1. Introduction
Some migratory birds cover astounding distances, e.g., up to 96,000 km per year for the Arctic tern [
1]. This performance is particularly noteworthy for birds that have a reduced weight and must face headwinds and sometimes-challenging weather conditions. To cover such distances, some migratory birds have developed either remarkable physiological capacities or collaborative strategies that allow them to fly in flocks to minimize their energy expenditure [
2,
3,
4]. Among these strategies, we could mention the V-shaped formation so characteristic of migratory bird flights [
5,
6]. Research has shown that flying in an organized group provides an energy advantage to birds using the eddy areas generated by their wings [
2,
3,
4,
7,
8]. However, a definitive understanding of the aerodynamic implications of these formations is still a matter of considerable debate. The aerodynamic models—developed as knowledge was gained—are of great interest to biologists seeking to understand the strategies and constraints of wild bird migration.
When birds fly in a V formation, their wings produce vortices that are generated due to pressure differences on the upper and lower sides of the wings. These wake vortices generate an upwash outside the wing tips and a downwash inside the wing tips [
7,
9]. Extensive scientific literature has suggested that migratory birds judiciously place their wingtips in upwash areas to minimize their energy expenditure [
4,
6,
10]. Many studies reported decreases in drag forces during V-flight, which could be related to the drafting technique, an aerodynamic phenomenon well-known in the field of sports [
11,
12,
13]. However, a decrease in drag forces can also be accompanied by a decrease in lift, the force required to sustain the bird in flight. Flying animals, unlike aircraft, generate both lift and thrust by flapping their wings. However, many birds also use gliding, i.e., flying without flapping their wings [
14]. More than 300 species of birds from various taxonomic groups undertake long migratory journeys alternating between flapping and gliding modes [
15]. Even today, measuring the aerodynamic coefficients of flying birds remains challenging. The evolution of the drag and lift of a bird is most often estimated from theoretical mathematical models. Experimental studies attempting to unravel the secrets of the aerodynamics of flying birds are scarce due to the difficulties involved in setting up experiments. However, some authors have applied the digital particle image velocimetry (DPIV) technique to analyses of bird flight [
16,
17]. This technique requires the use of a low turbulence wind tunnel and a cooperative and well-trained bird. One alternative to experimental techniques is computational fluid dynamics (CFD). Numerical simulations are often less expensive and restrictive than laboratory experiments and provide information on all flow quantities, including those that are not usually available from measurements. CFD has been used by scientists in an attempt to improve our knowledge of the mechanics of bird flight. Maeng et al. [
18] performed a numerical study on the energy conservation of Canada Geese in flight by modeling wing flapping dynamics. Beaumont et al. [
19] modeled a flapping wing of a Canada goose and studied the effects of altitude on the aerodynamic forces exerted on the wing. More recently, they used CFD to highlight the three-dimensional vortex structures developing in the wake of a Canada goose [
20].
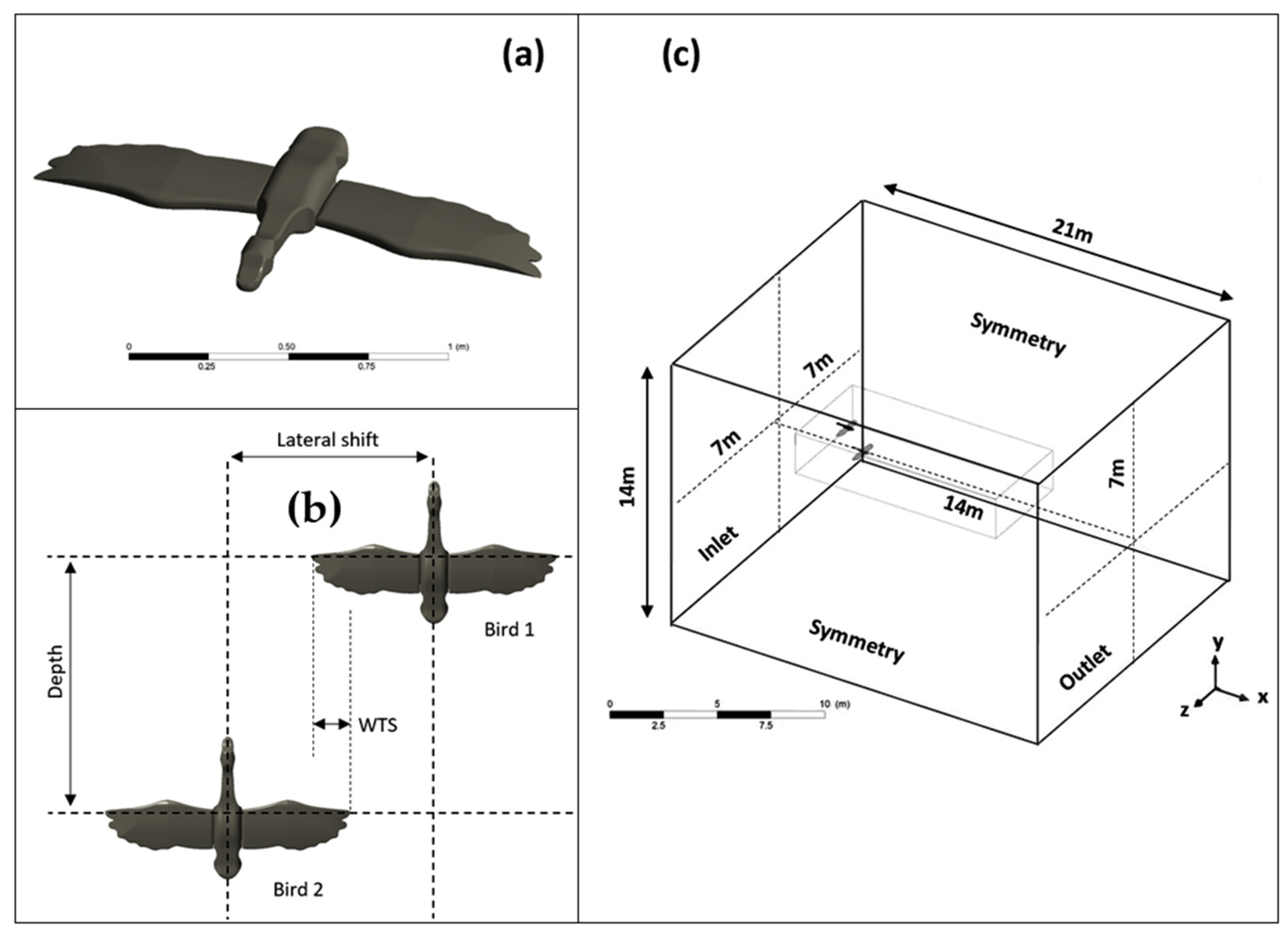
In this study, we evaluated the optimal position of Canada geese in a V-shaped formation. We investigated the influence of wing tip spacing (WTS) and depth on the aerodynamic forces exerted on two birds during gliding flight. We analyzed the vortex wake, the structure of which varied according to the positions of the birds in relation to each other. A computational code, based on the finite volume method, was used to highlight three-dimensional vortex structures developing in the wake of Canada geese flying at an altitude of 1000 m and a speed of 13.9 m/s. Ultimately, the results obtained for 2 birds could easily be extrapolated to a larger bird flock.
