
Integration of Physiological, Transcriptomic and Metabolomic Reveals Molecular Mechanism of Paraisaria dubia Response to Zn2+ Stress
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strain and Zn2+ Treatment
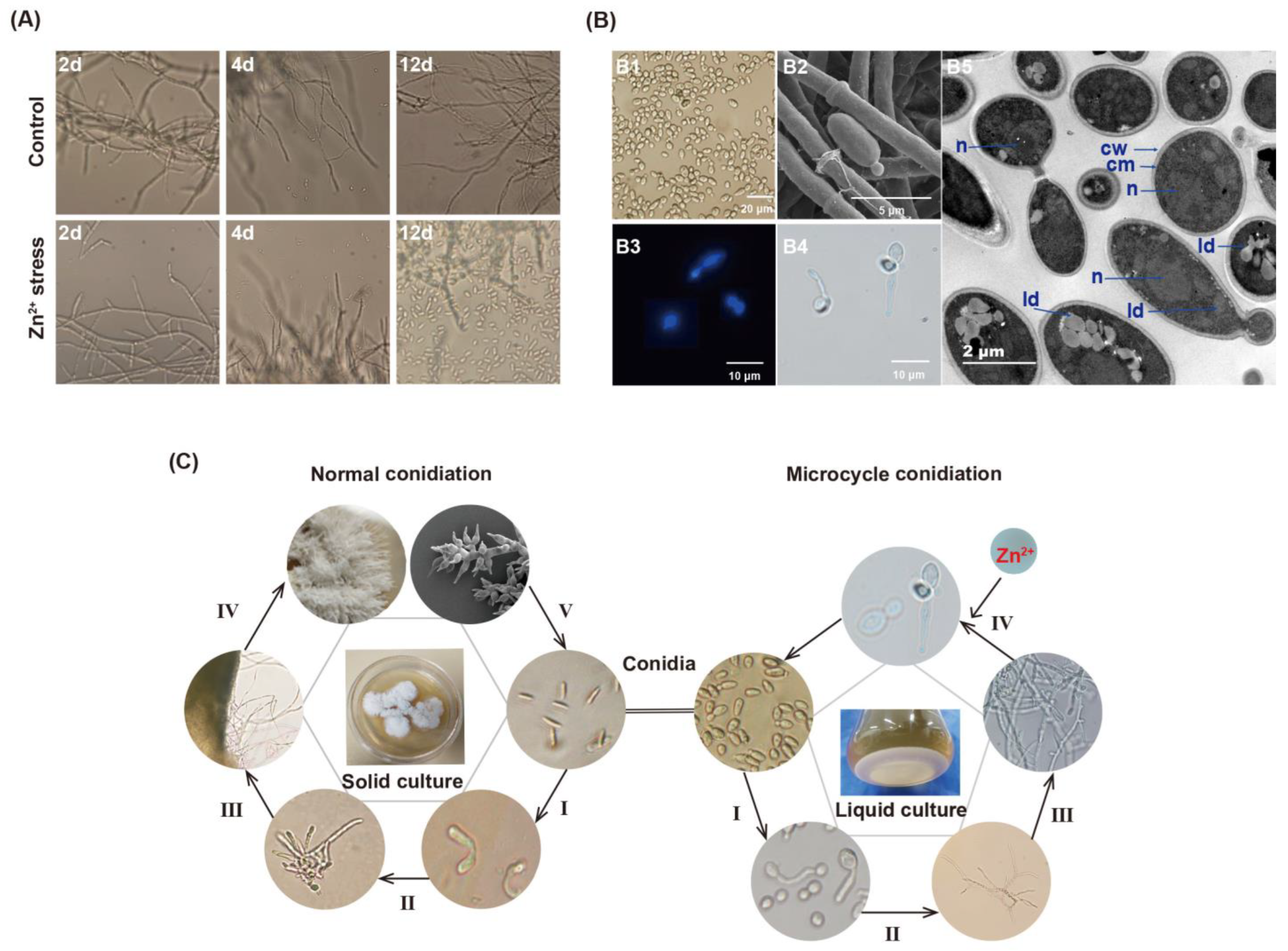
2.2. Microscopic Morphological Analysis
2.3. Preparation of IPS and EPS
2.4. Zn2+ Concentration Determination and Removal Rate
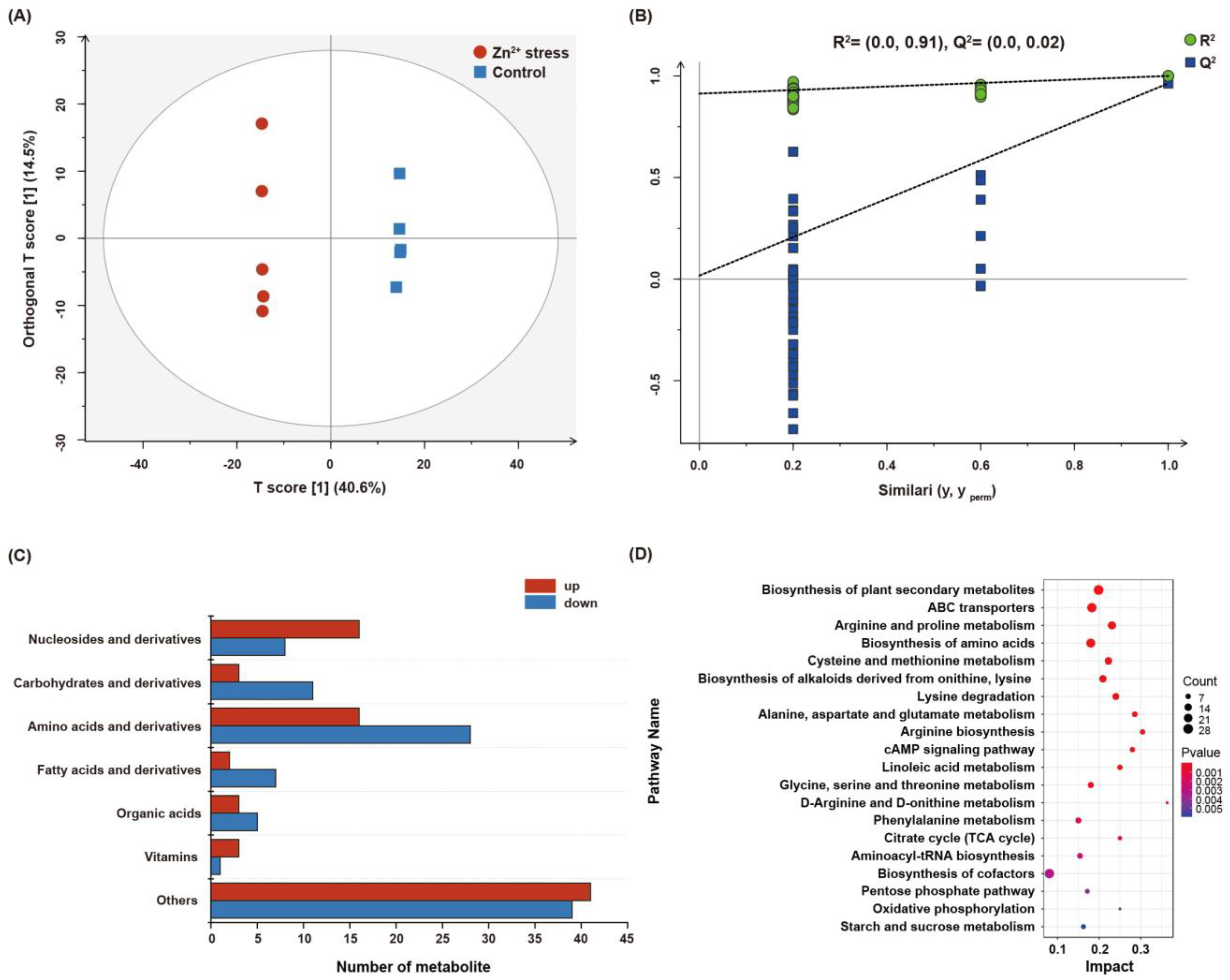
2.5. Nontargeted Metabolomic Profiling Analysis
2.6. Transcriptomic Analysis
2.7. Statistical Analysis
3. Results
3.1. Morphological Alterations of Mycelia and Reproduction of P. dubia under Zn2+ Stress
3.2. Physiological Characteristic and Zn2+ Removal Capacity of P. dubia under Zn2+ Stress
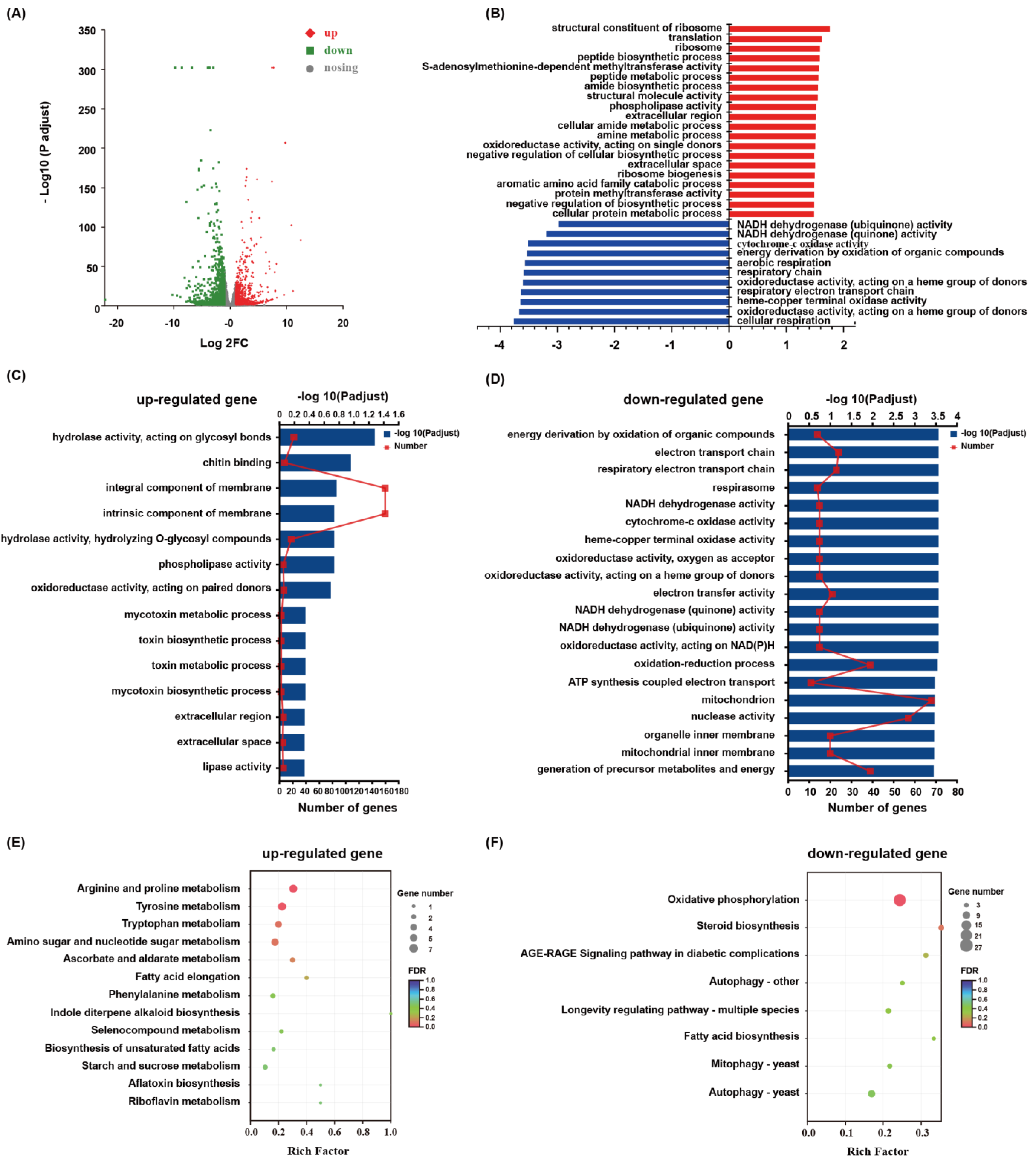
3.3. Transcriptomic Sequencing and DEGs Identification
3.4. Functional Annotation and Enrichment of DEGs
3.5. Critical Functional Genes in Response to Zn2+ Stress
3.6. Metabolic Changes of P. dubia under Zn2+ Stress
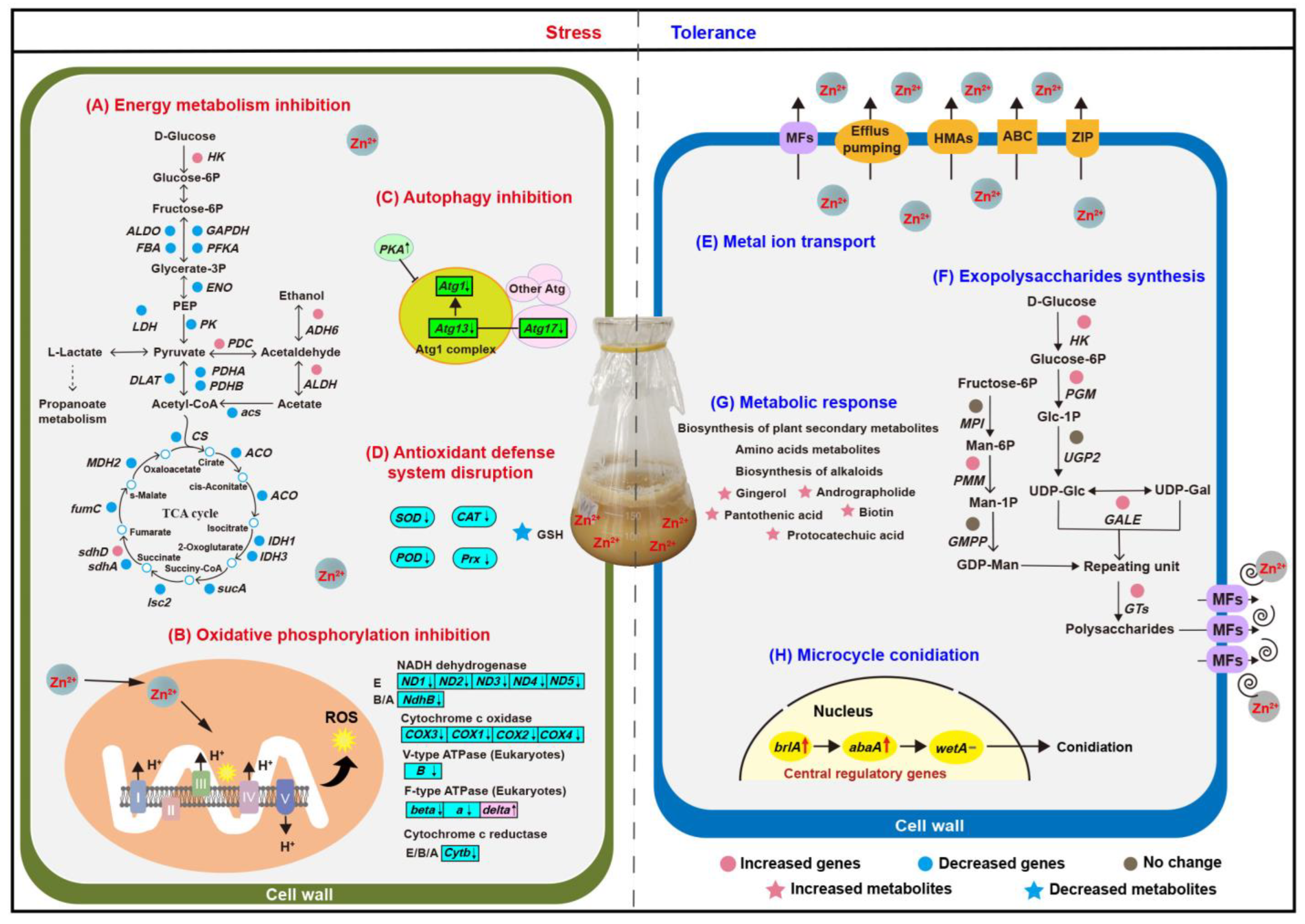
4. Discussion
4.1. Zn2+ Stress Affected in Energy Metabolism, Oxidative Stress, and Autophagy
4.2. Lead-Induced Detoxification and Defense Mechanisms
4.2.1. Metal Ion Transport Response to Zn2+ Stress
4.2.2. Exopolysaccharides Synthesis Promotion Response to Zn2+ Stress
4.2.3. Metabolic Response to Zn2+ Stress
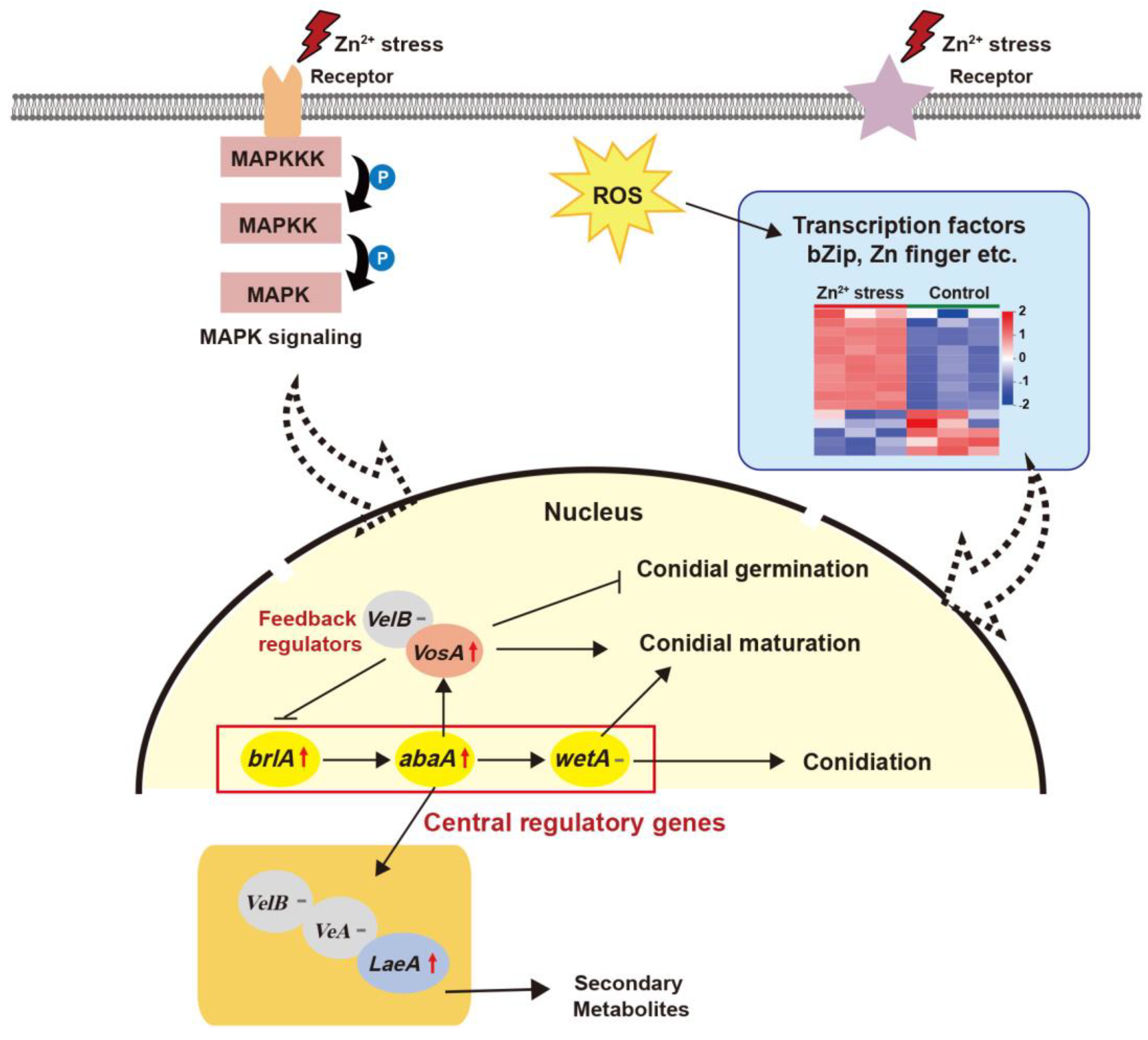
4.2.4. Reproduction Response Based on MC in Zn2+ Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jiang, D.; Tan, M.T.; Zheng, L.; Wu, H.F.; Li, Y.N.; Yan, S.C. Cd exposure-triggered metabolic disruption increases the susceptibility of Lymantria dispar (Lepidoptera: Erebidae) larvae to Mamestra brassicae nuclear polyhedrosis virus: A multi-omits study. Ecotoxicol. Environ. Saf. 2022, 232, 113280. [Google Scholar] [CrossRef]
- Bi, X.; Feng, X.; Yang, Y.; Qiu, G.; Li, G.; Li, F.; Liu, T.; Fu, Z.; Jin, Z. Environmental contamination of heavy metals from zinc smelting areas in Hezhang County, western Guizhou, China. Environ. Int. 2006, 32, 883–890. [Google Scholar] [CrossRef]
- Long, X.X.; Yang, X.E.; Ni, W.Z.; Ye, Z.Q.; He, Z.L.; Calvert, D.V.; Stoffella, J.P. Assessing zinc thresholds for phytotoxicity and potential dietary toxicity in selected vegetable crops. Commun. Soil Sci. Plan. 2003, 34, 1421–1434. [Google Scholar] [CrossRef]
- Arrivault, S.; Senger, T.; Kramer, U. The Arabidopsis metal tolerance protein AtMTP3 maintains metal homeostasis by mediating Zn exclusion from the shoot under Fe deficiency and Zn oversupply. Plant J. 2006, 46, 861–879. [Google Scholar] [CrossRef]
