1. Introduction
Entomopathogenic fungi (EPF) naturally infect insects and some EPF strains have been commercially produced as biopesticides for managing insect pests [
1,
2]. Their field persistence and efficacy may be affected by various abiotic and biotic factors, such as solar radiation, temperature, humidity/moisture levels, surface chemistry, and phylloplane microbiota [
2,
3,
4,
5]. However, widespread infection by EPF may occur under favorable conditions, e.g., high humidity, mild temperature, frequent rainfalls, and high host densities, which promote horizontal transmissions in pest populations, leading to epizootics [
2]. In 2017, a naturally occurring epizootic among whiteflies was observed in southern Georgia where high levels of whitefly infestations were noted in cotton and vegetable production [
6]. The fungus was isolated and identified as a new strain of
Cordyceps javanica (Wf GA17 strain) [
7,
8]. This new strain demonstrated high virulence to the sweetpotato whitefly,
Bemisia tabaci (Gennadius) MEAM1 (Middle East-Asia Minor 1, also known as biotype B) (Hemiptera: Aleyrodidae), and caused higher whitefly mortality and infection levels than commercial strains
C. javanica Apopka 97 (formerly
C. fumosorosea (Wize) Kepler, B. Shrestha and Spatafora Apopka 97),
Metarhizium brunneum Petch strain F52, and
Beauveria bassiana (Bals.-Criv.) Vuill. strain GHA [
7]. In addition, this strain was more tolerant to extreme temperatures than
C. javanica Apopka 97 [
9]. These studies suggest great potential for using
C. javanica Wf GA17 as biopesticides in pest management, especially against whiteflies in field applications.
The whitefly,
B. tabaci, infests a variety of crops (such as vegetables, ornamentals, and cotton) and may cause significant crop losses at high population densities [
10,
11]. The adults normally lay eggs on the leaf undersides; the eggs hatch into tiny crawlers that settle at a feeding location and develop through four immature stages (the fourth instar is also referred to pupa) before emerging into adults. Both adult and immature whiteflies feed on the plant phloem, reduces plant vigor and stunts the growth; meanwhile, it secrets honeydew that causes sooty mold on plant leaves and fruits, affecting crop growth and values. In addition, secondary damage may occur when whiteflies vector plant viruses, which can be more damaging than the direct feeding [
12,
13]. Whiteflies have been primarily managed with synthetic insecticides but have developed widespread resistance to many commonly used insecticides [
14,
15]. Under these circumstances, EPF has been considered as a viable solution to combat the resistance issues in managing whiteflies. Various species of EPF have been used for biological control of
B. tabaci and mostly belong to Hyphomycetes, such as
B. bassiana,
C. javanica (formerly
Isaria fumosorosea or
Paecilomyces fumosorosea),
Akanthomyces lecanii (Zimm.) Spatafora, Kepler and B. Shrestha (formerly
Lecanicillium lecanii or
Verticillium lecanii) [
1,
16]. In particular, whiteflies are on the insecticide label of several fungal biopesticides, including NOFLY
™ and PFR-97
™ (
C. javanica) and BotaniGard
®, BoteGHA
® and Velifer
® (
B. bassiana), available in the United States [
17].
The interest of using
C. javanica in whitefly control started in the 1990s, when the strain Apopka97 was isolated and tested for its high virulence to
B. tabaci and other pests [
18,
19,
20]. Studies on the use of
C. javanica for controlling
B. tabaci has been reviewed by Faria and Wraight [
16], Lacey et al. [
21] and Zimmermann [
22]. Many research on the efficacy of
C. javanica against
B. tabaci has been conducted in laboratory and greenhouse conditions except a few field studies (e.g., [
23,
24,
25,
26]), and data on field efficacy need to be expanded. In addition, the selection of isolates is a key consideration for managing
B. tabaci with EPF [
16]. Given the higher efficacy and environmental tolerance of
C. javanica Wf GA17 compared to the commercial strain Apopka 97 (in PFR-97
™) in previous studies [
7,
9], a potential advantage of the new strain may exist but needs to be verified under field conditions. Hence, the objective of this study was to evaluate the efficacy of
C. javanica Wf GA17 blastospores for control of whiteflies and assess the post-application persistence under natural field conditions. This research complements previous laboratory studies and further explores the potential use of this new fungal strain in whitefly management, which will assist in directing future research to optimize field performance.
2. Materials and Methods
2.1. Preparation of Fungal Materials
Blastospores of
C. javanica Wf GA17 and Apopka97 were grown in 100 mL of liquid culture media containing 1 × 10
5 conidia/mL in a 250-mL baffled Erlenmeyer flask for four days using a rotary shaker incubator (INNOVA 4000, New Brunswick Scientific, Edison, NJ, USA) set at 28 °C and 350 rpm. Fungal cultures were sieved using a 120-mesh screen to removal fungal hyphae. Skim milk powder (Non-Fat Instant Dry Milk, Great Value, Walmart, Bentonville, AR, USA) was added to screened cultures at 10%
w/
w [
27]. Cultures were then spray dried using a Niro spray drier (Columbia, MD, USA) with a spinning disk atomizer, at ~85 °C inlet temperature, 47–51 °C outlet temperature, and 5.7 bar wheel atomizer pressure. Blastospore concentrations per gram of product were determined with a hemocytometer under 400× magnification. Blastospore germination percentages were determined using a 6 h liquid culture germination assay [
28]. Fungal blastospores were stored under refrigeration (~5 °C) until use.
