
Ectomycorrhizal Fungal Community and Ascoma Production in a Declining Tuber borchii Plantation

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plantation Site
2.2. Ascoma Collection
2.3. Ectomycorrhizal Sampling
2.4. Molecular Identification of Ectomycorrhizae
2.5. Statistical Analysis
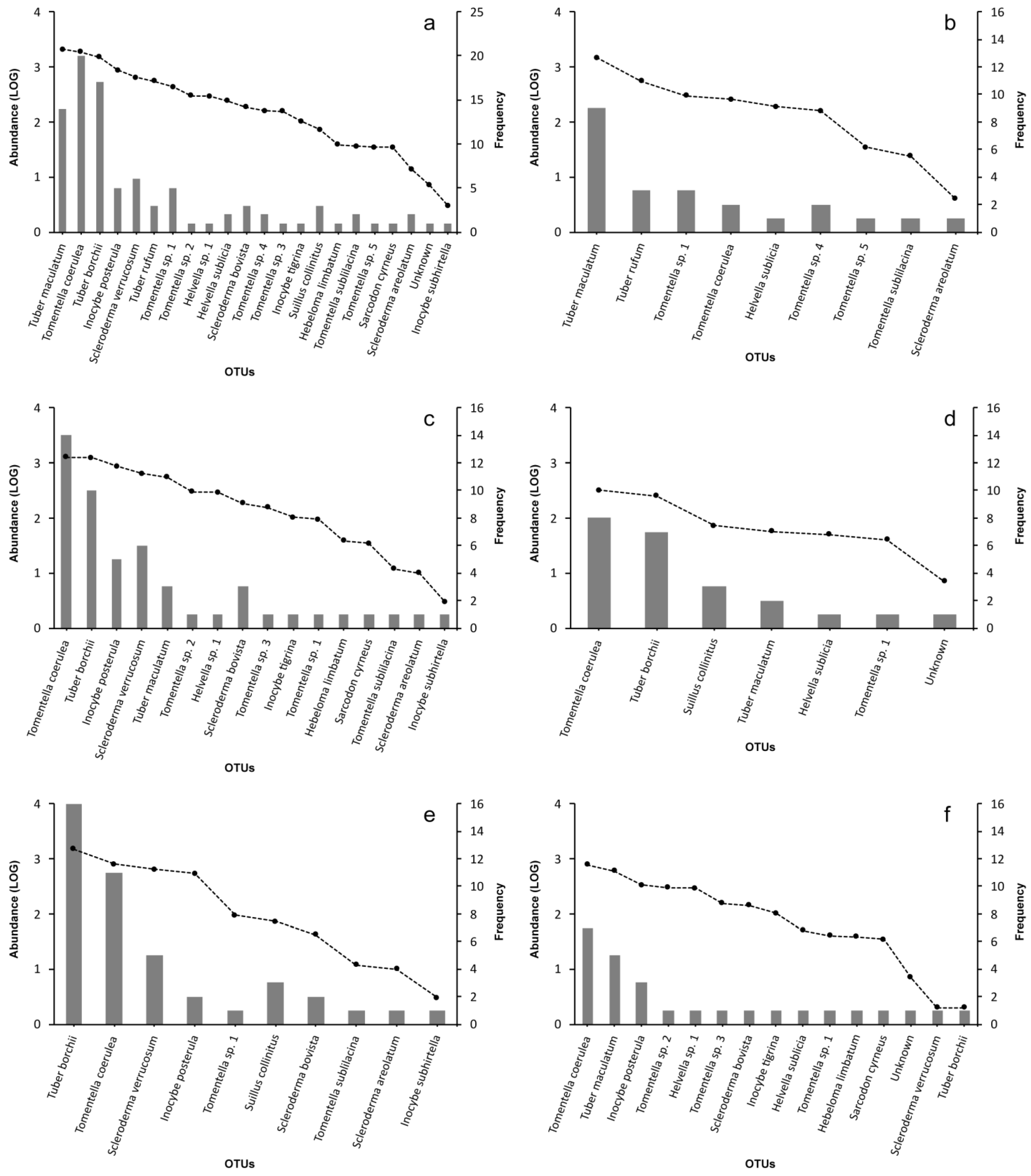
3. Results
3.1. Ascoma Production
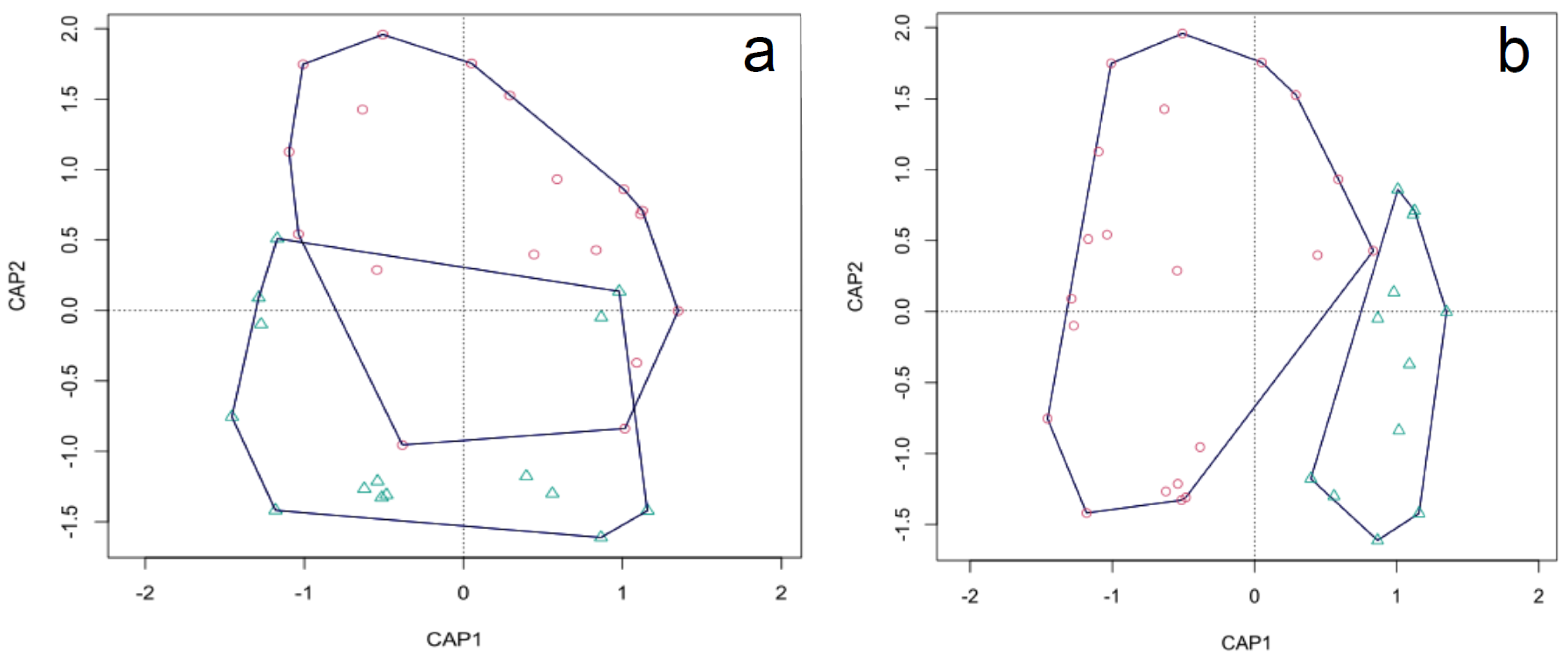
3.2. Whole ECM Fungal Community
3.3. Host Type (Pine vs. Hardwoods)
3.4. Sampling Position (FP vs. FA)
3.5. ECM Fungal Community in Proximity to Non-Inoculated C. avellana Seedlings (Samples NA)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hall, I.R.; Lyon, A.; Wang, Y.; Buchanan, P. A List of Putative Edible or Medicinal Ectomycorrhizal Mushrooms (Revised); Truffles and Mushrooms (Consulting) Limited: Dunedin, New Zealand, 2011. [Google Scholar]
- Zambonelli, A.; Iotti, M.; Boutahir, S.; Lancellotti, E.; Perini, C.; Pacioni, G. Ectomycorrhizal Fungal Communities of Edible Ectomycorrhizal Mushrooms. In Edible Ectomycorrhizal Mushrooms: Current Knowledge and Future Prospects; Zambonelli, A., Bonito, G., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2012; Volume 34, pp. 105–124. [Google Scholar]
- Karwa, A.; Varma, A.; Rai, M. Edible Ectomycorrhizal Fungi: Cultivation, Conservation and Challenges. In Diversity and biotechnology of Ectomycorrhizae; Rai, M., Varma, A., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2010; Volume 25, pp. 429–453. [Google Scholar]
- Iotti, M.; Piattoni, F.; Zambonelli, A. Techniques for Host Plant Inoculation with Truffles and Other Edible Ectomycorrhizal Mushrooms. In Edible Ectomycorrhizal Mushrooms: Current Knowledge and Future Prospects; Zambonelli, A., Bonito, G., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2012; Volume 34, pp. 145–161. [Google Scholar]
- Wang, Y.; Cummings, N.; Guerin-Laguette, A. Cultivation of Basidiomycete Edible Ectomycorrhizal Mushrooms: Tricholoma, Lactarius, and Rhizopogon. In Edible Ectomycorrhizal Mushrooms: Current Knowledge and Future Prospects; Zambonelli, A., Bonito, G., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2012; Volume 34, pp. 281–304. [Google Scholar]