- Natasha, N.; Shahid, M.; Bibi, I.; Iqbal, J.; Khalid, S.; Murtaza, B.; Bakhat, H.F.; Farooq, A.U.; Amjad, M.; Hammad, H.M.; et al. Zinc in soil-plant-human system: A data-analysis review. Sci. Total Environ. 2022, 20, 152024. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, H.; Carmona, V.M.V.; Inocencio, M.F.; Neto, A.E.F.; Cecilio, A.B.; Mauad, M. Soil Type and Zinc Doses in Agronomic Biofortification of Lettuce Genotypes. Agronomy 2020, 10, 124. [Google Scholar] [CrossRef] [Green Version]
- Aydin, S.S.; Gokce, E.; Buyuk, I.; Aras, S. Characterization of stress induced by copper and zinc on cucumber (Cucumis sativus L.) seedlings by means of molecular and population parameters. Mutat. Res. 2012, 746, 49–55. [Google Scholar] [CrossRef]
- Garg, N.; Singh, S. Arbuscular Mycorrhiza Rhizophagus irregularis and Silicon Modulate Growth, Proline Biosynthesis and Yield in Cajanus cajan L. Millsp (pigeonpea) Genotypes Under Cadmium and Zinc Stress. J. Plant Growth Regul. 2018, 37, 46–63. [Google Scholar] [CrossRef]
- Tewari, R.K.; Kumar, P.; Sharma, P.N. Morphology and physiology of zinc-stressed mulberry plants. J. Plant Nutr. Soil Sci. 2008, 171, 286–294. [Google Scholar] [CrossRef]
- Michael, P.I.; Krishnaswamy, M. The effect of zinc stress combined with high irradiance stress on membrane damage and antioxidative response in bean seedlings. Environ. Exp. Bot. 2011, 74, 171–177. [Google Scholar] [CrossRef]
- Ab Rhaman, S.M.S.; Naher, L.; Siddiquee, S. Mushroom Quality Related with Various Substrates' Bioaccumulation and Translocation of Heavy Metals. J. Fungi 2022, 8, 42. [Google Scholar] [CrossRef]
- Nordgren, A.; Baath, E.; Soderstrom, B. Microfungi and microbial activity along a heavy metal gradient. Appl. Environ. Microbiol. 1983, 45, 1829–1837. [Google Scholar] [CrossRef] [Green Version]
- Sung, G.H.; Hywel-Jones, N.L.; Sung, J.M.; Luangsa-Ard, J.J.; Shrestha, B.; Spatafora, J.W. Phylogenetic classification of Cordyceps and the clavicipitaceous fungi. Stud. Mycol. 2007, 57, 5–59. [Google Scholar] [CrossRef] [Green Version]
- Kepler, R.M.; Sung, G.H.; Harada, Y.; Tanaka, K.; Tanaka, E.; Hosoya, T.; Bischoff, J.F.; Spatafora, J.W. Host jumping onto close relatives and across kingdoms by Tyrannicordyceps (Clavicipitaceae) gen. nov. and Ustilaginoidea_(Clavicipitaceae). Am. J. Bot. 2012, 99, 552–561. [Google Scholar] [CrossRef]
- Kepler, R.; Ban, S.; Nakagiri, A.; Bischoff, J.; Hywel-Jones, N.; Owensby, C.A.; Spatafora, J.W. The phylogenetic placement of hypocrealean insect pathogens in the genus Polycephalomyces: An application of One Fungus One Name. Fungal. Biol. 2013, 117, 611–622. [Google Scholar] [CrossRef]