2.2. Field Efficacy in 2019
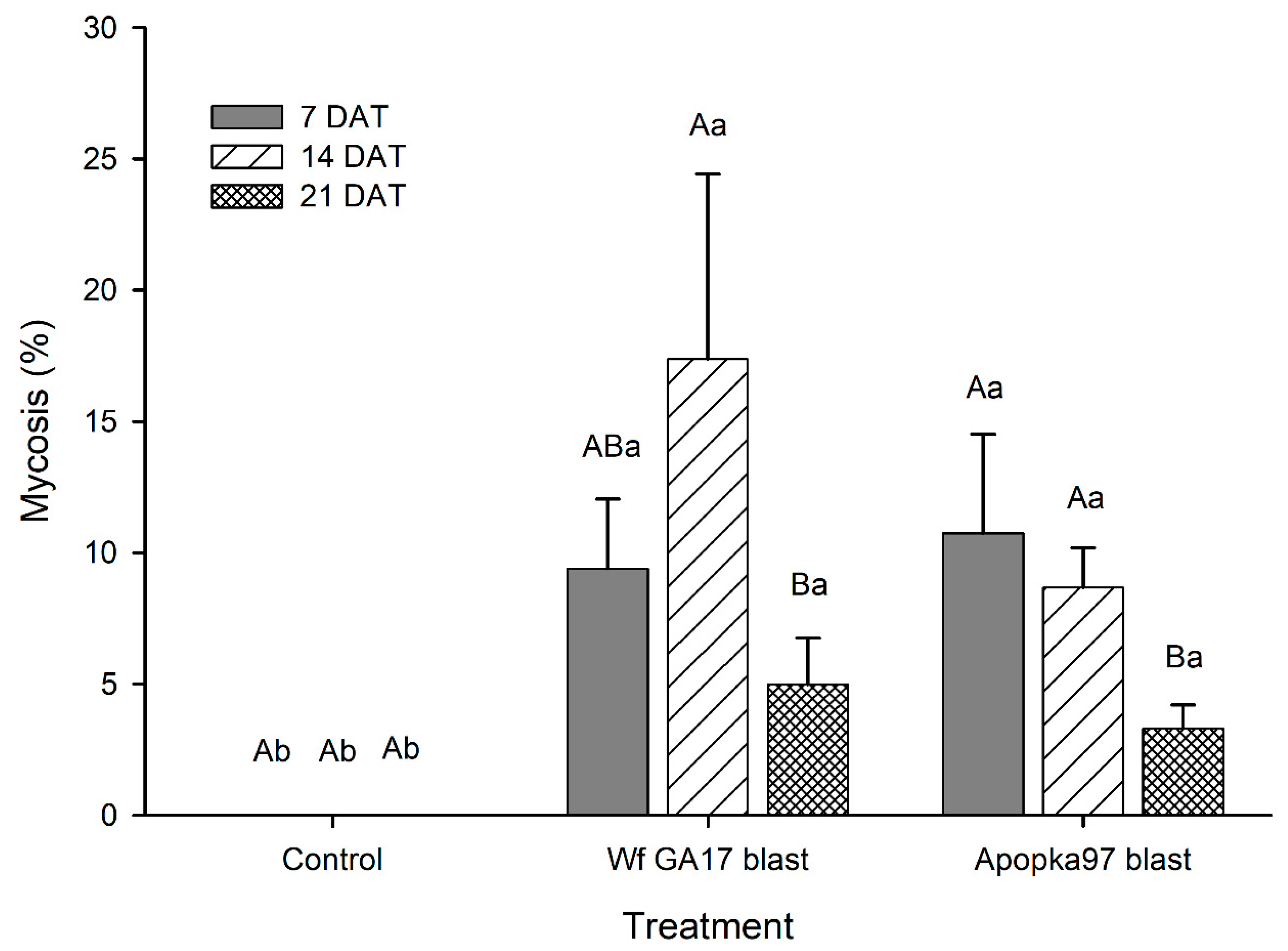
The field efficacy of blastospores of
C. javanica Wf GA17 versus Apopka97 against whiteflies was tested in a cotton field (31.5095, −83.6472) at the University of Georgia, Tifton, GA, USA. The treatments included
C. javanica Wf GA17 blastospores,
C. javanica Apopka97 blastospores, and an untreated control. Each treatment was replicated in four blocks using a randomized complete block design, and the test was conducted twice. Trial 1 was carried out on 12 August 2019. Fungal blastospores were mixed with water and sprayed at 2.5 × 10
12 viable blastospores/ha in the spray volume of 168 L/ha using a CO
2 backpack sprayer equipped with four hollow cone nozzles (ConeJet TXVS-6) at the pressure of 276 kPa. The application was made between 11:30 a.m. and 12:30 p.m. when the weather condition was sunny. The plot size was 2 m width by 2 m length, and there was a 10 m barrier between adjacent plots. After application, a field cage was placed onto each plot to prevent whitefly adults migrating to adjacent plots. The 5th true leaf (counting from top) was taken to count the number of live and dead whitefly immatures and adults at 0, 7, 14 and 21 days after treatment (DAT), and five leaves were randomly sampled from each plot. Percent fungal infection was also evaluated by examining the morphological characteristics, followed by molecular identification using the same procedure for identifying
C. javanica Wf GA17 [
7]. Trial 2 was conducted on 6 September 2019 in a different region of the same cotton field using the same treatments and procedures except that the application rate was doubled to 5 × 10
12 viable blastospores/ha. During the first and second trials, the average daily air temperature ranged 24–29 and 23–29 °C (maximum observed temperatures were 26–37 and 29–37 °C; minimum observed temperatures were 19–24 and 16–23 °C); the daily R.H. ranged 67–93% and 58–79% on average (maximum 92–99% and 83–99%; minimum 34–81% and 20–51%); the daily total solar radiation was 6–24 and 15–23 MJ/m
2; there was 133.6 and 0.1 mm of accumulated rainfall, respectively.