- Wang, D.; Zhang, J.L.; Wang, Y.; Zambonelli, A.; Hall, I.; Xiong, W.P. The Cultivation of Lactarius with Edible Mushrooms. Ital. J. Mycol. 2021, 50, 63–77. [Google Scholar]
- Mello, A. State of the Art of the Research on Boletus edulis. In Edible Ectomycorrhizal Mushrooms: Current Knowledge and Future Prospects; Zambonelli, A., Bonito, G., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2012; Volume 34, pp. 73–81. [Google Scholar]
- Zambonelli, A.; Iotti, M.; Murat, C. True Truffle (Tuber spp.) in the World: Soil Ecology, Systematics and Biochemistry; Soil Biology; Springer International Publishing: Cham, Switzerland, 2016; Volume 47. [Google Scholar]
- Kagan-Zur, V.; Roth-Bejerano, N.; Sitrit, Y.; Morte, A. Desert Truffles: Phylogeny, Physiology, Distribution and Domestication; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2014; Volume 38. [Google Scholar]
- Bach, C.; Beacco, P.; Cammaletti, P.; Babel-Chen, Z.; Levesque, E.; Todesco, F.; Cotton, C.; Robin, B.; Murat, C. First Production of Italian White Truffle (Tuber magnatum Pico) Ascocarps in an Orchard Outside Its Natural Range Distribution in France. Mycorrhiza 2021, 31, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Graziosi, S.; Hall, I.R.; Zambonelli, A. The Mysteries of the White Truffle: Its Biology, Ecology and Cultivation. Encyclopedia 2022, 2, 1959–1971. [Google Scholar] [CrossRef]
- Hall, I.R.; Brown, G.T.; Zambonelli, A. Taming the Truffle: The History Lore and Science of the Ultimate Mushroom; Timber Press: Portland, OR, USA, 2007. [Google Scholar]
- Zambonelli, A.; Iotti, M.; Puliga, F.; Hall, I.R. Enhancing White Truffle (Tuber magnatum Picco and T. borchii Vittad.) Cultivation through Biotechnology Innovation. In Advances in Plant Breeding Strategies: Vegetable Crops; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer International Publishing: Cham, Switzerland, 2021; Volume 10, pp. 505–532. [Google Scholar]
- Zambonelli, A.; Iotti, M.; Rossi, I.; Hall, I. Interactions between Tuber borchii and Other Ectomycorrhizal Fungi in a Field Plantation. Mycol. Res. 2000, 104, 698–702. [Google Scholar] [CrossRef]
