
The Frq–Frh Complex Light-Dependently Delays Sfl1-Induced Microsclerotia Formation in Verticillium dahliae

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioinformatic Methods
2.2. Cultivation of Microorganisms
2.3. Verification of Gene Annotation
2.4. Plasmid and Verticillium Strain Construction
2.5. Genomic DNA Extraction and Southern Hybridization
2.6. Cultivation of V. dahliae for Time Point Experiments
2.7. Western Experiments and Quantification of GFP Fusion Protein Levels
2.8. RNA Extraction, cDNA Synthesis and Transcript Level Quantification
2.9. Phenotypical Analysis
2.10. Quantification of Microsclerotia and Conidiospore Formation
2.11. Pathogenicity Assay on Tomato Plants
2.12. Confocal Microscopy
2.13. In Vitro Protein Pull-Down and LC/MS Analyses
3. Results
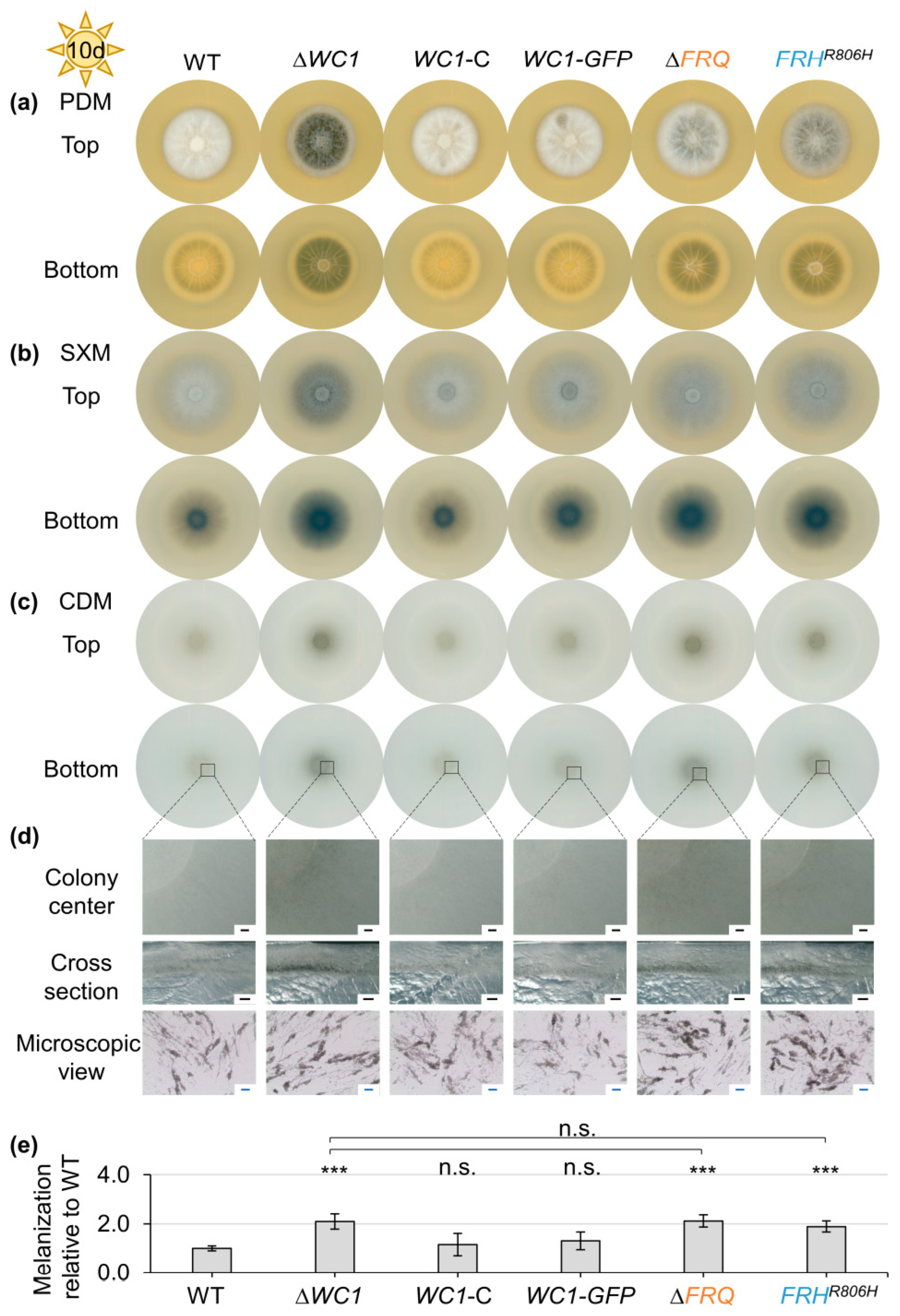
3.1. Light-Dependent Repression of Microsclerotia Formation and Induction of Aerial Hyphae Formation in V. dahliae Require FRQ, FRH, and WC1
3.2. V. dahliae FRQ and FRH Positively Affect Conidiation
3.3. Symptom Induction in V. dahliae Infected Tomato Plants Is Independent of FRQ, FRH, and WC1
3.4. The Frq–Frh Complex Formation in V. dahliae Depends on Frh Amino Acid Residue Arginine 806
3.5. Sfl1 Positively Affects Microsclerotia Formation Prior to Frq and Contributes to V. dahliae Spore Formation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Braunsdorf, C.; Mailänder-Sánchez, D.; Schaller, M. Fungal Sensing of Host Environment. Cell Microbiol. 2016, 18, 1188–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Fischer, R. Light Sensing and Responses in Fungi. Nat. Rev. Microbiol. 2019, 17, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Diernfellner, A.C.R.; Brunner, M. Phosphorylation Timers in the Neurospora crassa Circadian Clock. J. Mol. Biol. 2020, 432, 3449–3465. [Google Scholar] [CrossRef] [PubMed]
- Froehlich, A.C.; Liu, Y.; Loros, J.J.; Dunlap, J.C. White Collar-1, a Circadian Blue Light Photoreceptor, Binding to the frequency Promoter. Science 2002, 297, 815–819. [Google Scholar] [CrossRef]
- He, Q.; Cheng, P.; Yang, Y.; Wang, L.; Gardner, K.H.; Liu, Y. White Collar-1, a DNA Binding Transcription Factor and a Light Sensor. Science 2002, 297, 840–843. [Google Scholar] [CrossRef]
- Talora, C.; Franchi, L.; Linden, H.; Ballario, P.; Macino, G. Role of a White Collar-1-White Collar-2 Complex in Blue-Light Signal Transduction. EMBO J. 1999, 18, 4961–4968. [Google Scholar] [CrossRef]
- Hurley, J.M.; Dasgupta, A.; Emerson, J.M.; Zhou, X.; Ringelberg, C.S.; Knabe, N.; Lipzen, A.M.; Lindquist, E.A.; Daum, C.G.; Barry, K.W.; et al. Analysis of Clock-Regulated Genes in Neurospora Reveals Widespread Posttranscriptional Control of Metabolic Potential. Proc. Natl. Acad. Sci. USA 2014, 111, 16995–17002. [Google Scholar] [CrossRef]
- Garceau, N.Y.; Liu, Y.; Loros, J.J.; Dunlap, J.C. Alternative Initiation of Translation and Time-Specific Phosphorylation Yield Multiple Forms of the Essential Clock Protein FREQUENCY. Cell 1997, 89, 469–476. [Google Scholar] [CrossRef] [Green Version]
- Diernfellner, A.C.R.; Schafmeier, T.; Merrow, M.W.; Brunner, M. Molecular Mechanism of Temperature Sensing by the Circadian Clock of Neurospora crassa. Genes Dev. 2005, 19, 1968–1973. [Google Scholar] [CrossRef] [Green Version]
