
Gelation and Orientation Dynamics Induced by Contact of Protein Solution with Transglutaminase Solution
 , and
, and
Abstract
:
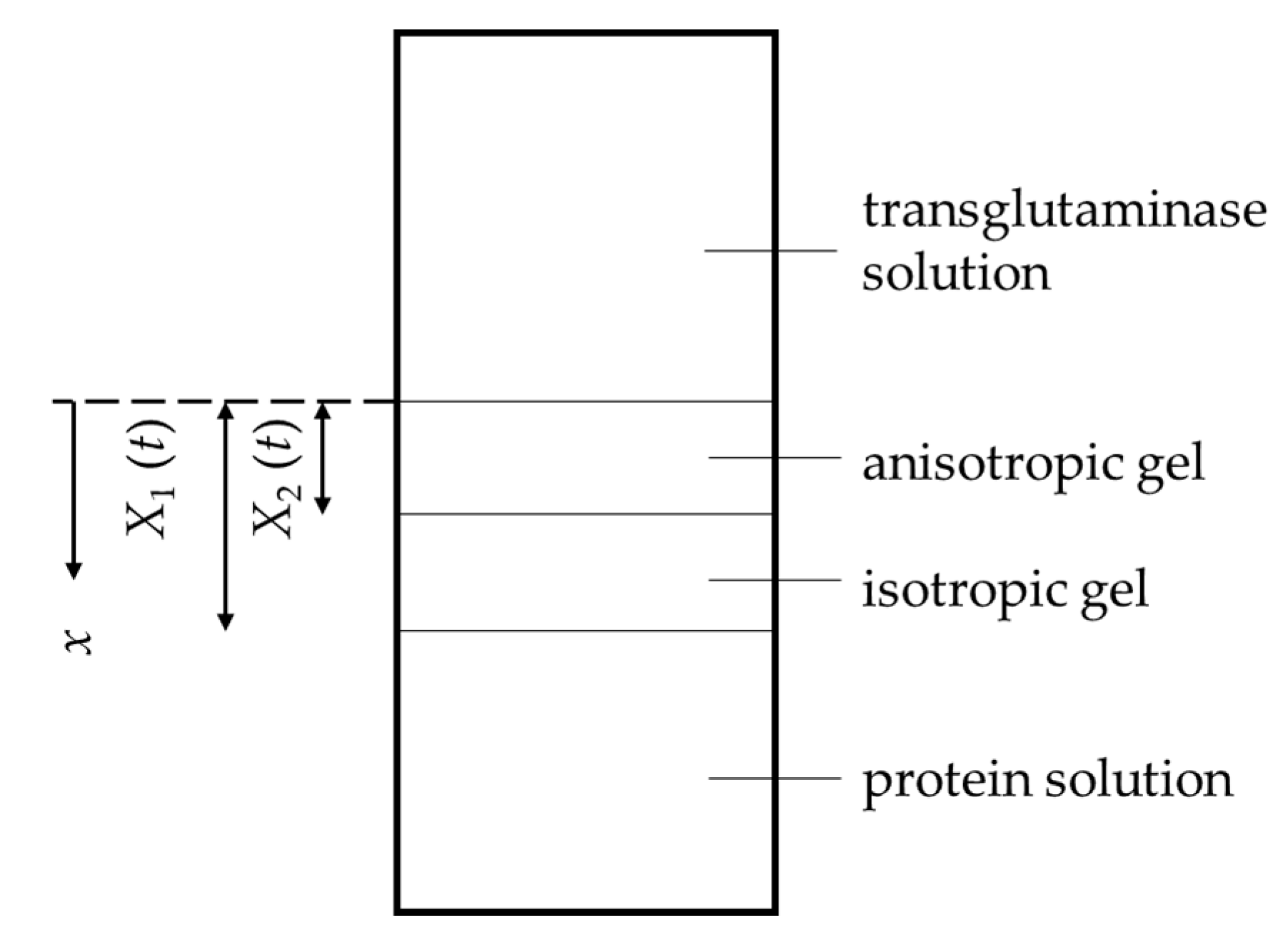
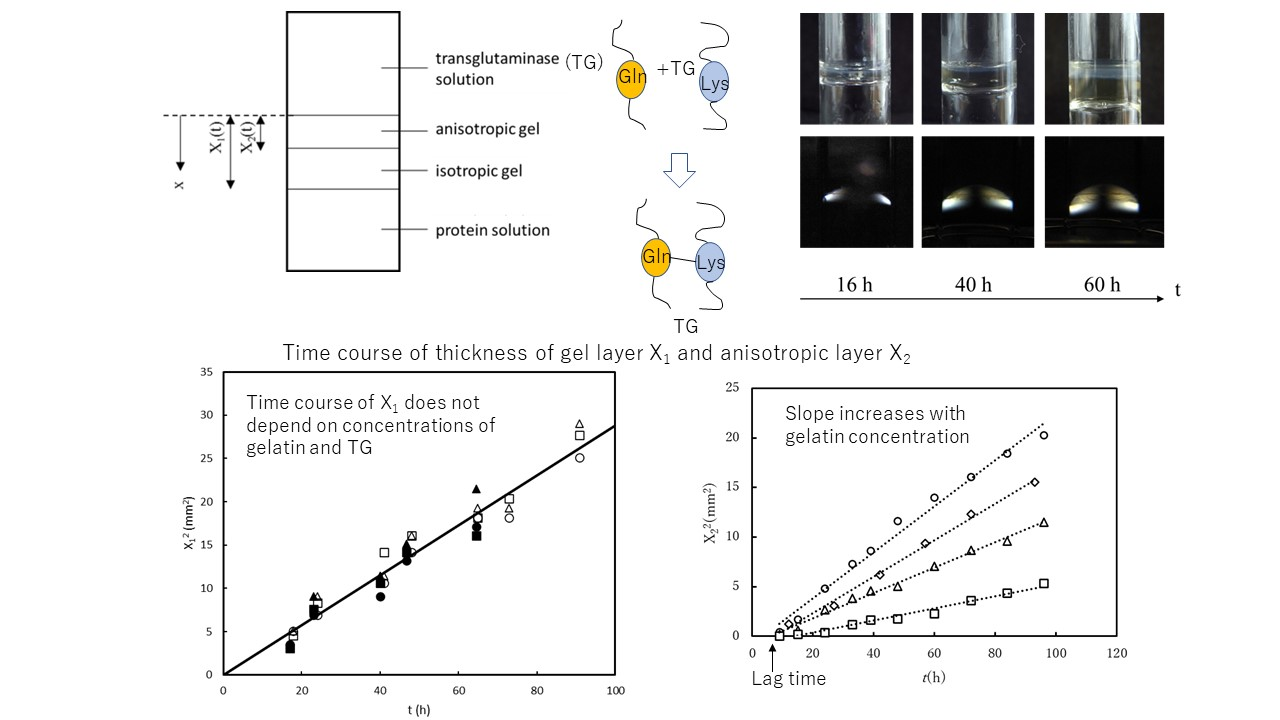{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Theory
2.1. Gelation Dynamics by a Contact of Protein Solution with Transglutaminase Solution
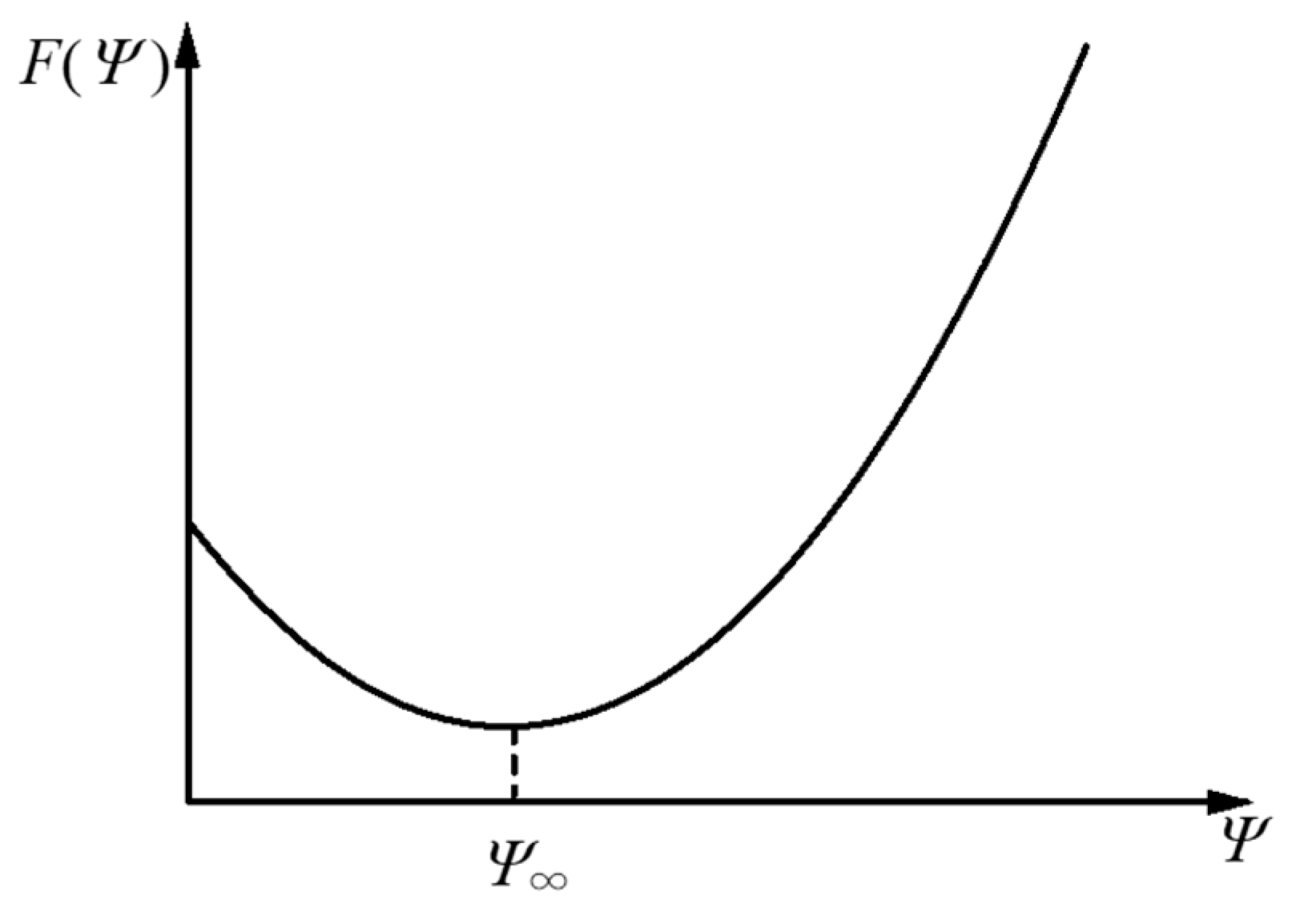
2.2. Orientation Dynamics Based on the Free-Energy Principle
3. Results and Discussion
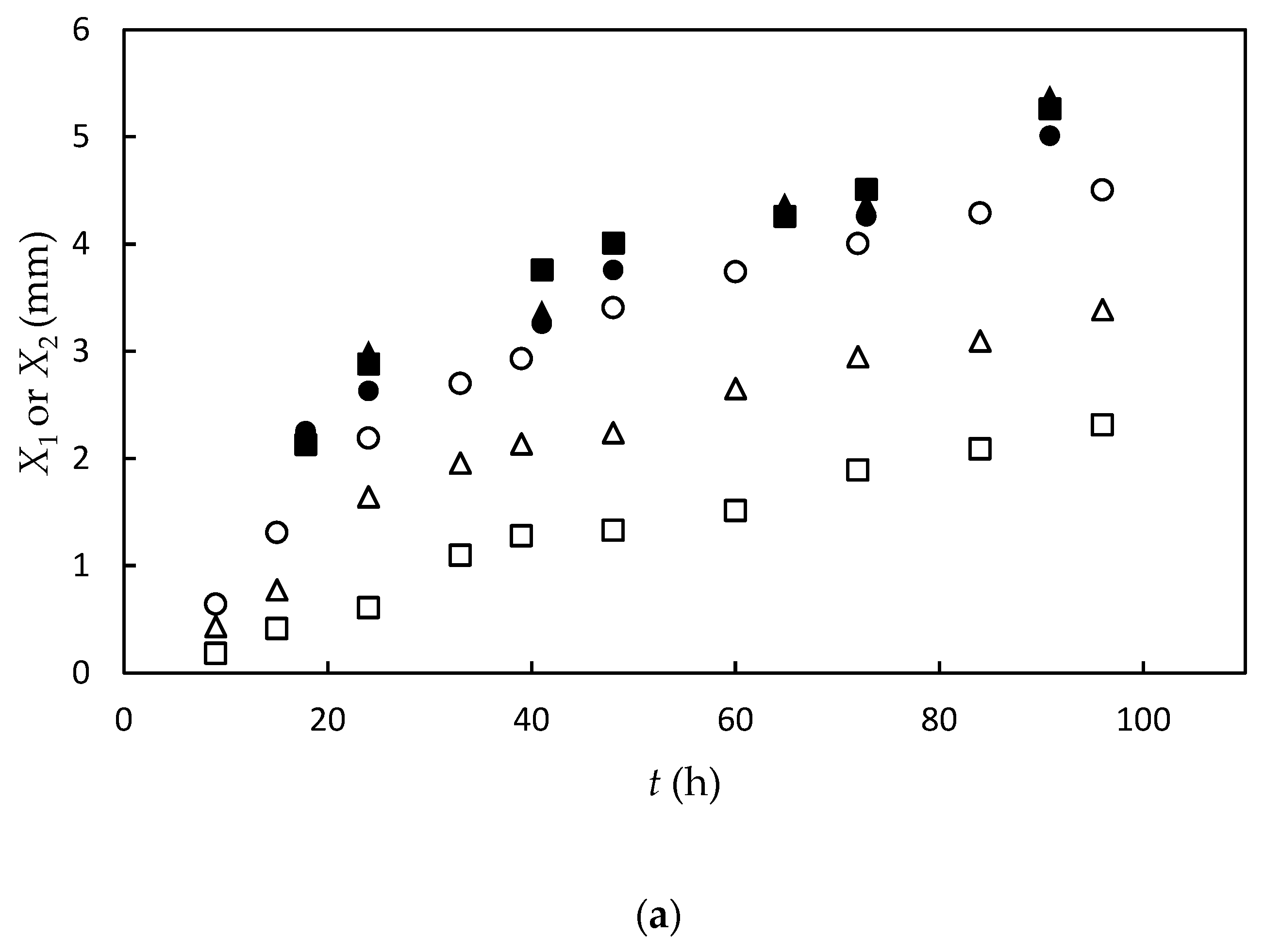
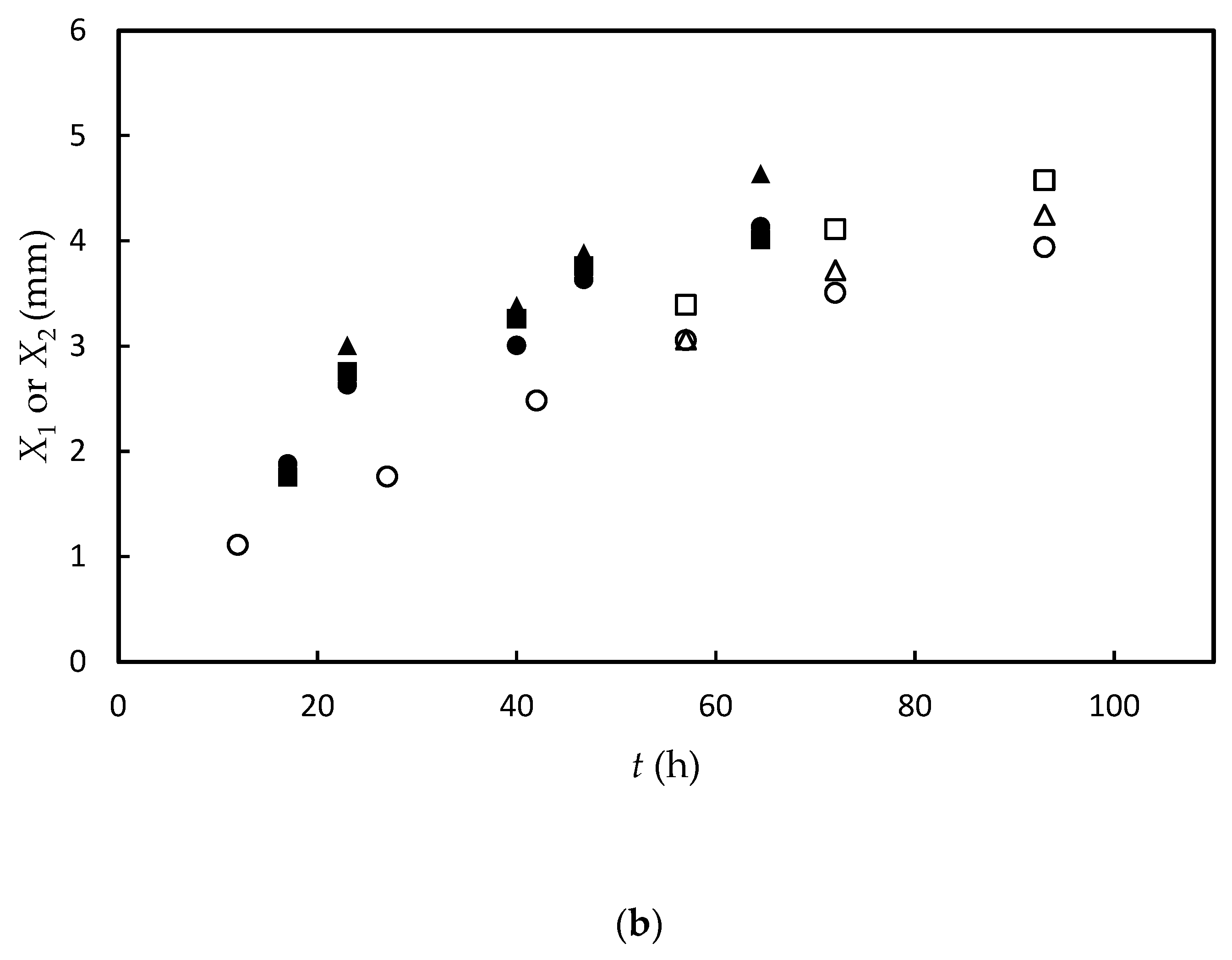
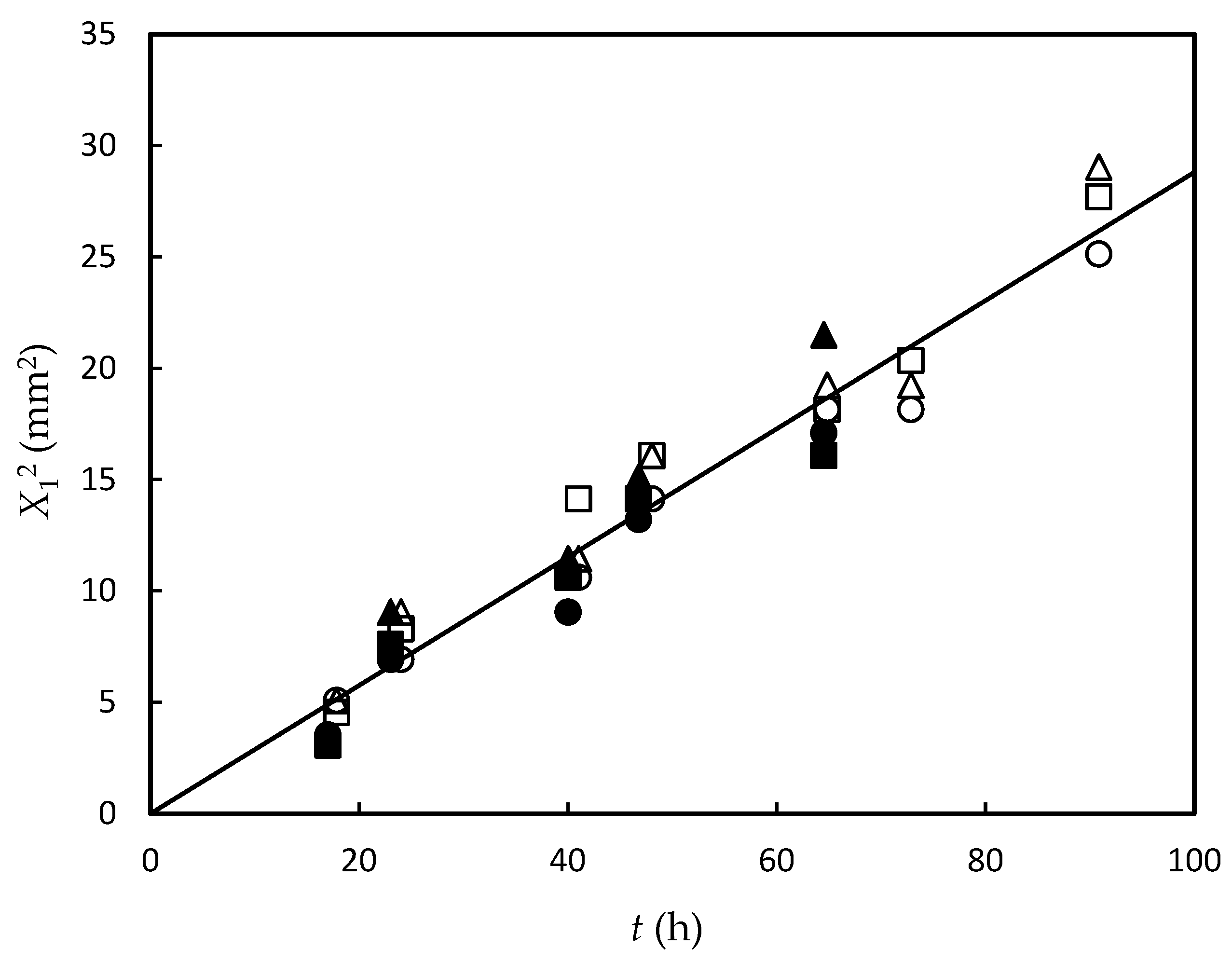
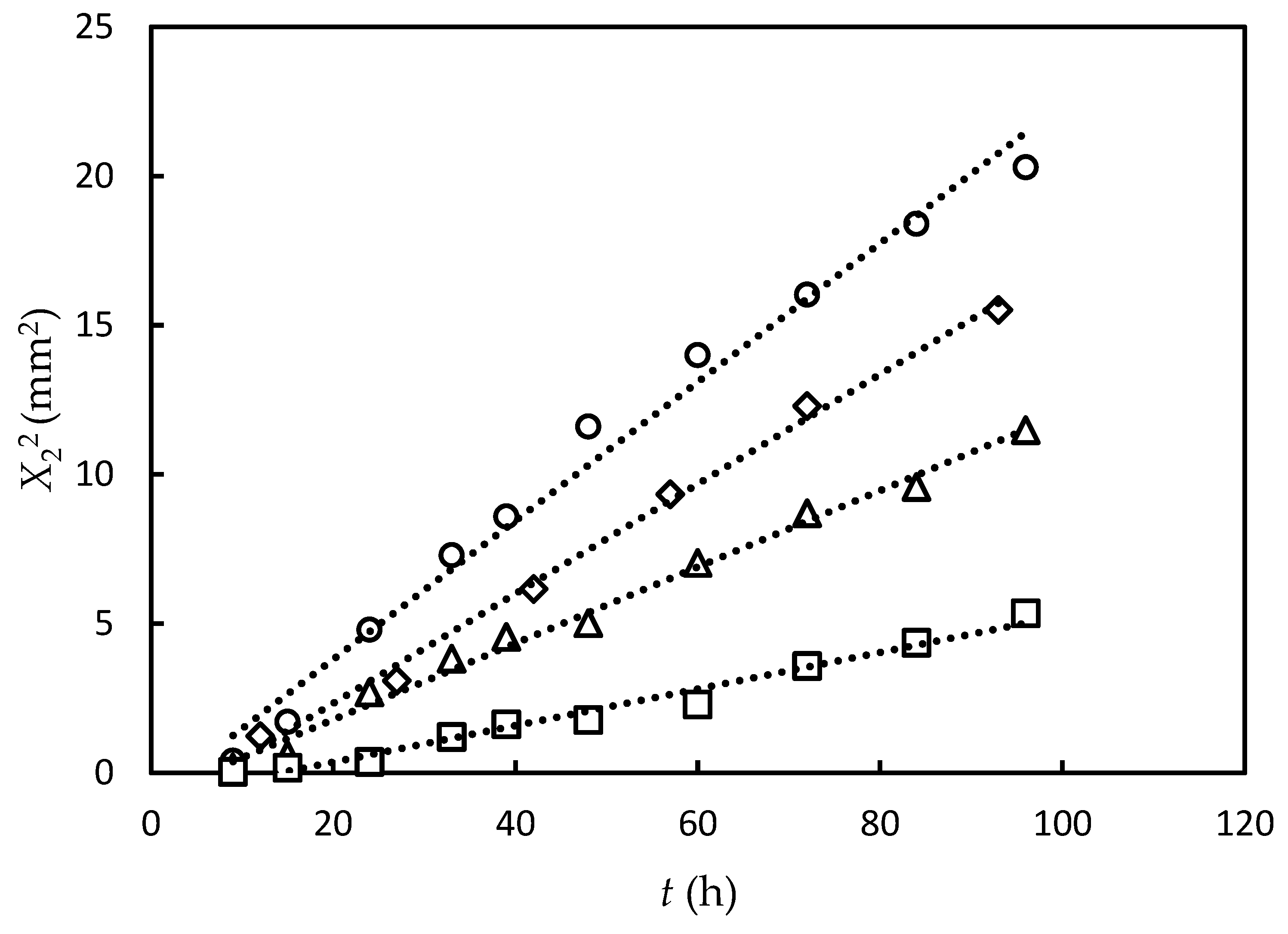
3.1. Experimental Results and Data Fitting
3.2. Discussion about Free-Energy Function and Orientation Dynamics
4. Conclusions
5. Materials and Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Flory, P.J. Principles of Polymer Chemistry; Cornell University Press: Ithaca, NY, USA, 1953. [Google Scholar]
- Gordon, M.; Ross-Murphy, S.B. The structure and properties of molecular trees and networks. Pure Appl. Chem. 1975, 43, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Stockmayer, W.H.; Zimm, B.H. When Polymer Science Looked Easy. Annu. Rev. Phys. Chem. 1984, 35, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Stauffer, J. Gelation in concentrated critically branched polymer solutions. Percolation scaling theory of intramolecular bond cycles. J. Chem. Soc. Faraday Trans. II 1976, 72, 1354–1364. [Google Scholar] [CrossRef]