- Iotti, M.; Lancellotti, E.; Hall, I.; Zambonelli, A. The Ectomycorrhizal Community in Natural Tuber borchii Grounds. FEMS Microbiol. Ecol. 2010, 72, 250–260. [Google Scholar] [CrossRef] [Green Version]
- Iotti, M.; Piattoni, F.; Leonardi, P.; Hall, I.R.; Zambonelli, A. First Evidence for Truffle Production from Plants Inoculated with Mycelial Pure Cultures. Mycorrhiza 2016, 26, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Zambonelli, A.; Morara, M. Le Specie Di Tuberales Dell’Emilia Romagna, Ecologia e Distribuzione. Nat. E Mont. 1984, 4, 9–32. [Google Scholar]
- Morara, M.; Saitta, A.; Venturella, G.; Iotti, M.; Gargano, M.L.; Zambonelli, A. I Funghi Ipogei Dell’Emilia Romagna e Della Sicilia a Confronto. In Annales Confederationis Europaeae Mycologiae Mediterraneensis 2009; Venturella, G., Gargano, M.L., Eds.; CEMM, 2012; pp. 53–67. [Google Scholar]
- Tositti, L.; Pieri, L.; Brattich, E.; Parmeggiani, S.; Ventura, F. Chemical Characteristics of Atmospheric Bulk Deposition in a Semi-Rural Area of the Po Valley (Italy). J. Atmos. Chem. 2018, 75, 97–121. [Google Scholar] [CrossRef]
- Pegler, D.N.; Spooner, B.M.; Young, T.W. British Truffles: A Revision of British Hypogeous Fungi; Royal Botanic Gardens: Kew, UK, 1993. [Google Scholar]
- Ceruti, A.; Fontana, A.; Nosenzo, C. Le Specie Europee del Genere Tuber: Una Revisione Storica; Museo Regionale di Scienze Naturali: Torino, Italy, 2003. [Google Scholar]
- Amicucci, A.; Zambonelli, A.; Giomaro, G.; Potenza, L.; Stocchi, V. Identification of Ectomycorrhizal Fungi of the Genus Tuber by Species-specific ITS Primers. Mol. Ecol. 1998, 7, 273–277. [Google Scholar] [CrossRef]
- Iotti, M.; Amicucci, A.; Bonito, G.; Bonuso, E.; Stocchi, V.; Zambonelli, A. Selection of a Set of Specific Primers for the Identification of Tuber rufum: A Truffle Species with High Genetic Variability. FEMS Microbiol. Lett. 2007, 277, 223–231. [Google Scholar] [CrossRef] [Green Version]
- Agerer, R. Colour Atlas of Ectomycorrhizae; Einhorn-Verlag: Schwäbisch Gmünd, Germany, 1997. [Google Scholar]
