
Transcription Factor VM1G_06867: A Requirement for Growth, Pathogenicity, Development, and Maintenance of Cell Wall Integrity in Valsa mali
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Bioinformatic Analysis
2.3. Identification, VM1G_06867 Gene Replacement, and Complementation
2.4. Double Deletion Genes VM1G 06867 and VmSom1
2.5. Growth Rate Determination
2.6. Conidial Production and Germination
2.7. Stress Sensitivity Assay
2.8. Analysis of the Effect of Different Carbon-Nitrogen on the Growth of Gene VM1G_06867
2.9. Infection Assays on Apple Fruit and Twigs
2.10. Statistical Analysis
3. Results
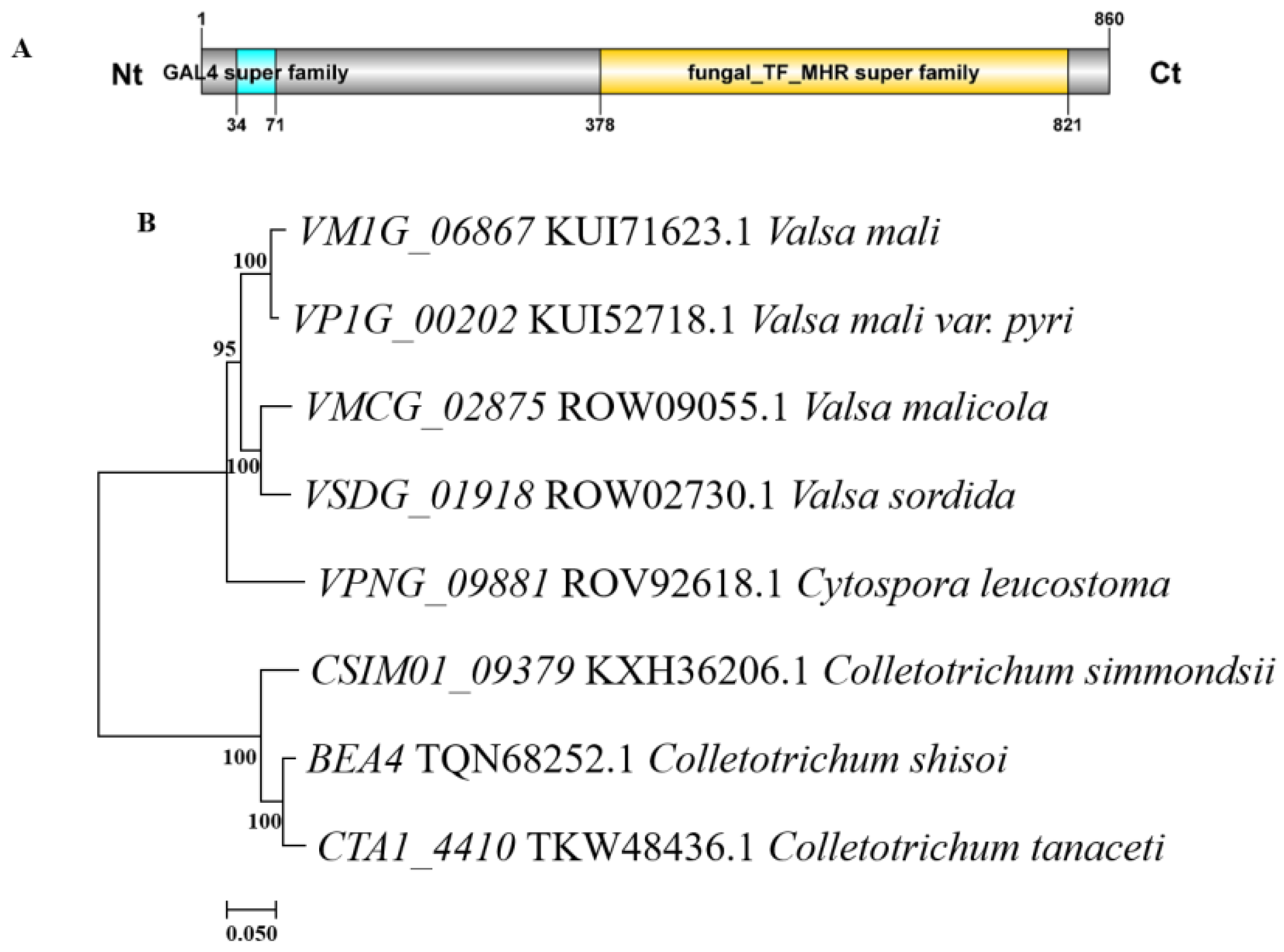
3.1. Bioinformatical Characterizations of VM1G_06867 in V. mali
3.2. Role of VM1G_06867 in Growth
3.3. Negative Regulation of Conidiation and Impact on Conidial Germination by VM1G_06867
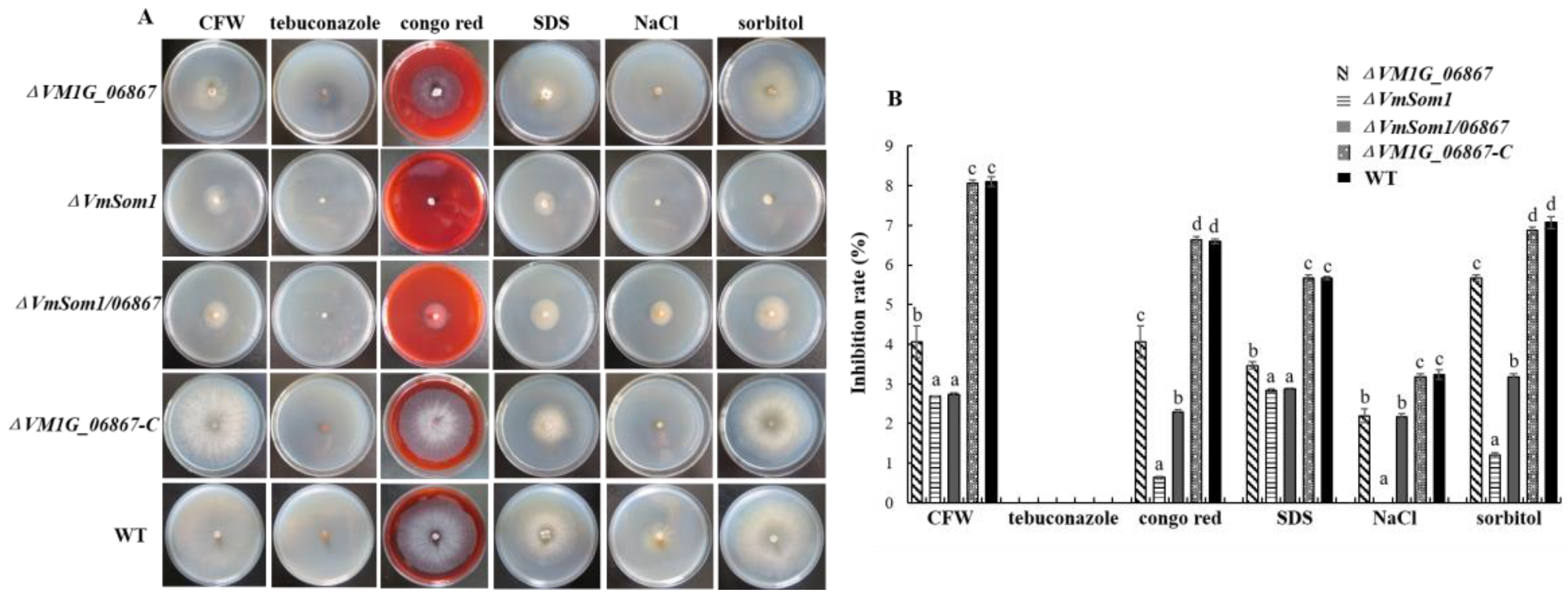
3.4. Stress Influence of VM1G_06867 on Response to Osmotic Stress and Cell-Wall Integrity Inhibitor
3.5. Impact of VM1G_06867 on Utilization of Carbon and Nitrogen Sources
3.6. Absence of VM1G_06867 in Pathogenicity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, C.X.; Li, C.; Li, B.H.; Li, G.F.; Dong, X.L.; Wang, G.P.; Zhang, Q.M. Toxins produced by Valsa mali var. mali and their relationship with Pathogenicity. Toxins 2014, 6, 1139–1154. [Google Scholar] [PubMed] [Green Version]
- Li, B.H.; Wang, C.X.; Dong, X.L. Research progress in apple diseases and problems in the diseases management in China. Plant Prot. 2013, 39, 46–54. [Google Scholar]
- Cui, X.Q.; Ding, Z.; Wu, L.; Che, Q.; Li, D.H.; Gu, Q.Q.; Zhu, T.J. Research progress on the pathway-specific transcription factors and their applocation in activating cryptic fungal secondary metabolites. Chin. J. Mar. Drugs 2017, 36, 75–82. [Google Scholar]
- Wu, Y.X.; Xu, L.S.; Yin, Z.Y.; Hao, F.; Huang, L.L. Transcription factor VmSeb1 is required for the growth, development, and virulence in Valsa mali. Microb. Pathog. 2018, 123, 132–138. [Google Scholar] [CrossRef]
- Shelest, E. Transcription factors in fungi. FEMS Microbiol. Lett. 2008, 286, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Benjamin, P.K.; Nancy, P.K. Key players in the regulation of fungal secondary metabolism. Fungal Biol. 2015, 2, 13–28. [Google Scholar]