- De Gennes, P.G. Scaling Concepts in Polymer Physics; Cornell University Press: Ithaca, NY, USA, 1979. [Google Scholar]
- Tanaka, F. Polymer Physics: Applications to Molecular Association and Thermoreversible Gelation; Cambridge University Press: New York, NY, USA, 2011. [Google Scholar]
- Matter, F.; Luna, A.L.; Niederberger, M. From colloidal dispersions to aerogels: How to master nanoparticle gelation. Nano Today 2020, 30, 100827. [Google Scholar] [CrossRef]
- Thiele, H. Ordered Coagulation and Gel Formation. Discuss. Faraday Soc. 1954, 18, 294–314. [Google Scholar] [CrossRef]
- Skjåk-Bræk, G.; Grasdalen, H.; Smisrød, O. Inhomogeneous polysaccharide ionic gels. Carbohydr. Polym. 1989, 10, 31–54. [Google Scholar] [CrossRef]
- Dobashi, T.; Nobe, M.; Yoshihara, H.; Yamamoto, T.; Konno, A. Liquid crystalline gel with refractive index gradient of curdlan. Langmuir 2004, 20, 6530–6534. [Google Scholar] [CrossRef]
- Dobashi, T.; Yamamoto, T. Anisotropic Gel Formation Induced by Dialysis. In Encyclopedia of Biocolloid and Biointerface Science; Ohshima, H., Ed.; Wiley: Hoboken, NJ, USA, 2016; pp. 487–497, Chap. 39. [Google Scholar]