3. Results
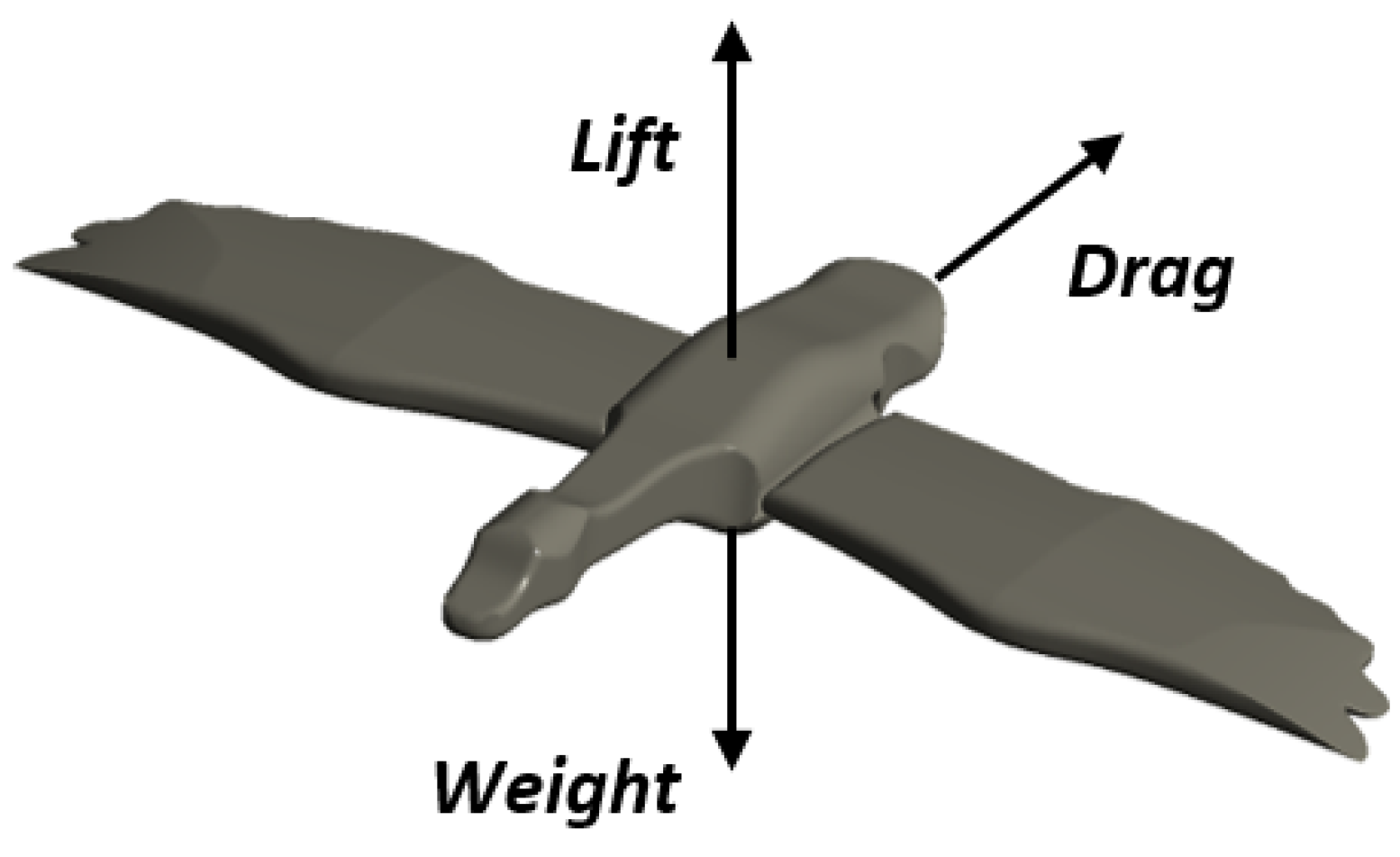
There are four different forces acting on a flying device in flight, whether it is a bird, a bat, an insect or an airplane: lift, thrust, drag and gravity (
Figure 3). Thrust must be equal to drag and lift must be equal to gravity in straight and level flight. In flight aerodynamics, several parameters can be used to quantify the flow characteristics. Hereafter, we presented the most representative ones. Two parameters that provide important information in the study of the aerodynamics of bird wings are the drag and lift coefficients, which are defined as follows:
and are the drag and lift forces; is the density of the fluid, is the forward speed and is the projected area (m2). Lift and drag are two components of the resulting aerodynamic force acting on the wing. Lift can be in any direction relative to gravity, since it is defined relative to the direction of flow rather than the direction of gravity.
