
Taxonomic and Phylogenetic Studies of Saprobic Fungi Associated with Mangifera indica in Yunnan, China

 ,
,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation
2.2. DNA Extraction, PCR and Sequencing
2.3. Phylogenetic Analyses
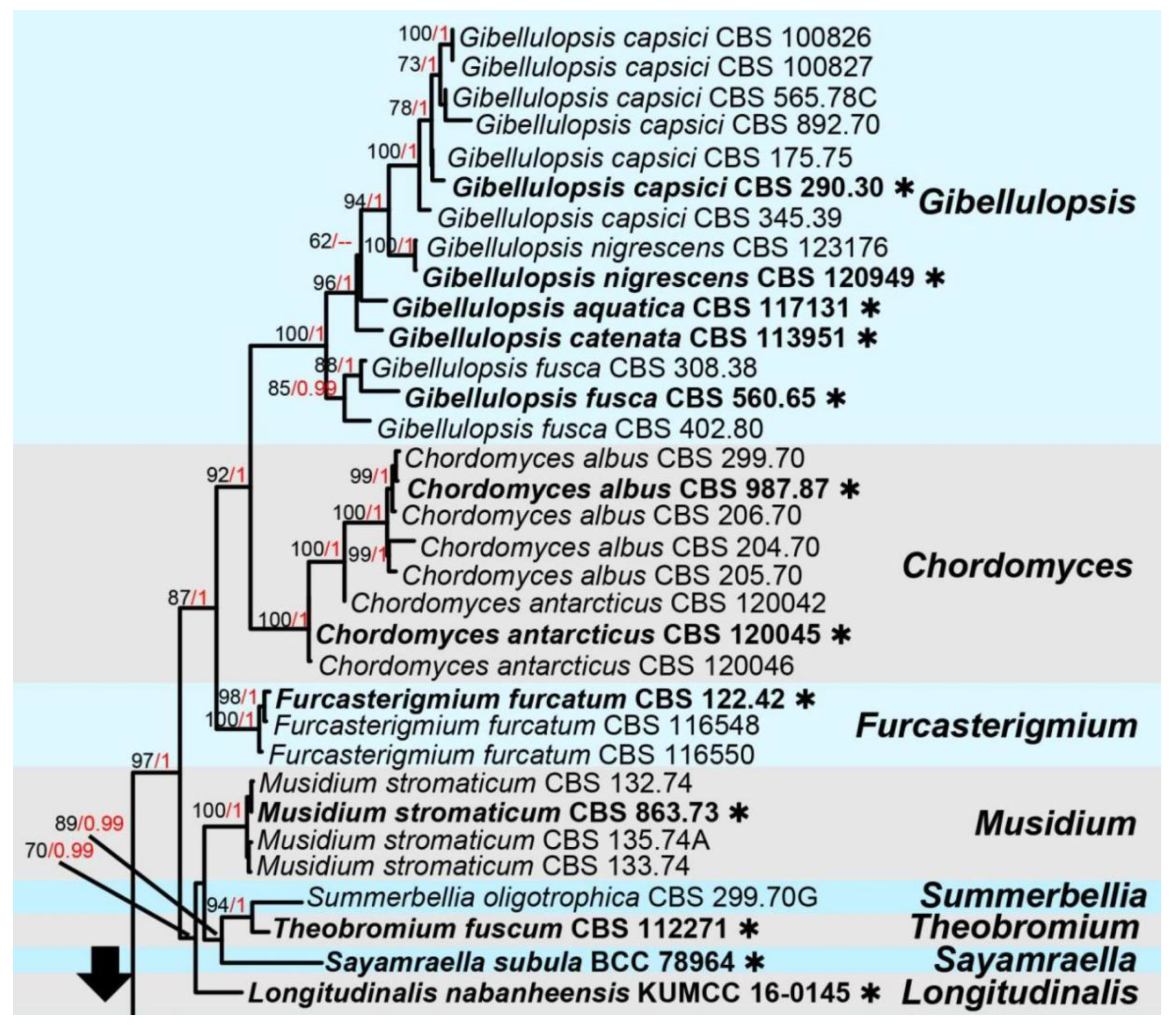
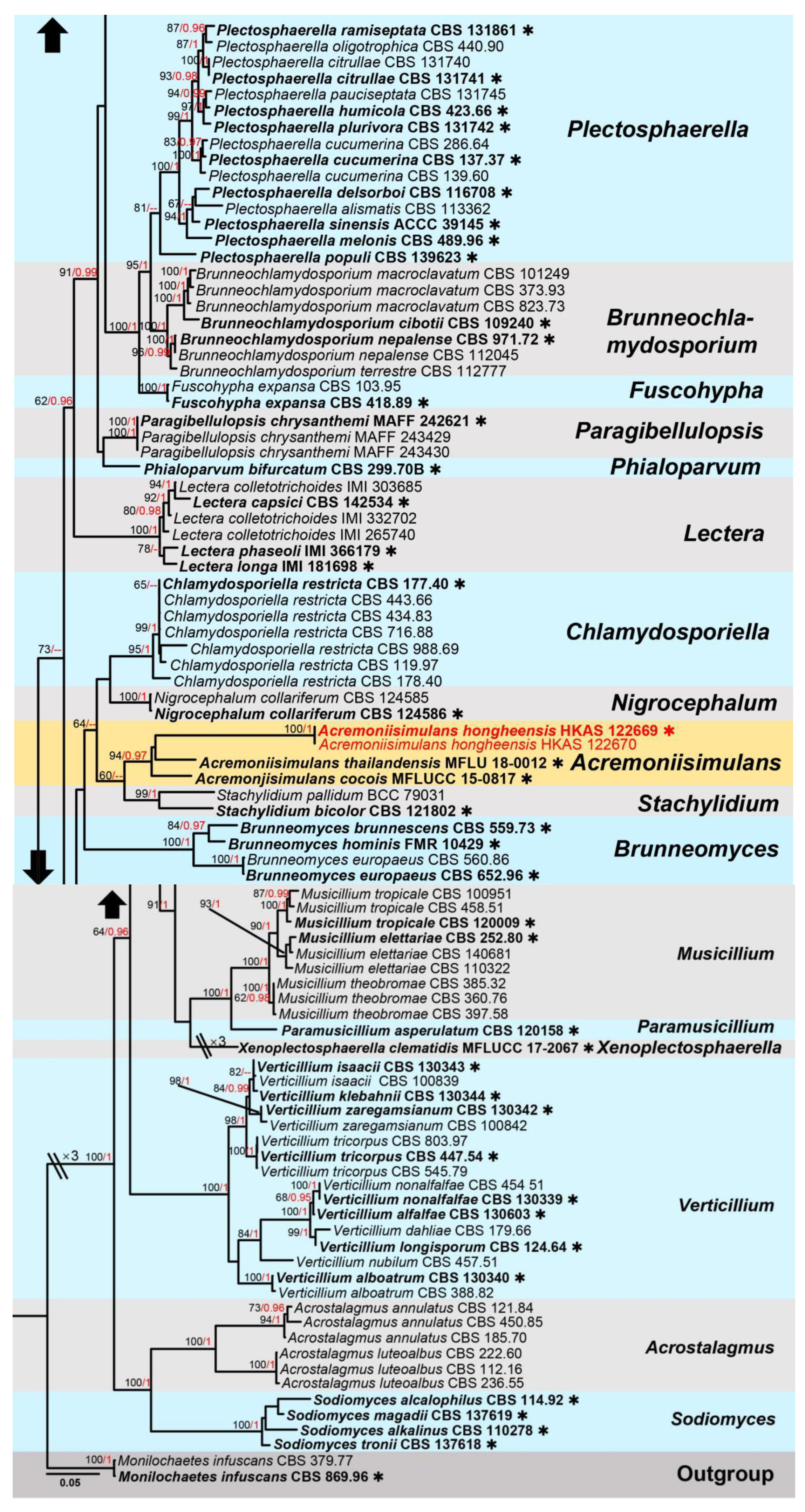
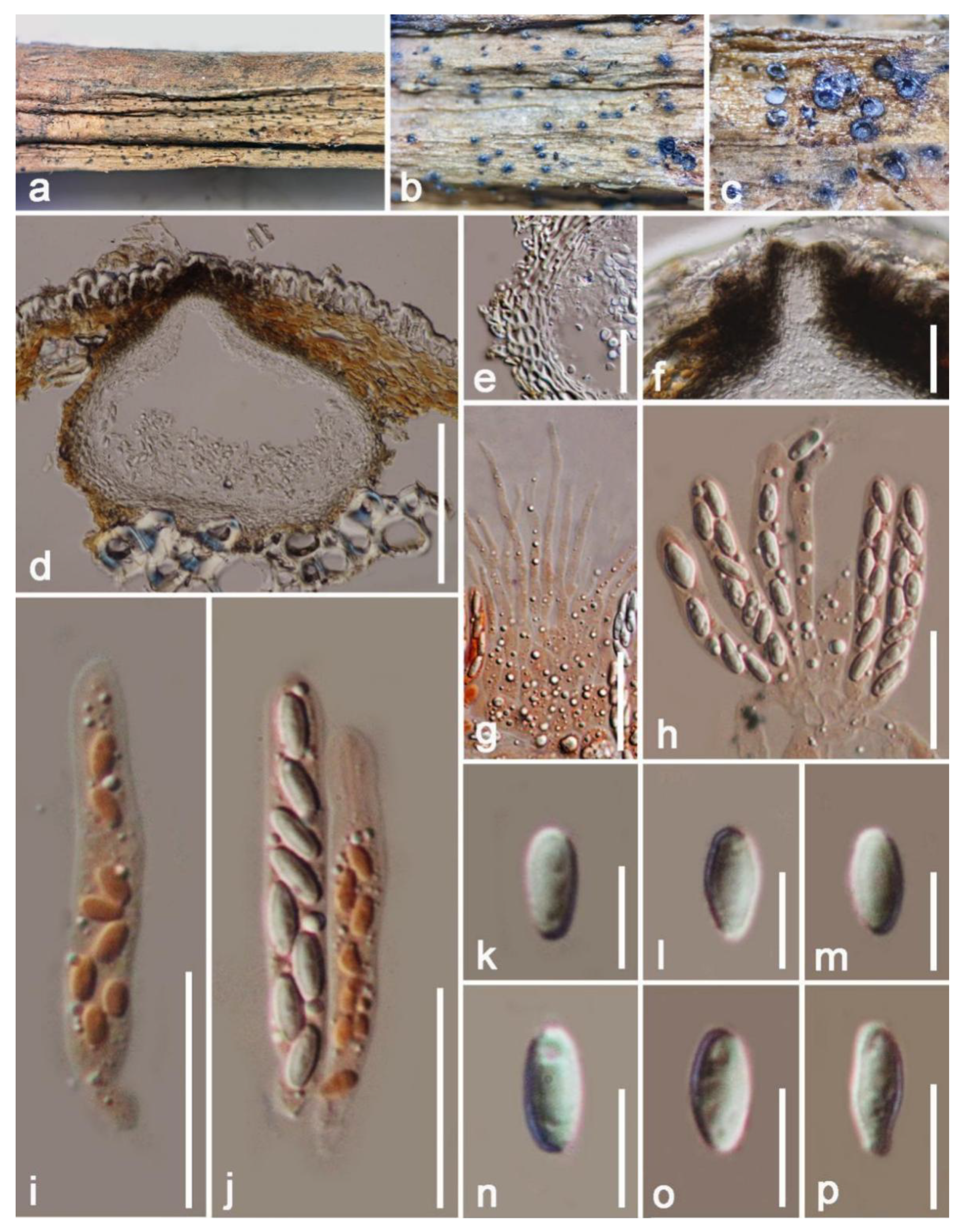
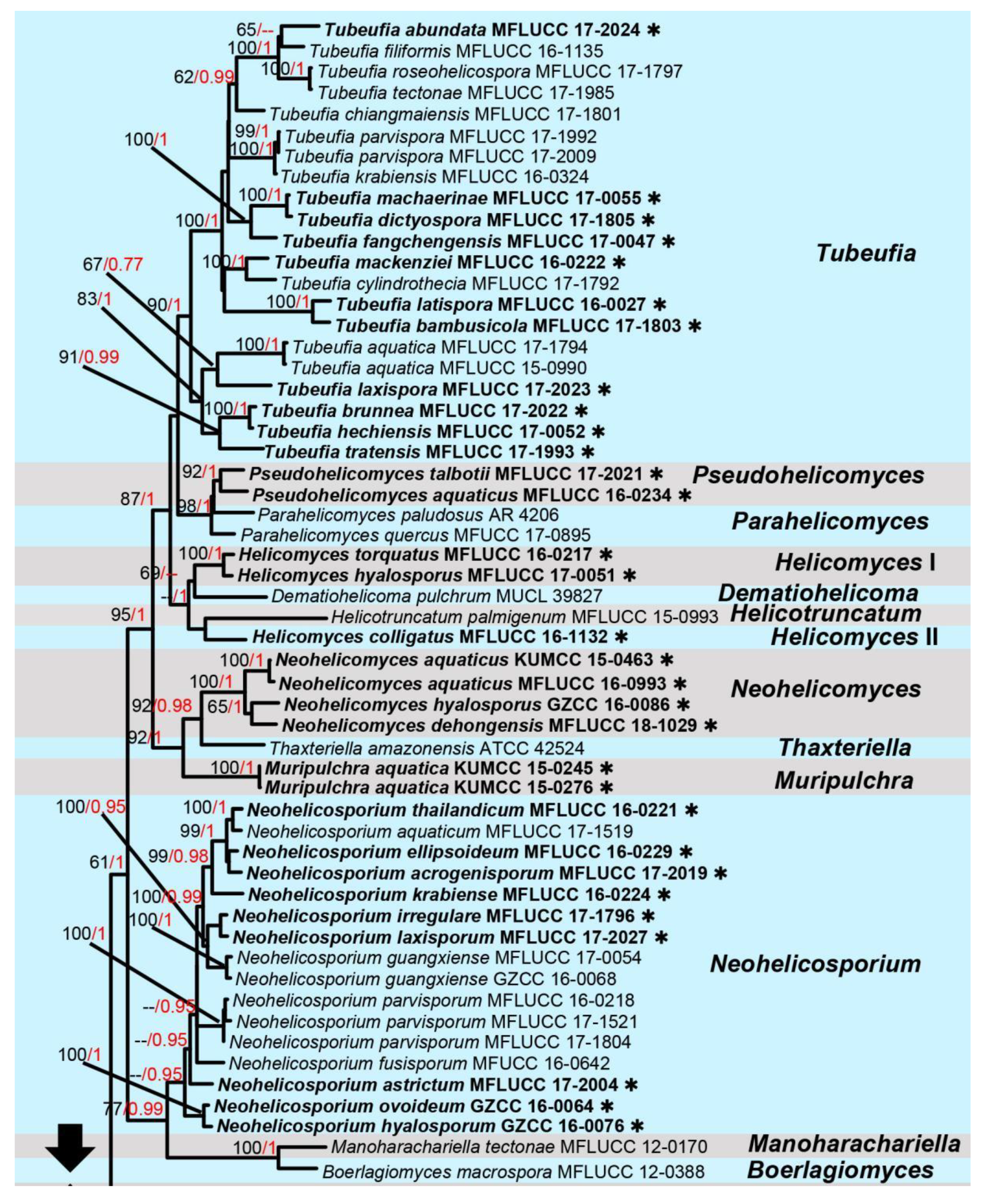
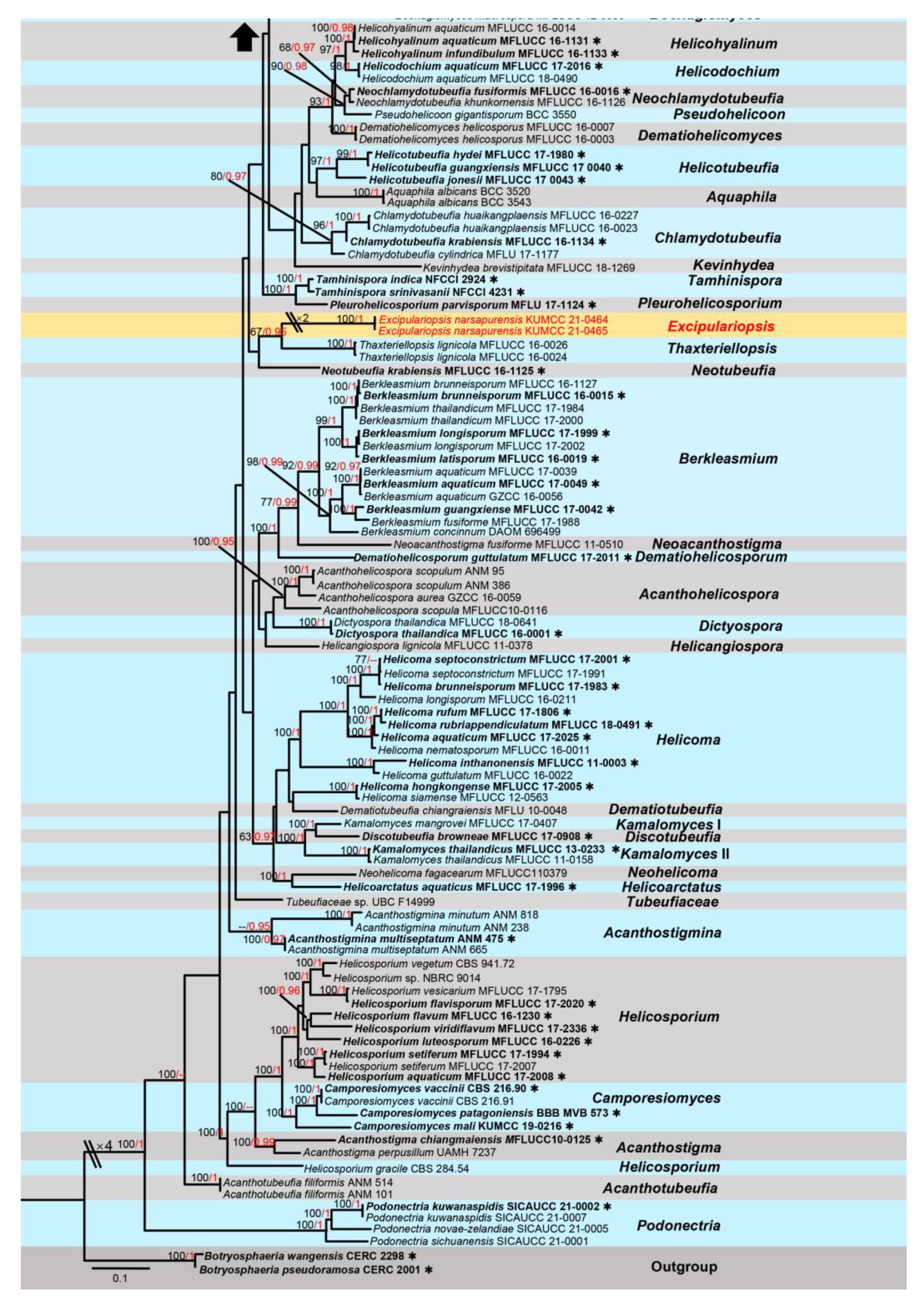
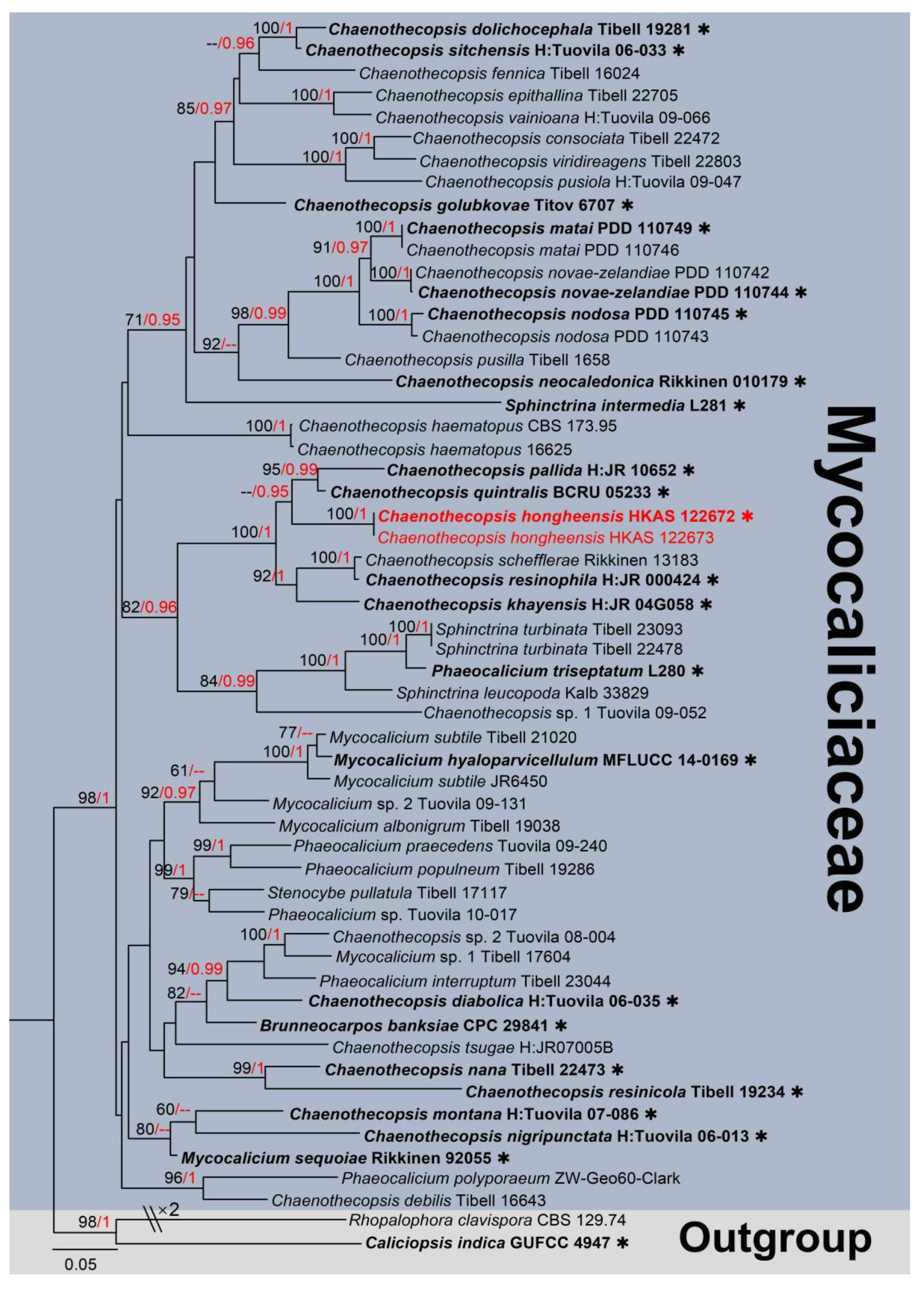
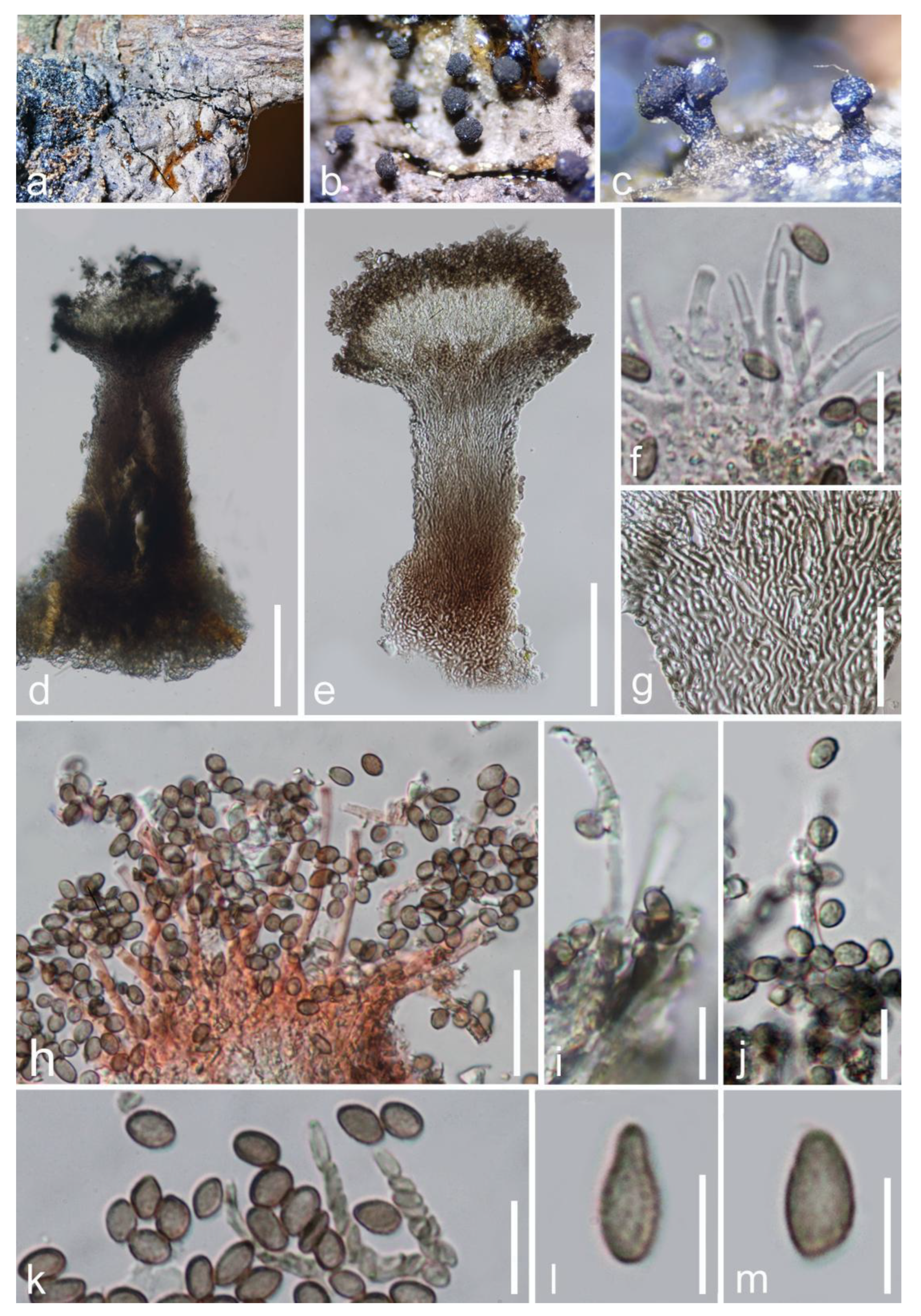
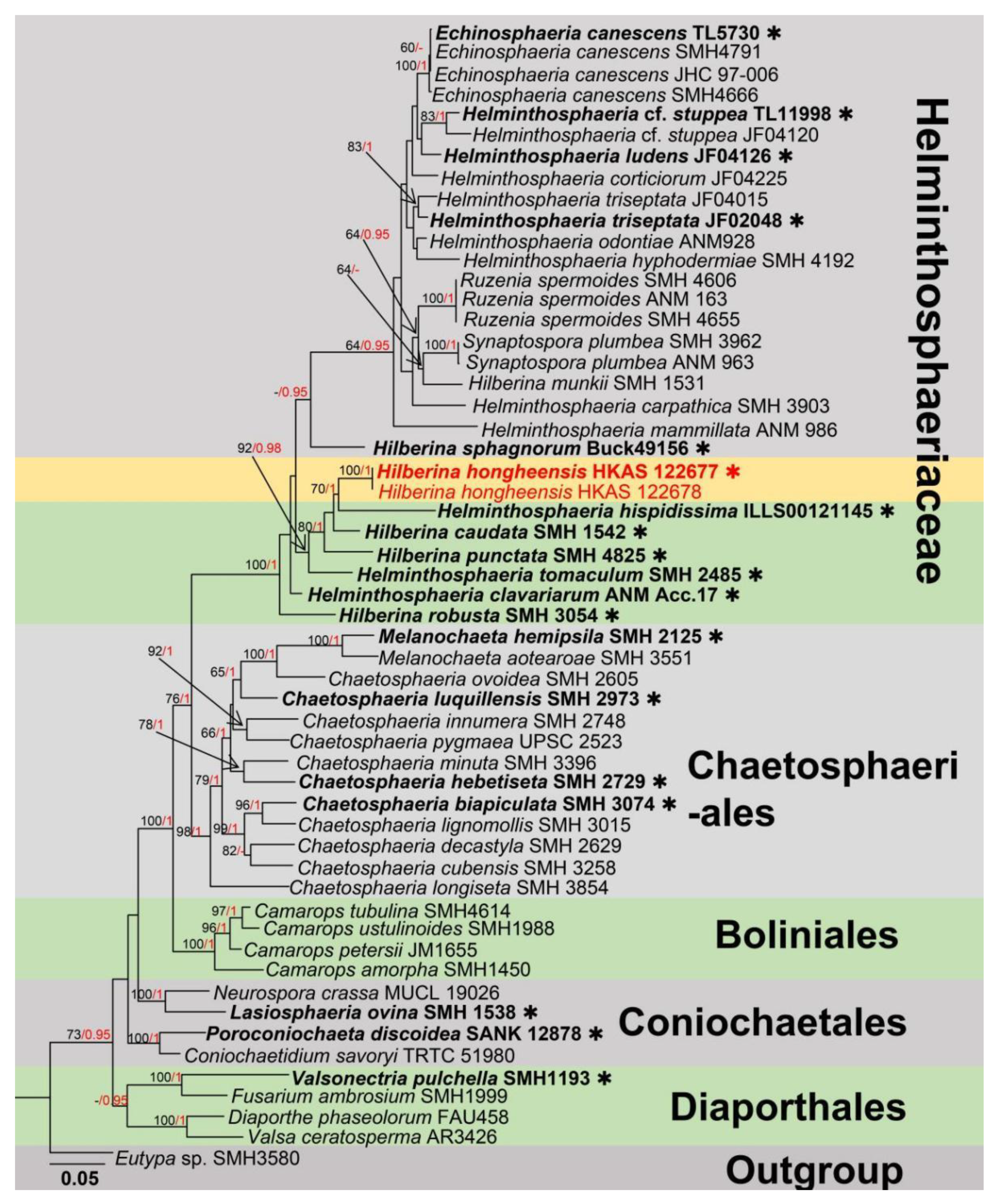
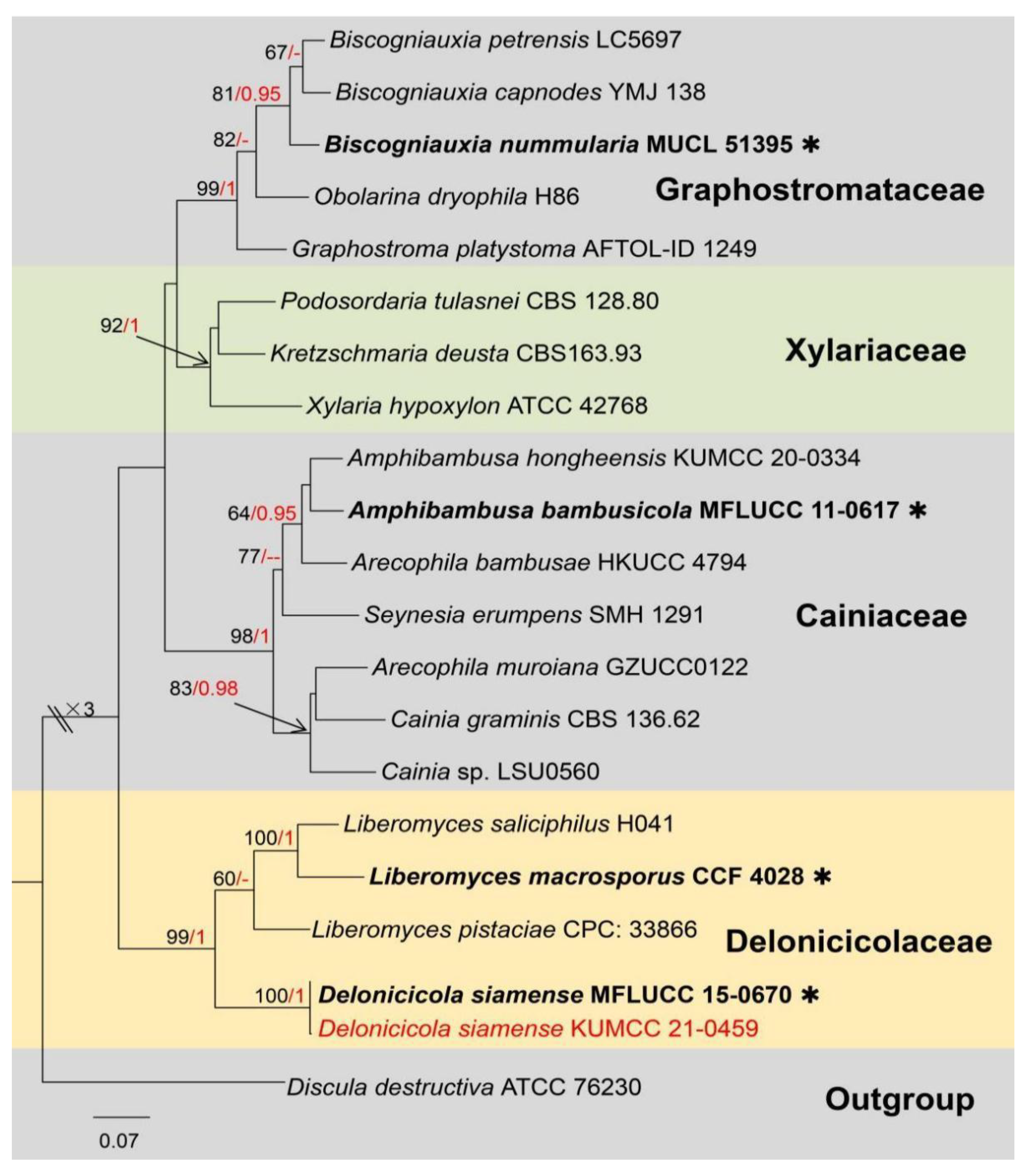
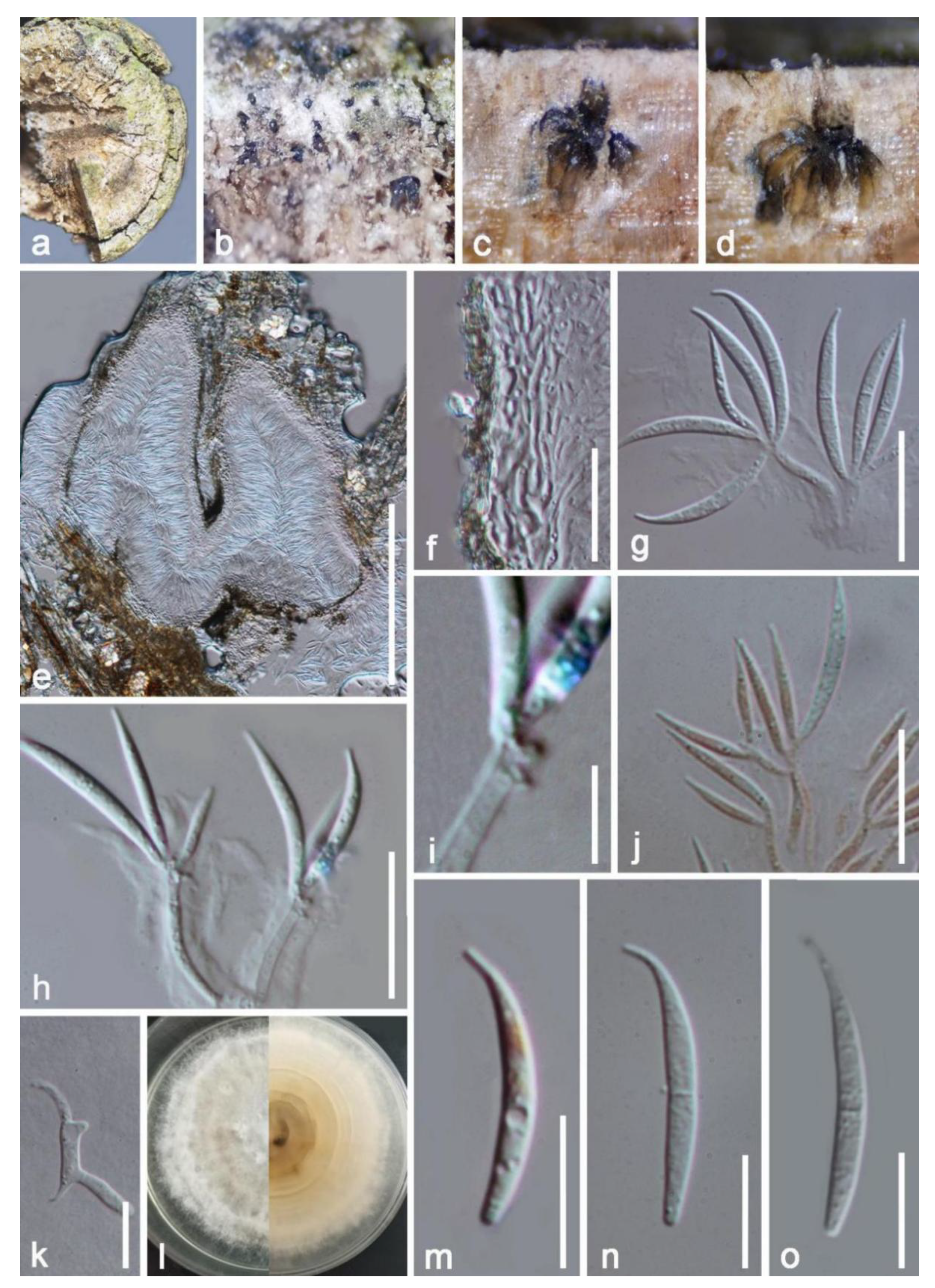
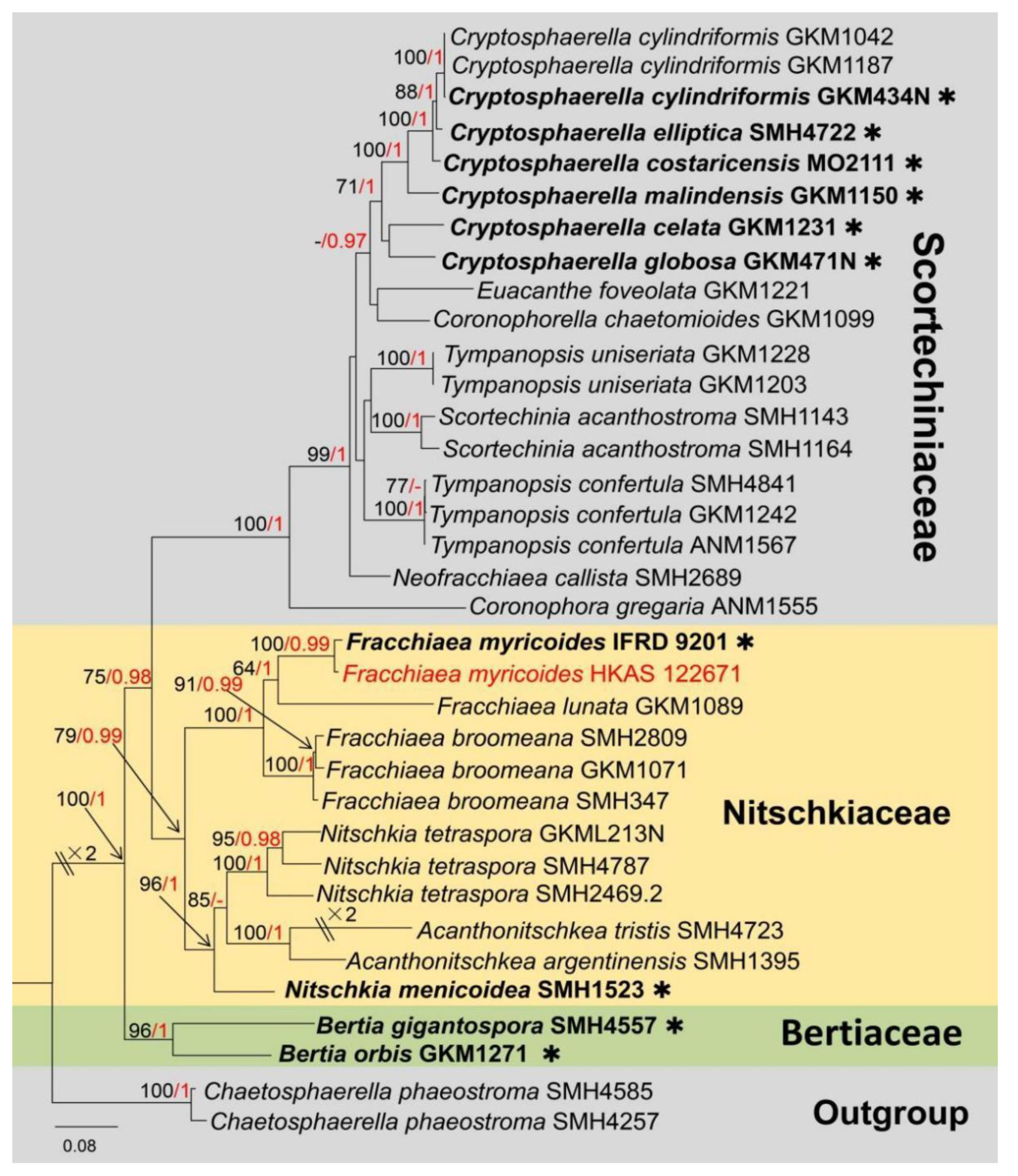
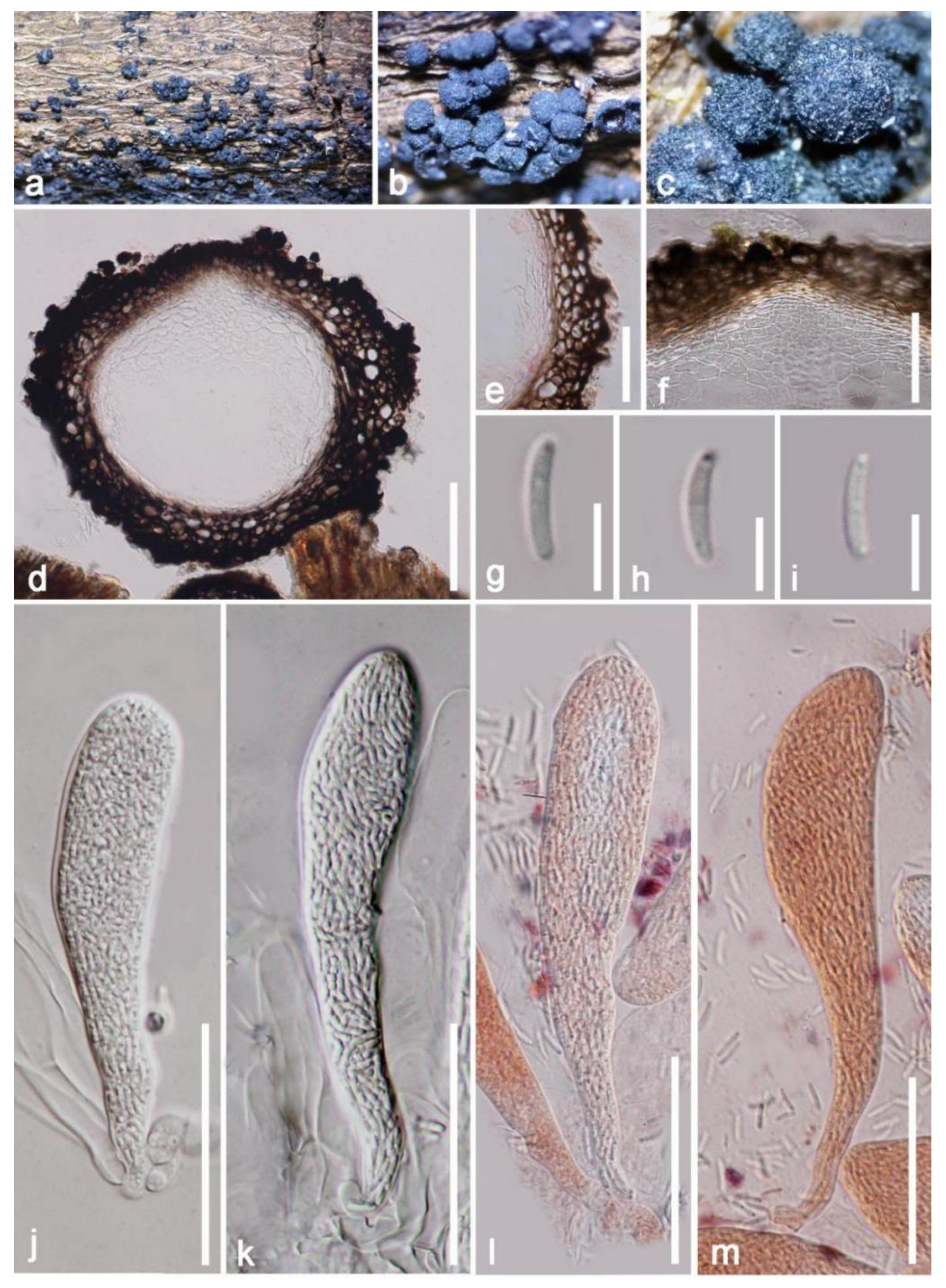
3. Results
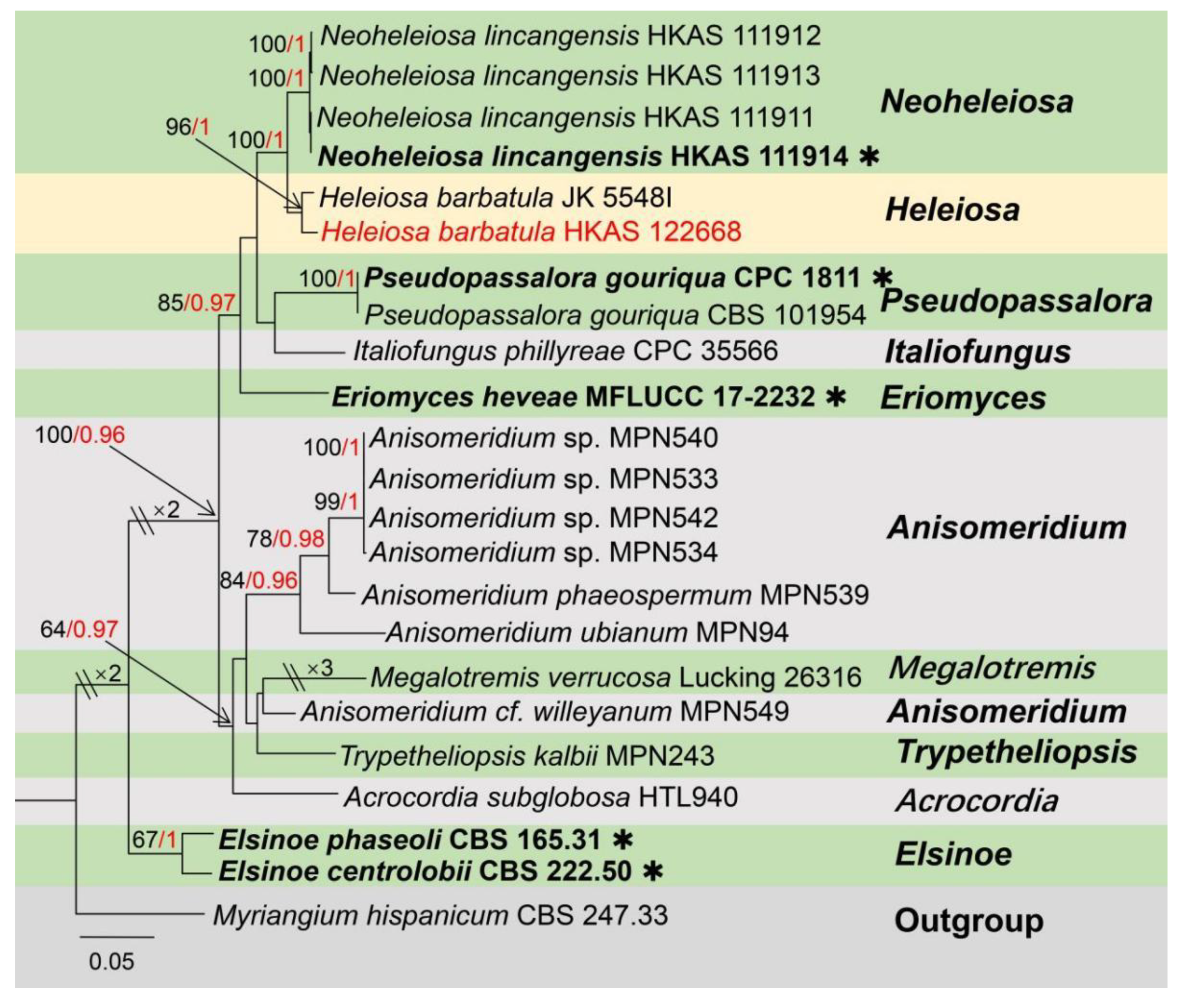
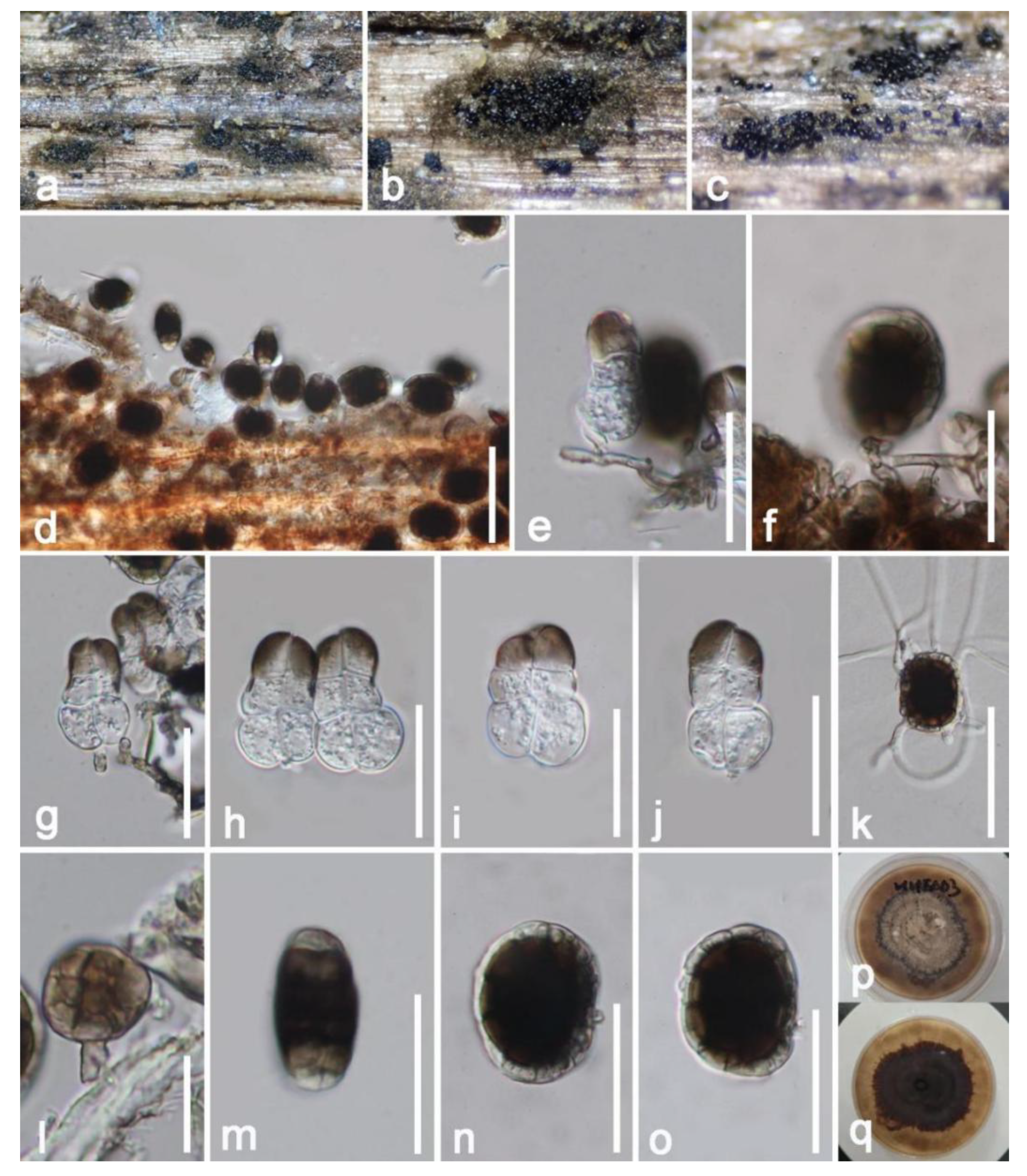