- Iotti, M.; Zambonelli, A. A Quick and Precise Technique for Identifying Ectomycorrhizas by PCR. Mycol. Res. 2006, 110, 60–65. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS Primers with Enhanced Specificity for Basidiomycetes—Application to the Identification of Mycorrhizae and Rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Bonuso, E.; Zambonelli, A.; Bergemann, S.E.; Iotti, M.; Garbelotto, M. Multilocus Phylogenetic and Coalescent Analyses Identify Two Cryptic Species in the Italian Bianchetto Truffle, Tuber borchii Vittad. Conserv. Genet. 2010, 11, 1453–1466. [Google Scholar] [CrossRef]
- Lancellotti, E.; Iotti, M.; Zambonelli, A.; Franceschini, A. The Puberulum Group Sensu Lato (Whitish Truffles). In True Truffle (Tuber spp.) in the World: Soil Ecology, Systematics and Biochemistry; Zambonelli, A., Iotti, M., Murat, C., Eds.; Soil Biology; Springer International Publishing: Cham, Switzerland, 2016; Volume 47, pp. 105–124. [Google Scholar]
- Anderson, M.J.; Willis, T.J. Canonical Analysis of Principal Coordinates: A Useful Method of Constrained Ordination for Ecology. Ecology 2003, 84, 511–525. [Google Scholar] [CrossRef]
- Oksanen, J. Ordination and Analysis of Dissimilarities: Tutorial with R and Vegan; University Tennessee: Knoxville, TN, USA, 2009. [Google Scholar]
- Murat, C.; Vizzini, A.; Bonfante, P.; Mello, A. Morphological and Molecular Typing of the Below-Ground Fungal Community in a Natural Tuber magnatum Truffle-Ground. FEMS Microbiol. Lett. 2005, 245, 307–313. [Google Scholar] [CrossRef]
- Leonardi, M.; Iotti, M.; Oddis, M.; Lalli, G.; Pacioni, G.; Leonardi, P.; Maccherini, S.; Perini, C.; Salerni, E.; Zambonelli, A. Assessment of Ectomycorrhizal Fungal Communities in the Natural Habitats of Tuber magnatum (Ascomycota, Pezizales). Mycorrhiza 2013, 23, 349–358. [Google Scholar] [CrossRef]
- Zambonelli, A.; Iotti, M.; Zinoni, F.; Dallavalle, E.; Hall, I.R. Effect of Mulching on Tuber uncinatum Ectomycorrhizas in an Experimental Truffière. N. Z. J. Crop Hortic. Sci. 2005, 33, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Benucci, G.M.N.; Raggi, L.; Albertini, E.; Grebenc, T.; Bencivenga, M.; Falcinelli, M.; Di Massimo, G. Ectomycorrhizal Communities in a Productive Tuber aestivum Vittad. Orchard: Composition, Host Influence and Species Replacement. FEMS Microbiol. Ecol. 2011, 76, 170–184. [Google Scholar] [CrossRef] [Green Version]
