
A Symbiotic Fungus Sistotrema Benefits Blueberry Rejuvenation and Abiotic Stress Tolerance
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Soil Sampling, Reinoculation, and Experimental Design
2.2. Soil DNA Extraction
2.3. ITS Gene Sequencing, Data Processing, and Taxonomic Assignments
2.4. Functional Predictions Based on ITS Regions
2.5. Data Visualization and Analysis
2.6. COG Functional Annotation
2.7. Directional Isolation of Fungal Strains and Reinoculation
2.8. Determination of Physiological Indexes
2.9. Statistical Analysis
3. Results
3.1. Diversity Analysis of Fungi in Rhizosphere and Non-Rhizosphere Soils of Different Blueberry Cultivars
3.2. Functional Prediction Based on Fungal ITS Sequences and Estimation of Blueberry Root Health Status
3.3. Rhizosphere Fungal Co-Occurrence Networks and Differences in Rhizosphere Fungi among Various Blueberry Cultivars
3.4. Enrichment Analysis of COG Functional Classes of Reinoculated Fungi
3.5. Effects of Reinoculation with Symbiotic Fungi on Chlorophyll Content and Root Activity of Blueberry under Drought and Deficiency Stress
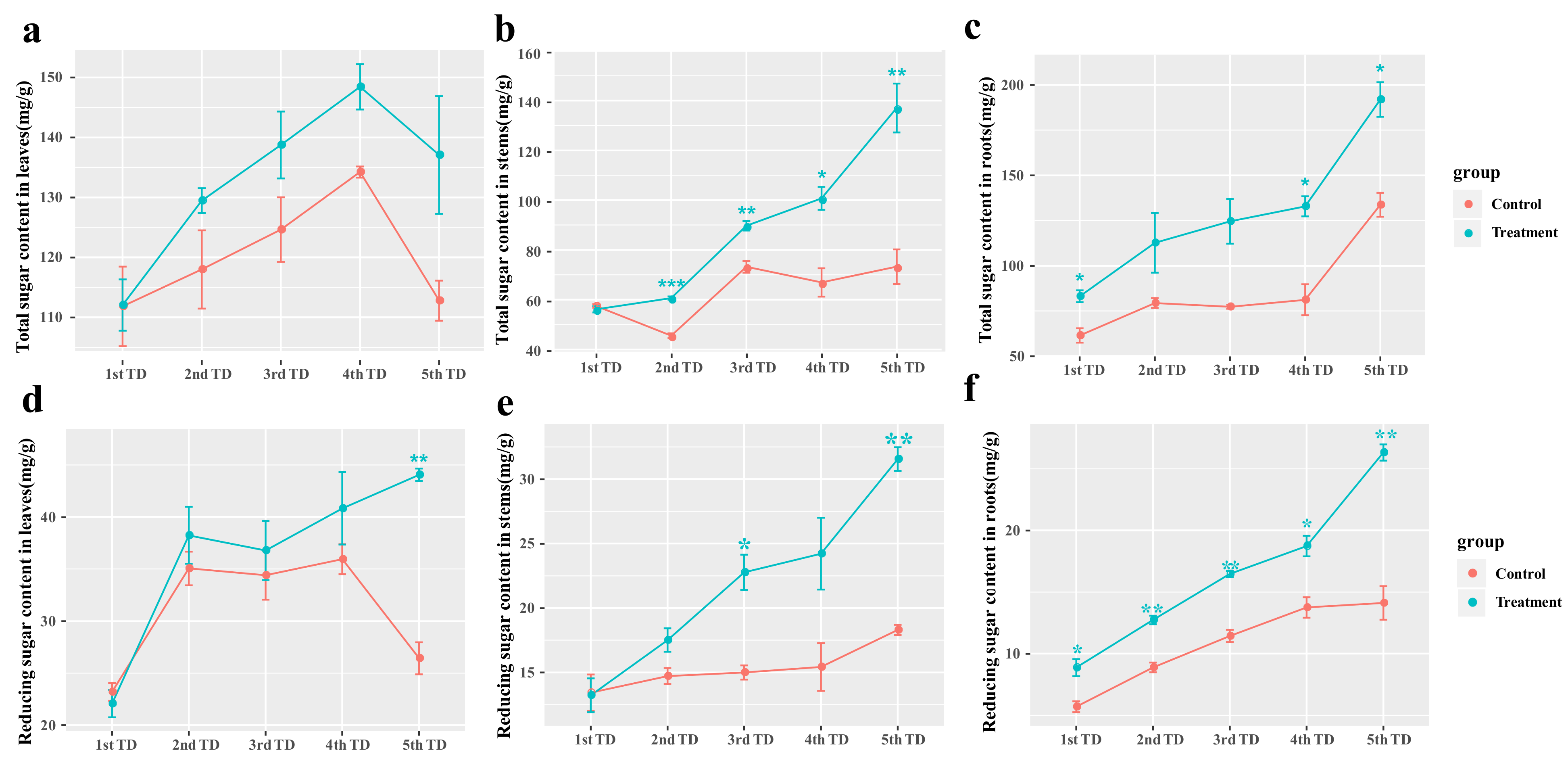
3.6. Effects of Symbiotic Fungi Reinoculation on Carbohydrate Synthesis in Blueberry under Drought and Nutrient-Deficiency Stress
3.7. Effects of Symbiotic Fungi Reinoculation on Soluble Iron and Nitrogen Contents in Rhizosphere Soil
3.8. Effects of Symbiotic Fungi Reinoculation on Plant Height, Rejuvenation, and Sprouting of Blueberry
4. Discussion
4.1. Differences of Fungal Diversity in Rhizosphere Soil under Different Growth States and Different Cultivars of Blueberry
4.2. Identification of Core Symbiotic Fungi Unique to ‘Premier’ Blueberry
4.3. Growth-Promoting Effect of Sistotrema Reinoculated onto Blueberry
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hinsinger, P.; Bengough, A.G.; Vetterlein, D.; Young, I.M. Rhizosphere: Biophysics, biogeochemistry and ecological relevance. Plant Soil. 2009, 321, 117–152. [Google Scholar] [CrossRef]
- Massart, S.; Martinez-Medina, M.; Jijakli, M.H. Biological control in the microbiome era: Challenges and opportunities. Biol. Control 2015, 89, 98–108. [Google Scholar] [CrossRef]
- Hosseyni Moghaddam, M.S.; Safaie, N.; Soltani, J.; Hagh-Doust, N. Desert-adapted fungal endophytes induce salinity and drought stress resistance in model crops. Plant Physiol. Biochem. 2021, 160, 225–238. [Google Scholar] [CrossRef]
- Mendes, R.; Kruijt, M.; De Bruijn, I.; Dekkers, E.; Van Der Voort, M.; Schneider, J.H.; Piceno, Y.M.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A. Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef] [PubMed]
- Zamioudis, C.; Pieterse, C.M. Modulation of host immunity by beneficial microbes. Mol. Plant Microbe Interact. 2012, 25, 139–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaparro, J.M.; Sheflin, A.M.; Manter, D.K.; Vivanco, J.M. Manipulating the soil microbiome to increase soil health and plant fertility. Biol. Fertil. Soils 2012, 48, 489–499. [Google Scholar] [CrossRef]
