholar] [CrossRef]
- Goel, A.; Halami, P.M.; Tamang, J.P. Genome analysis of Lactobacillus plantarum isolated from some Indian fermented foods for bacteriocin production and probiotic marker genes. Front. Microbiol. 2020, 11, 40. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.K.; Kumari, R.; Sanjukta, S.; Sahoo, D. Production of bioactive protein hydrolysate using the yeasts isolated from soft chhurpi. Bioresour. Technol. 2016, 1, 239–245. [Google Scholar] [CrossRef]
- Abedin, A.M.; Chourasia, R.; Phukon, L.C.; Singh, S.P.; Rai, A.K. Characterization of ACE inhibitory and antioxidant peptides in yak and cow milk hard chhurpi cheese of the Sikkim Himalayan region. Food Chem. 2022, 13, 100231. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 20th ed.; Association of Official Analytical Chemists: Rockville, MD, USA, 2016. [Google Scholar]
- Pratap, D.; Maurya, V.K.; Kumar, N.; Singh, R.; Upadhyay, A. Studies on physico-chemical and organoleptic properties of soymilk blended dahi (curd) with toned milk (cattle milk). Curr. Nutr. Food Sci. 2018, 14, 61–67. [Google Scholar] [CrossRef]
- Ali, K.; Mehmood, M.H.; Iqbal, M.A.; Masud, T.; Qazalbash, M.; Saleem, S.; Ahmed, S.; Tariq, M.R.; Safdar, W.; Nasir, M.A.; et al. Isolation and characterization of exopolysaccharide-producing strains of Lactobacillus bulgaricus from curd. Food Sci. Nutr. 2019, 7, 1207–1213. [Google Scholar] [CrossRef] [Green Version]
- Greppi, A.; Saubade, F.; Botta, C.; Humblot, C.; Guyot, J.P.; Cocolin, L. Potential probiotic Pichia kudriavzevii strains and their ability to enhance folate content of traditional cereal-based African fermented food. Food Microbiol. 2017, 62, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Zhang, Y.; Zhang, Y.; Liu, Y.; Wang, S.; Dong, X.; Wang, Y.; Zhang, H. Screening of potential probiotic properties of Lactobacillus fermentum isolated from traditional dairy products. Food Control 2010, 21, 695–701. [Google Scholar] [CrossRef]
- Fernandez-Pacheco, P.; Arévalo-Villena, M.; Bevilacqua, A.; Corbo, M.R.; Pérez, A.B. Probiotic characteristics in Saccharomyces cerevisiae strains: Properties for application in food industries. LWT-Food Sci. Technol. 2018, 97, 332–340. [Google Scholar] [CrossRef]
- Alkalbani, N.S.; Osaili, T.M.; Al-Nabulsi, A.A.; Obaid, R.S.; Olaimat, A.N.; Liu, S.Q.; Ayyash, M.M. In vitro characterization and identification of potential probiotic yeasts isolated from fermented dairy and non-dairy food products. J. Fungi 2022, 8, 544. [Google Scholar] [CrossRef]
- Umeh, S.O.; Agwuna, L.C.; Okafor, U.C. Yeasts from local sources: An alternative to the conventional brewer’s yeast. Int. J. Biotechnol. Food Sci. 2017, 30, 191–195. [Google Scholar]
- Kotzekidou, P. Identification of yeasts from black olives in rapid system microtitre plates. Food Microbiol. 1997, 14, 609–616. [Google Scholar] [CrossRef]