- Maki, Y.; Furusawa, K.; Yamamoto, T.; Dobashi, T. Structure formation in biopolymer gels induced by diffusion of gelling factors. J. Biorheol. 2018, 32, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.C.; Minamisawa, Y.; Furusawa, K.; Maki, Y.; Takeno, H.; Yamamoto, T.; Dobashi, T. Phase relationship and dynamics of anisotropic gelation of carboxymethylcellulose aqueous solution. Colloid Polym. Sci. 2010, 288, 695–701. [Google Scholar] [CrossRef]
- Furusawa, K.; Sato, S.; Masumoto, J.; Hanazaki, Y.; Maki, Y.; Dobashi, T.; Yamamoto, T.; Fukui, A.; Sasaki, N. Studies on the formation mechanism and the structure of the anisotropic collagen gel prepared by dialysis-induced anisotropic gelation. Biomacromolecules 2012, 13, 29–30. [Google Scholar] [CrossRef]
- Narita, T.; Tokita, M. Liesegang pattern formation κ-carrageenan gel. Langmuir 2006, 22, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Maki, Y.; Furusawa, K.; Yasuraoka, S.; Okamura, H.; Hosoya, N.; Sunaga, M.; Dobashi, T.; Sugimoto, Y.; Wakabayashi, K. Universality and specificity in molecular orientation in anisotropic gels prepared by diffusion method. Carbohydr. Polym. 2014, 108, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Mredha, M.T.I.; Zhang, X.; Nonoyama, T.; Nakajima, T.; Kurokawa, T.; Takagi, Y.; Gong, J.P. Swim bladder collagen forms hydrogel with macroscopic superstructure by diffusion induced fast gelation. J. Mater. Chem. B 2015, 3, 7658–7666. [Google Scholar] [CrossRef] [PubMed]