- Tong, L.L.; Wang, Y.; Yuan, L.; Liu, M.Z.; Du, Y.H.; Mu, X.Y.; Yang, Q.H.; Wei, S.X.; Li, J.Y.; Wang, M.A.; et al. Enhancement of polysaccharides production using microparticle enhanced technology by Paraisaria dubia. Microb. Cell Fact. 2022, 21, 12. [Google Scholar] [CrossRef]
- Perez-Villamares, J.C.; Burrola-Aguilar, C.; Aguilar-Miguel, X.; Sanjuan, T.; Jimenez-Sanchez, E. New records of entomopathogenous fungi of the genus Cordyceps s. l. (Ascomycota: Hypocreales) from the State of Mexico. Rev. Mex. Biodivers. 2017, 88, 773–783. [Google Scholar] [CrossRef]
- Tong, L.L.; Wang, Y.; Du, Y.H.; Yuan, L.; Liu, M.Z.; Mu, X.Y.; Chen, Z.L.; Zhang, Y.D.; He, S.J.; Li, X.J.; et al. Transcriptomic Analysis of Morphology Regulatory Mechanisms of Microparticles to Paraisaria dubia in Submerged Fermentation. Appl Biochem Biotechnol. 2022, 194, 4333–4347. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wang, S.; Hao, Q.X.; Kang, L.P.; Kang, C.Z.; Yang, J.; Yang, W.Z.; Jiang, J.Y.; Huang, L.Q.; Guo, L.P. Bioaccessibility and risk assessment of heavy metals, and analysis of arsenic speciation in Cordyceps sinensis. Chin. Med. 2018, 13, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Q.Y.; Zhu, Z.Y. Chemical structure and mechanism of polysaccharide on Pb2+ tolerance of Cordyceps militaris after Pb2+ domestication. Int. J. Biol. Macromol. 2020, 165, 958–969. [Google Scholar] [CrossRef]
- Qian, G.M.; Pan, G.F.; Guo, J.Y. Anti-inflammatory and antinociceptive effects of cordymin, a peptide purified from the medicinal mushroom Cordyceps sinensis. Nat. Prod. Res. 2012, 26, 2358–2362. [Google Scholar] [CrossRef]
- Wu, D.T.; Meng, L.Z.; Wang, L.Y.; Lv, G.P.; Cheong, K.L.; Hu, D.J.; Guan, J.; Zhao, J.; Li, S.P. Chain conformation and immunomodulatory activity of a hyperbranched polysaccharide from Cordyceps sinensis. Carbohydr. Polym. 2014, 110, 405–414. [Google Scholar] [CrossRef]
- Wang, J.Q.; Kan, L.J.; Nie, S.P.; Chen, H.H.; Cui, S.W.; Phillips, A.O.; Phillips, G.O.; Li, Y.J.; Xie, M.Y. A comparison of chemical composition, bioactive components and antioxidant activity of natural and cultured Cordyceps sinensis. LWT-Food Sci. Technol. 2015, 63, 2–7. [Google Scholar] [CrossRef]
- Ozenver, N.; Boulos, J.C.; Efferth, T. Activity of Cordycepin from Cordyceps sinensis Against Drug-Resistant Tumor Cells as Determined by Gene Expression and Drug Sensitivity Profiling. Nat. Prod. Commun. 2021, 16, 1934578X21993350. [Google Scholar] [CrossRef]