2.3. Field Efficacy and Post-Application Persistence in 2020
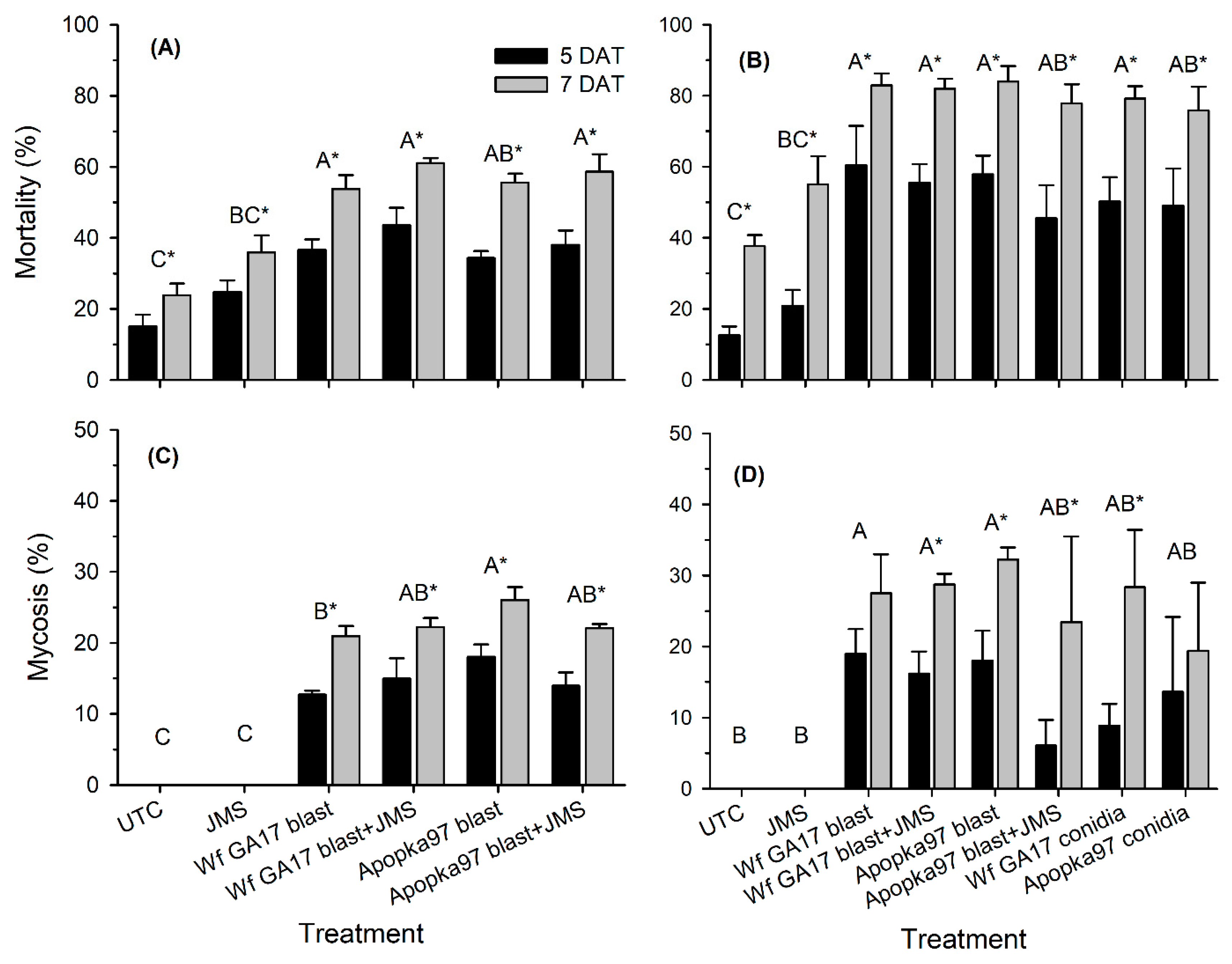
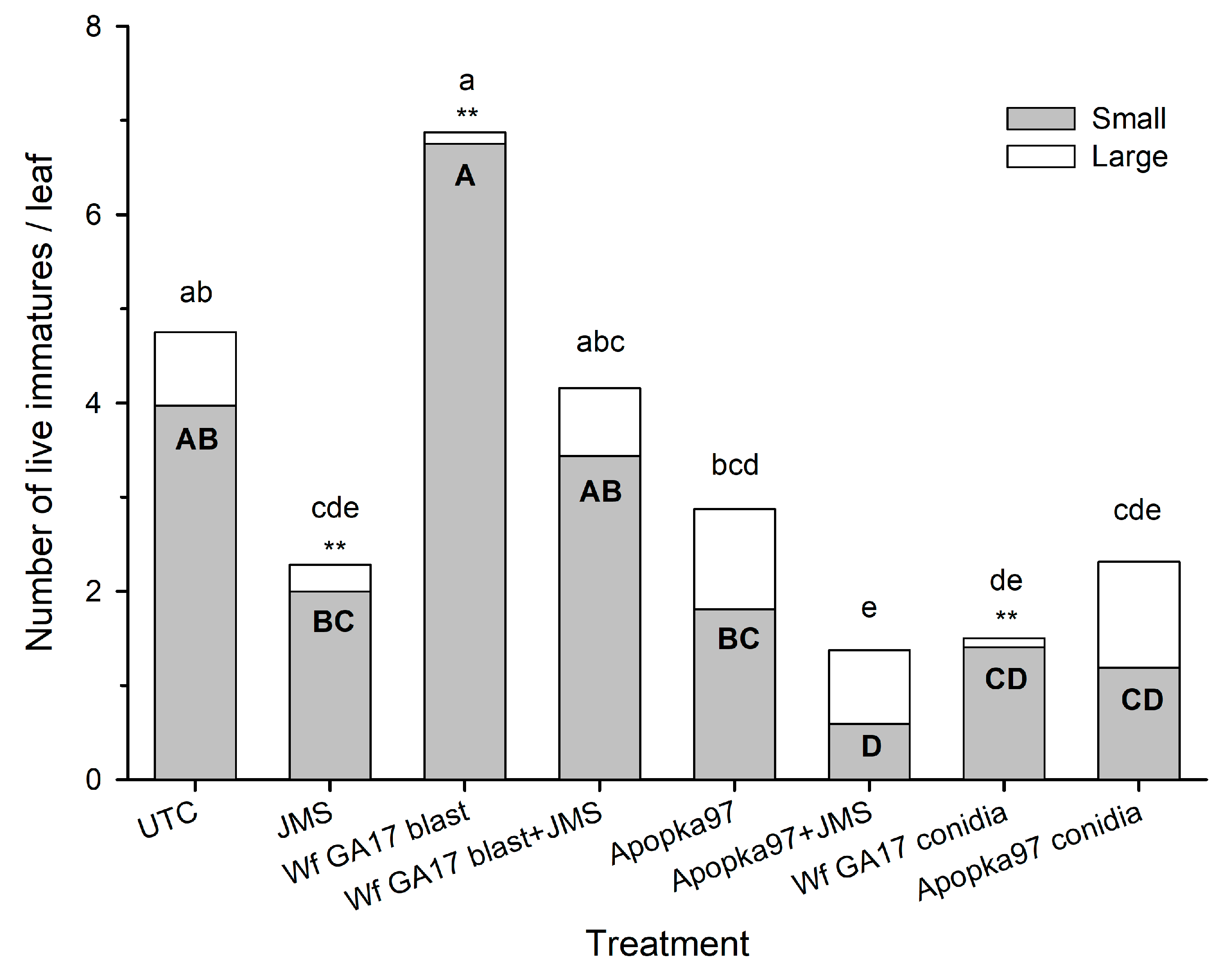
In 2020, a horticultural oil, JMS® stylet oil was added to blastospores to improve the control efficacy against whiteflies. Trial 1 was conducted in an irrigated cotton field (31.5261, −83.5464) at the University of Georgia (Tifton, GA, USA) at the stage of four nodes above white flower on 21 August 2020. There were six treatments: control, JMS oil alone, C. javanica Wf GA17 alone, C. javanica Wf GA17 plus JMS oil, C. javanica Apopka97 alone, and C. javanica Apopka97 plus JMS oil. Fungi C. javanica Wf GA17 and Apopka97 were applied at 5 × 1012 viable blastospores/ha (2.94 ×107 viable spores/mL); the JMS oil was used at 3%. Applications were made at 7 p.m. Treatments were sprayed in 168 L/ha using ConeJet TXVS-6 nozzles at 276 kPa. There were four treatment blocks, and each plot was consisted of 5.5 m width by 6.1 m length, and there was a 10 m buffer between adjacent plots. Five leaves at the 5th true leaf position were randomly taken from each plot at 0, 7, 14, 21 and 28 DAT. Live whitefly counts, mortality and percent mycosis were evaluated. For samples collected immediately after application (0 DAT), leaves were made into leaf discs placed onto 60-mm Petri dishes filled with 2% water agar to incubate fungal growth on whitefly immatures at 25 °C (light 14 h: dark 10 h), and whitely immature mortality and mycosis levels were assessed after 5 and 7 days of incubation. During the field test, the average daily temperature was between 22 and 29 °C, with the maximum between 24–35 °C and the minimum between 20 and 25 °C; there were 14 out of 28 days with rain, accumulating 178 mm of total rain; the R.H. ranged from a minimum of 29–91% to a maximum of 84–100% (average 66–97%); and the total daily solar radiation ranged 1–24 MJ/m2.