- Hayatsu, M.; Katsuyama, C.; Tago, K. Overview of recent researches on nitrifying microorganisms in soil. Soil. Sci. Plant Nutr. 2021, 67, 619–632. [Google Scholar] [CrossRef]
- Nguyen, T.T.N.; Xu, C.-Y.; Tahmasbian, I.; Che, R.; Xu, Z.; Zhou, X.; Wallace, H.M.; Bai, S.H. Effects of biochar on soil available inorganic nitrogen: A review and meta-analysis. Geoderma 2017, 288, 79–96. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Wang, X.M.; Ma, H.Y.; Yang, T.; Jia, Y.; Zhou, J.; Dai, C.C. Fungal endophyte Phomopsis liquidambari affects nitrogen transformation processes and related microorganisms in the rice rhizosphere. Front. Microbiol. 2015, 6, 982. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Luo, C.; Zhang, D.; Cai, X.; Jiang, L.; Zhao, X.; Zhang, G. Diversity of the active phenanthrene degraders in PAH-polluted soil is shaped by ryegrass rhizosphere and root exudates. Soil. Biol. Biochem. 2019, 128, 100–110. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Badri, D.V.; Bakker, M.G.; Sugiyama, A.; Manter, D.K.; Vivanco, J.M. Root exudation of phytochemicals in Arabidopsis follows specific patterns that are developmentally programmed and correlate with soil microbial functions. PLoS ONE 2013, 8, e55731. [Google Scholar] [CrossRef]
- Kellenberger, E. Exploring the unknown. The silent revolution of microbiology. EMBO Rep. 2001, 2, 5–7. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhu, Y.; Shao, T.; Long, X.; Gao, X.; Zhou, Z. Relationship between rhizosphere soil properties and disease severity in highbush blueberry (Vaccinium corymbosum). Agric. Ecosyst. Environ. Appl. Soil. Ecol. 2019, 137, 187–194. [Google Scholar] [CrossRef]
- Cai, B.; Vancov, T.; Si, H.; Yang, W.; Tong, K.; Chen, W.; Fang, Y. Isolation and Characterization of Endomycorrhizal Fungi Associated with Growth Promotion of Blueberry Plants. J. Fungi 2021, 7, 584. [Google Scholar] [CrossRef]
- Liu, X.M.; Xu, Q.L.; Li, Q.Q.; Zhang, H.; Xiao, J.X. Physiological responses of the two blueberry cultivars to inoculation with an arbuscular mycorrhizal fungus under low-temperature stress. J. Plant Nutr. 2017, 40, 2562–2570. [Google Scholar] [CrossRef]
- Bizabani, C.; Dames, J. Effects of inoculating Lachnum and Cadophora isolates on the growth of Vaccinium corymbosum. Microbiol. Res. 2015, 181, 68–74. [Google Scholar] [CrossRef]
- Bizabani, C.; Fontenla, S.; Dames, J.F. Ericoid fungal inoculation of blueberry under commercial production in South Africa. Sci. Hortic. 2016, 209, 173–177. [Google Scholar] [CrossRef]
- Watts-Williams, S.J.; Cavagnaro, T.R.; Tyerman, S.D. Variable effects of arbuscular mycorrhizal fungal inoculation on physiological and molecular measures of root and stomatal conductance of diverse Medicago truncatula accessions. Plant Cell Environ. 2019, 42, 285–294. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Mavrodi, O.V.; Hou, J.; Blackmon, C.; Babiker, E.M.; Mavrodi, D.V. Comparative Analysis of Rhizosphere Microbiomes of Southern Highbush Blueberry (Vaccinium corymbosum L.), Darrow’s Blueberry (V. darrowii Camp), and Rabbiteye Blueberry (V. virgatum Aiton). Front. Microbiol. 2020, 11, 370. [Google Scholar] [CrossRef] [Green Version]