When a bird flies straight and level, most of the lift is opposed to gravity [
32]. However, when a bird climbs, descends, or tilts in a turn, the lift is tilted away from the vertical.
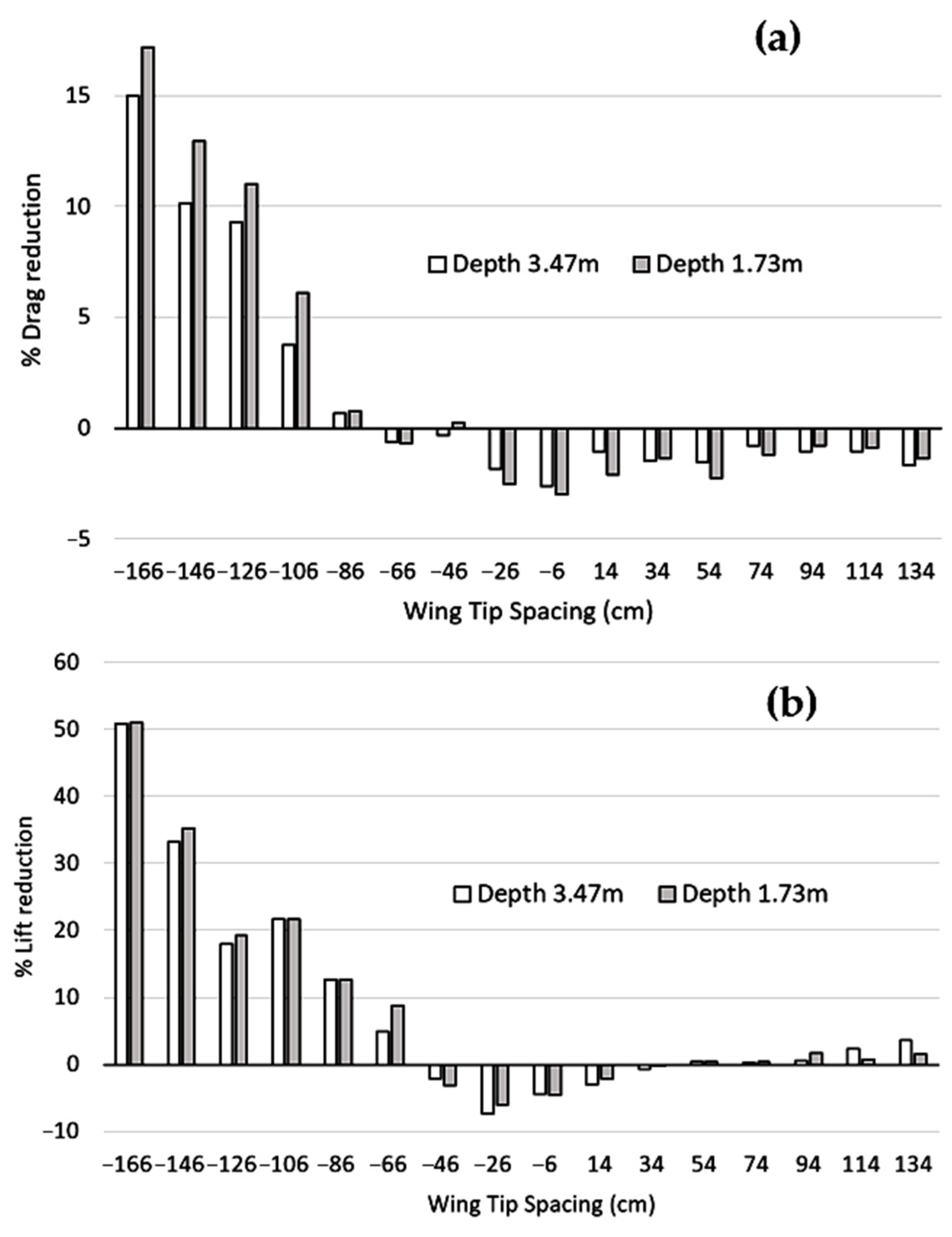
Figure 4 plots reductions or increases (in %) in drag and lift of Bird 2, as compared to Bird alone, as a function of WTS and depth. From this figure, we were able to draw several conclusions. First, a reduction in drag forces was associated with a reduction in lift forces. On the one hand, drag reduction is positive, since it reduces air resistance forces; on the other hand, lift reduction can be a challenge for the bird, since lift must always equal weight. We noted that the reduction of drag and lift forces decreased between −166 and −26 cm. The drag reduction was maximal at −166 cm when the birds were flying in line. In this situation, bird 2 took full advantage of the positive effects of drafting, an aerodynamic principle which consists in reducing air resistance forces by flying in the wake of the preceding bird. We also observed that the reduction of drag forces was higher for a depth of 1.73 m and for a small (or zero) wingtip spacing. For larger WTS, the difference between the two depths (3.47 and 1.73 m) was less significant. On the other hand, the depth seemed to have a smaller impact on lift than on drag.
Our results demonstrated that several forces act on a bird in flight. Flying in formation would allow birds to limit their energy expenditure by reducing aerodynamic drag and improving lift. In this case, it may be appropriate to reason in terms of aerodynamic efficiency, which defines the ratio between lift and drag. The L/D ratio is the amount of lift generated by a wing, divided by the drag it creates while moving through the air.