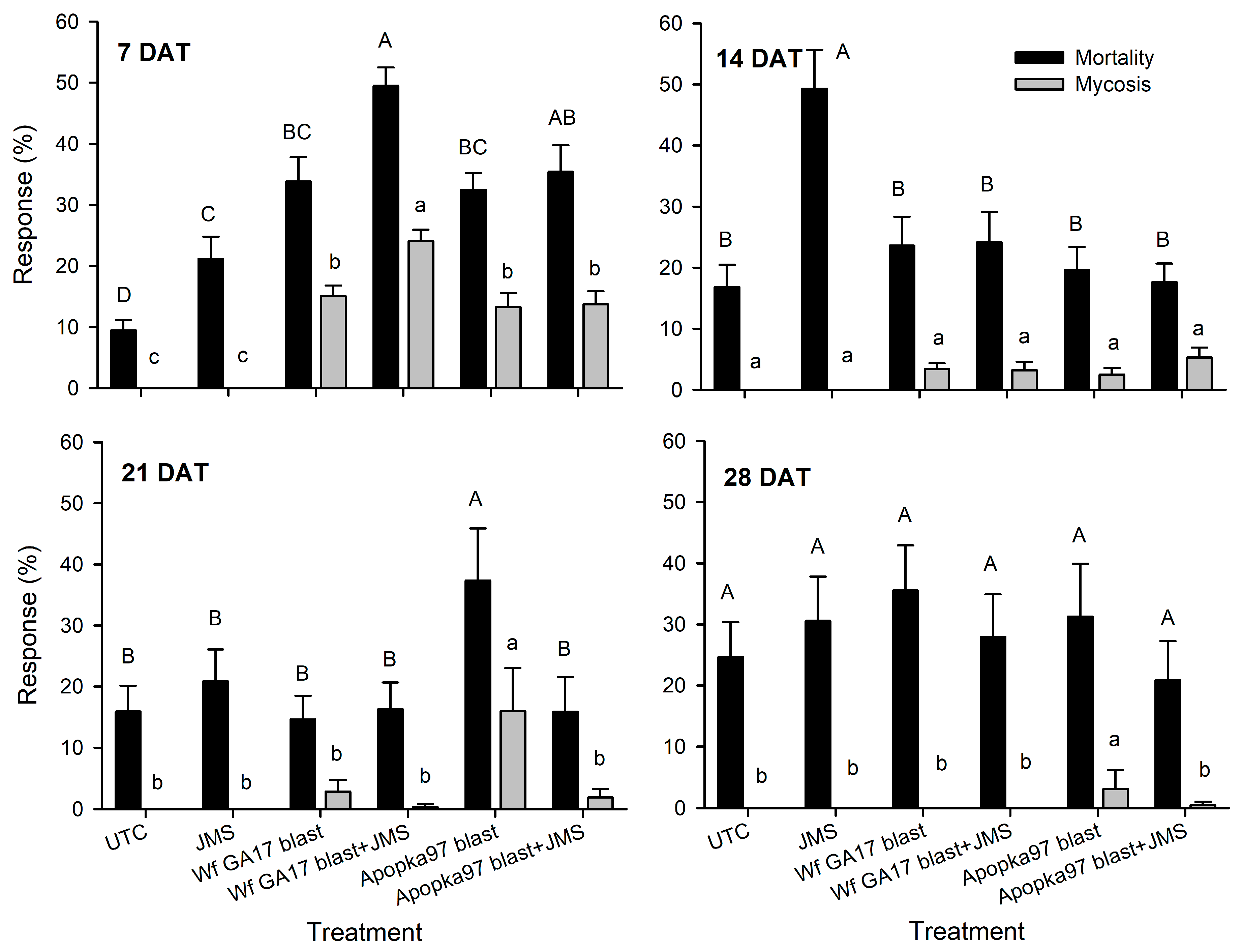
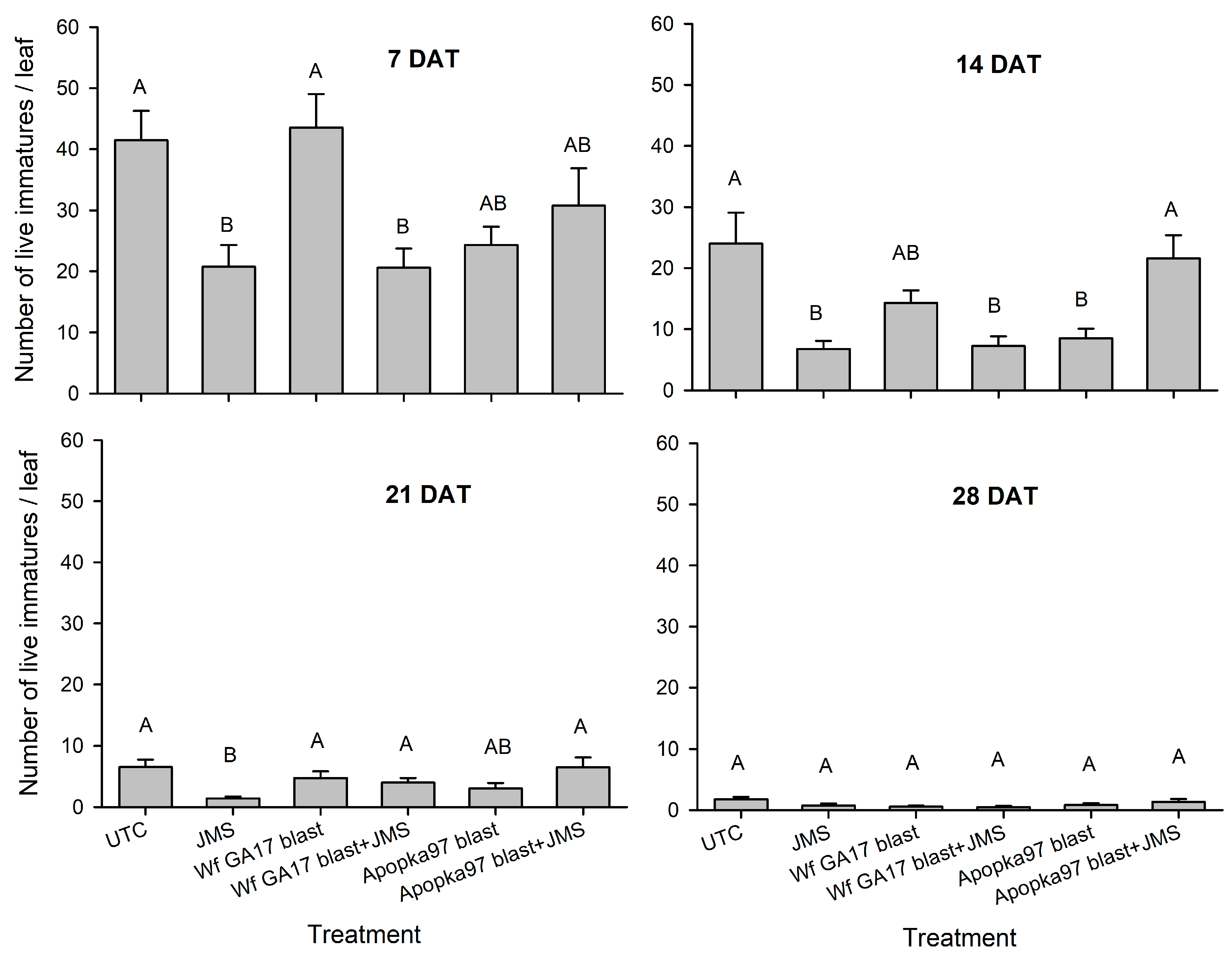
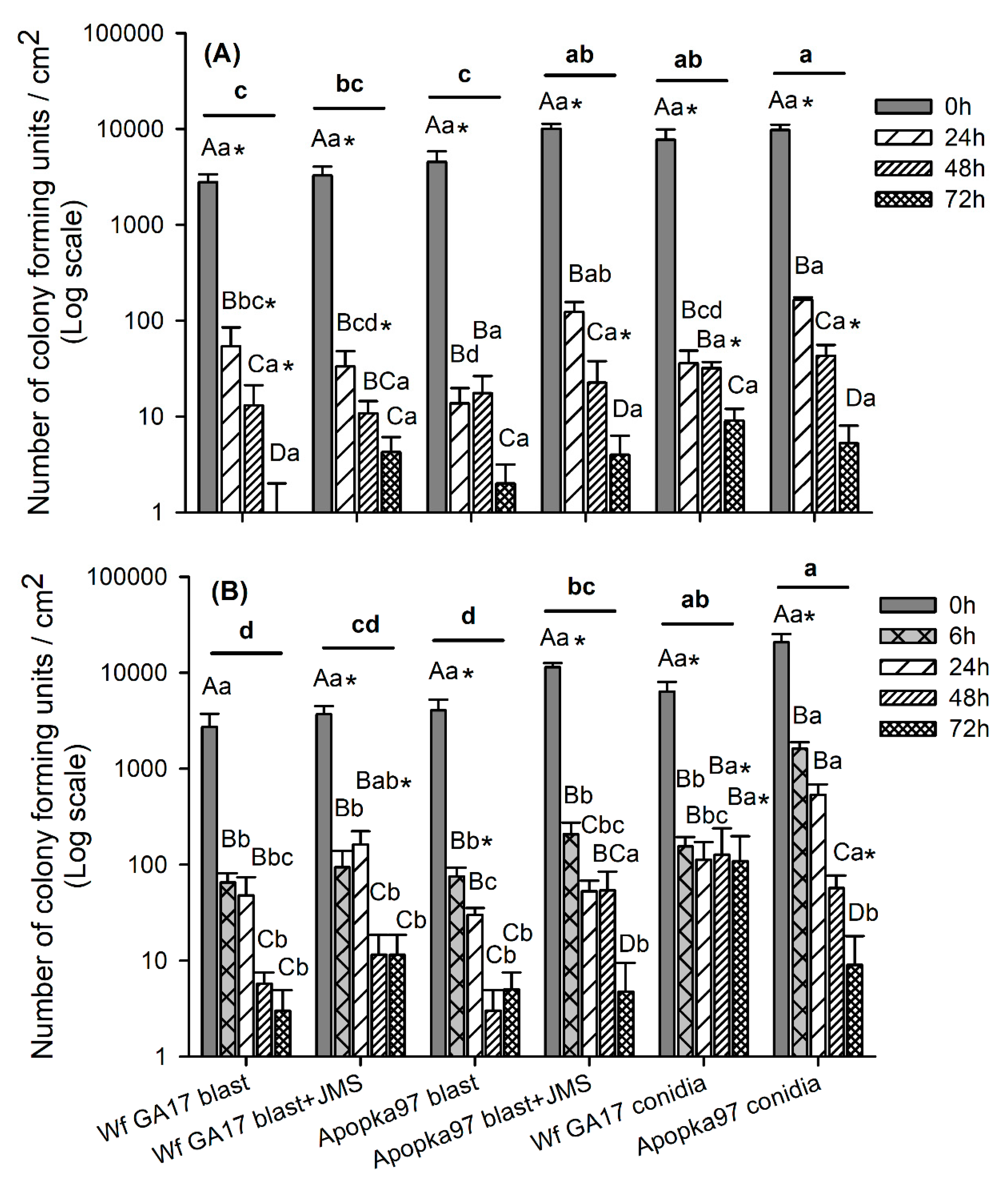
Trial 2 was carried out in a cotton field (31.5261, −83.5464) at Fort Valley State University (Fort Valley, GA, USA) at three nodes above white flower stage on 6 October 2020. In addition to the six treatments used in the previous trial, conidia of C. javanica Wf GA17 and Apopka97 (without JMS oil) were added. All fungal treatments were applied at 5 × 1012 viable spores/ha, and JMS oil applied at 3% at 10 a.m. The plot size and design were similar to the first trial, and the experimental procedures were similar except that eight leaves were sampled from each plot instead of five. Similar to the previous test, efficacy against whiteflies was evaluated at 0, 7, 14, 21 and 28 DAT. Additional samplings were taken at 0, 24, 48 and 72 h after application to evaluate fungal persistence post-application. For this purpose, a leaf disc (dia. 1.7 cm; area 2.27 cm2) was punctured from each leaf, and a total of eight leaf discs from eight leaves were used to determine fungal viability in each plot at each time. Each leaf disc was rinsed with approximately 1 mL of sterile 0.05% Silwet on each side, and the rinses of eight leaf discs from upper and lower leaf surfaces were collected separately. The collected rinses were diluted up to 10 times, and 0.1 mL of original or diluted suspension was spread onto a PDAY dodine plate (100-mm), which was incubated at 25 °C (light 14 h: dark 10 h) for 4 days to count the number of colony forming units (CFU). The number of CFUs per cm2 was evaluated for each plot over time. During the test (4 weeks), the average daily temperature ranged 9–24 °C (minimum 0–21 °C; maximum 17–31 °C); daily R.H. was 39–93% (minimum 19–75%; maximum 87–100%); there were 12 rainy days that produced 36 mm of total rain; the daily solar radiation was 7–17 MJ/m2.