- Xu, F.; Chen, P.; Li, H.; Qiao, S.Y.; Wang, J.X.; Wang, Y.; Wang, X.T.; Wu, B.H.; Liu, H.K.; Wang, C.; et al. Comparative transcriptome analysis reveals the differential response to cadmium stress of two Pleurotus fungi: Pleurotus cornucopiae and Pleurotus ostreatus. J. Hazard. Mater. 2021, 416, 125814. [Google Scholar] [CrossRef]
- Bo, T.; Miao, L.; Cheng, Z.; Qian, Z.; Jia, S.R. Metabolomic Analysis of Antimicrobial Mechanisms of ε-Poly-l-lysine on Saccharomyces cerevisiae. J. Agric. Food Chem. 2014, 62, 4454–4465. [Google Scholar] [CrossRef] [PubMed]
- Traxler, L.; Shrestha, J.; Richter, M.; Krause, K.; Schafer, T.; Kothe, E. Metal adaptation and transport in hyphae of the wood-rot fungus Schizophyllum commune. J. Hazard. Mater. 2022, 425, 127978. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Ying, J.L.; Xu, L.; Wang, Y.; Dong, J.H.; Chen, Y.L.; Tang, M.J.; Li, C.; Muleke, E.M.; Liu, L.W. Genome-wide sRNA and mRNA transcriptomic profiling insights into dynamic regulation of taproot thickening in radish (Raphanus sativus L.). BMC Plant Biol. 2020, 20, 373. [Google Scholar] [CrossRef] [PubMed]
- Sekiguchi, J.; Gaucher, G.M.; Costerton, J.W. Microcycle conidiation in Penicillium urticae: An ultrastructural investigation of conidiogenesis. Can. J. Microbiol. 1975, 21, 2069–2083. [Google Scholar] [CrossRef]
- Anderson, J.G.; Smith, J.E. The Production of Conidiophores and Conidia by Newly Germinated Conidia of Aspergillus niger (Microcycle conidiation). J. Gen. Microbiol. 1971, 69, 185–197. [Google Scholar] [CrossRef] [Green Version]
- Hu, T.; Sun, X.Y.; Zhao, Z.J.; Amombo, E.; Fu, J.M. High temperature damage to fatty acids and carbohydrate metabolism in tall fescue by coupling deep transcriptome and metabolome analysis. Ecotoxicol. Environ. Saf. 2020, 203, 110943. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Y.; Ren, Q.Q.; Lai, Y.H.; Peng, M.Y.; Zhang, J.; Yang, L.T.; Huang, Z.R.; Chen, L.S. Metabolomics combined with physiology and transcriptomics reveals how Citrus grandis leaves cope with copper-toxicity. Ecotoxicol. Environ. Saf. 2021, 223, 112579. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.W.; Liu, X.L.; Zhang, L.L.; Wang, Z.M.; Yuan, Y.; Li, J.; Li, Y.Y.; Huang, H.H.; Cao, X.; Fan, Z.Q. Toxicity mechanism of Nylon microplastics on Microcystis aeruginosa through three pathways: Photosynthesis, oxidative stress and energy metabolism. J. Hazard. Mater. 2022, 426, 128094. [Google Scholar] [CrossRef] [PubMed]