- Colot, H.V.; Loros, J.J.; Dunlap, J.C. Temperature-Modulated Alternative Splicing and Promoter Use in the Circadian Clock Gene frequency. Mol. Biol. Cell 2005, 16, 5563–5571. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Garceau, N.Y.; Loros, J.J.; Dunlap, J.C. Thermally Regulated Translational Control of FRQ Mediates Aspects of Temperature Responses in the Neurospora Circadian Clock. Cell 1997, 89, 477–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diernfellner, A.; Colot, H.V.; Dintsis, O.; Loros, J.J.; Dunlap, J.C.; Brunner, M. Long and Short Isoforms of Neurospora Clock Protein FRQ Support Temperature Compensated Circadian Rhythms. FEBS Lett. 2007, 581, 5759–5764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurley, J.M.; Larrondo, L.F.; Loros, J.J.; Dunlap, J.C. Conserved RNA Helicase FRH Acts Nonenzymatically to Support the Intrinsically Disordered Neurospora Clock Protein FRQ. Mol. Cell 2013, 52, 832–843. [Google Scholar] [CrossRef] [Green Version]
- Pelham, J.F.; Dunlap, J.C.; Hurley, J.M. Intrinsic Disorder Is an Essential Characteristic of Components in the Conserved Circadian Circuit. Cell Commun. Signal. 2020, 18, 181. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.; He, Q.; He, Q.; Wang, L.; Liu, Y. Regulation of the Neurospora Circadian Clock by an RNA Helicase. Genes Dev. 2005, 19, 234–241. [Google Scholar] [CrossRef] [Green Version]
- Cheng, P.; Yang, Y.; Heintzen, C.; Liu, Y. Coiled-Coil Domain-Mediated FRQ-FRQ Interaction Is Essential for Its Circadian Clock Function in Neurospora. EMBO J. 2001, 20, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Cha, J.; Yuan, H.; Liu, Y. Regulation of the Activity and Cellular Localization of the Circadian Clock Protein FRQ. J. Biol. Chem. 2011, 286, 11469–11478. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Cheng, P.; Liu, Y. Functional Significance of FRH in Regulating the Phosphorylation and Stability of Neurospora Circadian Clock Protein FRQ. J. Biol. Chem. 2010, 285, 11508–11515. [Google Scholar] [CrossRef] [Green Version]
- Schafmeier, T.; Haase, A.; Káldi, K.; Scholz, J.; Fuchs, M.; Brunner, M. Transcriptional Feedback of Neurospora Circadian Clock Gene by Phosphorylation-Dependent Inactivation of Its Transcription Factor. Cell 2005, 122, 235–246. [Google Scholar] [CrossRef] [Green Version]
- He, Q.; Cha, J.; He, Q.; Lee, H.-C.; Yang, Y.; Liu, Y. CKI and CKII Mediate the FREQUENCY-Dependent Phosphorylation of the WHITE COLLAR Complex to Close the Neurospora Circadian Negative Feedback Loop. Genes Dev. 2006, 20, 2552–2565. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Kettenbach, A.N.; Zhou, X.; Loros, J.J.; Dunlap, J.C. The Phospho-Code Determining Circadian Feedback Loop Closure and Output in Neurospora. Mol. Cell 2019, 74, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Cheng, P.; Yuan, H.; Liu, Y. The Exosome Regulates Circadian Gene Expression in a Posttranscriptional Negative Feedback Loop. Cell 2009, 138, 1236–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Q.; Cheng, P.; Yang, Y.; He, Q.; Yu, H.; Liu, Y. FWD1-Mediated Degradation of FREQUENCY in Neurospora Establishes a Conserved Mechanism for Circadian Clock Regulation. EMBO J. 2003, 22, 4421–4430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Q.; Liu, Y. Degradation of the Neurospora Circadian Clock Protein FREQUENCY through the Ubiquitin-Proteasome Pathway. Biochem. Soc. Trans. 2005, 33, 953–956. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.L.; Kettenbach, A.N.; Loros, J.J.; Gerber, S.A.; Dunlap, J.C. Quantitative Proteomics Reveals a Dynamic Interactome and Phase-Specific Phosphorylation in the Neurospora Circadian Clock. Mol. Cell 2009, 34, 354–363. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.-T.; Li, S.; Long, C.; Cha, J.; Huang, G.; Li, L.; Chen, S.; Liu, Y. Setting the Pace of the Neurospora Circadian Clock by Multiple Independent FRQ Phosphorylation Events. Proc. Natl. Acad. Sci. USA 2009, 106, 10722–10727. [Google Scholar] [CrossRef]