Taxonomic and Phylogenetic Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hyde, K.D.; Xu, J.C.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.S.; Brahamanage, R.S.; Stadler, M.; et al. The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Divers. 2019, 97, 1–136. [Google Scholar] [CrossRef] [Green Version]
- Phukhamsakda, C.; Nilsson, R.H.; Bhunjun, C.S.; de Farias, A.R.G.; Sun, Y.R.; Wijesinghe, S.N.; Raza, M.; Bao, D.F.; Lu, L.; Tibpromma, S.; et al. The numbers of fungi: Contributions from traditional taxonomic studies and challenges of metabarcoding. Fungal Divers. 2022, 114, 327–386. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Lücking, R. Fungal diversity revisited: 2.2 to 3.8 million species. Microbiol. Spectr. 2017, 5, 79–95. [Google Scholar] [CrossRef]
- Lücking, R.; Aime, M.C.; Robbertse, B.; Mille, A.N.; Aoki, T.; Ariyawansa, H.A.; Cardinali, G.; Crous, P.W.; Druzhinina, I.S.; Geiser, D.M.; et al. Fungal taxonomy and sequence-based nomenclature. Nat. Microbiol. 2021, 6, 540–548. [Google Scholar] [CrossRef]
- Baldrian, P.; Větrovský, T.; Lepinay, C.; Kohout, P. High-throughput sequencing view on the magnitude of global fungal diversity. Fungal Divers. 2021, 19, 1–9. [Google Scholar] [CrossRef]
- Douthett, B.D.G. The mango: Asia’s king of fruits. Ethnobot. Leafl. 2011, 2000, 4. [Google Scholar]
- Yadav, D.; Singh, S.P. Mango: History origin and distribution. J. Pharmacogn. Phytochem. 2017, 6, 1257–1262. [Google Scholar]
- Lauricella, M.; Emanuele, S.; Calvaruso, G.; Giuliano, M.; D’Anneo, A. Multifaceted health benefits of Mangifera indica L. (Mango): The inestimable value of orchards recently planted in Sicilian rural areas. Nutrients 2017, 9, 525. [Google Scholar] [CrossRef]