- Chung, H.; Choi, J.; Park, S.Y.; Jeon, J.; Lee, Y.H. Two conidiation-related Zn2Cys6 transcription factor genes in the rice blast fungus. Fungal Genet. Biol. 2013, 61, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.Z.; Waalwijk, C.; Wit, P.J.G.M.D.; Lee, T.V.D.; Tang, D.Z. EBR1, a novel Zn(2)Cys(6) transcription factor, affects virulence and apical dominance of the hyphal tip in Fusarium graminearum. Mol. Plant-Mol. Interact. MPMI 2011, 24, 1407–1418. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.P.; Cao, H.J.; Zhang, L.L.; Huang, P.Y.; Lin, F.C. Systematic analysis of Zn2Cys6 transcription factors required for development and pathogenicity by high-throughput gene knockout in the rice blast fungus. PLoS Pathog. 2014, 10, el004432. [Google Scholar] [CrossRef]
- Tanaka, A.; Cartwright, G.M.; Saikia, S.; Kayano, Y.; Takemoto, D.; Kato, M.; Tsuge, T.; Scott, B. ProA, a transcriptional regulator of fungal fruiting body development, regulates leaf hyphal network development in the Epichloe festucae-Lolium perenne symbiosis. Mol. Microbiol. 2013, 90, 551–568. [Google Scholar] [CrossRef]
- Boyce, K.J.; McLauchlan, A.; Schreider, L.; Andrianopoulos, A. Intracellular growth is dependent on tyrosine catabolism in the dimorphic fungal pathogen Penicillium marneffei. PLoS Pathog. 2015, 11, e1004790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rispail, N.; Soanes, D.M.; Ant, C.; Czajkowski, R.; Grünler, A.; Huguet, R.; Perez-Nadales, E.; Poli, A.; Sartorel, E.; Valiante, V.; et al. Comparative genomics of MAP kinase and calcium-calcineurin signaling components in plant and human pathogenic fungi. Fungal Genet. Biol. 2009, 46, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Tri-Thuc, B.; Harting, R.; Braus-Stromeyer, S.A.; Van-Tuan, T.; Leonard, M.; Hoefer, A.; Abelmann, A.; Bakti, F.; Valerius, O.; Schlueter, R.; et al. Verticillium dahliae transcription factors Som1 and Vta3 control microsclerotia formation and sequential steps of plant root penetration and colonisation to induce disease. New Phytol. 2018, 221, 2138–2159. [Google Scholar]
- Yan, X.; Li, Y.; Yue, X.F.; Wang, C.C.; Que, Y.W.; Kong, D.D.; Ma, Z.H.; Talbot, N.J.; Wang, Z.Y. Two novel transcriptional regulators are essential for infection-related morphogenesis and pathogenicity of the rice blast fungus Magnaporthe oryzae. PLoS Pathog. 2011, 7, e1002385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.J.; Sasse, C.; Gerke, J.; Valerius, O.; Irmer, H.; Frauendorf, H.; Heinekamp, T.; Strβburger, M.; Tran, V.T.; Herzog, B.; et al. Transcription factor SomA is required for adhesion, development and virulence of the human pathogen Aspergillus fumigatus. PLoS Pathog. 2015, 11, e1005205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.; Jin, K.; Xia, Y. Involvement of MaSom1, a downstream transcriptional factor of cAMP/PKA pathway, in conidial yield, stress tolerances, and virulence in Metarhizium acridum. Appl. Microbiol. Biotechnol. 2018, 102, 5611–5623. [Google Scholar] [CrossRef]