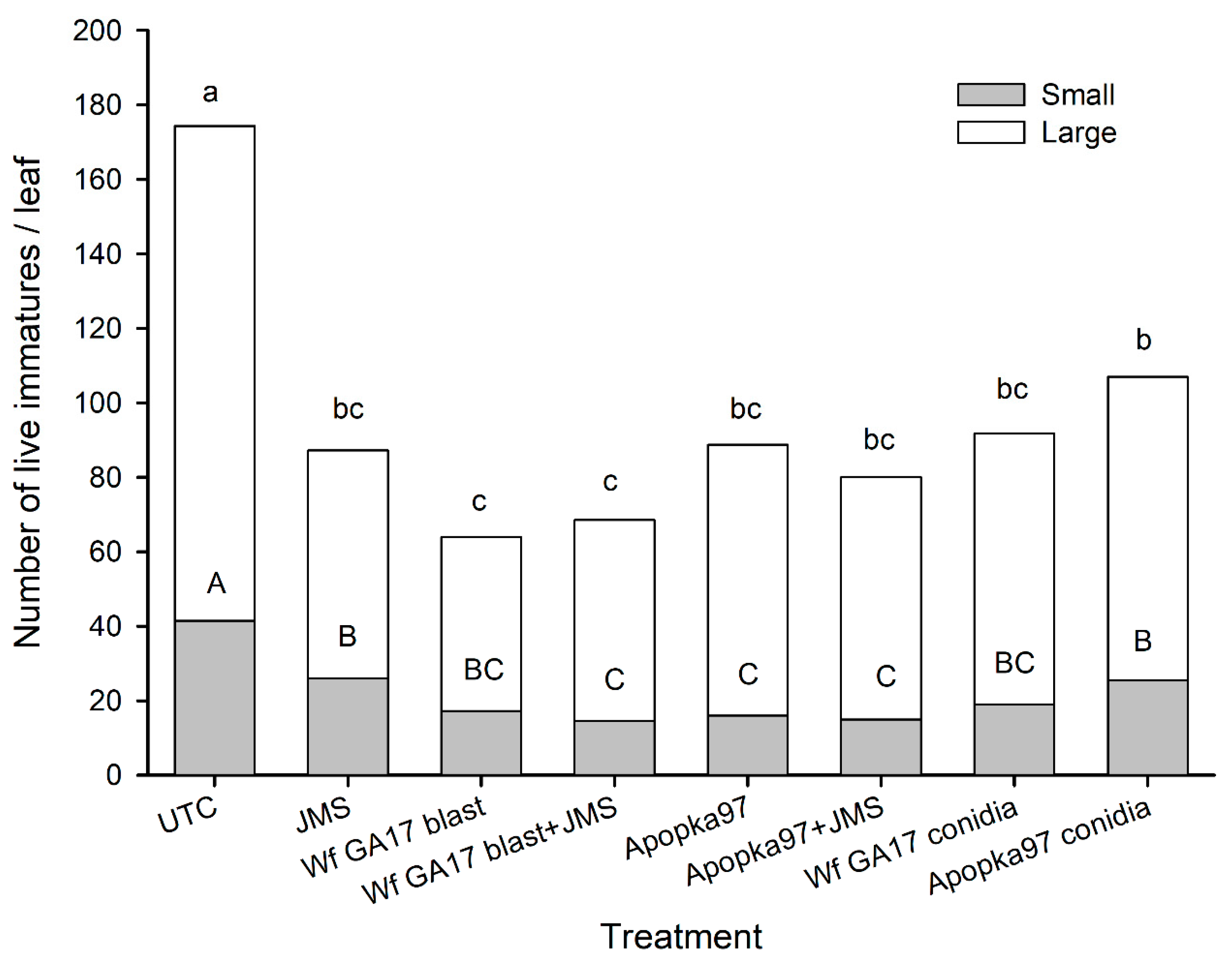
Trial 3 was implemented in a snap bean field (31.5261, −83.5464) infested with whiteflies on a research farm at University of Georgia (Tifton, GA, USA) on 30 October 2020. The eight treatments and experimental procedures were the same as the second trial, except that the fungal rate was doubled to 1 × 1013 viable spores/ha (5.88 × 107 viable spores/mL), and the plot size was 3 m width by 6 m length, with 3 m by 4 m buffer between adjacent plots. Treatments were sprayed at 9 a.m. Five leaves were sampled at 0, 7, 14 and 21 DAT (28 DAT sampling was not made due to frost) to assess live whitefly immature counts, whitefly mortality and mycosis levels for control efficacy. Additional samplings were made at 0, 6, 24, 48 and 72 h after application to check the post-application persistence of fungal treatments. During the 3-week period, the average air temperature was 10–25 °C (3–23 °C low and 16–30 °C high); the R.H. ranged 12–91% low and 71–100% high, with a daily average of 40–95%; six rainy days produced only 10 mm of rainfall in total; the daily solar radiation was 7–17 MJ/m2.
2.4. Data Analysis
All data were analyzed using the software SAS version 9.4 [
29]. For leaf samples collected immediately after treatment and incubated for various times, whitefly mortality and mycosis levels were analyzed with repeated measures with Proc Mixed procedure with block as a random factor; residual plots were checked for normality. For other samplings, immature mortality and mycosis levels were analyzed with Proc GLM using beta distribution and logit function; count data (live immatures, CFU) were analyzed with negative binomial distribution and log function. Means between treatments were separated by Tukey’s test at α = 0.05.