- Yeates, C.; Gillings, M.R.; Davison, A.D.; Altavilla, N.; Veal, D.A. Methods for microbial DNA extraction from soil for PCR amplification. Biol. Proced. Online 1998, 1, 40–47. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, R.H.; Larsson, K.H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glockner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2019, 47, D259–D264. [Google Scholar] [CrossRef] [PubMed]
- Alonso, R.M.; San-Martin, M.I.; Sotres, A.; Escapa, A. Graphene oxide electrodeposited electrode enhances start-up and selective enrichment of exoelectrogens in bioelectrochemical systems. Sci. Rep. 2017, 7, 13726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Hanbo, C.; Boutros, P.C. VennDiagram: A package for the generation of highly-customizable Venn and Euler diagrams in R. BMC Bioinf. 2011, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Conway, J.R.; Lex, A.; Gehlenborg, N. UpSetR: An R package for the visualization of intersecting sets and their properties. Bioinformatics 2017, 33, 2938–2940. [Google Scholar] [CrossRef] [Green Version]
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. Complex. Syst. 2006, 1695. [Google Scholar]
- Dong, L.; Xu, J.; Feng, G.; Li, X.; Chen, S. Soil bacterial and fungal community dynamics in relation to Panax notoginseng death rate in a continuous cropping system. Sci. Rep. 2016, 6, 31802. [Google Scholar] [CrossRef] [Green Version]
- Huerta-Cepas, J.; Forslund, K.; Coelho, L.P.; Szklarczyk, D.; Jensen, L.J.; von Mering, C.; Bork, P. Fast Genome-Wide Functional Annotation through Orthology Assignment by eggNOG-Mapper. Mol. Biol. Evol. 2017, 34, 2115–2122. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansouri, H.; Salari, F. Influence of mevinolin on chloroplast terpenoids in Cannabis sativa. Physiol. Mol. Biol. Plants 2014, 20, 273–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, E.; Yang, X.; Li, T.; Liu, D.; Jin, X.; Meng, F. Effect of Pb toxicity on root morphology, physiology and ultrastructure in the two ecotypes of Elsholtzia argyi. J. Hazard. Mater. 2007, 147, 806–816. [Google Scholar] [CrossRef] [PubMed]
- Leyva, A.; Quintana, A.; Sanchez, M.; Rodriguez, E.N.; Cremata, J.; Sanchez, J.C. Rapid and sensitive anthrone-sulfuric acid assay in microplate format to quantify carbohydrate in biopharmaceutical products: Method development and validation. Biologicals 2008, 36, 134–141. [Google Scholar] [CrossRef]
- Ballesteros, L.F.; Teixeira, J.A.; Mussatto, S.I. Extraction of polysaccharides by autohydrolysis of spent coffee grounds and evaluation of their antioxidant activity. Carbohydr. Polym. 2017, 157, 258–266. [Google Scholar] [CrossRef] [Green Version]
- Jalali, M.; Beygi, M.; Jalali, M.; Buss, W. Background levels of DTPA-extractable trace elements in calcareous soils and prediction of trace element availability based on common soil properties. J. Geochem. Explor. 2022, 241, 107073. [Google Scholar] [CrossRef]