- Oliveira, V.A.; Vicente, M.A.; Fietto, L.G.; de Miranda Castro, I.; Coutrim, M.X.; Schüller, D.; Alves, H.; Casal, M.; de Oliveira Santos, J.; Araújo, L.D.; et al. Biochemical and molecular characterization of Saccharomyces cerevisiae strains obtained from sugar-cane juice fermentations and their impact in cachaca production. Appl. Environ. Microbiol. 2008, 74, 693–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renshaw, M.A.; Olds, B.P.; Jerde, C.L.; McVeigh, M.M.; Lodge, D.M. The room temperature preservation of filtered environmental DNA samples and assimilation into a phenol–chloroform–isoamyl alcohol DNA extraction. Mol. Ecol. Resour. 2015, 15, 168–176. [Google Scholar] [CrossRef] [Green Version]
- Palla, M.; Blandino, M.; Grassi, A.; Giordano, D.; Sgherri, C.; Quartacci, M.F.; Reyneri, A.; Agnolucci, M.; Giovannetti, M. Characterization and selection of functional yeast strains during sourdough fermentation of different cereal wholegrain flours. Sci. Rep. 2020, 10, 12856. [Google Scholar] [CrossRef] [PubMed]
- Ogunremi, O.; Sanni, A.; Agrawal, R. Probiotic potentials of yeasts isolated from some cereal-based Nigerian traditional fermented food products. J. Appl. Microbiol. 2015, 119, 797–808. [Google Scholar] [CrossRef] [PubMed]
- Fadda, M.E.; Mossa, V.; Deplano, M.; Pisano, M.B.; Cosentino, S. In vitro screening of Kluyveromyces strains isolated from Fiore Sardo cheese for potential use as probiotics. LWT-Food Sci. Technol. 2017, 75, 100–106. [Google Scholar] [CrossRef]
- Vera-Pingitore, E.; Jimenez, M.E.; Dallagnol, A.; Belfiore, C.; Fontana, C.; Fontana, P.; von Wright, A.; Vignolo, G.; Plumed-Ferrer, C. Screening and characterization of potential probiotic and starter bacteria for plant fermentations. LWT-Food Sci. Technol. 2016, 1, 288–294. [Google Scholar] [CrossRef]
- Wilson, R.A.; Jenkinson, J.M.; Gibson, R.P.; Littlechild, J.A.; Wang, Z.Y.; Talbot, N.J. Tps1 regulates the pentose phosphate pathway, nitrogen metabolism and fungal virulence. EMBO J. 2007, 26, 3673–3685. [Google Scholar] [CrossRef] [Green Version]
- Marinangeli, P.; Angelozzi, D.; Ciani, M.; Clementi, F.; Mannazzu, I. Minisatellites in Saccharomyces cerevisiae genes encoding cell wall proteins: A new way towards wine strain characterisation. FEMS Yeast Res. 2004, 4, 427–435. [Google Scholar] [CrossRef] [Green Version]
- Mannazzu, I.; Simonetti, E.; Marinangeli, P.; Guerra, E.; Budroni, M.; Thangavelu, M.; Clementi, F. SED1 gene length and sequence polymorphisms in feral strains of Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2002, 68, 5437–5444. [Google Scholar] [CrossRef] [Green Version]