4. Discussion
Previously, laboratory tests showed superior virulence of
C. javanica Wf GA17 against whiteflies [
7]. Here, results under field conditions suggest that the efficacy of Wf GA17 was not different from the commercial Apopka97 strain. In 2019 field trials, neither fungus provided significant control and the infection levels were very low (below 20% maximum), when fungal blastospores applied alone. In 2020 trials, when leaf samples were collected immediately following application (minimum outdoor exposure to environmental conditions), both fungal strains applied alone or mixed with JMS oil achieved considerate immature mortality. Nonetheless, in follow-up field samplings, whiteflies were mostly killed within 7 days. Among treatments, the combination of Wf GA17 blastospores and JMS oil appeared to be most effective, which was consistently shown in 2020 trial 1 (cotton) and trial 3 (snap bean). However, the pattern was different in 2020 trial 2 (cotton), in which Apopka97 blastospores combined with JMS was most suppressive to whiteflies, while Wf GA17 blastospores + JMS seemed to be ineffective. It was observed that JMS oil alone caused significant control and reduced whitefly populations, as shown in all three field trials in 2020. When fungal blastospores applied alone, the efficacy of Wf GA17 and Apopka97 fluctuated with trials; they reduced whitefly numbers in some trials (e.g., 2020 trial 3) but were ineffective in others. The difference between blastospores applied alone and in combination with JMS only appeared for Wf GA17 in 2020 trial 1 and Apopka97 in 2020 trial 2.