- Maki, Y.; Ito, K.; Hosoya, N.; Yoneyama, C.; Furusawa, K.; Yamamoto, T.; Dobashi, T.; Sugimoto, Y.; Wakabayashi, K. Anisotropic Structure of Calcium-Induced Alginate Gels by Optional and Small-Angle X-ray Scattering Measurements. Biomacromolecules 2011, 12, 2145–2152. [Google Scholar] [CrossRef]
- Maki, Y.; Furusawa, K.; Dobashi, T.; Sugimoto, Y.; Wakabayashi, K. Small-angle X-ray and light scattering analysis of multi-layered Curdlan gels prepared by a diffusion method. Carbohydr. Polym. 2017, 155, 136–145. [Google Scholar] [CrossRef]
- Rivas-Araziza, R.; Alcouffe, P.; Rochas, C.; Montembault, A.; David, L. Micron range morphology of physical chitosan hydrogels. Langmuir 2010, 26, 17495–17504. [Google Scholar] [CrossRef]
- Yang, W.Y.; Furukawa, H.; Gong, J.P. Highly Extensible Double-Network Gels with Self-Assembling Anisotropic Structure. Adv. Mater. 2008, 20, 4499–4503. [Google Scholar] [CrossRef]
- Despang, F.; Dittrich, R.; Gelinsky, M. Novel Biomaterials with Parallel Aligned Pore Channels by Directed Ionotropic Gelation of Alginate: Mimicking the Anisotropic Structure of Bone Tissue. In Advances in Biomimetics; Chapter 17; George, A., Ed.; Intech: London, UK, 2011. [Google Scholar]
- Mredha, M.T.I.; Kitamura, N.; Nonomiya, T.; Wada, S.; Goto, K.; Zhang, X.; Nakajima, T.; Kurokawa, T.; Takagi, Y.; Yasuda, K.; et al. Anisotropic tough double network hydrogel from fish collagen and its spontaneous in vivo bonding to bone. Biomaterials 2017, 132, 85–95. [Google Scholar] [CrossRef]