- Caudle, K.E.; Barker, K.S.; Wiederhold, N.P.; Xu, L.; Homayouni, R.; Rogers, P.D. Genome wide expression profile analysis of the Candida glabrata Pdr1 regulon. Eukaryot. Cell 2011, 10, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.S.; Choo, Y.M.; Lee, K.S.; Yoon, H.J.; Kim, I.; Je, Y.H.; Sohn, H.D.; Jin, B.R. Cloning and expression profiling of four antibacterial peptide genes from the bumblebee Bombus ignitus. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 15, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Younis, G.; Awad, A.; Dawod, R.E.; Yousef, N.E. Antimicrobial activity of yeasts against some pathogenic bacteria. Vet. World. 2017, 10, 979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damas-Buenrostro, L.C.; Gracia-González, G.; Hernández-Luna, C.E.; Galán-Wong, L.J.; Pereyra-Alférez, B.; Sierra-Benavides, J.A. Detection of FLO genes in lager and wild yeast strains. J. Am. Soc. Brew. Chem. 2008, 66, 184–187. [Google Scholar] [CrossRef]
- Nutaratat, P.; Boontham, W.; Khunnamwong, P. A novel yeast genus and two novel species isolated from pineapple leaves in Thailand: Savitreella phatthalungensis gen. nov., sp. nov. and Goffeauzyma siamensis sp. nov. J. Fungi 2022, 8, 118. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- García-Burgos, M.; Moreno-Fernández, J.; Alférez, M.J.M.; Díaz-Castro, J.; López-Aliaga, I. New perspectives in fermented dairy products and their health relevance. J. Funct. Foods 2020, 72, 104059. [Google Scholar] [CrossRef]
- Widyastuti, Y.; Febrisiantosa, A.; Tidona, F. Health-Promoting properties of lactobacilli in fermented dairy products. Front. Microbiol. 2021, 12, 673890. [Google Scholar] [CrossRef]
- Staniszewski, A.; Kordowska-Wiater, M. Probiotic and potentially probiotic yeasts-characteristics and food application. Foods 2021, 10, 1306. [Google Scholar] [CrossRef]
- Hatoum, R.; Labrie, S.; Fliss, I. Antimicrobial and probiotic properties of yeasts: From fundamental to novel applications. Front. Microbiol. 2012, 3, 421. [Google Scholar] [CrossRef] [Green Version]
- Suharja, A.; Henriksson, A.; Liu, S.Q. Impact of Saccharomyces cerevisiae on Viability of probiotic Lactobacillus rhamnosus in fermented milk under ambient conditions. J. Food Process. Preserv. 2014, 38, 326–337. [Google Scholar] [CrossRef]
- Buldo, P.; Sokolowsky, M.; Hoegholm, T. The role of starter cultures on oral processing properties of different fermented milk products. Food Hydrocoll. 2021, 114, 106571. [Google Scholar] [CrossRef]
- Markowiak, P.; Śliżewska, K. Effects of probiotics, prebiotics, and synbiotics on human health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Moradi, R.; Nosrati, R.; Zare, H.; Tahmasebi, T.; Saderi, H.; Owlia, P. Screening and characterization of in-vitro probiotic criteria of Saccharomyces and Kluyveromyces strains. Iran J. Microbiol. 2018, 10, 123–131. [Google Scholar]
- Kechagia, M.; Basoulis, D.; Konstantopoulou, S.; Dimitriadi, D.; Gyftopoulou, K.; Skarmoutsou, N.; Fakiri, E.M. Health benefits of probiotics: A review. ISRN Nutr. 2013, 2013, 481651. [Google Scholar] [CrossRef] [PubMed]