- Safari Sinegani, A.A.; Safari Sinegani, M. Chemical fractionation and bioavailability of Fe, Mn, Pb, and Cd in soils around Meyghan Lake, Arak, Iran. Int. J. Environ. Sci. Technol. 2018, 16, 3297–3308. [Google Scholar] [CrossRef]
- Manolikaki, I.; Diamadopoulos, E. Agronomic potential of biochar prepared from brewery byproducts. J. Environ. Manag. 2020, 255, 109856. [Google Scholar] [CrossRef]
- Clément, C.-C.; Cambouris, A.N.; Ziadi, N.; Zebarth, B.J.; Karam, A.; Naeth, M.A. Nitrogen source and rate effects on residual soil nitrate and overwinter NO3-N losses for irrigated potatoes on sandy soils. Can. J. Soil. Sci. 2020, 100, 44–57. [Google Scholar] [CrossRef]
- Zang, Y.-X.; Chun, I.-J.; Zhang, L.-L.; Hong, S.-B.; Zheng, W.-W.; Xu, K. Effect of gibberellic acid application on plant growth attributes, return bloom, and fruit quality of rabbiteye blueberry. Sci. Hortic. 2016, 200, 13–18. [Google Scholar] [CrossRef]
- Jin, Q.; Zhang, Y.; Ma, Y.; Sun, H.; Guan, Y.; Liu, Z.; Ye, Q.; Zhang, Y.; Shao, C.; Mu, P.; et al. The composition and function of the soil microbial community and its driving factors before and after cultivation of Panax ginseng in farmland of different ages. Ecol. Indic. 2022, 145, 109748. [Google Scholar] [CrossRef]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Han, M.; Hu, Y.; Li, Z.; Liu, C.; Wang, X.; Tian, Q.; Jiao, W.; Hu, J.; Liu, L.; et al. Effects of Continuous Cropping of Sweet Potato on the Fungal Community Structure in Rhizospheric Soil. Front. Microbiol. 2019, 10, 2269. [Google Scholar] [CrossRef] [PubMed]
- Sasse, J.; Martinoia, E.; Northen, T. Feed Your Friends: Do Plant Exudates Shape the Root Microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoeksema, J.D.; Chaudhary, V.B.; Gehring, C.A.; Johnson, N.C.; Karst, J.; Koide, R.T.; Pringle, A.; Zabinski, C.; Bever, J.D.; Moore, J.C.; et al. A meta-analysis of context-dependency in plant response to inoculation with mycorrhizal fungi. Ecol. Lett. 2010, 13, 394–407. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Zhang, Z.; Zhou, L. Advances in the studies on symbiotic arbuscular mycorrhizal fungi of traditional Chinese medicinal plants. BIOCELL 2022, 46, 2559–2573. [Google Scholar] [CrossRef]
- Bradley, E.L.; Okmen, B.; Doehlemann, G.; Henrissat, B.; Bradshaw, R.E.; Mesarich, C.H. Secreted Glycoside Hydrolase Proteins as Effectors and Invasion Patterns of Plant-Associated Fungi and Oomycetes. Front. Plant Sci. 2022, 13, 853106. [Google Scholar] [CrossRef]
- Schröder, I.; Johnson, E.; de Vries, S. Microbial ferric iron reductases. FEMS Microbiol. Rev. 2003, 27, 427–447. [Google Scholar] [CrossRef] [Green Version]
- Howitt, S.M.; Udvardi, M.K. Structure, function and regulation of ammonium transporters in plants. Biochim. Biophys. Acta Biomembr. 2000, 1465, 152–170. [Google Scholar] [CrossRef] [Green Version]
- Su, Z.; Zhang, Y.; Zhao, R.; Zhou, J. Enhancement of dissimilatory nitrate/nitrite reduction to ammonium of Escherichia coli sp. SZQ1 by ascorbic acid: Mechanism and performance. Sci. Total Environ. 2022, 853, 158423. [Google Scholar] [CrossRef]
