
Cyanobacterial Variability in Lichen Cephalodia




Abstract
:1. Introduction
2. Materials and Methods
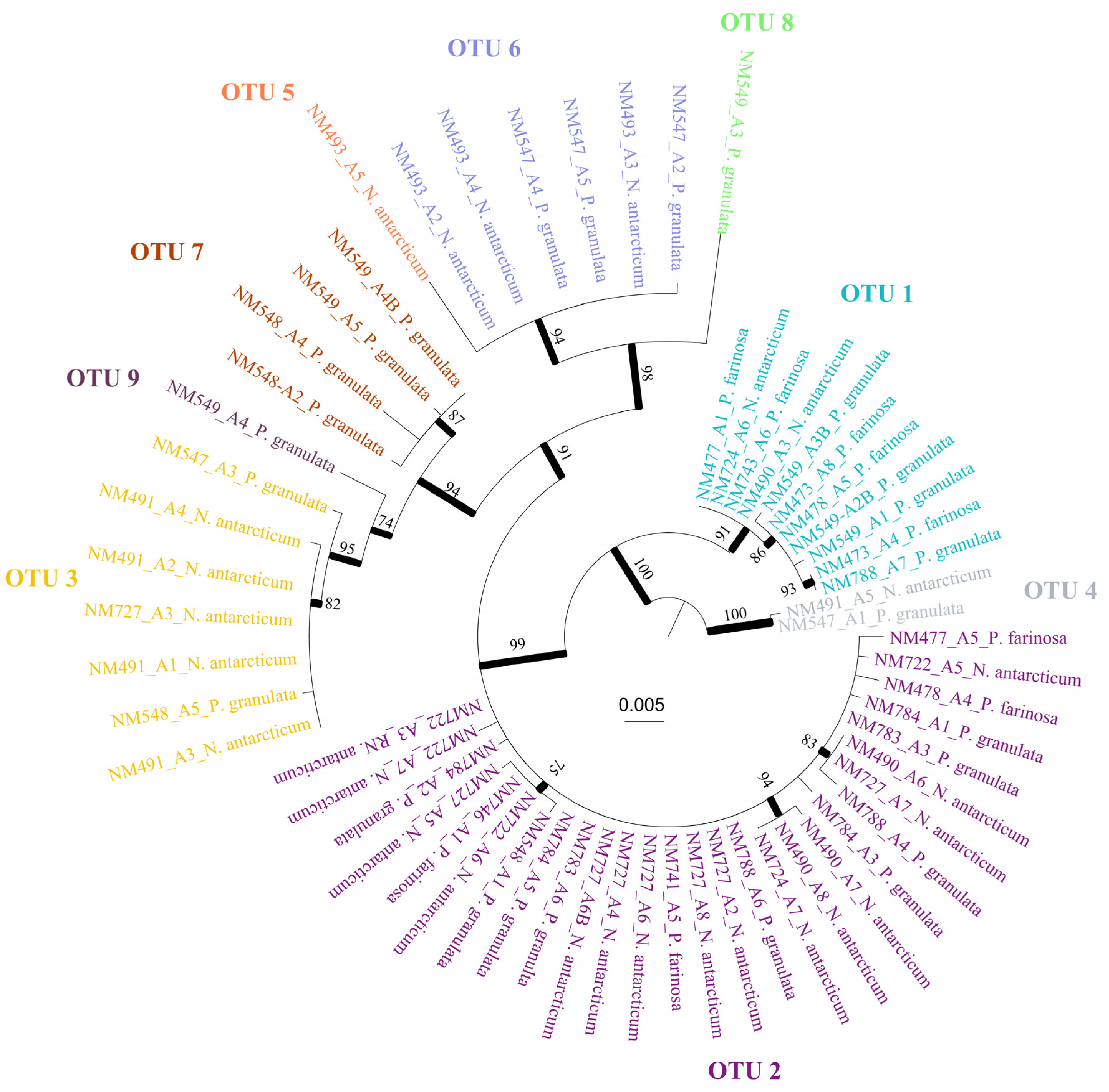
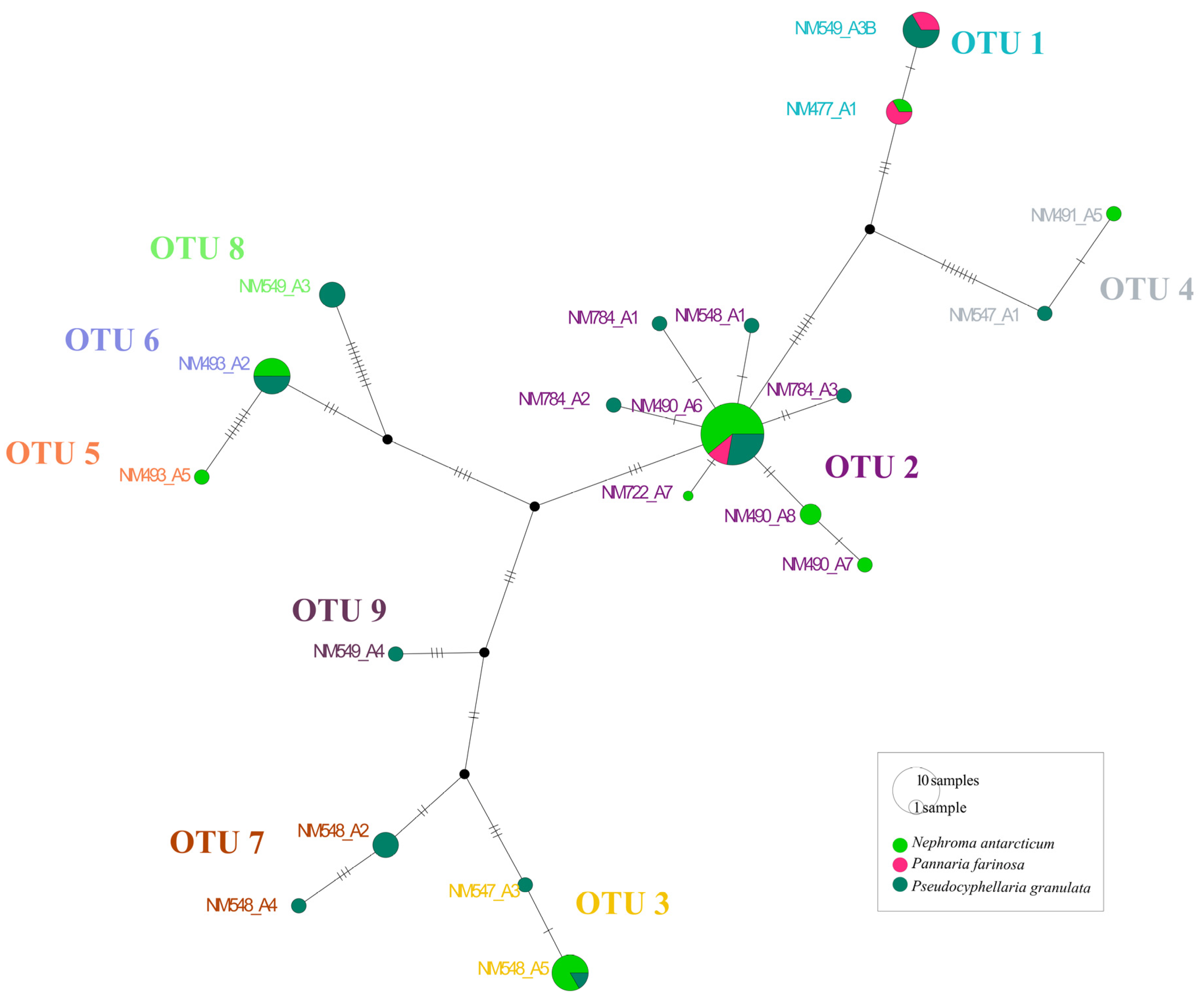
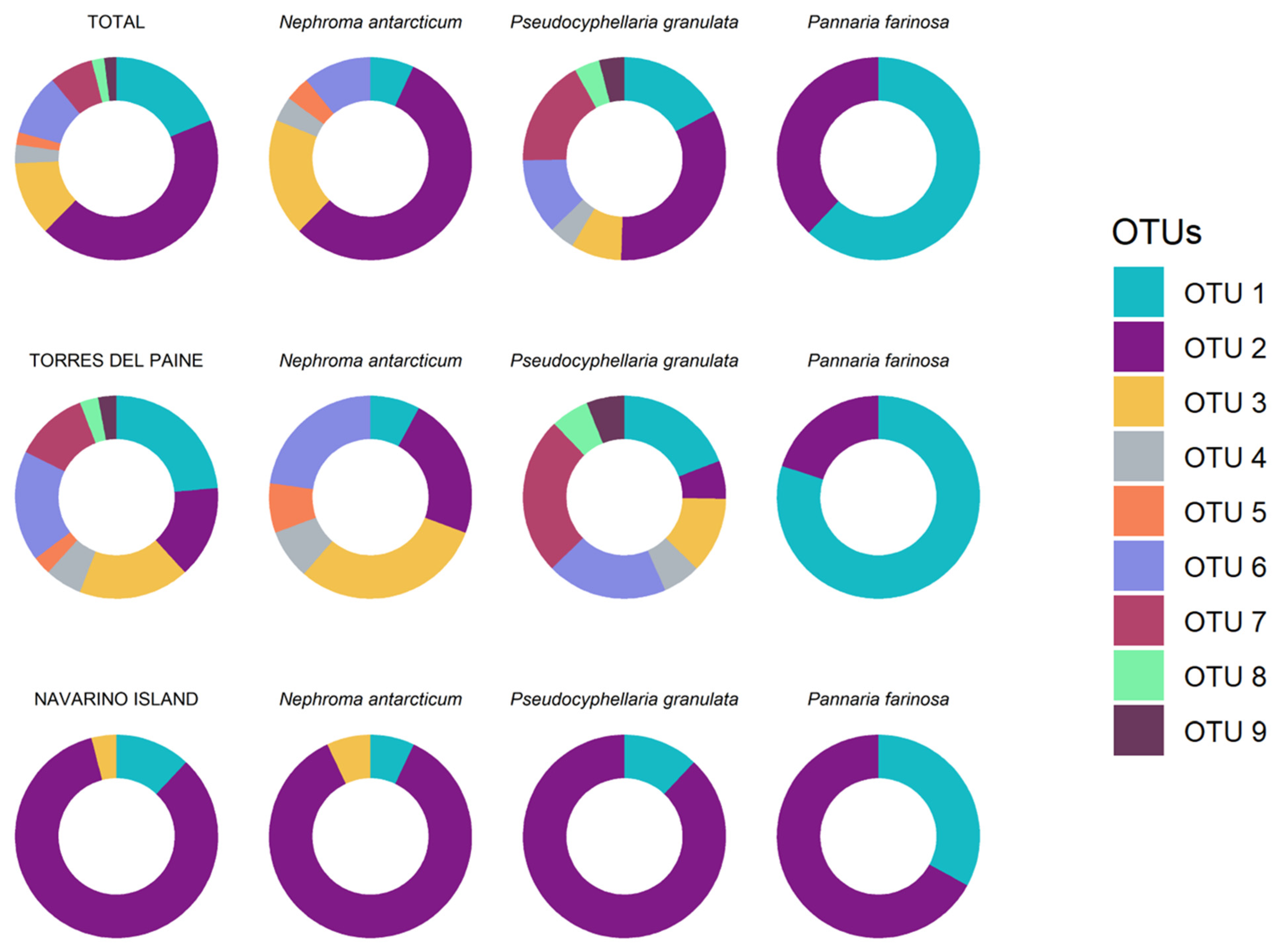
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the Fungi, 10th ed.; Cabi: Wallingford, UK, 2008; p. 640. [Google Scholar]
- Ahmadjian, V. Lichens are more important than you think. BioScience 1995, 45, 123–124. [Google Scholar] [CrossRef]
- Kershaw, K.A. Physiological Ecology of Lichens; Cambridge University Press: Cambridge, UK, 1985; p. 293. [Google Scholar]
- Rai, A.N. Nitrogen metabolism. In CRC Handbook of Lichenology, 1st ed.; Galun, M., Ed.; CRC Press Inc.: Boca Raton, FL, USA, 1998; Volume 1, pp. 201–237. [Google Scholar]
- Stanton, D.E.; Ormond, A.; Koch, N.M.; Colesie, C. Lichen ecophysiology in a changing climate. Am. J. Bot. 2023, 110, e16131. [Google Scholar] [CrossRef] [PubMed]
- Peksa, O.; Škaloud, P. Do photobionts influence the ecology of lichens? A case study of environmental preferences in symbiotic green alga Asterochloris (Trebouxiophyceae). Mol. Ecol. 2011, 20, 3936–3948. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Mendoza, F.; Domaschke, S.; García, M.A.; Jordan, P.; Martín, M.P.; Printzen, C. Population structure of mycobionts and photobionts of the widespread lichen Cetraria aculeata. Mol. Ecol. 2011, 20, 1208–1232. [Google Scholar] [CrossRef] [PubMed]
- Rolshausen, G.; Grande, F.D.; Sadowska-Deś, A.D.; Otte, J.; Schmitt, I. Quantifying the climatic niche of symbiont partners in a lichen symbiosis indicates mutualist mediated niche expansions. Ecography 2018, 41, 1380–1392. [Google Scholar] [CrossRef] [Green Version]
- Rolshausen, G.; Hallman, U.; Grande, F.D.; Otte, J.; Knudsen, K.; Schmitt, I. Expanding the mutualistic niche: Parallel symbiont turnover along climatic gradients. Proc. R. Soc. B 2020, 287, 20192311. [Google Scholar] [CrossRef] [Green Version]
- Muggia, L.; Pérez-Ortega, S.; Kopun, T.; Zellnig, G.; Grube, M. Photobiont selectivity leads to ecological tolerance and evolutionary divergence in a polymorphic complex of lichenized fungi. Ann. Bot. 2014, 114, 463–475. [Google Scholar] [CrossRef] [Green Version]