- Benucci, G.M.N.; Raggi, L.; Albertini, E.; Gógán Csorbai, A.; Donnini, D. Assessment of Ectomycorrhizal Biodiversity in Tuber macrosporum Productive Sites. Mycorrhiza 2014, 24, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Belfiori, B.; Riccioni, C.; Tempesta, S.; Pasqualetti, M.; Paolocci, F.; Rubini, A. Comparison of Ectomycorrhizal Communities in Natural and Cultivated Tuber melanosporum Truffle Grounds. FEMS Microbiol. Ecol. 2012, 81, 547–561. [Google Scholar] [CrossRef] [PubMed]
- Otsing, E.; Tedersoo, L. Temporal Dynamics of Ectomycorrhizal Fungi and Persistence of Tuber melanosporum in Inoculated Quercus robur Seedlings in North Europe. Mycorrhiza 2015, 25, 61–66. [Google Scholar] [CrossRef]
- Taschen, E.; Sauve, M.; Taudiere, A.; Parlade, J.; Selosse, M.A.; Richard, F. Whose Truffle Is This? Distribution Patterns of Ectomycorrhizal Fungal Diversity in Tuber melanosporum Brûlés Developed in Multi-host Mediterranean Plant Communities. Environ. Microbiol. 2015, 17, 2747–2761. [Google Scholar] [CrossRef] [PubMed]
- Queralt, M.; Parlade, J.; Pera, J.; De Miguel, A.M. Seasonal Dynamics of Extraradical Mycelium and Mycorrhizas in a Black Truffle (Tuber melanosporum) Plantation. Mycorrhiza 2017, 27, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, M.; Iotti, M.; Pacioni, G.; Hall, I.R.; Zambonelli, A. Truffles: Biodiversity, Ecological Significances, and Biotechnological Applications. In Industrially Important Fungi for Sustainable Development: Biodiversity and Ecological Perspectives; Abdel-Azeem, A.M., Yadav, A.N., Yadav, N., Usmani, Z., Eds.; Springer International Publishing: Cham, Switzerland, 2021; Volume 1, pp. 107–146. [Google Scholar]
- Hilszczańska, D.; Szmidla, H.; Horak, J.; Rosa-Gruszecka, A. Ectomycorrhizal Communities in a Tuber aestivum Vittad. Orchard in Poland. Open Life Sci. 2016, 11, 348–357. [Google Scholar] [CrossRef]