Generating lift with less drag directly results in better energy economy, better climb performance and better glide.
Figure 5 represents the evolution of the C
L/C
D Ratio for the single bird and for bird 2 as a function of WTS for a depth of 1.73 m and 3.47 m, respectively.
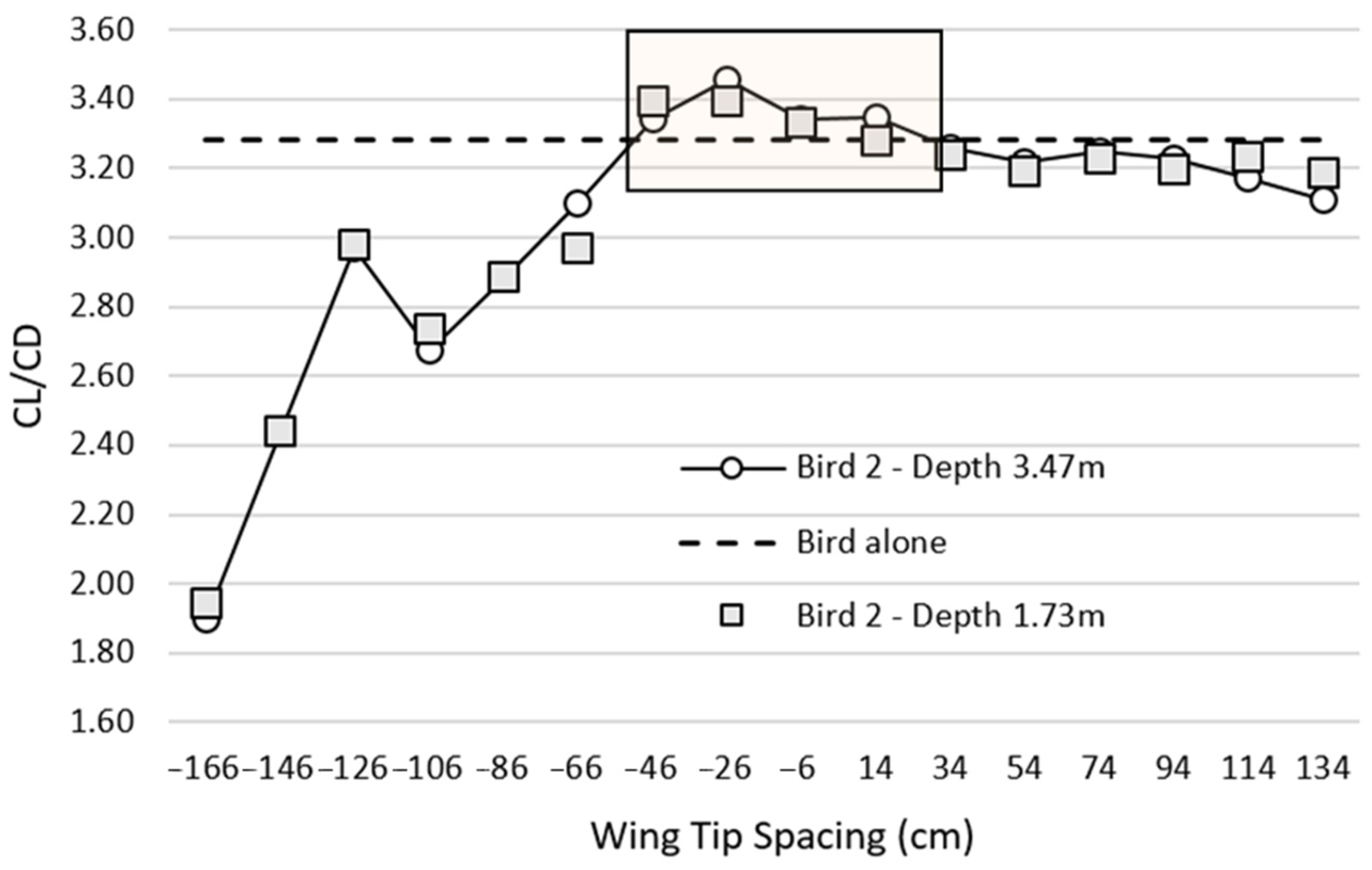
In the present study, the L/D ratio was used to determine the optimal WTS. For the single bird, the L/D ratio was 3.28. Therefore, in order for flocking to be more valuable than flying alone, the value of the L/D ratio of the second bird must be greater than 3.28.
Figure 5 suggests that, compared to flying alone, flying in formation improved the aerodynamic efficiency of the second bird for a WTS between −46 and 14 cm. The most interesting position from an aerodynamic perspective was −26 cm with an L/D = 3.46 ratio 5.5% higher than the L/D ratio of the bird alone. Our study showed that it was necessary to laterally overlap the wingtips of the leader bird and the bird in its wake by 26 cm (WTS = −26 cm) to achieve the best aerodynamic efficiency. Hainsworth [
10] reported that Canada Geese with depths greater than 5 m had a WTS of 54.5 cm and geese with depths less than 5 m had a median WTS of −28.2 cm. Given that, in our study, the depth between the two birds was less than 5 m, our results were consistent with the literature [
10].