- Leavitt, S.D.; Kraichak, E.; Nelsen, M.P.; Altermann, S.; Divakar, P.K.; Alors, D.; Esslinger, T.L.; Crespo, A.; Lumbsch, T. Fungal specificity and selectivity for algae play a major role in determining lichen partnerships across diverse ecogeographic regions in the lichen-forming family Parmeliaceae (Ascomycota). Mol. Ecol. 2015, 24, 3779–3797. [Google Scholar] [CrossRef]
- Magain, N.; Miadlikowska, J.; Goffinet, B.; Sérusiaux, E.; Lutzoni, F. Macroevolution of specificity in cyanolichens of the genus Peltigera Section Polydactylon (Lecanoromycetes, Ascomycota). Syst. Biol. 2017, 66, 74–99. [Google Scholar]
- Blüthgen, N.; Menzel, F.; Blüthgen, N. Measuring specialization in species interaction networks. BMC Ecol. 2006, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Sahli, H.F.; Conner, J.K. Characterizing ecological generalization in plant-pollination systems. Oecologia 2006, 148, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Arribas, C.; Prieto, M.; Aragón, G.; López-Angulo, J.; Escudero, A.; Martínez, I. Specialization: A new multidimensional and integrative perspective. Ecology 2023, 13, e10296. [Google Scholar]
- Fox, L.R.; Morrow, P.A. Specialization: Species property or local phenomenon? Science 1981, 211, 887–893. [Google Scholar] [CrossRef] [PubMed]
- Fedrowitz, K.; Kaasalainen, U.; Rikkinen, J. Geographic mosaic of symbiont selectivity in a genus of epiphytic cyanolichens. Ecol. Evol. 2012, 2, 2291–2303. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Magain, N.; Miadlikowska, J.; Coyle, J.R.; Truong, C.; Lutzoni, F. Bioclimatic factors at an intrabiome scale are more limiting than cyanobiont availability for the lichen-forming genus Peltigera. Am. J. Bot. 2018, 105, 1198–1211. [Google Scholar] [CrossRef] [Green Version]
- Herrera, P.; Suárez, J.P.; Sánchez-Rodríguez, A.; Molina, M.C.; Prieto, M.; Méndez, M. Many broadly-shared mycobionts characterize mycorrhizal interactions of two coexisting epiphytic orchids in a high elevation tropical forest. Fungal Ecol. 2019, 39, 26–36. [Google Scholar] [CrossRef]