- Gao, A.; Chen, Y.; Luo, R.; Huang, J.; Zhao, Z.; Wang, W.; Wang, Y.; Dang, Z. Development status of Chinese mango industry in 2018. Adv. Agric. Hortic. Entomol. 2019, 1, 21–60. [Google Scholar]
- Zhang, C.X.; Xie, D.H.; Chen, Y.F.; Bai, T.Q.; Ni, Z.G. The development status of Yunnan mango industry. China Fruits 2020, 6, 112–117. (In Chinese) [Google Scholar]
- Hyde, K.D.; Bussaban, B.; Paulus, B.; Crous, P.W.; Lee, S.; Mckenzie, E.H.; Photita, W.; Lumyong, S. Diversity of saprobic microfungi. Biodivers. Conserv. 2007, 16, 7–35. [Google Scholar] [CrossRef]
- Taïbi, A.; Diop, A.; Leneveu-Jenvrin, C.; Broussolle, V.; Lortal, S.; Méot, J.M.; Soria, C.; Chillet, M.; Lechaudel, M.; Meile, J.C.; et al. Dynamics of bacterial and fungal communities of mango: From the tree to ready-to-Eat products. Food Microbiol. 2022, 108, 104095. [Google Scholar] [CrossRef] [PubMed]
- Tennakoon, D.S.; Kuo, C.H.; Jeewon, R.; Thambugala, K.M.; Hyde, K.D. Saprobic Lophiostomataceae (Dothideomycetes): Pseudolophiostoma mangiferae sp. nov. and Neovaginatisporafuckelii, a new record from Mangifera indica. Phytotaxa 2018, 364, 157–171. [Google Scholar] [CrossRef]
- Guo, Z.; Yu, Z.; Li, Q.; Tang, L.; Guo, T.; Hsiang, T.; Huang, S.; Mo, J.; Luo, S. Fusarium species associated with leaf spots of mango in China. Microb. Pathog. 2021, 150, 104736. [Google Scholar] [CrossRef] [PubMed]
- Tamakaew, N.; Karunarathna, A.; Pakdeeniti, P.; Withee, P.; Senwanna, C.; Haituk, S.; Cheewangkoon, R. New species of Cercosporoid on Mango in Thailand. Chiang Mai J. Sci. 2022, 49, 684–1692. [Google Scholar] [CrossRef]
- Yang, E.F.; Tibpromma, S.; Karunarathna, S.C.; Phookamsak, R.; Xu, J.C.; Zhao, Z.X.; Karunanayake, C.; Promputtha, I. Taxonomy and phylogeny of novel and extant taxa in Pleosporales associated with Mangifera indica from Yunnan, China (Series I). J. Fungi 2022, 8, 152. [Google Scholar] [CrossRef]
- Yang, E.F.; Karunarathna, S.C.; Dai, D.Q.; Stephenson, S.L.; Elgorban, A.M.; Al-Rejaie, S.; Promputtha, I.; Tibpromma, S. Taxonomy and phylogeny of fungi associated with Mangifera indica from Yunnan, China. J. Fungi 2022, 8, 1249. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Rathnayaka, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B.; Hurdeal, V.G.; Pem, D.; Dissanayake, L.S.; Wijesinghe, S.N.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Wang, F.; Wang, K.; Cai, L.; Zhao, M.; Kirk, P.M.; Fan, G.; Sun, Q.; Li, B.; Wang, S.; Yu, Z.; et al. Fungal Names. 2023. Available online: https://nmdc.cn/fungalnames/ (accessed on 3 February 2023).