- Wang, B.; Stevenson, E.-L.; Dunlap, J.C. Functional Analysis of 110 Phosphorylation Sites on the Circadian Clock Protein FRQ Identifies Clusters Determining Period Length and Temperature Compensation. G3 Genes Genomes Genet. 2023, 13, jkac334. [Google Scholar] [CrossRef]
- Chen, X.; Liu, X.; Gan, X.; Li, S.; Ma, H.; Zhang, L.; Wang, P.; Li, Y.; Huang, T.; Yang, X.; et al. Differential Regulation of Phosphorylation, Structure and Stability of Circadian Clock Protein FRQ Isoforms. J. Biol. Chem. 2023, 299, 104597. [Google Scholar] [CrossRef]
- Shi, M.; Collett, M.; Loros, J.J.; Dunlap, J.C. FRQ-Interacting RNA Helicase Mediates Negative and Positive Feedback in the Neurospora Circadian Clock. Genetics 2010, 184, 351–361. [Google Scholar] [CrossRef] [Green Version]
- Conrad, K.S.; Hurley, J.M.; Widom, J.; Ringelberg, C.S.; Loros, J.J.; Dunlap, J.C.; Crane, B.R. Structure of the Frequency-interacting RNA Helicase: A Protein Interaction Hub for the Circadian Clock. EMBO J. 2016, 35, 1707–1719. [Google Scholar] [CrossRef] [Green Version]
- Salichos, L.; Rokas, A. The Diversity and Evolution of Circadian Clock Proteins in Fungi. Mycologia 2010, 102, 269–278. [Google Scholar] [CrossRef] [Green Version]
- Valim, H.F.; Dal Grande, F.; Otte, J.; Singh, G.; Merges, D.; Schmitt, I. Identification and Expression of Functionally Conserved Circadian Clock Genes in Lichen-Forming Fungi. Sci. Rep. 2022, 12, 15884. [Google Scholar] [CrossRef]
- Tong, S.-M.; Wang, D.-Y.; Cai, Q.; Ying, S.-H.; Feng, M.-G. Opposite Nuclear Dynamics of Two FRH-Dominated Frequency Proteins Orchestrate Non-Rhythmic Conidiation in Beauveria bassiana. Cells 2020, 9, 626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, S.-M.; Gao, B.-J.; Peng, H.; Feng, M.-G. Essential Roles of Two FRQ Proteins (Frq1 and Frq2) in Beauveria bassiana’s Virulence, Infection Cycle, and Calcofluor-Specific Signaling. Appl. Environ. Microbiol. 2021, 87, e02545-20. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.-Y.; Yu, L.; Luo, X.-C.; Ying, S.-H.; Feng, M.-G. Co-Regulatory Roles of WC1 and WC2 in Asexual Development and Photoreactivation of Beauveria bassiana. J. Fungi 2023, 9, 290. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Guo, C.-T.; Tong, S.-M.; Ying, S.-H.; Feng, M.-G. Two White Collar Proteins Protect Fungal Cells from Solar UV Damage by Their Interactions with Two Photolyases in Metarhizium robertsii. Environ. Microbiol. 2021, 23, 4925–4938. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Zhang, Y.-L.; Ying, S.-H.; Feng, M.-G. The Essential and the Nonessential Roles of Four Clock Elements in the Circadian Rhythm of Metarhizium robertsii. J. Biol. Chem. 2022, 8, 558. [Google Scholar] [CrossRef]
- de Leone, M.J.; Hernando, C.E.; Mora-García, S.; Yanovsky, M.J. It’s a Matter of Time: The Role of Transcriptional Regulation in the Circadian Clock-Pathogen Crosstalk in Plants. Transcription 2020, 11, 100–116. [Google Scholar] [CrossRef]
- Butt, G.R.; Qayyum, Z.A.; Jones, M.A. Plant Defence Mechanisms Are Modulated by the Circadian System. Biology 2020, 9, 454. [Google Scholar] [CrossRef]
- Hevia, M.A.; Canessa, P.; Müller-Esparza, H.; Larrondo, L.F. A Circadian Oscillator in the Fungus Botrytis cinerea Regulates Virulence When Infecting Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2015, 112, 8744–8749. [Google Scholar] [CrossRef]