- Rodríguez-Arribas, C.; Martínez, I.; Aragón, G.; Zamorano-Elgueta, C.; Cavieres, L.; Prieto, M. Specialization patterns in symbiotic associations: A community perspective over spatial scales. Ecol. Evol. 2023, 13, e10296. [Google Scholar] [CrossRef]
- Paulsrud, P.; Rikkinen, J.; Lindblad, P. Cyanobiont specificity in some Nostoc-containing lichens and in a Peltigera aphthosa photosymbiodeme. New Phytol. 1998, 139, 517–524. [Google Scholar] [CrossRef]
- Piercey-Normore, M.D.; DePriest, P.T. Algal switching among lichen symbioses. Am. J. Bot. 2001, 88, 1490–1498. [Google Scholar] [CrossRef]
- Myllys, L.; Stenroos, S.; Thell, A.; Kuusinen, M. High cyanobiont selectivity of epiphytic lichens in old growth boreal forest of Finland. New Phytol. 2007, 173, 621–629. [Google Scholar] [CrossRef]
- O’Brien, H.E.; Miadlikowska, J.; Lutzoni, F. Assessing population structure and host specialization in lichenized cyanobacteria. New Phytol. 2013, 198, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Wirtz, N.; Lumbsch, T.; Green, T.G.A.; Turk, R.; Pintado, A.; Sancho, L.; Schroeter, B. Lichen fungi have low cyanobiont selectivity in maritime Antarctica. New Phytol. 2003, 160, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Muggia, L.; Vancurova, L.; Škaloud, P.; Peksa, O.; Wedin, M.; Grube, M. The symbiotic playground of lichen thalli—A highly flexible photobiont association in rock-inhabiting lichens. FEMS Microbiol. Ecol. 2013, 85, 313–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadowska-Deś, A.D.; Dal Grande, F.; Lumbsch, H.T.; Beck, A.; Otte, J.; Hur, J.S.; Kim, J.A.; Schmitt, I. Integrating coalescent and phylogenetic approaches to delimit species in the lichen photobiont Trebouxia. Mol. Phylogenet. Evol. 2014, 76, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Paulsrud, P.; Rikkinen, J.; Lindblad, P. Field investigations on cyanobacterial specificity in Peltigera aphthosa. New Phytol. 2001, 152, 117–123. [Google Scholar] [CrossRef]
- Pardo De la Hoz, C.J.; Magain, N.; Lutzoni, F.; Goward, T.; Restrepo, S.; Miadlikowska, J. Contrasting symbiotic patterns in two closely related lineages of trimembered lichens of the genus Peltigera. Front. Microbiol. 2019, 9, 2770. [Google Scholar] [CrossRef] [Green Version]