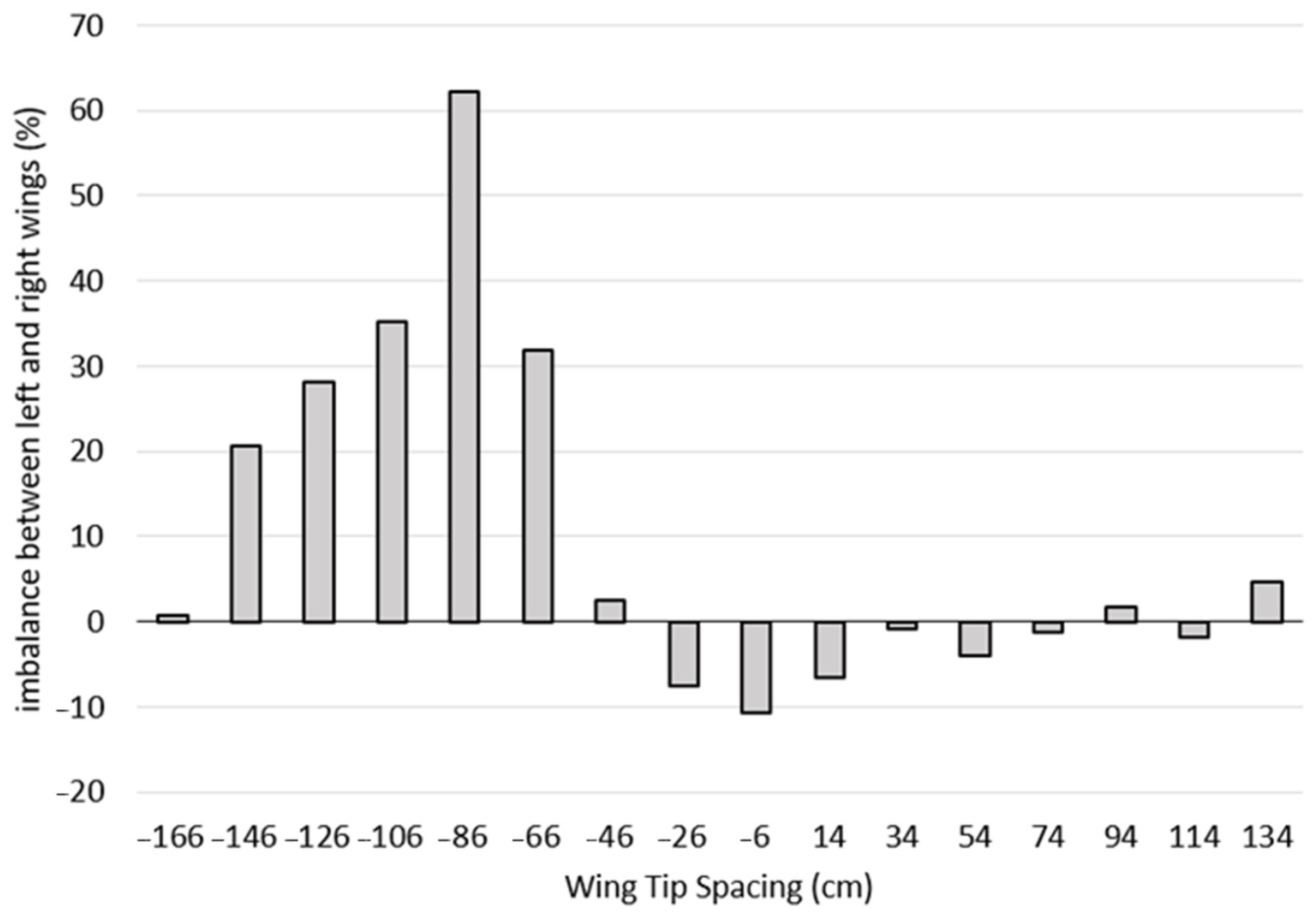
Figure 6 represents the lift imbalance (%) between the right and left wings of the second bird as a function of WTS. The lift imbalance was between 20% and 62% for a WTS between −146 and −66 cm, while the maximum imbalance of 62% was reached at −86 cm.
The lift imbalance remained significant for WTS between −46 and 134 cm but remained below 11%. For WTS, estimated to be optimal at −26 cm, the imbalance between the two wings was 7.6%, although the lift advantage was found at 7.3%. In order to illustrate the force imbalance phenomenon as clearly as possible,
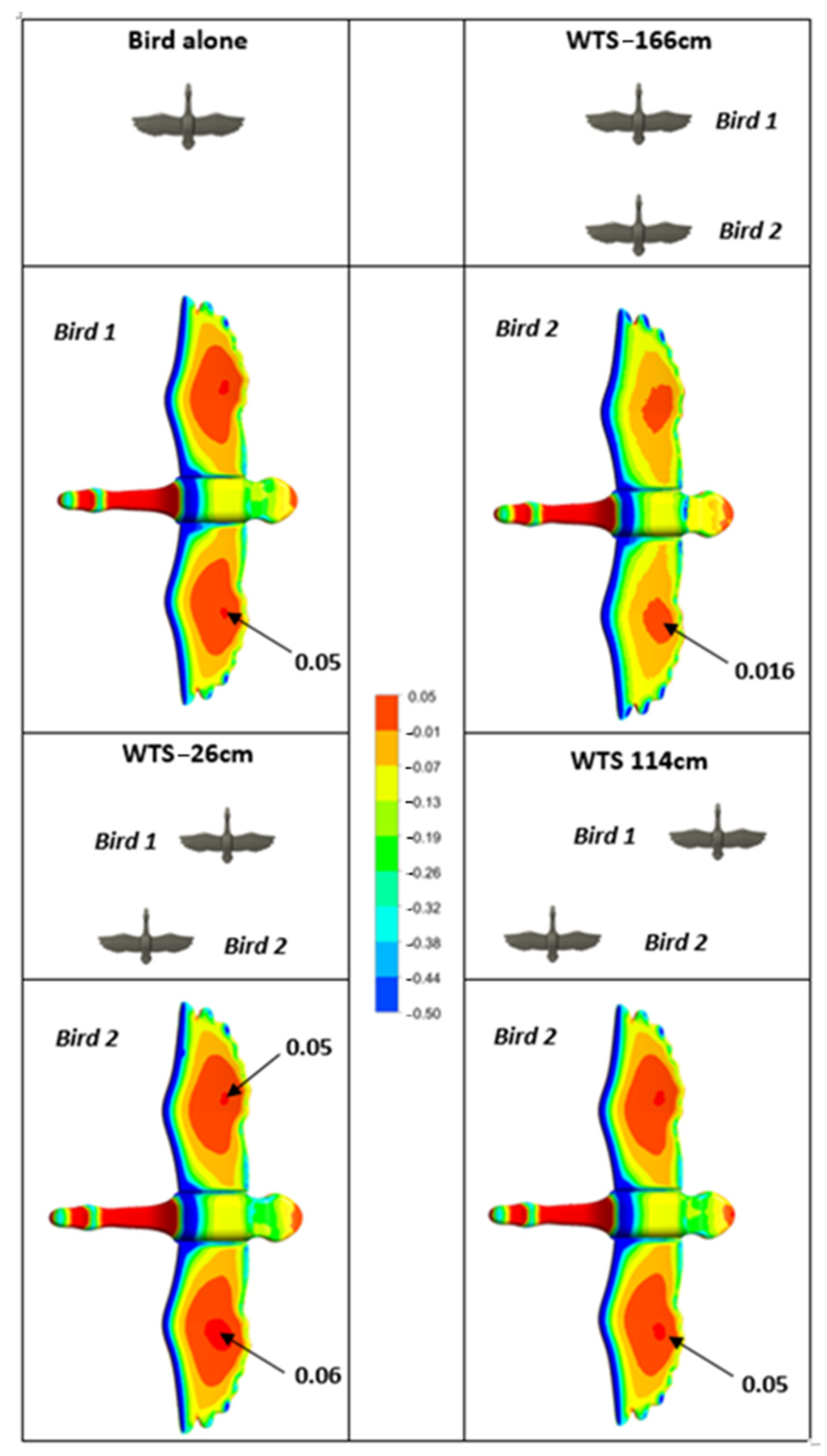
Figure 7 shows a comparison of pressure distributions on the underside of the birds’ wings between the single bird and the second bird for WTS of −26, −166 and 114 cm, respectively.
Note that the pressure coefficient range was purposely limited to the interval [−0.5; 0.05] in order to highlight the slightest change in pressure. The pressure coefficient is defined as follows:
where
is the static pressure,
is the reference static pressure (i.e., atmospheric pressure),
is the air density and
is the speed of the body through the fluid (m.s
−1).