- Yin, Z.Y.; Liu, H.Q.; Li, Z.P.; Ke, X.W.; Dou, D.L.; Gao, X.N.; Song, N.; Dai, Q.Q.; Wu, Y.X.; Xu, J.R.; et al. Genome sequence of Valsa canker pathogens uncovers a potential adaptation of colonization of woody bark. New Phytol. 2015, 208, 1202–1216. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Alan, F.; Sudhir, K. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.H.; Hamari, Z.; Han, K.H.; Seo, J.A.; Reyes-Domínguez, Y.; Scazzocchio, C. Double-joint PCR: A PCR-based molecular tool for gene manipulations in filamentous fungi. Fungal Genet. Biol. 2004, 41, 973–981. [Google Scholar] [CrossRef]
- Chen, L.; Lun, Y.Y.; Sun, G.W.; Zhao, Y.T.; He, B.L.; Wang, H.K.; Liu, H.X. Preparation of protoplasm and optimization of regeneration conditions of Valsa mali. Sci. Agric. Sin. 2014, 46, 109–112. [Google Scholar]
- Zhou, X.Y.; Zhang, H.F.; Li, G.T.; Shaw, B.; Xu, J.R. The Cyclase-associated protein Cap1 is important for proper regulation of infection-related morphogenesis in Magnaporthe oryzae. PLoS Pathog. 2012, 8, 503–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhtar, A.; Becker, P.B. The histone H4 acetyltransferase MOF uses a C2HC zinc finger for substrate recognition. EMBO Rep. 2001, 2, 113–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cass, D.; Hotchko, R.; Barber, P.; Jones, K.; Gates, D.P.; Berglund, J.A. The four Zn fingers of MBNL1 provide a flexible platform for recognition of its RNA binding elements. BMC Mol. Biol. 2011, 12, 20. [Google Scholar] [CrossRef] [Green Version]
- Son, H.; Sco, Y.S.; Min, K.; Park, A.R.; Lcc, J.; Jin, J.M.; Lin, Y.; Cao, P.; Hong, S.Y.; Kim, E.K.; et al. Agmhenome-based functiona analysis of transcription factors in the cereal head blight fungus, Fusarium gaminearum. PLoS Pathog. 2011, 7, e100231. [Google Scholar] [CrossRef]
- Cao, H.J. Analysis of Transcription Factor Function of Zn2Cys6 and bHLH Family in Magnaporthe oryzae; Zhejiang University: Hangzhou, China, 2015. [Google Scholar]
- Bugeja, H.E.; Hynes, M.J.; Andrianopoulos, A. HgrA is necessary and suffiffifficient to drive hyphal growth in the dimorphic pathogen Penicillium marneffffei. Mol. Microbiol. 2013, 88, 998–1014. [Google Scholar] [CrossRef]