- Elvebakk, A.; Papaefthimiou, D.; Robertsen, E.; Liaimer, A. Phylogenetic patterns among Nostoc cyanobionts within bi- and tripartite lichens of the genus Pannaria. J. Phycol. 2008, 44, 1049–1059. [Google Scholar] [CrossRef]
- Paulsrud, P.; Rikkinen, J.; Lindblad, P. Spatial patterns of photobiont diversity in some Nostoc-containing lichens. New Phytol. 2000, 146, 291–299. [Google Scholar] [CrossRef]
- Bačkor, M.; Peksa, O.; Škaloud, P.; Bačkorová, M. Photobiont diversity in lichens from metal-rich substrata based on ITS rDNA sequences. Ecotoxicol. Environ. Saf. 2010, 73, 603–612. [Google Scholar] [CrossRef]
- Moya, P.; Molins, A.; Martinez-Alberola, F.; Muggia, L.; Barreno, E. Unexpected associated microalgal diversity in the lichen Ramalina farinacea is uncovered by pyrosequencing analyses. PLoS ONE 2017, 12, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Vančurová, L.; Muggia, L.; Peksa, O.; Řídká, T.; Škaloud, P. The complexity of symbiotic interactions influences the ecological amplitude of the host: A case study in Stereocaulon (lichenized Ascomycota). Mol. Ecol. 2018, 27, 3016–3033. [Google Scholar] [CrossRef] [PubMed]
- Onuț-Brännström, I.; Benjamin, M.; Scofield, D.G.; Heiðmarsson, S.; Andersson, M.G.I.; Lindström, E.S.; Johannesson, H. Sharing of photobionts in sympatric populations of Thamnolia and Cetraria lichens: Evidence from high-throughput sequencing. Sci. Rep. 2018, 8, 4406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peksa, O.; Gebouská, T.; Škvorová, Z.; Vančurová, L.; Škaloud, P. The guilds in green algal lichens-an insight into the life of terrestrial symbiotic communities. FEMS Microbiol. Ecol. 2022, 98, fiac008. [Google Scholar] [CrossRef] [PubMed]
- Rikkinen, J.; Oksanen, I.; Lohtander, K. Lichen guilds share related cyanobacterial symbionts. Science 2002, 297, 357. [Google Scholar] [CrossRef] [Green Version]
- Rikkinen, J. Ecological and evolutionary role of photobiont-mediated guilds in lichens. Symbiosis 2003, 34, 99–110. [Google Scholar]