- Yahata, S.; Furusawa, K.; Nagao, K.; Nakajima, M.; Fukuda, T. Effects of Three-Dimensional Culture of Mouse Calvaria-Derived Osteoblastic Cells in a Collagen Gel with a Multichannel Structure on the Morphogenesis Behaviors of Engineered Bone Tissues. ACS Biomater. Sci. Eng. 2017, 3, 3414–3424. [Google Scholar] [CrossRef]
- Koh, I.; Furusawa, K.; Haga, H. Anisotropic Multi-channel Collagen Gel (MCCG) Guides the Growth Direction of the Neurite-like Processes of PC12 Cells. Sci. Rep. 2018, 8, 13901. [Google Scholar] [CrossRef] [Green Version]
- Furusawa, K.; Wakamatsu, M.; Dobashi, T.; Yamamoto, T. Adsorption Kinetics of Carcinogens to DNA Liquid Crystalline Gel Beads. Langmuir 2007, 23, 10081–10087. [Google Scholar] [CrossRef]
- Qiao, L.; Du, C.; Gong, J.P.; Wu, Z.L.; Zheng, Q. Programmed diffusion induces anisotropic superstructures in hydrogels with high mechano-optical sensitivity. Adv. Mater. Technol. 2019, 4, 1900665. [Google Scholar] [CrossRef]
- Nobe, M.; Dobashi, T.; Yamamoto, T. Dynamics in Dialysis Process for Liquid Crystalline Gle Formation. Langmuir 2005, 21, 8155–8160. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, K.; Minamisawa, Y.; Dobashi, T.; Yamamoto, T. Dynamics of liquid crystalline gelation of DNA. J. Phys. Chem. B 2007, 111, 14423–14430. [Google Scholar] [CrossRef]