- Chang, P.K.; Scharfenstein, L.L.; Luo, M.; Mahoney, N.; Molyneux, R.J.; Yu, J.J.; Brown, R.L.; Campbell, B.C. Loss of msnA, a putative stress regulatory gene, in Aspergillus parasiticus and Aspergillus flflavus increased production of conidia, aflflatoxins and kojic acid. Toxins 2011, 3, 82–104. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, A.; Ohm, R.A.; Oxiles, L.; Brooks, F.; Lawrence, C.B.; Grigoriev, I.V.; Cho, Y. A zinc-fifinger-family transcription factor, AbVf19, is required for the induction of a gene subset important for virulence in Alternaria brassicicola. Mol. Plant Microb. Interact. 2012, 25, 443–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, K.R.; Wouw, A.P.V.D.; Gardiner, D.M.; Elliott, C.E.; Idnurm, A. A conserved Zn2Cys6 transcription factor, identified in a spontaneous mutant from in vitro passaging, is involved in pathogenicity of the blackleg fungus Leptosphaeria maculans. Fungal Biol. 2021, 125, 541–550. [Google Scholar] [CrossRef]
- Samantaray, S.; Neubauer, M.; Helmschrott, C.; Wagener, J. Role of the Guanine Nucleotide Exchange Factor Rom2 in Cell Wall Integrity Maintenance of Aspergillus fumigatus. Eukaryot. Cell 2013, 12, 288–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, T.; Inamine, T.; Yamauchi, S.; Nagayoshi, Y.; Saijo, T.; Izumikawa, K.; Seki, M.; Kakeya, H.; Yamamoto, Y.; Yanajihara, K.; et al. Role of the Slt2 mitogen-activated protein kinase pathway in cell wall integrity and virulence in Candida glabrata. FEMS Yeast Res. 2010, 10, 343–352. [Google Scholar] [CrossRef] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diao, Y.; Jin, J.; Xiong, X.; Yu, C.; Tian, Y.; Li, D.; Liu, H. Transcription Factor VM1G_06867: A Requirement for Growth, Pathogenicity, Development, and Maintenance of Cell Wall Integrity in Valsa mali. J. Fungi 2023, 9, 692. https://doi.org/10.3390/jof9060692
Diao Y, Jin J, Xiong X, Yu C, Tian Y, Li D, Liu H. Transcription Factor VM1G_06867: A Requirement for Growth, Pathogenicity, Development, and Maintenance of Cell Wall Integrity in Valsa mali. Journal of Fungi. 2023; 9(6):692. https://doi.org/10.3390/jof9060692
Chicago/Turabian StyleDiao, Yufei, Jiyang Jin, Xiong Xiong, Chengming Yu, Yehan Tian, Duochuan Li, and Huixiang Liu. 2023. "Transcription Factor VM1G_06867: A Requirement for Growth, Pathogenicity, Development, and Maintenance of Cell Wall Integrity in Valsa mali" Journal of Fungi 9, no. 6: 692. https://doi.org/10.3390/jof9060692