- Yahr, R.; Vilgalys, R.; DePriest, P.T. Geographic variation in algal partners of Cladonia subtenuis (Cladoniaceae) highlights the dynamic nature of a lichen symbiosis. New Phytol. 2006, 171, 847–860. [Google Scholar] [CrossRef]
- Otálora, M.A.G.; Martínez, I.; O’Brien, H.; Molina, M.C.; Aragón, G.; Lutzoni, F. Multiple origins of high reciprocal specificity at an intercontinental spatial scale among gelatinous lichens (Collemataceae, Lecanoromycetes). Mol. Phylogenet. Evol. 2010, 56, 1089–1095. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Dal Grande, F.; Divakar, P.K.; Otte, J.; Crespo, A.; Schmitt, I. Fungal–algal association patterns in lichen symbiosis linked to macroclimate. New Phytol. 2017, 214, 317–329. [Google Scholar] [CrossRef]
- Mark, K.; Laanisto, L.; Bueno, C.G.; Niinemets, Ü.; Keller, C.; Scheidegger, C. Contrasting co-occurrence patterns of photobiont and cystobasidiomycete yeast associated with common epiphytic lichen species. New Phytol. 2020, 227, 1362–1375. [Google Scholar] [CrossRef] [PubMed]
- Rudi, K.; Skulberg, O.M.; Jakobsen, K.S. Evolution of cyanobacteria by exchange of genetic material among phyletically related strains. J. Bacteriol. 1998, 180, 3453–3461. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [Green Version]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clement, M.; Snell, Q.; Walker, P.; Posada, D.; Crandall, K. TCS: Estimating gene genealogies. In Proceedings of the Parallel and Distributed Processing Symposium, International Proceedings, Ft. Lauderdale, FL, USA, 15–19 April 2002; Volume 2, p. 184. [Google Scholar]
- Piercey-Normore, M.D.; Coxson, D.; Goward, T.; Goffinet, B. Phylogenetic position of a Pacific North American endemic cyanolichen, Nephroma occultum (Ascomycota, Peltigerales). Lichenologist 2006, 38, 441–456. [Google Scholar] [CrossRef]
- del Campo, E.M.; Gimeno, J.; de Nova, J.P.G.; Casano, L.M.; Gasulla, F.; García-Breijo, F.; Reig, F.J.; Barreno, E. South European populations of Ramalina farinacea (L.) Ach. Share different Trebouxia algae. Bibl. Lichenol. 2010, 105, 247–256. [Google Scholar]