- Zhong, J.; Gu, J.; Guo, Y.; You, S.; Liao, F.; Chen, W.; Guo, W. Blueberry VcLon1 protease increases iron use efficiency by alleviating chloroplast oxidative stress. Plant Soil. 2019, 445, 533–548. [Google Scholar] [CrossRef]
- Larsen, C.N.; Krantz, B.A.; Wilkinson, K.D. Substrate Specificity of Deubiquitinating Enzymes: Ubiquitin C-Terminal Hydrolases. Biochemistry 1998, 37, 3358–3368. [Google Scholar] [CrossRef] [PubMed]
- Qihua, L.; Weihua, H.; Amy, B.; Paul, J. Chloroplast Biogenesis Is Regulated by Direct Action of the Ubiquitin-Proteasome System. Science 2012, 338, 655–659. [Google Scholar] [CrossRef]
- Hernandez-Martinez, F.; Briones-Roblero, C.I.; Nelson, D.R.; Rivera-Orduna, F.N.; Zuniga, G. Cytochrome P450 complement (CYPome) of Candida oregonensis, a gut-associated yeast of bark beetle, Dendroctonus rhizophagus. Fungal Biol. 2016, 120, 1077–1089. [Google Scholar] [CrossRef]
- Hagh-Doust, N.; Färkkilä, S.M.A.; Hosseyni Moghaddam, M.S.; Tedersoo, L. Symbiotic fungi as biotechnological tools: Methodological challenges and relative benefits in agriculture and forestry. Fungal Biol. Rev. 2022, 42, 34–55. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blueberry Variety | Growth State | Plant Character | Sampling Site | Sample Code |
|---|---|---|---|---|
| ‘O’Neal’ | Weak growth | Flourishing | Rhizosphere | O |
| Flourishing | Nonrhizosphere | OK | ||
| ‘Sharpblue’ | Medium growth | Flourishing | Rhizosphere | XG |
| Flourishing | Nonrhizosphere | XGK | ||
| Weak | Rhizosphere | XB | ||
| Weak | Nonrhizosphere | XBK | ||
| ‘Premier’ | Strong growth | Flourishing | Rhizosphere | JG |
| Flourishing | Nonrhizosphere | JGK | ||
| Weak | Rhizosphere | JB | ||
| Weak | Nonrhizosphere | JBK |
| Blueberry Variety | Classification | Plant Character | Treatment |
|---|---|---|---|
| 1-year-old ‘Gulfcoast’ | Southern highbush blueberry | Normal growth | Drought and nutrient-deficiency stress |
| 3-year-old ‘Star’ | Southern highbush blueberry | Normal growth | Normal treatment |
| Stunted | Normal treatment | ||
| Dying | Normal treatment | ||
| 3-year-old ‘Jewel’ | Southern highbush blueberry | Stunted | Normal treatment |
| Dying | Normal treatment |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, Y.; Zhan, X.; Wang, K.; Zhong, J.; Liao, F.; Chen, W.; Guo, W. A Symbiotic Fungus Sistotrema Benefits Blueberry Rejuvenation and Abiotic Stress Tolerance. J. Fungi 2023, 9, 779. https://doi.org/10.3390/jof9070779
Ye Y, Zhan X, Wang K, Zhong J, Liao F, Chen W, Guo W. A Symbiotic Fungus Sistotrema Benefits Blueberry Rejuvenation and Abiotic Stress Tolerance. Journal of Fungi. 2023; 9(7):779. https://doi.org/10.3390/jof9070779
Chicago/Turabian StyleYe, Yu, Xufang Zhan, Kai Wang, Jingya Zhong, Fanglei Liao, Wenrong Chen, and Weidong Guo. 2023. "A Symbiotic Fungus Sistotrema Benefits Blueberry Rejuvenation and Abiotic Stress Tolerance" Journal of Fungi 9, no. 7: 779. https://doi.org/10.3390/jof9070779