- Yamamoto, T.; Tomita, N.; Maki, Y.; Dobashi, T. Dynamics in the Process of Formation of Anisotropic Chitosan Hydrogel. J. Phys. Chem. B 2010, 114, 10002–10009. [Google Scholar] [CrossRef] [PubMed]
- Rokugawa, I.; Tomita, N.; Dobashi, T.; Yamamoto, T. One-Dimensional Growth of Hydrogel by a Contact of Chitosan Solution with High-pH Solution. Soft Mater. 2014, 12, 36–41. [Google Scholar] [CrossRef]
- Pisano, J.J.; Finlayson, J.S.; Peyton, M.P. Chemical and enzymic detection of protein cross-links. Measurement of epsilon-(gamma-glutamyl)lysine in Fibrin Polymerized by Factor XIII. Biochemistry 1969, 8, 871–876. [Google Scholar] [CrossRef]
- Connellan, J.H.; Chung, S.I.; Whetzel, N.K.; Bradely, C.M.; Folk, J.E. Structural Properties of Guinea Pig Liver Transglutaminase. J. Biol. Chem. 1971, 246, 1093–1098. [Google Scholar] [CrossRef]
- Sieffring, G.H.; Apostol, A.B.; Velasco, P.T.; Lonard, L. Enzymatic Basis for the Ca2+-induced Cross-linking of Membrane Proteins in Intact Human Erythrocytes. Biochemistry 1978, 17, 2598–2604. [Google Scholar] [CrossRef]
- Seki, N.; Uno, K.; Lee, N.H.; Kimura, I.; Toyoda, K.; Fujita, T.; Arai, K. Transglutaminase Activity in Alaska Pollack Muscle and Surimi, and its Reaction with Myosin, B. J. Nippon Suisan Gakkaishi 1990, 56, 125–132. [Google Scholar] [CrossRef]