- Blázquez, M.; Hernández-Moreno, L.S.; Gasulla, F.; Pérez-Vargas, I.; Pérez-Ortega, S. The role of photobionts as drivers of diversification in an island radiation of lichen-forming Fungi. Front. Microbiol. 2022, 12, 784182. [Google Scholar] [CrossRef] [PubMed]
- Jordan, W.P.; Rickson, F.R. Cyanophyte cephalodia in the lichen genus Nephroma. Am. J. Bot. 1971, 58, 562–568. [Google Scholar] [CrossRef]
- Tschermak-Woess, E. The algal partner. In CRC Handbook of Lichenology, 1st ed.; Galun, M., Ed.; CRC Press Inc.: Boca Raton, FL, USA, 1998; Volume 1, pp. 39–92. [Google Scholar]
- Casano, L.M.; del Campo, E.M.; García-Breijo, F.J.; Reig-Armiñana, J.; Gasulla, F.; Del Hoyo, A.; Guéra, A.; Barreno, E. Two Trebouxia algae with different physiological performances are ever-present in lichen thalli of Ramalina farinacea. Coexistence versus competition? Environ. Microbiol. 2011, 13, 806–818. [Google Scholar] [CrossRef] [PubMed]
- Lehr, H.; Galun, M.; Ott, S.; Jahns, H.M.; Fleminger, G. Cephalodia of the lichen Peltigera aphthosa (L.) Willd. Specific recognition of the compatible photobiont. Symbiosis 2000, 29, 357–365. [Google Scholar]
- Rikkinen, J. Molecular studies on cyanobacterial diversity in lichen symbioses. MycoKeys 2013, 6, 3–32. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, H.E.; Miadlikowska, J.; Lutzoni, F. Assessing host specialization in symbiotic cyanobacteria associated with four closely related species of the lichen fungus Peltigera. Eur. J. Phycol. 2005, 40, 363–378. [Google Scholar] [CrossRef]
- Cardós, J.L.H.; Prieto, M.; Jylhä, M.; Aragón, G.; Molina, M.C.; Martínez, I.; Rikkinen, J. A case study on the re-establishment of the cyanolichen symbiosis: Where do the compatible photobionts come from? An. Bot. 2019, 124, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.N.; Soderback, E.; Bergman, B. Cyanobacterium–plant symbioses. New Phytol. 2000, 147, 449–481. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Species | Thallus | Cephalodia | OTU ASAP | Forest | Voucher | GenBank |
|---|---|---|---|---|---|---|
| Nephroma antarcticum | NM490 | A3 | 1 | TP | ARAN20283 | OR344442 |
| Nephroma antarcticum | NM490 | A6 | 2 | TP | OR344444 | |