- Canessa, P.; Schumacher, J.; Hevia, M.A.; Tudzynski, P.; Larrondo, L.F. Assessing the Effects of Light on Differentiation and Virulence of the Plant Pathogen Botrytis cinerea: Characterization of the White Collar Complex. PLoS ONE 2013, 8, e84223. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.-B.; Chen, N.; Zhu, X.-M.; Liang, S.; Li, L.; Wang, J.-Y.; Lu, J.-P.; Lin, F.-C.; Liu, X.-H. F-Box Proteins MoFwd1, MoCdc4 and MoFbx15 Regulate Development and Pathogenicity in the Rice Blast Fungus Magnaporthe oryzae. Environ. Microbiol. 2019, 21, 3027–3045. [Google Scholar] [CrossRef]
- Tiley, A.M.M.; Lawless, C.; Pilo, P.; Karki, S.J.; Lu, J.; Long, Z.; Gibriel, H.; Bailey, A.M.; Feechan, A. The Zymoseptoria tritici white collar-1 Gene, ZtWco-1, Is Required for Development and Virulence on Wheat. Fungal Genet. Biol. 2022, 161, 103715. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Singh, P.; Park, J.; Park, S.; Friedman, A.; Zheng, T.; Lee, Y.-H.; Lee, K. Genetic and Molecular Characterization of a Blue Light Photoreceptor MGWC-1 in Magnaporth oryzae. Fungal Genet. Biol. 2011, 48, 400–407. [Google Scholar] [CrossRef]
- Lee, K.; Singh, P.; Chung, W.-C.; Ash, J.; Kim, T.S.; Hang, L.; Park, S. Light Regulation of Asexual Development in the Rice Blast Fungus, Magnaporthe oryzae. Fungal Genet. Biol. 2006, 43, 694–706. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, H.-K.; Lee, S.; Yun, S.-H. The White Collar Complex Is Involved in Sexual Development of Fusarium graminearum. PLoS ONE 2015, 10, e0120293. [Google Scholar] [CrossRef]
- Ruiz-Roldán, M.C.; Garre, V.; Guarro, J.; Mariné, M.; Roncero, M.I.G. Role of the White Collar 1 Photoreceptor in Carotenogenesis, UV Resistance, Hydrophobicity, and Virulence of Fusarium oxysporum. Eukaryot. Cell 2008, 7, 1227–1230. [Google Scholar] [CrossRef] [Green Version]
- Cascant-Lopez, E.; Crosthwaite, S.K.; Johnson, L.J.; Harrison, R.J. No Evidence That Homologs of Key Circadian Clock Genes Direct Circadian Programs of Development or mRNA Abundance in Verticillium dahliae. Front. Microbiol. 2020, 11, 1977. [Google Scholar] [CrossRef]
- Pardo-Medina, J.; Limón, M.C.; Avalos, J. Fusarium Photoreceptors. J. Fungi 2023, 9, 319. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, J. Tools for Botrytis cinerea: New Expression Vectors Make the Gray Mold Fungus More Accessible to Cell Biology Approaches. Fungal Genet. Biol. 2012, 49, 483–497. [Google Scholar] [CrossRef]
- Fradin, E.F.; Thomma, B.P.H.J. Physiology and Molecular Aspects of Verticillium Wilt Diseases Caused by V. dahliae and V. albo-atrum. Mol. Plant Pathol. 2006, 7, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Klosterman, S.J.; Atallah, Z.K.; Vallad, G.E.; Subbarao, K.V. Diversity, Pathogenicity, and Management of Verticillium Species. Annu. Rev. Phytopathol. 2009, 47, 39–62. [Google Scholar] [CrossRef] [Green Version]
- Pegg, G.F.; Brady, B.L. Verticillium Wilts; CABI Publishing: Wallingford, UK, 2002; ISBN 978-1-84593-322-7. [Google Scholar]
- Carroll, C.L.; Carter, C.A.; Goodhue, R.E.; Lawell, C.-Y.C.L.; Subbarao, K.V. A Review of Control Options and Externalities for Verticillium Wilts. Phytopathology 2018, 108, 160–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eynck, C.; Koopmann, B.; Grunewaldt-Stoecker, G.; Karlovsky, P.; von Tiedemann, A. Differential Interactions of Verticillium longisporum and V. dahliae with Brassica napus Detected with Molecular and Histological Techniques. Eur. J. Plant Pathol. 2007, 118, 259–274. [Google Scholar] [CrossRef]
- Tian, J.; Kong, Z. Live-Cell Imaging Elaborating Epidermal Invasion and Vascular Proliferation/Colonization Strategy of Verticillium dahliae in Host Plants. Mol. Plant Pathol. 2022, 23, 895–900. [Google Scholar] [CrossRef]