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and tef1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Glez-Peña, D.; Gómez-Blanco, D.; Reboiro-Jato, M.; Fdez-Riverola, F.; Posada, D. FALTER: Program oriented conversion of DNA and protein alignments. Nucleic Acids Res. 2010, 38, 14–18. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES science gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erixon, P.; Svennblad, B.; Britton, T.; Oxelman, B. Reliability of Bayesian posterior probabilities and bootstrap frequencies in phylogenetics. Syst. Biol. 2003, 52, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Zhaxybayeva, O.; Gogarten, J.P. Bootstrap, Bayesian probability and maximum likelihood mapping: Exploring new tools for comparative genome analysis. BMC Genom. 2002, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.; Wilgenbusch, J.C.; Warren, D.L.; Swofford, D.L. AWTY (Are we there yet?): A system for graphical exploration of MCMC convergence in Bayesian phylogenetics. Bioinformatics 2008, 24, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree v1. 4.0. A Graphical Viewer of Phylogenetic Trees. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 3 January 2023).
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.C.; Bhat, D.J.; Phillips, A.J.L.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.S.; Dissanayake, A.J.; Tennakoon, D.S.; et al. Fungal diversity notes 840–928: Micro-fungi associated with Pandanaceae. Fungal Divers. 2018, 93, 1–160. [Google Scholar] [CrossRef]
- Konta, S.; Tibpromma, S.; Karunarathna, S.C.; Samarakoon, M.C.; Steven, L.S.; Mapook, A.; Boonmee, S.; Senwanna, C.; Balasuriya, A.; Eungwanichayapant, P.D.; et al. Morphology and multigene phylogeny reveal ten novel taxa in Ascomycota from terrestrial palm substrates (Arecaceae) in Thailand. Mycosphere 2023, 14, 107–152. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Saxena, R.K.; Erdoğdu, M.; Selçuk, F.; Rajeshkumar, K.C. Outline of Fungi and fungus-like taxa–2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Spooner, B.M.; Kirk, P.M. Taxonomic notes on Excipularia and Scolicosporium. Trans. Br. Mycol. Soc. 1982, 78, 247–257. [Google Scholar] [CrossRef]
- Pem, D.; Jeewon, R.; Bhat, D.J.; Doilom, M.; Boonmee, S.; Hongsanan, S.; Promputtha, I.; Xu, J.C.; Hyde, K.D. Mycosphere notes 275–324: A morpho-taxonomic revision and typification of obscure Dothideomycetes genera (incertaesedis). Mycosphere 2019, 10, 1115–1246. [Google Scholar] [CrossRef]
- Dubey, R.; Pandey, A.D. Statistical analysis of foliicolous fungal biodiversity of Konkan region, Maharashtra, India: A novel approach. Plant Pathol. Quar. 2019, 9, 77–115. [Google Scholar] [CrossRef]
- Rajeshkumar, K.C.; Bhat, D.J.; Lad, S.S.; Wijayawardene, N.W.; Singh, S.K.; Pandkar, M.R.; Maurya, D.K.; Ashtekar, N.D.; Hyde, K.D. Morphology and phylogeny of Tamhinispora srinivasanii sp. nov. (Tubeufiaceae) from northern Western Ghats, India. Phytotaxa 2018, 346, 113–120. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Tibpromma, S.; Wanasinghe, D.N.; Thambugala, K.M.; Tian, Q.; Wang, Y.; Fu, L. Towards incorporating asexual fungi in a natural classification: Checklist and notes 2012–2016. Mycosphere 2017, 8, 1457–1555. [Google Scholar] [CrossRef]
- Tibell, L.; Wedin, M. Mycocaliciales, a new order for nonlichenized calicioid fungi. Mycologia 2000, 92, 577–581. [Google Scholar] [CrossRef]
- Kirk, P.M. Index Fungorum. Available online: http://www.indexfungorum.org/names/names.asp (accessed on 3 February 2023).
- Tibell, L. Calicioid lichens and fungi. Nord. Lichen Flora 1999, 1, 2–94. [Google Scholar]
- Tuovila, H.; Schmidt, A.R.; Beimforde, C.; Dörfelt, H.; Grabenhorst, H.; Rikkinen, J. Stuck in time—A new Chaenothecopsis species with proliferating ascomata from Cunninghamia resin and its fossil ancestors in European amber. Fungal Divers. 2013, 58, 199–213. [Google Scholar] [CrossRef] [Green Version]
- Sukhomlyn, M.M.; Heluta, V.P.; Perkovsky, E.E.; Ignatov, M.S.; Vasilenko, D.V. First record of fungus of the family Mycocaliciaceae in rovno amber (Ukraine). Paleontol. J. 2021, 55, 684–690. [Google Scholar] [CrossRef]
- Tuovila, H.; Larsson, P.; Rikkinen, J. Three resinicolous north American species of Mycocaliciales in Europe with a re-evaluation of Chaenothecopsis oregana Rikkinen. Karstenia 2011, 51, 37–49. [Google Scholar] [CrossRef]
- Tuovila, H.; Davey, M.L.; Yan, L.; Huhtinen, S.; Rikkinen, J. New resinicolous Chaenothecopsis species from China. Mycologia 2014, 106, 989–1003. [Google Scholar] [CrossRef] [PubMed]
- Temu, S.G.; Tibell, S.; Tibuhwa, D.D.; Tibell, L. Crustose calicioid lichens and fungi in mountain cloud forests of Tanzania. Microorganisms 2019, 7, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beimforde, C.; Schmidt, A.R.; Tuovila, H.; Kaulfuss, U.; Germer, J.; Lee, W.G.; Rikkinen, J. Chaenothecopsis (Mycocaliciales, Ascomycota) from exudates of endemic New Zealand Podocarpaceae. MycoKeys 2023, 95, 101–129. [Google Scholar] [CrossRef] [PubMed]
- Rikkinen, J.; Tuovila, H.; Beimforde, C.; Seyfullah, L.; Perrichot, V.; Schmidt, A.R. Chaenothecopsis neocaledonica sp. nov.: The first resinicolous fungus from an araucarian conifer. Phytotaxa 2014, 173, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Fuckel, L. Symbolae Mycologicae. Beiträge zur Kenntniss der Rheinischen Pilze; Jahrbücher des Nassauischen Vereins für Naturkunde; Wiesbaden, J. Niedner: London, UK, 1870; pp. 1–459. [Google Scholar]
- Miller, A.N.; Huhndorf, S.M. A natural classification of Lasiosphaeria based on nuclear LSU rDNA sequences. Mycol. Res. 2004, 108, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.N.; Huhndorf, S.M.; Fournier, J. Phylogenetic relationships of five uncommon species of Lasiosphaeria and three new species in the Helminthosphaeriaceae (Sordariomycetes). Mycologia 2014, 106, 505–524. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Restrepo, M.; Giraldo, A.; van Doorn, R.; Wingfield, M.J.; Groenewald, J.Z.; Barreto, R.W.; Colmán, A.A.; Mansur, P.S.C.; Crous, P.W. The genera of fungi–G6: Arthrographis, Kramasamuha, Melnikomyces, Thysanorea, and Verruconis. Fungal Syst. Evol. 2020, 6, 1–24. [Google Scholar] [CrossRef]
- Kirk, P.M. Species Fungorum. Available online: https://www.speciesfungorum.org/ (accessed on 3 January 2023).