- Nostro, A.; Cannatelli, M.A.; Crisafi, G.; Musolino, A.D.; Procopio, F.; Alonzo, V. Modifications of hydrophobicity, in vitro adherence and cellular aggregation of Streptococcus mutans by Helichrysum italicum extract. Lett. Appl. Microbiol. 2004, 38, 423–427. [Google Scholar] [CrossRef]
- de Oliveira Coelho, B.; Fiorda-Mello, F.; de Melo Pereira, G.V.; Thomaz-Soccol, V.; Rakshit, S.K.; de Carvalho, J.C.; Soccol, C.R. In vitro probiotic properties and dna protection activity of yeast and lactic acid bacteria isolated from a honey-based kefir beverage. Foods 2019, 8, 485. [Google Scholar] [CrossRef] [Green Version]
- Kumura, H.; Tanoue, Y.; Tsukahara, M.; Tanaka, T.; Shimazaki, K. Screening of dairy yeast strains for probiotic applications. J. Dairy Sci. 2004, 87, 4050–4056. [Google Scholar] [CrossRef] [Green Version]
- Cissé, H.; Kagambèga, B.; Sawadogo, A.; Tankoano, A.; Sangaré, G.; Traoré, Y.; Ouoba, I.I.L.; Savadogo, A. Molecular characterization of Bacillus, lactic acid bacteria and yeast as potential probiotic isolated from fermented food. Sci. Afr. 2019, 6, 00175. [Google Scholar] [CrossRef]
- Nahidul-Islam, S.M.; Kuda, T.; Takahashi, H.; Kimura, B. Bacterial and fungal microbiota in traditional Bangladeshi fermented milk products analysed by culture-dependent and culture-independent methods. Food Res. Int. 2018, 111, 431–437. [Google Scholar] [CrossRef]
- Pérez, P.F.; Minnaard, Y.; Disalvo, E.A.; De Antoni, G.L. Surface properties of bifidobacterial strains of human origin. Appl. Environ. Microbiol. 1998, 64, 21–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syal, P.; Vohra, A. Probiotic potential of yeasts isolated from traditional Indian fermented foods. Int. J. Microbiol. Res. 2013, 5, 390. [Google Scholar] [CrossRef] [Green Version]
- Bonatsou, S.; Karamouza, M.; Zoumpopoulou, G.; Mavrogonatou, E.; Kletsas, D.; Papadimitriou, K.; Tsakalidou, E.; Nychas, G.J.E.; Panagou, E.Ζ. Evaluating the probiotic potential and technological characteristics of yeasts implicated in cv. Kalamata natural black olive fermentation. Int. J. Food Microbiol. 2018, 271, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Suvarna, S.; Dsouza, J.; Ragavan, M.L.; Das, N. Potential probiotic characterization and effect of encapsulation of probiotic yeast strains on survival in simulated gastrointestinal tract condition. Food Sci. Biotechnol. 2018, 27, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Fakruddin, M.; Hossain, M.N.; Ahmed, M.M. Antimicrobial and antioxidant activities of Saccharomyces cerevisiae IFST062013, a potential probiotic. BMC Complement. Altern. Med. 2017, 17, 64. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Gómez, J.G.; López-Bonilla, A.; Trejo-Tapia, G.; Ávila-Reyes, S.V.; Jiménez-Aparicio, A.R.; Hernández-Sánchez, H. In vitro bile salt hydrolase (BSH) activity screening of different probiotic microorganisms. Foods 2021, 10, 674. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, K.; Zoumpopoulou, G.; Foligné, B.; Alexandraki, V.; Kazou, M.; Pot, B.; Tsakalidou, E. Discovering probiotic microorganisms: In vitro, in vivo, genetic and omics approaches. Front. Microbiol. 2015, 6, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turpin, W.; Humblot, C.; Guyot, J.P. Genetic screening of functional properties of lactic acid bacteria in a fermented pearl millet slurry and in the metagenome of fermented starchy foods. Appl. Environ. Microbiol. 2011, 77, 8722–8734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turpin, W.; Humblot, C.; Noordine, M.L.; Thomas, M.; Guyot, J.P. Lactobacillaceae and cell adhesion: Genomic and functional screening. PLoS ONE 2012, 7, e38034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawahata, M.; Masaki, K.; Fujii, T.; Iefuji, H. Yeast genes involved in response to lactic acid and acetic acid: Acidic conditions caused by the organic acids in Saccharomyces cerevisiae cultures induce expression of intracellular metal metabolism genes regulated by Aft1p. FEMS Yeast Res. 2006, 6, 924–936. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Probiotic Traits | PrimersF:5′-3′R:3′-5′ | Annealing Temperature (°C) | Amplicon Size (bp) | References |
|---|---|---|---|---|---|
| TPS1 | Acid tolerance | F- ATGACTACGGATAACG R- TCAGTTTTTGGTGGCAGAGG | 65 | 1600 | [59] |
| HSP150 | Acid tolerance | F- CACTTTGACTCCAACAGCCACTTACA R- TACCGGACAAACATTGGTAGAAGACA | 65 | 781 | [60] |
| SED1 | Acid tolerance | F- ATGAAATTATCAACTGTCCTATTATCTGCCGG R- TTATAAGAATAACATAGCAACACCAGCCAAACC | 64 | 950–1300 | [61] |
| YIM1 | Bile tolerance | F- CAAGAAATGGACCCCCGAGT R- TGCGTGGAAGCACCATATAC | 64 | 51 | [62] |
| PDR1 | Bile tolerance | F- TTTGACTCTGTTATGAGCGATTACG R- TTCGGATTTTTCTGTGACAATGG | 64 | 51 | [62] |
| YOR1 | Bile tolerance | F- CCATCGGTGCTTGTGTAATGTTA R- TTGAGAGGCGTGGAAAAAATG | 64 | 68 | [62] |
| ERG3 | Bile tolerance | F- AAGCGTGTGAACAAGGAC R- GCGTAGGTCTTCTCTGTGA | 64 | 68 | [62] |
| EPA1 | Bile tolerance | F- AACCGCAAGAAAATCCTCCT R- GGACTGGAAGTGGGGTATGA | 64 | 60 | [62] |
| Apid | Antimicrobial activity | F- ATGAAGAATTTTATCTTCGCTATT R- TCAGTAATATAATTCCTCATCAGC | 48 | 752 | [63] |
| khs | Antimicrobial activity | F- AAGCATCCGAAACAGTACT R- TCAAGGATGCTGCTGCTAAGCTG | 53 | 919 | [64] |
| pelA | Antimicrobial activity | F- ATCGAATTCATGAAGTTCACTGCTGCTTTC R- ACGGAATTCGCAGCTCGTGGTGGAGCCAGT | 55 | 727 | [64] |
| FLO1 | Adhesion | F- ATGACAATGCCTCATCGC R- CTTCCACCCCATGGCTTGATACCGTC | 52 | 596 | [65] |
| FLO5 | Adhesion | F- GACAATTGCACACCACTGC F- CCTGTCATTTCTAGGGTTACG | 52 | 423 | [65] |
| FLO10 | Adhesion | F- AACTGGTACCTACACATTTGGC R- GGCAATACCACACTAACAGG | 52 | 270 | [65] |
| FLO11 | Adhesion | F- CCTTGTTCAACTGGTACTGGCG R- TAGAATACAACTGGAAGAGCG | 52 | 749 | [65] |