The authors hypothesize that the inconsistent and low field efficacy of fungi were due to short fungal persistence. This hypothesis was verified in later persistence tests, in which most spores lost viability within 24 h (2020 trial 2) and even as short as 6 h (2020 trial 3) post-application regardless of propagule form (conidia or blastospores) and application method (alone or combined with JMS). Undoubtedly, the rapid decline in viability of fungal propagules dramatically reduced the activity of fungi in effective infection of whiteflies. Environmental factors, such as temperature, solar radiation, humidity and rainfall play a significant role in EPF persistence and control success [
2,
3,
4,
5]. High temperature is detrimental to fungal spores and may reduce the activity and infection, as shown in the previous report that the virulence of
C. javanica Wf GA17 was greatly reduced at above 30 °C and at ≥35 °C most conidia lost viability in short time [
9]. The weather was hot and dry in 2019 during test periods, especially in trial 2 where there was only 0.1 mm rain in total; in both trials there were high temperatures (close to 35 °C or above), strong solar radiation and no rain in several days following application (although trial 1 had rain afterwards), which adversely affected the survival and efficacy of both fungal strains. In 2020, environmental conditions during the first cotton test were relatively mild and the maximum temperature did not exceed 35 °C; in particular, there were frequent rainfalls, weak solar radiation and mild temperature (below 30 °C) in several days post-treatment application, which might have created an environment suitable for fungal activities, although rain might have also washed spores off leaf substrates, reducing propagules that could have taken effect. In 2020 trials 2 and 3 (conducted in October–November, 2020), the temperature dropped and solar radiation was weaker; still, the conditions were dry, which combined with other unknown factors might have compromised the fungal efficacy.