| Nephroma antarcticum | NM490 | A7 | 2 | TP | OR344445 | |
| Nephroma antarcticum | NM490 | A8 | 2 | TP | OR344446 | |
| Nephroma antarcticum | NM491 | A3 | 3 | TP | ARAN20284 | OR344447 |
| Nephroma antarcticum | NM491 | A1 | 3 | TP | OR344449 | |
| Nephroma antarcticum | NM491 | A2 | 3 | TP | OR344450 | |
| Nephroma antarcticum | NM491 | A4 | 3 | TP | OR344451 | |
| Nephroma antarcticum | NM491 | A5 | 4 | TP | OR344448 | |
| Nephroma antarcticum | NM493 | A5 | 5 | TP | ARAN20285 | OR344452 |
| Nephroma antarcticum | NM493 | A2 | 6 | TP | OR344454 | |
| Nephroma antarcticum | NM493 | A3 | 6 | TP | OR344455 | |
| Nephroma antarcticum | NM493 | A4 | 6 | TP | OR344456 | |
| Nephroma antarcticum | NM722 | A3 | 2 | IN | ARAN20292 | OR344474 |
| Nephroma antarcticum | NM722 | A6 | 2 | IN | OR344476 | |
| Nephroma antarcticum | NM722 | A5 | 2 | IN | OR344475 | |
| Nephroma antarcticum | NM722 | A7 | 2 | IN | OR344477 | |
| Nephroma antarcticum | NM724 | A6 | 1 | IN | ARAN20293 | OR344478 |
| Nephroma antarcticum | NM724 | A7 | 2 | IN | OR344479 | |
| Nephroma antarcticum | NM727 | A3 | 3 | IN | ARAN20294 | OR344481 |
| Nephroma antarcticum | NM727 | A2 | 2 | IN | OR344480 | |
| Nephroma antarcticum | NM727 | A4 | 2 | IN | OR344482 | |
| Nephroma antarcticum | NM727 | A6 | 2 | IN | OR344484 | |
| Nephroma antarcticum | NM727 | A8 | 2 | IN | OR344486 | |
| Nephroma antarcticum | NM727 | A5 | 2 | IN | OR344483 | |
| Nephroma antarcticum | NM727 | A7 | 2 | IN | OR344485 | |
| Pannaria farinosa | NM473 | A4 | 1 | TP | OR344431 | |
| Pannaria farinosa | NM473 | A8 | 1 | TP | ARAN20286 | OR344432 |
| Pannaria farinosa | NM477 | A1 | 1 | TP | ARAN20287 | OR344433 |
| Pannaria farinosa | NM477 | A2 | 2 | TP | OR344434 | |
| Pannaria farinosa | NM478 | A5 | 1 | TP | ARAN20288 | OR344441 |
| Pannaria farinosa | NM741 | A5 | 2 | IN | ARAN20295 | OR344487 |
| Pannaria farinosa | NM743 | A6 | 1 | IN | ARAN20296 | OR344488 |
| Pannaria farinosa | NM746 | A1 | 2 | IN | ARAN20297 | OR344489 |
| Pseudocyphellaria granulata | NM547 | A4 | 6 | TP | ARAN20289 | OR344460 |
| Pseudocyphellaria granulata | NM547 | A5 | 6 | TP | OR344461 | |