- Mandal, P.K.; Roy, R.G.; Samkaria, A. Oxidative Stress: Glutathione and Its Potential to Protect Methionine-35 of Aβ Peptide from Oxidation. Acs Omega 2022, 7, 27052–27061. [Google Scholar] [CrossRef]
- Farres, M.; Pina, B.; Tauler, R. LC-MS based metabolomics and chemometrics study of the toxic effects of copper on Saccharomyces cerevisiae. Metallomics 2016, 8, 790–798. [Google Scholar] [CrossRef]
- He, R.J.; Wang, Z.; Cui, M.; Liu, S.; Wu, W.; Chen, M.; Wu, Y.C.; Qu, Y.J.; Lin, H.; Chen, S.; et al. HIF1A Alleviates compression-induced apoptosis of nucleus pulposus derived stem cells via upregulating autophagy. Autophagy 2021, 17, 3338–3360. [Google Scholar] [CrossRef]
- Remus, D.; Beuron, F.; Tolun, G.; Griffith, J.D.; Morris, E.P.; Diffley, J.F.X. Concerted Loading of Mcm2-7 Double Hexamers around DNA during DNA Replication Origin Licensing. Cell 2009, 139, 719–730. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.R.; Yang, Y.; Wu, J.R. Activation of DNA damage checkpoints in CHO cells requires a certain level of DNA damage. Biochem. Biophys. Res. Commun. 2001, 287, 775–780. [Google Scholar] [CrossRef]
- Zhu, S.; Li, X.; Dang, B.R.; Wu, F.; Wang, C.M.; Lin, C.J. Lycium Barbarum polysaccharide protects HaCaT cells from PM2.5-induced apoptosis via inhibiting oxidative stress, ER stress and autophagy. Redox Rep. 2022, 27, 32–44. [Google Scholar] [CrossRef]
- Mizushima, N. The role of the Atg1/ULK1 complex in autophagy regulation. Curr Opin Cell Biol. 2010, 22, 132–139. [Google Scholar] [CrossRef]
- Narendrula-Kotha, R.; Theriault, G.; Mehes-Smith, M.; Kalubi, K.; Nkongolo, K. Metal Toxicity and Resistance in Plants and Microorganisms in Terrestrial Ecosystems. Rev Environ Contam Toxicol. 2020, 249, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Krishna, T.P.A.; Maharajan, T.; Roch, G.V.; Ignacimuthu, S.; Ceasar, S.A. Structure, Function, Regulation and Phylogenetic Relationship of ZIP Family Transporters of Plants. Front. Plant Sci. 2020, 11, 662. [Google Scholar] [CrossRef] [PubMed]
- Gustin, J.L.; Loureiro, M.E.; Kim, D.; Na, G.; Tikhonova, M.; Salt, D.E. MTP1-dependent Zn sequestration into shoot vacuoles suggests dual roles in Zn tolerance and accumulation in Zn-hyperaccumulating plants. Plant J. 2009, 57, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Guerrero, M.; Benabdellah, K.; Valderas, A.; Azcon-Aguilar, C.; Ferrol, N. GintABC1 encodes a putative ABC transporter of the MRP subfamily induced by Cu, Cd, and oxidative stress in Glomus intraradices. Mycorrhiza 2010, 20, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Leng, Y.; Li, Y.; Wen, Y.; Zhao, H.; Wang, Q.; Li, S.W. Transcriptome analysis provides molecular evidences for growth and adaptation of plant roots in cadimium-contaminated environments. Ecotoxicol. Environ. Saf. 2020, 204, 111098. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Prasad, M.N.V. Plant-lead interactions: Transport, toxicity, tolerance, and detoxification mechanisms. Ecotoxicol. Environ. Saf. 2018, 166, 401–418. [Google Scholar] [CrossRef]
- Lu, H.Y.; Liu, S.Y.; Zhang, S.L.; Chen, Q.H. Light Irradiation Coupled with Exogenous Metal Ions to Enhance Exopolysaccharide Synthesis from Agaricus sinodeliciosus ZJU-TP-08 in Liquid Fermentation. J. Fungi. 2021, 7, 992. [Google Scholar] [CrossRef]
- Yang, S.L.; Yang, X.; Zhang, H. Extracellular polysaccharide biosynthesis in Cordyceps. Crit. Rev. Microbiol. 2020, 46, 359–380. [Google Scholar] [CrossRef]
- Yuan, Y.; Imtiaz, M.; Rizwan, M.; Dai, Z.H.; Hossain, M.M.; Zhang, Y.H.; Huang, H.L.; Tu, S.X. The role and its transcriptome mechanisms of cell wall polysaccharides in vanadium detoxication of rice. J. Hazard. Mater. 2022, 425, 127966. [Google Scholar] [CrossRef]
- Li, Q.Z.; Chen, J.; Liu, J.Y.; Yu, H.L.; Zhang, L.J.; Song, C.Y.; Li, Y.; Jiang, N.; Tan, Q.; Shang, X.D.; et al. De novo Sequencing and Comparative Transcriptome Analyses Provide First Insights into Polysaccharide Biosynthesis During Fruiting Body Development of Lentinula edodes. Front. Microbiol. 2021, 12, 627099. [Google Scholar] [CrossRef]
- Xu, Y.; Du, H.M.; Huang, B.R. Identification of Metabolites Associated with Superior Heat Tolerance in Thermal Bentgrass through Metabolic Profiling. Crop. Sci. 2013, 53, 1626–1635. [Google Scholar] [CrossRef]
- Hausler, R.E.; Ludewig, F.; Krueger, S. Amino acids—A life between metabolism and signaling. Plant Sci. 2014, 229, 225–237. [Google Scholar] [CrossRef]
- Jung, B.; Kim, S.; Lee, J. Microcyle Conidiation in Filamentous Fungi. Mycobiology 2014, 42, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Hanlin, R.T.J.M. Microcycle conidiation—A review. J. Gen. Microbiol. 1994, 35, 113–123. [Google Scholar] [CrossRef]
- Bosch, A.; Yantorno, O.J. Microcycle conidiation in the entomopathogenic fungus Beauveria bassiana bals. (vuill.). Process Biochem. 1999, 34, 707–716. [Google Scholar] [CrossRef]