- McDermott, M.K.; Chen, T.; Williams, C.M.; Markley, K.M.; Payne, G.F. Mechanical Properties of Biomimetic Tissue Adhesive Based on the Microbial Transglutaminase-catalyzed Crosslinking of Gelatin. Biomacromolecules 2004, 5, 1270–1279. [Google Scholar] [CrossRef] [PubMed]
- Ariëns, R.A.S.; Lai, T.-S.; Weisel, J.W.; Greenberg, C.S.; Grant, P.J. Role of Factor XIII in Fibrin Clot Formation and Effects of Genetic Polymorphisms. Blood 2002, 100, 743–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisel, J.W.; Litvinov, R.I. Mechanisms of Fibrin Polymerization and Clinical Implications. Blood 2013, 121, 1712–1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorand, L.; Graham, R.M. Transglutaminases: Crosslinking Enzymes with Pleiotropic Functions. Nat. Rev. Mol. Cell Biol. 2003, 4, 140–156. [Google Scholar] [CrossRef]
- Wilhelmus, M.M.; de Jager, M.; Bakker, E.N.; Drukarch, B. Tissue Transglutaminase in Alzheimer’s Disease: Involvement in Pathogenesis and its Potential as a Therapeutic Target. J. Alzheimer’s Dis. 2014, 42, S289–S303. [Google Scholar] [CrossRef]
- Kuraishi, C.; Yamazaki, K.; Susa, Y. Transglutaminase: Its Utilization in the Food Industry. Food Rev. Int. 2001, 17, 221–246. [Google Scholar] [CrossRef]
- Dobashi, T.; Yamamoto, T. Analysis of heterogeneous gelation dynamics and their application to blood coagulation. Gels 2018, 4, 59. [Google Scholar] [CrossRef] [Green Version]