- Luo, J.; Yin, J.F.; Cai, L.; Zhang, K.Q.; Hyde, K.D. Freshwater fungi in Lake Dianchi, a heavily polluted lake in Yunnan, China. Fungal Divers. 2004, 16, 93–112. [Google Scholar]
- Carneirod, A.D.A.; Gusmão, L.F.P.; Miller, A.N. Brazilian semi-arid ascomycetes III: New records of Dothideomycetes and Sordariomycetes. Nova Hedwig. 2017, 105, 487–494. [Google Scholar] [CrossRef]
- Perera, R.H.; Maharachchikumbura, S.S.N.; Jones, E.B.G.; Bahkali, A.H.; Elgorban, A.M.; Liu, J.K.; Liu, Z.Y.; Hyde, K.D. Delonicicola siamense gen. & sp. nov. (Delonicicolaceae fam. nov., Delonicicolales ord. nov.), a saprobic species from Delonix regia seed pods. Cryptogam. Mycol. 2017, 38, 321–340. [Google Scholar] [CrossRef]
- Pažoutová, S.; Šrůtka, P.; Holuša, J.; Chudíčková, M.; Kubátová, A.; Kolařík, M. Liberomyces gen. nov. with two new species of endophytic coelomycetes from broadleaf trees. Mycologia 2012, 10, 198–210. [Google Scholar] [CrossRef]
- Voglmayr, H.; Aguirre-Hudson, M.B.; Wagner, H.G.; Tello, S.; Jaklitsch, W.M. Lichens or endophytes? The enigmatic genus Leptosillia in the Leptosilliaceae fam. nov. (Xylariales), and Furfurella gen. nov.(Delonicicolaceae). Persoonia 2019, 42, 228–260. [Google Scholar] [CrossRef] [Green Version]
- Saccardo, P.A. Fungi Veneti novi vel critici. Series I. Nuovo Giornale Botanico Italiano; Nabu Press: Florence, Italy, 1873; Volume 5, pp. 269–298. [Google Scholar]
- Fitzpatrick, H.M. The genus Fracchiaea. Mycologia 1924, 16, 101–114. [Google Scholar] [CrossRef]
- Huang, S.K.; Hyde, K.D.; Maharachchikumbura, S.S.N.; McKenzie, E.H.C.; Wen, T.C. Taxonomic studies of Coronophorales and Niessliaceae (Hypocreomycetidae). Mycosphere 2021, 12, 875–992. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–491: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Kohlmeyer, J.; Volkmann-Kohlmeyer, B.; Eriksson, O.E. Fungi on juncus roemerianus. 8. new bitunicate ascomycetes. Canad. J. Bot. 1996, 74, 1830–1840. [Google Scholar] [CrossRef]
- Suetrong, S.; Schoch, C.L.; Spatafora, J.W.; Kohlmeyer, J.; Volkmann-Kohlmeyer, B.; Sakayaroj, J.; Phongpaichit, S.; Tanaka, K.; Hirayama, K.; Jones, E.B.G. Molecular systematics of the marine Dothideomycetes. Stud. Mycol. 2009, 64, 155–173. [Google Scholar] [CrossRef]
- Nelsen, M.P.; Lücking, R.; Mbatchou, J.S.; Andrew, C.J.; Spielmann, A.A.; Lumbsch, H.T. New insights into relationships of lichen-forming Dothideomycetes. Fungal Divers. 2011, 51, 155–162. [Google Scholar] [CrossRef]
- Mortimer, P.E.; Jeewon, R.; Xu, J.C.; Lumyong, S.; Wanasinghe, D.N. Morpho-phylo taxonomy of novel Dothideomycetous fungi associated with dead woody twigs in Yunnan Province, China. Front. Microbiol. 2021, 12, 654683. [Google Scholar] [CrossRef]
- Spegazzini, C. Mycetes Argentinenses (Series V). An. Mus. Nac. Hist. Nat. Buenos Aires Ser. 1911, 3, 329–467. [Google Scholar]
- Koukol, O.; Delgado, G.; Hofmann, T.A.; Piepenbring, M. Panama, a hot spot for Hermatomyces (Hermatomycetaceae, Pleosporales) with five new species, and a critical synopsis of the genus. IMA Fungus 2018, 9, 107–141. [Google Scholar] [CrossRef] [Green Version]
- Phukhamsakda, C.; McKenzie, E.H.C.; Phillips, A.J.L.; Jones, E.B.G.; Bhat, D.J.; Stadler, M.; Bhunjun, C.S.; Wanasinghe, D.N.; Thongbai, B.; Camporesi, E.; et al. Microfungi associated with Clematis (Ranunculaceae) with an integrated approach to delimiting species boundaries. Fungal Divers. 2020, 102, 1–203. [Google Scholar] [CrossRef]
- Chang, H.S. Notes on Taiwan dematiaceous hyphomycetes, some species of the genera Exserticlava, Craspedodidymum and Hermatomyces. Bot. Bull. Acad. Sin. 1995, 36, 243–246. [Google Scholar]
- Ren, G.C.; Wanasinghe, D.N.; Monkai, J.; Mortimer, P.E.; Hyde, K.D.; Xu, J.C.; Pang, A.; Gui, H. Novel saprobic Hermatomyces species (Hermatomycetaceae, Pleosporales) from China (Yunnan Province) and Thailand. MycoKeys 2021, 82, 57. [Google Scholar] [CrossRef] [PubMed]
- Prasher, I.B.; Prasher, S. Hermatomyces indicus sp. nov. (Hyphomycetes) from India. Nova Hedwig. 2014, 99, 551–556. [Google Scholar] [CrossRef]
- Doilom, M.; Dissanayake, A.J.; Wanasinghe, D.N.; Boonmee, S.; Liu, J.K.; Bhat, D.J.; Taylor, J.E.; Bahkali, A.H.; McKenzie, E.H.C.; Hyde, K.D. Microfungi on Tectona grandis (teak) in Northern Thailand. Fungal Divers. 2016, 82, 107–182. [Google Scholar] [CrossRef]
- Boddy, L. Interspecific combative interactions between wood-decaying basidiomycetes. FEMS Microbiol. Ecol. 2000, 31, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Falconer, R.E.; Bown, J.L.; White, N.A.; Crawford, J.W. Modelling interactions in fungi. J. R. Soc. Interface 2008, 5, 603–615. [Google Scholar] [CrossRef] [Green Version]
- Promputtha, I.; Hyde, K.D.; McKenzie, E.H.; Peberdy, J.F.; Lumyong, S. Can leaf degrading enzymes provide evidence that endophytic fungi becoming saprobes? Fungal Divers. 2010, 41, 89–99. [Google Scholar] [CrossRef]
- Promputtha, I.; Lumyong, S.; Dhanasekaran, V.; McKenzie, E.H.C.; Hyde, K.D.; Jeewon, R.A. Phylogenetic evaluation of whether endophytes become saprotrophs at host senescence. Microb. Ecol. 2007, 53, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Botrel, D.A.; Laborde, M.C.F.; Medeiros, F.H.V.D.; Resende, M.L.V.D.; Ribeiro Júnio, P.M.; Pascholati, S.F.; Gusmão, L.F.P. Saprobic fungi as biocontrol agents of halo blight (Pseudomonas syringae pv. garcae) in coffee clones. Coffee Sci. 2018, 13, 283–291. [Google Scholar] [CrossRef]
- Monkai, J.; Tibpromma, S.; Manowong, A.; Mapook, A.; Norphanphoun, C.; Hyde, K.D.; Promputtha, I. Discovery of three novel Cytospora species in Thailand and their antagonistic potential. Diversity 2021, 13, 488. [Google Scholar] [CrossRef]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Definition | Primer | Annealing (PCR) | Reference |
|---|---|---|---|---|
| ITS | Internal transcribed spacer | ITS4 | a 55 °C, 50 s b | [20] |
| ITS5 | ||||
| LSU | Large subunit 28S | LR0R | a 55 °C, 50 s b | [21] |
| LR5 | ||||
| rpb2 | DNA-dependent RNA polymerase II largest subunit | 5F | a 57 °C, 55 s b | [22] |
| 7cR | ||||
| SSU | The partial18S small subunit | NS1 | a 55 °C, 50 s b | [20] |
| NS4 | ||||
| tef1-α | Translation elongation factor 1 alpha | 983F | a 56 °C, 50 s b | [23] |
| 2218R | ||||
| tub2 | Beta-tubulin 2 | Bt2a | a 52 °C, 50 s b | [24] |
| Bt2b |
| Class | Order | Family | Species | Herbarium No. |
|---|---|---|---|---|
| Dothideomycetes | Botryosphaeriales | Aplosporellaceae | Aplosporellaartocarpi | HKAS 122656 |
| Botryosphaeriaceae | Lasiodiplodia theobromae | HKAS 122660 | ||
| Botryosphaeriaceae | Lasiodiplodiapseudotheobromae | HKAS 122658 | ||
| Monoblastiales | Monoblastiaceae | Heleiosabarbatula | HKAS 122668 | |
| Pleosporales | Hermatomycetaceae | Hermatomyces indicus | HKAS 122676 | |
| Incertaesedis | Crassipariesquadrisporus | HKAS 122192 | ||
| Incertaesedis | Mangifericomes hongheensis | HKAS 122188 | ||
| Massarinaceae | Vaginatisporaamygdali | HKAS 122195 | ||
| Melanommataceae | Byssosphaeriasiamensis | HKAS 122197 | ||
| Neomassariaceae | Neomassaria hongheensis | HKAS 122191 | ||
| Paradictyoarthriniaceae | Paradictyoarthriniumdiffractum | HKAS 122194 | ||
| Parabambusicolaceae | Paramonodictys hongheensis | HKAS 122190 | ||
| Parabambusicolaceae | Paramonodictys yunnanensis | HKAS 122189 | ||
| Phaeoseptaceae | Phaeoseptummali | HKAS 122193 | ||
| Torulaceae | Torulafici | HKAS 122196 | ||
| Tubeufiales | Tubeufiaceae | Excipulariopsisnarsapurensis | HKAS 122680 | |
| Eurotiomycete | Chaetothyriales | Cyphellophoraceae | Cyphellophora hongheensis | HKAS 122661 |
| Mycocaliciales | Mycocaliciaceae | Chaenothecopsis hongheensis | HKAS 122672 | |
| Sordariomycetes | Calosphaeriales | Pleurostomataceae | Pleurostomaootheca | HKAS 122679 |
| Coronophorales | Nitschkiaceae | Fracchiaeamyricoides | HKAS 122671 | |
| Delonicicolales | Delonicicolaceae | Delonicicolasiamense | HKAS 122662 | |
| Diaporthales | Diaporthaceae | Diaporthe hongheensis | HKAS 122657 | |
| Glomerellales | Plectosphaerellaceae | Acremoniisimulans hongheensis | HKAS 122669 | |
| Sordariales | Helminthosphaeriaceae | Hilberina hongheensis | HKAS 122677 | |
| Xylariales | Diatrypaceae | Mangifericola hongheensis | HKAS 122665 | |
| Diatrypaceae | Paraeutypellacitricola | HKAS 122667 | ||
| Hypoxylaceae | Hypoxylon hongheensis | HKAS 122663 | ||
| Hypoxylaceae | Hypomontagnellamonticulosa | HKAS 122664 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, E.-F.; Dai, D.-Q.; Bhat, J.D.; Dawoud, T.M.; Promputtha, I.; Adikaram, N.; Stephenson, S.L.; Karunarathna, S.C.; Tibpromma, S. Taxonomic and Phylogenetic Studies of Saprobic Fungi Associated with Mangifera indica in Yunnan, China. J. Fungi 2023, 9, 680. https://doi.org/10.3390/jof9060680
Yang E-F, Dai D-Q, Bhat JD, Dawoud TM, Promputtha I, Adikaram N, Stephenson SL, Karunarathna SC, Tibpromma S. Taxonomic and Phylogenetic Studies of Saprobic Fungi Associated with Mangifera indica in Yunnan, China. Journal of Fungi. 2023; 9(6):680. https://doi.org/10.3390/jof9060680
Chicago/Turabian StyleYang, Er-Fu, Dong-Qin Dai, Jayarama D. Bhat, Turki M. Dawoud, Itthayakorn Promputtha, Nimal Adikaram, Steven L. Stephenson, Samantha C. Karunarathna, and Saowaluck Tibpromma. 2023. "Taxonomic and Phylogenetic Studies of Saprobic Fungi Associated with Mangifera indica in Yunnan, China" Journal of Fungi 9, no. 6: 680. https://doi.org/10.3390/jof9060680