Pressure is the force acting on a given surface. The lift force is transferred by the pressure, which acts perpendicular to the surface of the wing. Thus, the net force is expressed as pressure differences. The direction of the net force implies that the average pressure on the upper surface of the wing is lower than the average pressure on the lower surface [
33]. In summary, the lowest pressure occurs on the top surface of the wing and the highest pressure on the underside of the wing.
The pressure distribution under the birds clearly showed that being sheltered (WTS = −166 cm) implied a decrease in the pressure under the wings of the second bird. This would result in reduced drag forces, but also reduced lift forces, as confirmed in
Figure 4. On the other hand, when the two birds were laterally offset at WTS = −26 cm, we observed a 17% increase in pressure on the underside of the right wing of the second bird. This local increase in pressure improved lift by 7.3%. The distribution of pressures also revealed the pressure imbalance between the right and left wings of bird 2. As mentioned previously, this pressure imbalance induced an imbalance of lift and drag. Beyond a certain offset, the pressure under the wings of the second bird was equivalent to that of the bird alone and the imbalance of forces decreased.
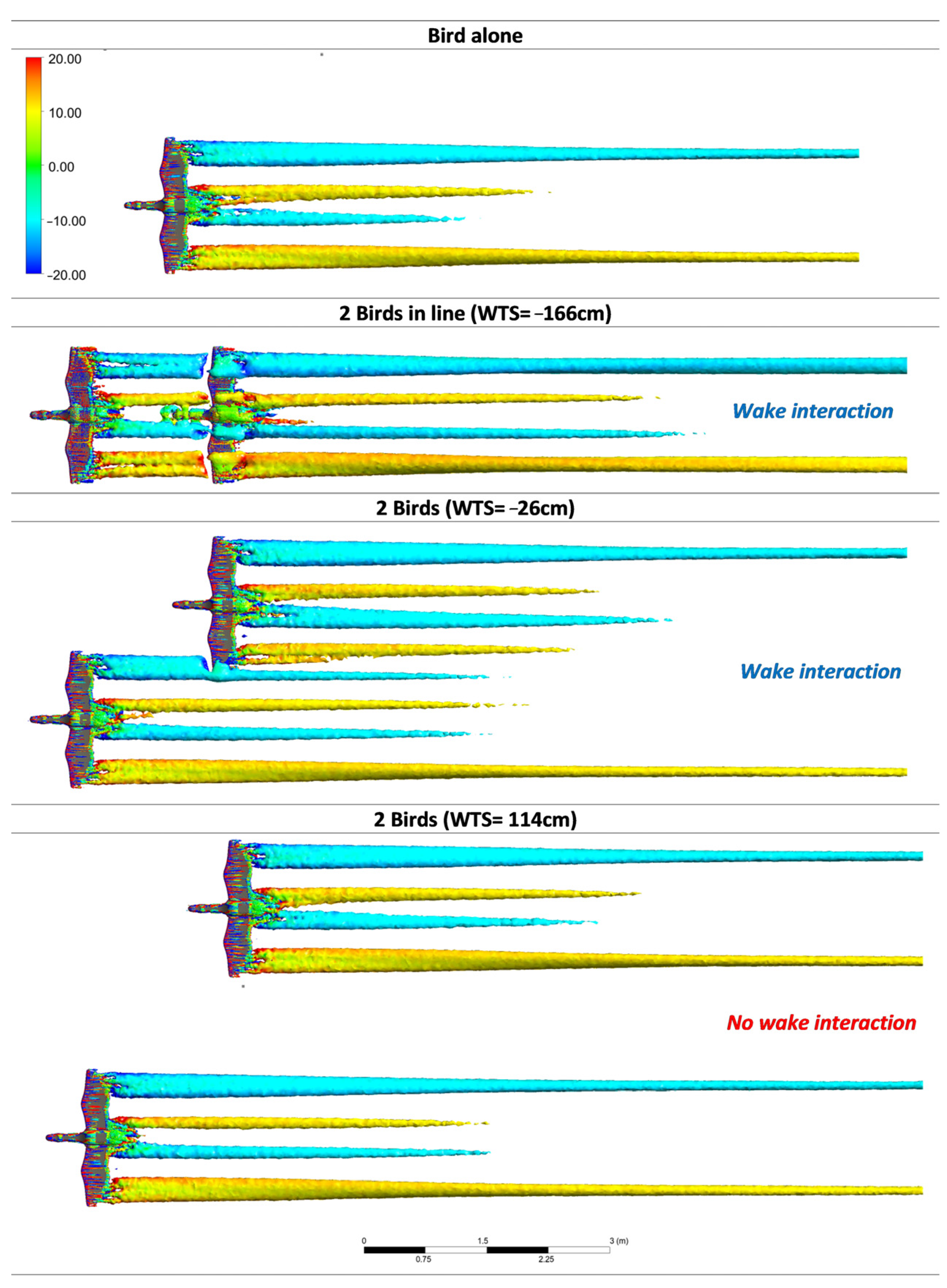