- Liu, J.; Cao, Y.; Xia, Y. Mmc, a gene involved in microcycle conidiation of the entomopathogenic fungus Metarhizium anisopliae. J. Invertebr. Pathol. 2010, 105, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Maheshwari, R. Microcycle conidiation and its genetic basis in Neurospora crassa. J. Gen. Microbiol. 1991, 137, 2103–2115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrestha, B.; Han, S.-K.; Yoon, K.-S.; Sung, J.-M. Morphological Characteristics of Conidiogenesis in Cordyceps militaris. Mycobiology 2005, 33, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Park, H.S.; Yu, J.H. Genetic control of asexual sporulation in filamentous fungi. Curr. Opin. Microbiol. 2012, 15, 669–677. [Google Scholar] [CrossRef]
- de Vries, R.P.; Riley, R.; Wiebenga, A.; Aguilar-Osorio, G.; Amillis, S.; Uchima, C.A.; Anderluh, G.; Asadollahi, M.; Askin, M.; Barry, K.; et al. Comparative genomics reveals high biological diversity and specific adaptations in the industrially and medically important fungal genus Aspergillus. Genome Biol. 2017, 18, 28. [Google Scholar] [CrossRef] [Green Version]
- Wendland, J.; Dunkler, A.; Walther, A. Characterization of alpha-factor pheromone and pheromone receptor genes of Ashbya gossypii. FEMS Yeast Res. 2011, 11, 418–429. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.Y.; Wang, F.; Lano, N.; Liu, B.; Hu, C.C.; Xue, W.; Zhang, Z.Y.; Li, S.J. The Zn(II)2Cys6-Type Transcription Factor ADA-6 Regulates Conidiation, Sexual Development, and Oxidative Stress Response in Neurospora crassa. Front. Microbiol. 2019, 10, 750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, Y.E.; Cho, H.J.; Lee, M.K.; Park, H.S. Characterizing the role of Zn cluster family transcription factor ZcfA in governing development in two Aspergillus species. PLoS ONE 2020, 15, e0228643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.Y.; Mead, M.E.; Lee, M.K.; Neuhaus, G.F.; Adpressa, D.A.; Martien, J.I.; Son, Y.E.; Moon, H.; Amador-Noguez, D.; Han, K.H.; et al. Transcriptomic, Protein-DNA Interaction, and Metabolomic Studies of VosA, VelB, and WetA in Aspergillus nidulans Asexual Spores. mBio 2021, 12, e03128-20. [Google Scholar] [CrossRef]
- Bayram, O.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.J.; Keller, N.P.; Yu, J.H.; et al. VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Ni, M.; Jeong, K.C.; Kim, Y.H.; Yu, J.H. The Role, Interaction and Regulation of the Velvet Regulator VelB in Aspergillus nidulans. PLoS ONE 2012, 7, 45935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsitsigiannis, D.I.; Kowieski, T.M.; Zarnowski, R.; Keller, N.P. Endogenous lipogenic regulators of spore balance in Aspergillus nidulans. Eukaryot Cell 2004, 3, 1398–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalid, A.R.; Lv, X.; Naeem, M.; Mehmood, K.; Shaheen, H.; Dong, P.; Qiu, D.; Ren, M. Autophagy Related Gene (ATG3) is a Key Regulator for Cell Growth, Development, and Virulence of Fusarium oxysporum. Genes 2019, 10, 658. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.Y.; Wang, J.J.; Feng, M.G.; Ying, S.H. Autophagy-related gene ATG7 participates in the asexual development, stress response and virulence of filamentous insect pathogenic fungus Beauveria bassiana. Curr. Genet. 2019, 65, 1015–1024. [Google Scholar] [CrossRef]
- Deniaud, E.; Le Gall, L.; Rusig, A.M.; Lahaye, M. Initial observations on glycoside deposition in cell walls of Palmaria palmata (L.) Kuntze (Rhodophyta) during spore germination. Bot. Mar. 2006, 49, 266–269. [Google Scholar] [CrossRef]
- Zhang, S.Z.; Xia, Y.X.; Keyhani, N.O. Contribution of the gas1 Gene of the Entomopathogenic Fungus Beauveria bassiana, Encoding a Putative Glycosylphosphatidylinositol-Anchored beta-1,3-Glucanosyltransferase, to Conidial Thermotolerance and Virulence. Appl. Environ. Microbiol. 2011, 77, 2676–2684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, W.; Yamashita, S.; Ohta, A.; Horiuchi, H. Functional Differentiation of Chitin Synthases in Yarrowia lipolytica. Biosci. Biotechnol. Biochem. 2013, 77, 1275–1281. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mycelia Biomass (g/L) | IPS (mg/g) | EPS (mg/L) | Conidia (g/L) | Zn2+ Concentration (mg/g) | |||
|---|---|---|---|---|---|---|---|
| Mycelia | EPS | Conidia | |||||
| Zn2+ stress | 6 ± 1.4 a | 135 ± 5.3 a | 233 ± 14.8 a | 0.162 ± 0.0062 | 11.9 ± 1.2 a | 13.2 ± 1.6 a | 6.2 ± 0.9 |
| Control | 13 ± 1.8 b | 119 ± 6.6 a | 181 ± 13.5 b | ND | 0.1 ± 0.004 b | 0.095 ± 0.003 b | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Tong, L.-L.; Yuan, L.; Liu, M.-Z.; Du, Y.-H.; Yang, L.-H.; Ren, B.; Guo, D.-S. Integration of Physiological, Transcriptomic and Metabolomic Reveals Molecular Mechanism of Paraisaria dubia Response to Zn2+ Stress. J. Fungi 2023, 9, 693. https://doi.org/10.3390/jof9070693
Wang Y, Tong L-L, Yuan L, Liu M-Z, Du Y-H, Yang L-H, Ren B, Guo D-S. Integration of Physiological, Transcriptomic and Metabolomic Reveals Molecular Mechanism of Paraisaria dubia Response to Zn2+ Stress. Journal of Fungi. 2023; 9(7):693. https://doi.org/10.3390/jof9070693
Chicago/Turabian StyleWang, Yue, Ling-Ling Tong, Li Yuan, Meng-Zhen Liu, Yuan-Hang Du, Lin-Hui Yang, Bo Ren, and Dong-Sheng Guo. 2023. "Integration of Physiological, Transcriptomic and Metabolomic Reveals Molecular Mechanism of Paraisaria dubia Response to Zn2+ Stress" Journal of Fungi 9, no. 7: 693. https://doi.org/10.3390/jof9070693