- Ragnelli, A.M.; Aimola, P.; Maione, M.; Zarivi, O.; Leonardi, M.; Pacioni, G. The Cell Death Phenomenon during Tuber Ectomycorrhiza Morphogenesis. Plant Biosyst. 2014, 148, 473–482. [Google Scholar] [CrossRef]
- Mrak, T.; Grebenc, T.; Friedrich, S.; Münzenberger, B. Description, Identification, and Growth of Tuber borchii Vittad. Mycorrhized Pinus sylvestris L. Seedlings on Different Lime Contents; Slovenia Forestry Institute: Ljubljana, Slovenia, 2022; preprint article. [Google Scholar]
- Gardin, L.I. Tartufi Minori in Toscana: Gli Ambienti di Crescita dei Tartufi Marzuolo e Scorzone; Quaderno ARSIA 1/2005; ARSIA, Regione Toscana: Florence, Italy, 2005. [Google Scholar]
- Ori, F.; Leonardi, P.; Stagnini, E.; Balestrini, V.; Iotti, M.; Zambonelli, A. Is Tuber brumale a Threat to T. melanosporum and T. aestivum Plantations? iForest 2018, 11, 775. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Community Type | Root Samples (n) | Ectomyc. (n) | R | D | J |
|---|---|---|---|---|---|
| Whole | 41 | 9592 | 22 | 2.176 | 0.739 |
| Hardwoods | 17 | 5808 | 16 a | 2.155 a | 0.777 a |
| Pine | 15 | 798 | 7 b | 1.483 b | 0.762 b |
| Fruiting points (FP samples) | 17 | 3718 | 10 | 1.543 | 0.770 |
| Fruiting areas (FA samples) | 12 | 2888 | 15 | 2.081 | 0.768 |
| Non-inoculated trees (NT samples) | 12 | 2986 | 9 | 1.538 | 0.517 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ori, F.; Leonardi, M.; Puliga, F.; Lancellotti, E.; Pacioni, G.; Iotti, M.; Zambonelli, A. Ectomycorrhizal Fungal Community and Ascoma Production in a Declining Tuber borchii Plantation. J. Fungi 2023, 9, 678. https://doi.org/10.3390/jof9060678
Ori F, Leonardi M, Puliga F, Lancellotti E, Pacioni G, Iotti M, Zambonelli A. Ectomycorrhizal Fungal Community and Ascoma Production in a Declining Tuber borchii Plantation. Journal of Fungi. 2023; 9(6):678. https://doi.org/10.3390/jof9060678
Chicago/Turabian StyleOri, Francesca, Marco Leonardi, Federico Puliga, Enrico Lancellotti, Giovanni Pacioni, Mirco Iotti, and Alessandra Zambonelli. 2023. "Ectomycorrhizal Fungal Community and Ascoma Production in a Declining Tuber borchii Plantation" Journal of Fungi 9, no. 6: 678. https://doi.org/10.3390/jof9060678