- Singh, S.; Braus-Stromeyer, S.A.; Timpner, C.; Tran, V.-T.; Lohaus, G.; Reusche, M.; Knüfer, J.; Teichmann, T.; von Tiedemann, A.; Braus, G.H. Silencing of Vlaro2 for Chorismate Synthase Revealed That the Phytopathogen Verticillium longisporum Induces the Cross-Pathway Control in the Xylem. Appl. Microbiol. Biotechnol. 2010, 85, 1961–1976. [Google Scholar] [CrossRef] [Green Version]
- Maurus, I.; Harting, R.; Herrfurth, C.; Starke, J.; Nagel, A.; Mohnike, L.; Chen, Y.-Y.; Schmitt, K.; Bastakis, E.; Süß, M.T.; et al. Verticillium dahliae Vta3 Promotes ELV1 Virulence Factor Gene Expression in Xylem Sap, but Tames Mtf1-Mediated Late Stages of Fungus-Plant Interactions and Microsclerotia Formation. PLoS Pathog. 2023, 19, e1011100. [Google Scholar] [CrossRef]
- Maurus, I.; Leonard, M.; Nagel, A.; Starke, J.; Kronstad, J.W.; Harting, R.; Braus, G.H. Tomato Xylem Sap Hydrophobins Vdh4 and Vdh5 Are Important for Late Stages of Verticillium dahliae Plant Infection. J. Fungi 2022, 8, 1252. [Google Scholar] [CrossRef]
- Leonard, M.; Kühn, A.; Harting, R.; Maurus, I.; Nagel, A.; Starke, J.; Kusch, H.; Valerius, O.; Feussner, K.; Feussner, I.; et al. Verticillium longisporum Elicits Media-Dependent Secretome Responses with Capacity to Distinguish between Plant-Related Environments. Front. Microbiol. 2020, 11, 1876. [Google Scholar] [CrossRef]
- Bui, T.-T.; Harting, R.; Braus-Stromeyer, S.A.; Tran, V.-T.; Leonard, M.; Höfer, A.; Abelmann, A.; Bakti, F.; Valerius, O.; Schlüter, R.; et al. Verticillium dahliae Transcription Factors Som1 and Vta3 Control Microsclerotia Formation and Sequential Steps of Plant Root Penetration and Colonisation to Induce Disease. New Phytol. 2019, 221, 2138–2159. [Google Scholar] [CrossRef]
- Tian, L.; Yu, J.; Wang, Y.; Tian, C. The C2H2 Transcription Factor VdMsn2 Controls Hyphal Growth, Microsclerotia Formation, and Virulence of Verticillium dahliae. Fungal Biol. 2017, 121, 1001–1010. [Google Scholar] [CrossRef]
- Lai, M.; Cheng, Z.; Xiao, L.; Klosterman, S.J.; Wang, Y. The bZip Transcription Factor VdMRTF1 Is a Negative Regulator of Melanin Biosynthesis and Virulence in Verticillium dahliae. Microbiol. Spectr. 2022, 10, e0258121. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Mao, H.; Wei, Y.; Cai, J.; Xie, C.; Sui, A.; Yang, X.; Dong, J. The Fungal-Specific Transcription Factor Vdpf Influences Conidia Production, Melanized Microsclerotia Formation and Pathogenicity in Verticillium dahliae. Mol. Plant Pathol. 2016, 17, 1364–1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Qin, J.; Cao, Y.; Subbarao, K.V.; Chen, J.; Mandal, M.K.; Xu, X.; Shang, W.; Hu, X. Transcription Factor VdCf2 Regulates Growth, Pathogenicity, and the Expression of a Putative Secondary Metabolism Gene Cluster in Verticillium dahliae. Appl. Environ. Microbiol. 2022, 88, e0138522. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Jin, X.; Klosterman, S.J.; Wang, Y. Convergent and Distinctive Functions of Transcription Factors VdYap1, VdAtf1, and VdSkn7 in the Regulation of Nitrosative Stress Resistance, Microsclerotia Formation, and Virulence in Verticillium dahliae. Mol. Plant Pathol. 2020, 21, 1451–1466. [Google Scholar] [CrossRef] [PubMed]
- Tran, V.-T.; Braus-Stromeyer, S.A.; Kusch, H.; Reusche, M.; Kaever, A.; Kühn, A.; Valerius, O.; Landesfeind, M.; Aßhauer, K.; Tech, M.; et al. Verticillium Transcription Activator of Adhesion Vta2 Suppresses Microsclerotia Formation and Is Required for Systemic Infection of Plant Roots. New Phytol. 2014, 202, 565–581. [Google Scholar] [CrossRef]