To highlight the structure of the vortex wake,
Figure 8 depicts the vorticity isosurfaces for the bird alone, the birds in line (WTS = −166 cm) and the birds offset by WTS = −26 cm and 114 cm, respectively. The isosurfaces provided insight into the three-dimensional structure of the vortex wake while giving quantitative information about the vortex intensity (vorticity).
Note that the vorticity range (vortex intensity in the direction of the bird’s motion) was purposely limited to the interval [−20; 20] to highlight any change in vortex intensity. The red and blue colors indicated that the vortices had opposite directions of rotation, with one vortex at the wingtip flowing clockwise and the other counterclockwise.
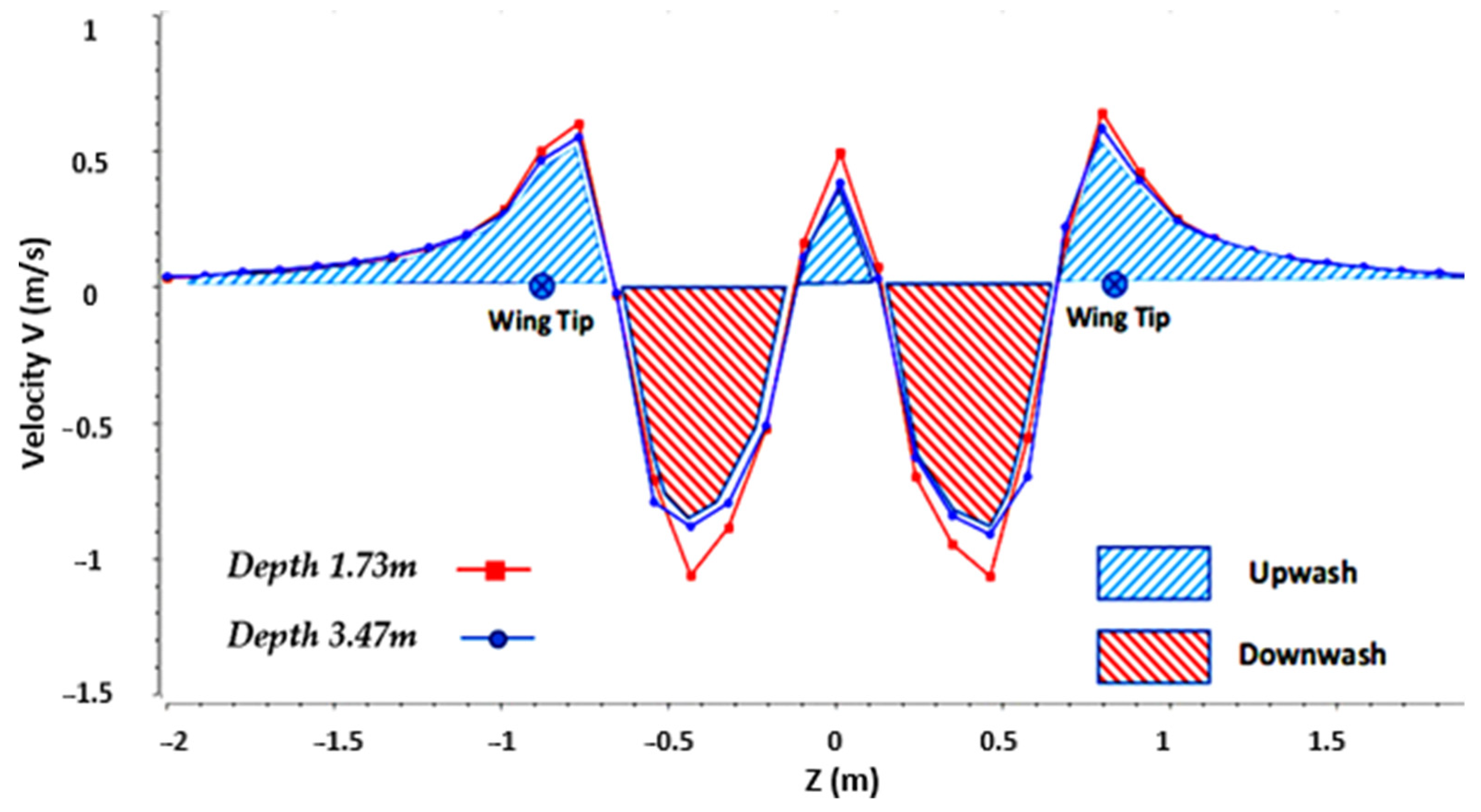
Figure 8 reveals that the wake consisted of a pair of wingtip vortices and a pair of tail vortices. In addition, we plotted (
Figure 9) the vertical velocity profile along an axis passing through the core of the vortex flow for different depths (3.47 m and 1.73 m). The velocity profile (
Figure 9) gave us valuable information about the flow dynamics in the wake of the bird alone. First, as shown in the figure, the absolute velocities were slightly higher for the shortest depth because the vortex energy dissipated as the wake moved away.