Interestingly, in the snap bean test (2020 trial 3) treatment effects did not show until at 21 DAT. Additionally, later phase infections were observed at 21 and 28 DAT in 2020 trial 1, despite at low levels. This was probably because it took longer for fungal propagules to be effective under low temperatures, or spores became dormant for a period of time and resumed activities when conditions were favorable, or the fungus propagated on hosts to reach a higher level to cause infection. Pick et al. [
30] found that the number of CFUs of
C. javanica Apopka97 (
I. fumosorosea of PFR-97™) declined at 7 days post-treatment compared with 0-day in weekly samples of field-caged citrus leaves but increased after 35 days, and there was a residue effect in causing mortality (up to 40%) of the Asian citrus psyllid,
Diaphorina citri Kuwayama, over a 35-day experimental period. Conversely, Avery et al. [
31] reported that in the field assessment of PFR-97™ against the invasive ficus whitefly,
Singhiella simplex (Singh), the CFU number increased from day 1 to day 7 post-treatment and then declined. It was suspected that the increased fungal activity was associated with germination of blastospores on leaf surfaces under favorable environmental conditions in Florida, and possibly also biotic factors such as arthropod hosts, while declined CFUs in other observations might be due to rainfall, biodegradation or other factors.
Blastospores differ greatly from conidia in structure and properties. Blastospores are thin-walled and hydrophilic whereas conidia have thicker walls and are hydrophobic. Blastospores are vegetative propagules growing in the haemocoel of infected hosts for many EPF including
C. javanica, while conidium is the most common type of propagule occurring naturally; the former germinate faster and are more virulent than conidia, but because of structural differences they are also more vulnerable to environmental stresses and less stable [
32,
33,
34,
35]. For example, blastospores of
I. fumosorosea SFP-198 were less thermotolerant than conidia and the addition of corn oil enhanced the thermotolerance [
36]. In this study, for both
C. javanica Wf GA17 and Apopka97 strains, conidia consistently persisted better than blastospores applied alone and reduced whitefly populations, as shown in both field persistence tests (2020 trials 2 and 3). However, there was a similarly high reduction in CFUs to be received by target hosts, and the two strains were not different in field persistence regardless of propagule form (conidia or blastospores) and application method (alone or combined with JMS) in both trials. Additionally, it was consistently shown that JMS oil improved persistence of Apopka97 blastospores but not Wf GA17. This effect was rather limited as most blastospores still lost activity rapidly even with the addition of oil. In terms of efficacy, Wf GA17 conidia were more effective than blastospores in reducing whiteflies in 2020 trial 2 but did not work similarly in 2020 trial 3; such a difference between propagules did not occur in Apopka97.
In addition to short persistence caused by environmental stress, lack of adequate coverage might be another reason explaining for the limited control efficacy. The mode of action of EPF requires spore adhesion to insect cuticles via direct contact to initiate the infection process, followed by the formation of appressoria penetrating the insect cuticle, mycelial growth that kills the host, and the production of asexual conidia that disperse to start the next infection cycle [
2]. Most spores (80–90%) were recovered from the upper surface of cotton leaves, while whitefly immatures develop on the lower leaf surface. Hence, most spores applied to leaf substrates lacked adequate contact with the target whiteflies and were ineffective in killing the pest. Despite strong evidence that most spores landed on upper leaf surface, after only a few hours there were more viable spores on the lower leaf surface, strongly suggesting that spores lost viability soon after application and those delivered to the lower leaf surface were protected from direct sunlight exposure and persisted longer, consistent with the report of Jaronski [
3]. In snap bean, there was no difference between leaf surfaces in spore persistence, which is expected because snap bean leaves are not parallel with the ground as with cotton leaves. To improve control efficacy, it is critical to leverage spore coverage and dispersion. Spores in the tests were delivered using a 4-nozzle bloom sprayer. Other types of sprayers, such as electrostatic and air blast sprayers with better coverage may be considered for future tests.
Given the short persistence of
C. javanica in the field, multiple applications and/or increased rate may be desirable to boost the control levels, especially for new growths of pests that were not exposed to fungi in previous sprays. Wraight et al. [
23] reported that in small-scale field trials, multiple applications of unformulated conidia of
P. fumosoroseus and
B. bassiana made at intervals of 4–5 days at 5 × 10
13 conidia/ha using an electrostatic air-assist sprayer achieved over 90% control of large nymphs of
B. argentifolii (
B. tabaci) on cucumbers and cantaloupe melons, and those applied at 1.25 × 10
13 conidia/ha at 4- to 5-day intervals reduced numbers of large nymphs by 85% in cantaloupe, although the fungi were ineffective against whitefly adults. Similarly, Jaronski and Lord [
37] found that multiple applications of
B. bassiana GHA at 2.5 × 10
13 conidia/ha in a weekly interval provided over 70% control of
B. argentifolii nymphs in an irrigated cantaloupe field in southern California. Additionally, Ou et al. [
24] reported 55% mortality of
B. tabaci caused by
C. javanica at 1 × 10
8 conidia/mL in a semi-field (cage) study at 15 days post-application in cotton, but the application rate was approximately 5 times of the label rate of similar products (PFR-97). This research mostly used the rate of 5 × 10
12 viable spores/ha (except 2019 trial 1 used 2.5 × 10
12 viable spores/ha and 2020 trial 3 used 1 × 10
13 viable spores/ha) in a single application. The lower rate and lack of repeated applications might have at least partially accounted for the inconsistent and low control levels. However, given that the used rates were close to the label rate of PFR-97, undoubtedly, increasing rate and application times would increase the material costs, while both efficacy and costs are essential considerations in strategic pest management that requires optimum control with affordable costs.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