| AGA1 | Adhesion | F- GTGACGATAACCAAGACAAACGATGCAA R- CCGTTTCATGCATACTGGTTAATGTGCT | 64 | 1198 | [60] |
| District | Sample | pH | Moisture Content (%) | Viscosity (cP) | Titratable Acidity (%) | Microbial Load (log10 cfu/mL or gm) |
|---|---|---|---|---|---|---|
| East Sikkim | Dahi (n = 10) | 3.88 ± 0.01 to 4.84 ± 0.01 | 86.54 ± 0.24 to 96.95 ± 0.58 | 176.55 ± 0.001 to 493.55 ± 0.01 | 0.80 ± 0.01 to 0.91 ± 0.01 | 4.73 ± 0.10 to 7.46 ± 0.02 |
| Chhurpi (n = 10) | 3.95 ± 0.01 to 5.13 ± 0.01 | 55.79 ± 0.59 to 67.42 ± 2.20 | 130.09 ± 0.04 to 175.20 ± 0.05 | 0.63 ± 0.04 to 0.92 ± 0.01 | 5.17 ± 0.02 to 7.48 ± 0.02 | |
| West Sikkim | Dahi (n = 10) | 4.32 ± 0.01 to 4.49 ± 0.01 | 93.86 ± 0.02 to 96.78 ± 0.02 | 428.16 ± 0.04 to 487.71 ± 0.05 | 0.83 ± 0.02 to 0.91 ± 0.02 | 6.29 ± 0.05 to 7.31 ± 0.02 |
| Chhurpi (n = 10) | 4.63 ± 0.01 to 5.13 ± 0.01 | 58.01 ± 0.66 to 61.37 ± 0.49 | 131.56 ± 0.02 to 139.95 ± 0.04 | 0.67 ± 0.03 to 0.79 ± 0.02 | 5.45 ± 0.02 to 6.66 ± 0.02 | |
| North Sikkim | Dahi (n = 10) | 4.13 ± 0.01 to 4.48 ± 0.01 | 93.09 ± 1.65 to 97.2 ± 0.07 | 230.19 ± 0.03 to 485.59 ± 0.04 | 0.83 ± 0.02 to 0.91 ± 0.01 | 5.1 ± 0.05 to 7.46 ± 0.02 |
| Chhurpi (n = 10) | 4.15 ± 0.01 to 5.14 ± 0.01 | 55.94 ± 0.35 to 60.94 ± 0.02 | 175.23 ± 0.02 to 135.55 ± 0.05 | 0.66 ± 0.02 to 0.88 ± 0.01 | 5.27 ± 0.02 to 6.65 ± 0.01 | |
| South Sikkim | Dahi (n = 10) | 4.53 ± 0.01 to 4.53 ± 0.01 | 96.78 ± 0.02 to 96.80 ± 0.03 | 485.33 ± 0.23 to 487.64 ± 0.08 | 0.85 ± 0.04 to 0.88 ± 0.03 | 6.33 ± 0.04 to 6.36 ± 0.01 |
| Chhurpi (n = 10) | 4.02 ± 0.01 to 4.47 ± 0.01 | 56.91 ± 0.04 to 59.58 ± 0.04 | 130.69 ± 0.15 to 140.68 ± 0.15 | 0.83 ± 0.01 to 0.86 ± 0.02 | 6.36 ± 0.01 to 7.46 ± 0.03 |
| Temperature | pH | Nitrate Reduction Test | H2S Production Test | Sugar Fermentation | Tentative Identity (No. of Isolates) (%) | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 25 °C | 30 °C | 37 °C | 40 °C | 45 °C | pH 2.0 | pH 3.0 | pH 4.0 | Lactose | Maltose | Glucose | Galactose | Arabinose | Mannose | Rhamnose | Raffinose | Ribose | Xylose | Sucrose | Trehalose | Mellibiose | |||
| +(13) | +(13) | +(13) | +(3) −(10) | +(1) −(12) | +(13) | +(13) | +(13) | −(13) | +(13) | −(13) | +(13) | +(13) | +(13) | +(13) | +(13) | +(13) | +(13) | +(13) | −(13) | +(13) | +(13) | −(13) | Saccharomyces (13) (65%) |
| +(3) | +(3) | +(3) | −(3) | −(3) | +(3) | +(3) | +(3) | −(3) | +(1) v(2) | −(3) | +(3) | +(3) | +(3) | +(3) | −(3) | +(3) | +(3) | v(3) | +(3) | +(3) | +(3) | v(3) | Pichia (3) (15%) |
| +(2) | +(2) | +(2) | +(1) −(1) | +(1) −(1) | +(2) | +(2) | +(2) | +(2) | +(2) | +(2) | +(2) | +(2) | +(2) | −(2) | −(2) | +(2) | +(2) | +(2) | −(2) | +(2) | +(2) | −(2) | Kluyveromyces (2) (10%) |
| +(2) | +(2) | +(2) | −(2) | −(2) | +(2) | +(2) | +(2) | −(2) | −(2) | −(2) | +(2) | +(2) | +(2) | +(2) | v(2) | +(2) | +(2) | v(2) | −(2) | +(2) | +(2) | v(2) | Debaryomyces (2) (10%) |