- Harting, R.; Höfer, A.; Tran, V.-T.; Weinhold, L.-M.; Barghahn, S.; Schlüter, R.; Braus, G.H. The Vta1 Transcriptional Regulator Is Required for Microsclerotia Melanization in Verticillium dahliae. Fungal Biol. 2020, 124, 490–500. [Google Scholar] [CrossRef]
- Starke, J.; Harting, R.; Maurus, I.; Leonard, M.; Bremenkamp, R.; Heimel, K.; Kronstad, J.W.; Braus, G.H. Unfolded Protein Response and Scaffold Independent Pheromone MAP Kinase Signaling Control Verticillium dahliae Growth, Development, and Plant Pathogenesis. J. Fungi 2021, 7, 305. [Google Scholar] [CrossRef]
- Höfer, A.M.; Harting, R.; Aßmann, N.F.; Gerke, J.; Schmitt, K.; Starke, J.; Bayram, Ö.; Tran, V.-T.; Valerius, O.; Braus-Stromeyer, S.A.; et al. The Velvet Protein Vel1 Controls Initial Plant Root Colonization and Conidia Formation for Xylem Distribution in Verticillium Wilt. PLoS Genet. 2021, 17, e1009434. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, X.; Fang, Y.; Anchieta, A.; Goldman, P.H.; Hernandez, G.; Klosterman, S.J. Transcription Factor VdCmr1 Is Required for Pigment Production, Protection from UV Irradiation, and Regulates Expression of Melanin Biosynthetic Genes in Verticillium dahliae. Microbiology 2018, 164, 685–696. [Google Scholar] [CrossRef]
- Xiong, D.; Wang, Y.; Tian, L.; Tian, C. MADS-Box Transcription Factor VdMcm1 Regulates Conidiation, Microsclerotia Formation, Pathogenicity, and Secondary Metabolism of Verticillium dahliae. Front. Microbiol. 2016, 7, 1192. [Google Scholar] [CrossRef] [Green Version]
- Sarmiento-Villamil, J.L.; Prieto, P.; Klosterman, S.J.; García-Pedrajas, M.D. Characterization of Two Homeodomain Transcription Factors with Critical but Distinct Roles in Virulence in the Vascular Pathogen Verticillium dahliae. Mol. Plant Pathol. 2018, 19, 986–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarmiento-Villamil, J.L.; García-Pedrajas, N.E.; Baeza-Montañez, L.; García-Pedrajas, M.D. The APSES Transcription Factor Vst1 Is a Key Regulator of Development in Microsclerotium- and Resting Mycelium-Producing Verticillium Species. Mol. Plant Pathol. 2018, 19, 59–76. [Google Scholar] [CrossRef] [Green Version]
- Fradin, E.F.; Zhang, Z.; Juarez Ayala, J.C.; Castroverde, C.D.M.; Nazar, R.N.; Robb, J.; Liu, C.-M.; Thomma, B.P.H.J. Genetic Dissection of Verticillium Wilt Resistance Mediated by Tomato Ve1. Plant Physiol. 2009, 150, 320–332. [Google Scholar] [CrossRef] [Green Version]
- Yates, A.D.; Allen, J.; Amode, R.M.; Azov, A.G.; Barba, M.; Becerra, A.; Bhai, J.; Campbell, L.I.; Carbajo Martinez, M.; Chakiachvili, M.; et al. Ensembl Genomes 2022: An Expanding Genome Resource for Non-Vertebrates. Nucleic Acids Res. 2022, 50, D996–D1003. [Google Scholar] [CrossRef] [PubMed]
- Basenko, E.Y.; Pulman, J.A.; Shanmugasundram, A.; Harb, O.S.; Crouch, K.; Starns, D.; Warrenfeltz, S.; Aurrecoechea, C.; Stoeckert, C.J., Jr.; Kissinger, J.C.; et al. FungiDB: An Integrated Bioinformatic Resource for Fungi and Oomycetes. J. Fungi 2018, 4, 39. [Google Scholar] [CrossRef] [Green Version]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2022, 50, D20–D26. [Google Scholar] [CrossRef] [PubMed]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L.; et al. InterPro in 2022. Nucleic Acids Res. 2023, 51, D418–D427. [Google Scholar] [CrossRef]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and Sequence Analysis Tools Services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, Scalable Generation of High-Quality Protein Multiple Sequence Alignments Using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Thumuluri, V.; Almagro Armenteros, J.J.; Rosenberg Johansen, A.; Nielsen, H.; Winther, O. DeepLoc 2.0: Multi-Label Subcellular Localization Prediction Using Protein Language Models. Nucleic Acids Res. 2022, 50, W228–W234. [Google Scholar] [CrossRef] [PubMed]