The velocity profile in
Figure 9 clearly highlights the upwash zones, in which the vertical velocities are positive, and the downwash zones, in which the vertical velocities are negative.
4. Discussion
The findings of our study revealed that drag was significantly reduced when birds flew in line. In this case, the closer the birds were to each other, the more the drag was reduced (−17% compared to the bird flying alone for a depth of 1.73 m against −15% for a depth of 3.47 m). Another interesting note is that, when the birds flew in a line, the leader bird also gained benefits from the presence of the second bird in its wake, with about an 8% reduction in drag compared to the bird alone. This observation has already been made in other sports such as cycling [
13] or running [
11] and was consistent with the results of Hainsworth [
10], who reported that the leaders also benefited from a reduction in drag, but in smaller proportions than the rest of the group. The results also suggested that the more the birds shifted laterally, relative to each other, the lower the aerodynamic gain. Indeed, birds flying in line in the wake of preceding birds would benefit from a low-pressure zone, allowing them to minimize their drag. In such a case, the birds practice drafting, an aerodynamic phenomenon well known in sports [
11,
13]. As they emerge from this aerodynamic shelter, the birds increase the aerodynamic forces that oppose their movement. Our calculations showed that the optimum offset to obtain a more favorable L/D ratio than that of the bird alone was around −26 cm. This optimal position implied that the tips of the birds’ wings overlapped laterally (see
Figure 8) to obtain better aerodynamic efficiency, as confirmed by Badgerow and Hainsworth [
4].
In this study, we monitored the forces exerted on each of the wings of the birds. This highlighted a phenomenon that has seen little discussion in the literature, yet is evident when birds are observed flying in a V formation. When the bird in the wake of the preceding bird shifts laterally, one of its two wings is exposed to the airflow, which induces an imbalance in the forces applied to the left and right wings. These results, which might seem counter-intuitive at first sight, could be explained by a wake interaction that would induce a local modification of the dynamics of the flow in the vicinity of the birds. This wake interaction will, depending on the position of one bird relative to another, prove beneficial or, on the contrary, be disadvantageous from an aerodynamic point of view. As shown in
Figure 9, the phenomenon was particularly visible for birds in line and for a WTS of −26 cm. Even if the phenomenon were not visible in
Figure 9, the wake of bird 1 continued to disturb the aerodynamic behavior of bird 2 for a significant lateral shift (see
Figure 4). The topology of the flow, as illustrated in this study, was consistent with observations performed on the wake of other gliding bird species, such as the swift [
23]. The pressure differential between the lower and upper surfaces of the wings is at the origin of a reversal of the air flow behind the wing which causes the rotation of the air masses downstream of the ends of the wings. Once the reversal is complete, the wake consists of two main contra-rotating cylindrical vortices. Green [
33] highlighted three mechanisms to describe the origin of wingtip vortices. The first and most common mechanism is pressure imbalance at the tip, during the process of generating lift. The pressure difference accelerates the flow from the pressure side (lower surface) to the suction side (upper surface), which leads to the formation of a wingtip vortex. The strength of the vortex depends on the magnitude of the pressure difference, which in turn depends on the angle of attack. The two wingtip vortices do not merge as they flow in opposite directions. The wingtip vortices rise from the wingtips and tend to dip and roll over each other downstream of the wing. Again, the end vortices eventually dissipate, their energy being transformed by viscosity. These vortices imply that the air located inside the vortex system, in the immediate wake of the bird, moves downwards (downwash), while the air located on the sides, outside the vortex system, moves up (upwash). Theoretically, a bird positioning itself in the upwash zone of the preceding bird would benefit from the upward flow, which would allow it to improve its lift [
3,
8]. The results obtained for the birds in line and those which implied a significant reduction in drag and lift for the second bird could seem counter-intuitive at first sight.
However, if we observe the velocity profile, as shown in
Figure 9, we see that, in absolute value, the velocities in the downwash zone were greater than the velocities in the upwash zone. Thus, when the birds flew one behind the other, part of the body of the second bird was subjected to downward flows of greater intensity than the upward flows, which largely explains the loss of lift estimated by our results. Consequently, the only way for the second bird to benefit from the upwash zones and to improve its lift is to shift laterally to an optimum which corresponds to the peak of positive speed. However, shifting involves an imbalance of forces between the left wing and the right wing. This imbalance is highlighted in
Figure 6 and
Figure 7. To our knowledge, no scientific study has reported this imbalance of forces between the two wings of a migratory bird. We therefore hypothesized that a bird must implement physiological and biomechanical adaptation mechanisms to counter this aerodynamic phenomenon.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}