| Product | Isolate Code | Identity | Type Species (% Similarity) | GenBank Accession Number |
|---|---|---|---|---|
| Dahi | DAO-17 | Saccharomyces cerevisiae | Saccharomyces cerevisiae NRRL Y-12632 (99.07) | OP071251 |
| DJT-2 | Saccharomyces cerevisiae | Saccharomyces cerevisiae NRRL Y-12632 (99.83) | OP071252 | |
| DKL-13 | Saccharomyces cerevisiae | Saccharomyces cerevisiae NRRL Y-12632 (99.83) | OP071253 | |
| DMS-15 | Saccharomyces cerevisiae | Saccharomyces cerevisiae NRRL Y-12632 (100.00) | OP071254 | |
| DPA-41 | Kluyveromyces marxianus | Kluyveromyces marxianus NRRL Y-8281 (99.82) | OP071255 | |
| DRC-41 | Saccharomyces cerevisiae | Saccharomyces cerevisiae NRRL Y-12632 (99.64) | OP071256 | |
| DRC-42 | Saccharomyces cerevisiae | Saccharomyces cerevisiae NRRL Y-12632 (99.80) | OP071257 | |
| DTL-6 | Kluyveromyces lactis | Kluyveromyces lactis NRRL Y-8279 (100.00) | OP071258 | |
| Chhurpi | CPA-55 | Debaryomyces prosopidis | Debaryomyces prosopidis JCM 9913 (100.00) | OP479882 |
| CCG-4 | Debaryomyces prosopidis | Debaryomyces prosopidis JCM 9913 (100.00) | OP479875 | |
| CGI-29 | Saccharomyces cerevisiae | Saccharomyces cerevisiae NRRL Y-12632 (99.43) | OP479876 | |
| CKL-10 | Saccharomyces cerevisiae | Saccharomyces cerevisiae NRRL Y-12632 (99.19) | OP479877 | |
| CMB-19 | Kazachstania exigua | Kazachstania exigua CBS 379 (99.81) | OP479878 | |
| CMN-3 | Meyerozyma guilliermondii | Meyerozyma guilliermondii NRRL Y-2075 (99.55) | OP479879 | |
| CMNB-4 | Kazachstania exigua | Kazachstania exigua CBS 379 (99.64) | OP479880 | |
| CMS-3 | Pichia bruneiensis | Pichia bruneiensis CBS 12611 (99.77) | OP479881437 | |
| CPAL-16 | Pichia bruneiensis | Pichia bruneiensis CBS 12611 (99.63) | OP479883 | |
| CRC-35 | Saccharomyces cerevisiae | Saccharomyces cerevisiae NRRL Y-12632 (100) | OP479884 | |
| CBG-9 | Yarrowia lipolytica | Yarrowia lipolytica NRRL YB-423 (99.19) | OP479874 | |
| CNT-3 | Pichia kudriavzevii | Pichia kudriavzevii NRRL Y-5396 (100.00) | OP561448 |
| Product | Yeast | Hydrophobicity (%) | Co-Aggregation (%) | Auto-Aggregation(%) | Antimicrobial Activity | BSH Activity | Lysozyme Tolerance (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Escherichia coli KL96 MTCC 1583 | Salmonella enterica subsp. enterica ser. typhimurium MTCC 3223 | Staphylococcus aureus subsp. aureus MTCC 740 | Bacillus cereus MTCC 1272 | Escherichia coli KL96 MTCC 1583 | Salmonella enterica subsp. enterica ser. typhimurium MTCC 3223 | Staphylococcus aureus subsp. aureus MTCC 740 | Bacillus cereus MTCC 1272 | Sodium Cholate | Sodium Taurocholate | Sodium Glycocholate | |||||
| Dahi | Saccharomyces cerevisiae DAO-17 | 92.98 | 55.38 | 73.08 | 49.00 | 77.62 | 91.06 | − | − | ++ | − | − | − | + | 88 |
| Saccharomyces cerevisiae DJT-2 | 97.54 | 59.29 | 73.98 | 70.00 | 67.38 | 65.55 | − | − | − | − | − | − | + | 91 | |