- Marx, A.; Backes, C.; Meese, E.; Lenhof, H.-P.; Keller, A. EDISON-WMW: Exact Dynamic Programing Solution of the Wilcoxon-Mann-Whitney Test. Genom. Proteom. Bioinform. 2016, 14, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertani, G. Studies on Lysogenesis. I. The Mode of Phage Liberation by Lysogenic Escherichia coli. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Jessee, J.; Bloom, F.R. Plasmid Transformation of Escherichia coli and Other Bacteria. Meth. Enzymol. 1991, 204, 63–113. [Google Scholar] [CrossRef]
- Inoue, H.; Nojima, H.; Okayama, H. High Efficiency Transformation of Escherichia coli with Plasmids. Gene 1990, 96, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Lazo, G.R.; Stein, P.A.; Ludwig, R.A. A DNA Transformation-Competent Arabidopsis Genomic Library in Agrobacterium. Nat. Biotechnol. 1991, 9, 963–967. [Google Scholar] [CrossRef] [PubMed]
- Jyothishwaran, G.; Kotresha, D.; Selvaraj, T.; Srideshikan, S.M.; Rajvanshi, P.K.; Jayabaskaran, C. A Modified Freeze-Thaw Method for Efficient Transformation of Agrobacterium tumefaciens. Curr. Sci. 2007, 93, 770–772. [Google Scholar]
- Mullins, E.D.; Chen, X.; Romaine, P.; Raina, R.; Geiser, D.M.; Kang, S. Agrobacterium-Mediated Transformation of Fusarium oxysporum: An Efficient Tool for Insertional Mutagenesis and Gene Transfer. Phytopathology 2001, 91, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Kolar, M.; Punt, P.J.; van den Hondel, C.A.M.J.J.; Schwab, H. Transformation of Penicillium chrysogenum Using Dominant Selection Markers and Expression of an Escherichia coli LacZ Fusion Gene. Gene 1988, 62, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Meister, C.; Thieme, K.G.; Thieme, S.; Köhler, A.M.; Schmitt, K.; Valerius, O.; Braus, G.H. COP9 Signalosome Interaction with Uspa/Usp15 Deubiquitinase Controls VeA-Mediated Fungal Multicellular Development. Biomolecules 2019, 9, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Rappsilber, J.; Mann, M.; Ishihama, Y. Protocol for Micro-Purification, Enrichment, Pre-Fractionation and Storage of Peptides for Proteomics Using StageTips. Nat. Protoc. 2007, 2, 1896–1906. [Google Scholar] [CrossRef]
- Rappsilber, J.; Ishihama, Y.; Mann, M. Stop and Go Extraction Tips for Matrix-Assisted Laser Desorption/Ionization, Nanoelectrospray, and LC/MS Sample Pretreatment in Proteomics. Anal. Chem. 2003, 75, 663–670. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant Enables High Peptide Identification Rates, Individualized p.p.b.-Range Mass Accuracies and Proteome-Wide Protein Quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus Computational Platform for Comprehensive Analysis of (Prote)Omics Data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE Database Resources in 2022: A Hub for Mass Spectrometry-Based Proteomics Evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Liu, X.; Li, H.; Fan, Y.; Duan, J.; Liu, Y.; He, Q. Transcription Factor CBF-1 Is Critical for Circadian Gene Expression by Modulating WHITE COLLAR Complex Recruitment to the frq Locus. PLoS Genet. 2018, 14, e1007570. [Google Scholar] [CrossRef] [PubMed]
- Crosthwaite, S.K.; Dunlap, J.C.; Loros, J.J. Neurospora wc-1 and wc-2: Transcription, Photoresponses, and the Origins of Circadian Rhythmicity. Science 1997, 276, 763–769. [Google Scholar] [CrossRef]