- Landau, L.D.; Lifshitz, E.M.; Pitaevskii, L.P. Statistical Physics 3rd Edition Part 1; Pergamon Press: Oxford, UK, 1980; Chapter XIV. [Google Scholar]
- Hohenberg, P.C.; Halperin, B.I. Theory of Dynamic Critical Phenomena. Rev. Mod. Phys. 1977, 49, 435–479. [Google Scholar] [CrossRef]
- Hohenberg, P.C.; Krekhov, A.P. An Introduction to the Ginzburg-Landau Theory of Phase Transitions and Nonequilibrium Patterns. Phys. Rep. 2015, 572, 1–42. [Google Scholar] [CrossRef] [Green Version]
- Furusawa, K.; Narazaki, Y.; Tomita, N.; Dobashi, T.; Sasaki, N.; Yamamoto, T. Effect of pH on Anisotropic Gelation of DNA Induced by Aluminum Cations. J. Phys. Chem. B 2010, 114, 13923–13932. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kakinoki, K.; Kurasawa, R.; Maki, Y.; Dobashi, T.; Yamamoto, T. Gelation and Orientation Dynamics Induced by Contact of Protein Solution with Transglutaminase Solution. Gels 2023, 9, 478. https://doi.org/10.3390/gels9060478
Kakinoki K, Kurasawa R, Maki Y, Dobashi T, Yamamoto T. Gelation and Orientation Dynamics Induced by Contact of Protein Solution with Transglutaminase Solution. Gels. 2023; 9(6):478. https://doi.org/10.3390/gels9060478
Chicago/Turabian StyleKakinoki, Kasumi, Ryuta Kurasawa, Yasuyuki Maki, Toshiaki Dobashi, and Takao Yamamoto. 2023. "Gelation and Orientation Dynamics Induced by Contact of Protein Solution with Transglutaminase Solution" Gels 9, no. 6: 478. https://doi.org/10.3390/gels9060478