| Saccharomyces cerevisiae DKL-13 | 81.95 | 57.00 | 54.00 | 44.32 | 31.53 | 92.73 | ++ | − | ++ | ++ | − | − | − | 82 | |
| Saccharomyces cerevisiae DMS-15 | 80.08 | 72.30 | 68.88 | 57.60 | 68.05 | 90.97 | − | − | − | − | − | − | − | 79 | |
| Kluyveromyces marxianus DPA-41 | 87.50 | 38.27 | 58.57 | 38.38 | 45.11 | 85.25 | − | − | − | − | ++ | ++ | ++ | 85 | |
| Saccharomyces cerevisiae DRC-41 | 81.25 | 58.08 | 27.25 | 45.77 | 42.23 | 69.42 | − | − | − | − | − | − | − | 75 | |
| Saccharomyces cerevisiae DRC-42 | 91.85 | 23.00 | 64.86 | 50.24 | 42.48 | 93.88 | − | − | − | − | − | − | − | 80 | |
| Kluyveromyces lactis DTL-6 | 94.28 | 43.59 | 64.54 | 52.22 | 38.57 | 82.45 | − | − | − | − | − | − | − | 88 | |
| Chhurpi | Debaryomyces prosopidis CPA-55 | 98.33 | 45.29 | 72.50 | 36.10 | 71.63 | 76.42 | − | − | − | − | − | − | + | 85 |
| Debaryomyces prosopidis CCG-4 | 81.85 | 61.72 | 54.34 | 59.53 | 64.28 | 61.25 | − | ++ | − | − | − | − | − | 93 | |
| Saccharomyces cerevisiae CGI-29 | 80.55 | 82.22 | 73.80 | 41.77 | 78.13 | 91.69 | − | − | − | − | − | − | + | 95 | |
| Saccharomyces cerevisiae CKL-10 | 82.03 | 74.35 | 56.72 | 24.55 | 65.98 | 65.69 | ++ | ++ | ++ | ++ | − | − | + | 93 | |
| Kazachstania exigua CMB-19 | 80.05 | 74.01 | 64.13 | 35.60 | 70.16 | 68.36 | ++ | ++ | ++ | ++ | − | − | − | 89 | |
| Meyerozyma guilliermondii CMN-3 | 80.85 | 79.47 | 54.70 | 36.41 | 69.39 | 68.44 | − | − | − | − | + | ++ | ++ | 91 | |
| Kazachstania exigua CMNB-4 | 81.85 | 70.61 | 65.24 | 38.88 | 70.90 | 69.24 | ++ | ++ | ++ | ++ | − | − | − | 88 | |
| Pichia bruneiensis CMS-3 | 94.32 | 82.19 | 62.80 | 66.41 | 62.17 | 75.26 | − | − | − | − | − | − | − | 85 | |
| Pichia bruneiensis CPAL-16 | 92.85 | 82.04 | 77.97 | 65.17 | 75.03 | 71.25 | − | − | ++ | ++ | − | − | − | 89 | |
| Saccharomyces cerevisiae CRC-35 | 84.04 | 79.36 | 68.01 | 51.46 | 76.94 | 68.42 | − | − | − | − | − | − | + | 87 | |
| Yarrowia lipolytica CBG-9 | 88.40 | 75.21 | 49.36 | 62.32 | 68.33 | 72.42 | − | ++ | − | − | − | − | − | 89 | |
| Pichia kudriavzevii CNT-3 | 90.13 | 81.01 | 57.19 | 77.24 | 73.25 | 75.42 | + | + | − | − | ++ | ++ | ++ | 84 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lama, S.; Tamang, J.P. Isolation of Yeasts from Some Homemade Fermented Cow-Milk Products of Sikkim and Their Probiotic Characteristics. Fermentation 2022, 8, 664. https://doi.org/10.3390/fermentation8120664
Lama S, Tamang JP. Isolation of Yeasts from Some Homemade Fermented Cow-Milk Products of Sikkim and Their Probiotic Characteristics. Fermentation. 2022; 8(12):664. https://doi.org/10.3390/fermentation8120664
Chicago/Turabian StyleLama, Sonam, and Jyoti Prakash Tamang. 2022. "Isolation of Yeasts from Some Homemade Fermented Cow-Milk Products of Sikkim and Their Probiotic Characteristics" Fermentation 8, no. 12: 664. https://doi.org/10.3390/fermentation8120664