- Gai, K.; Cao, X.; Dong, Q.; Ding, Z.; Wei, Y.; Liu, Y.; Liu, X.; He, Q. Transcriptional Repression of frequency by the IEC-1-INO80 Complex Is Required for Normal Neurospora Circadian Clock Function. PLoS Genet. 2017, 13, e1006732. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Li, H.; Liu, Q.; Niu, Y.; Hu, Q.; Deng, H.; Cha, J.; Wang, Y.; Liu, Y.; He, Q. Role for Protein Kinase A in the Neurospora Circadian Clock by Regulating White Collar-Independent frequency Transcription through Phosphorylation of RCM-1. Mol. Cell. Biol. 2015, 35, 2088–2102. [Google Scholar] [CrossRef] [Green Version]
- Sun, G.; Zhou, Z.; Liu, X.; Gai, K.; Liu, Q.; Cha, J.; Kaleri, F.N.; Wang, Y.; He, Q. Suppression of WHITE COLLAR-Independent frequency Transcription by Histone H3 Lysine 36 Methyltransferase SET-2 Is Necessary for Clock Function in Neurospora. J. Biol. Chem. 2016, 291, 11055–11063. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Liu, X.; Hu, Q.; Zhang, N.; Sun, G.; Cha, J.; Wang, Y.; Liu, Y.; He, Q. Suppression of WC-Independent frequency Transcription by RCO-1 Is Essential for Neurospora Circadian Clock. Proc. Natl. Acad. Sci. USA 2013, 110, E4867–E4874. [Google Scholar] [CrossRef]
- Luo, C.; Loros, J.J.; Dunlap, J.C. Nuclear Localization Is Required for Function of the Essential Clock Protein FRQ. EMBO J. 1998, 17, 1228–1235. [Google Scholar] [CrossRef] [Green Version]
- Tsvetkov, P.; Reuven, N.; Shaul, Y. The Nanny Model for IDPs. Nat. Chem. Biol. 2009, 5, 778–781. [Google Scholar] [CrossRef]
- Liang, S.; Hitomi, M.; Hu, Y.-H.; Liu, Y.; Tartakoff, A.M. A DEAD-Box-Family Protein Is Required for Nucleocytoplasmic Transport of Yeast mRNA. Mol. Cell. Biol. 1996, 16, 5139–5146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de la Cruz, J.; Kressler, D.; Tollervey, D.; Linder, P. Dob1p (Mtr4p) Is a Putative ATP-Dependent RNA Helicase Required for the 3′ End Formation of 5.8S rRNA in Saccharomyces cerevisiae. EMBO J. 1998, 17, 1128–1140. [Google Scholar] [CrossRef] [Green Version]
- Reiter, F.; Wienerroither, S.; Stark, A. Combinatorial Function of Transcription Factors and Cofactors. Curr. Opin. Genet. Dev. 2017, 43, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Covert, S.F.; Kapoor, P.; Lee, M.; Briley, A.; Nairn, C.J. Agrobacterium tumefaciens-Mediated Transformation of Fusarium circinatum. Mycol. Res. 2001, 105, 259–264. [Google Scholar] [CrossRef]
- Zhou, L.; Zhao, J.; Guo, W.; Zhang, T. Functional Analysis of Autophagy Genes via Agrobacterium-Mediated Transformation in the Vascular Wilt Fungus Verticillium dahliae. J. Genet. Genomics 2013, 40, 421–431. [Google Scholar] [CrossRef] [PubMed]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagel, A.; Leonard, M.; Maurus, I.; Starke, J.; Schmitt, K.; Valerius, O.; Harting, R.; Braus, G.H. The Frq–Frh Complex Light-Dependently Delays Sfl1-Induced Microsclerotia Formation in Verticillium dahliae. J. Fungi 2023, 9, 725. https://doi.org/10.3390/jof9070725
Nagel A, Leonard M, Maurus I, Starke J, Schmitt K, Valerius O, Harting R, Braus GH. The Frq–Frh Complex Light-Dependently Delays Sfl1-Induced Microsclerotia Formation in Verticillium dahliae. Journal of Fungi. 2023; 9(7):725. https://doi.org/10.3390/jof9070725
Chicago/Turabian StyleNagel, Alexandra, Miriam Leonard, Isabel Maurus, Jessica Starke, Kerstin Schmitt, Oliver Valerius, Rebekka Harting, and Gerhard H. Braus. 2023. "The Frq–Frh Complex Light-Dependently Delays Sfl1-Induced Microsclerotia Formation in Verticillium dahliae" Journal of Fungi 9, no. 7: 725. https://doi.org/10.3390/jof9070725