| Pseudocyphellaria granulata | NM547 | A2 | 6 | TP | OR344458 | |
| Pseudocyphellaria granulata | NM547 | A3 | 3 | TP | OR344459 | |
| Pseudocyphellaria granulata | NM547 | A1 | 4 | TP | OR344457 | |
| Pseudocyphellaria granulata | NM548 | A5 | 3 | TP | ARAN20290 | OR344465 |
| Pseudocyphellaria granulata | NM548 | A2 | 7 | TP | OR344462 | |
| Pseudocyphellaria granulata | NM548 | A4 | 7 | TP | OR344463 | |
| Pseudocyphellaria granulata | NM548 | A1 | 2 | TP | OR344464 | |
| Pseudocyphellaria granulata | NM549 | A4B | 7 | TP | ARAN20291 | OR344469 |
| Pseudocyphellaria granulata | NM549 | A5 | 7 | TP | OR344470 | |
| Pseudocyphellaria granulata | NM549 | A3 | 8 | TP | OR344471 | |
| Pseudocyphellaria granulata | NM549 | A4 | 9 | TP | OR344472 | |
| Pseudocyphellaria granulata | NM549 | A2B | 1 | TP | OR344467 | |
| Pseudocyphellaria granulata | NM549 | A3B | 1 | TP | OR344468 | |
| Pseudocyphellaria granulata | NM549 | A1 | 1 | TP | OR344466 | |
| Pseudocyphellaria granulata | NM783 | A3 | 2 | IN | ARAN20298 | OR344491 |
| Pseudocyphellaria granulata | NM783 | A6 | 2 | IN | OR344492 | |
| Pseudocyphellaria granulata | NM784 | A5 | 2 | IN | ARAN20299 | OR344496 |
| Pseudocyphellaria granulata | NM784 | A2 | 2 | IN | OR344494 | |
| Pseudocyphellaria granulata | NM784 | A1 | 2 | IN | OR344493 | |
| Pseudocyphellaria granulata | NM784 | A3 | 2 | IN | OR344495 | |
| Pseudocyphellaria granulata | NM788 | A7 | 1 | IN | ARAN20300 | OR344500 |
| Pseudocyphellaria granulata | NM788 | A6 | 2 | IN | OR344499 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prieto, M.; Montané, N.; Aragón, G.; Martínez, I.; Rodríguez-Arribas, C. Cyanobacterial Variability in Lichen Cephalodia. J. Fungi 2023, 9, 826. https://doi.org/10.3390/jof9080826
Prieto M, Montané N, Aragón G, Martínez I, Rodríguez-Arribas C. Cyanobacterial Variability in Lichen Cephalodia. Journal of Fungi. 2023; 9(8):826. https://doi.org/10.3390/jof9080826
Chicago/Turabian StylePrieto, Maria, Natalia Montané, Gregorio Aragón, Isabel Martínez, and Clara Rodríguez-Arribas. 2023. "Cyanobacterial Variability in Lichen Cephalodia" Journal of Fungi 9, no. 8: 826. https://doi.org/10.3390/jof9080826