

The Luminous Fungi of Japan

1
Department of Environmental Biology, Chubu University, Kasugai 487-8501, Aichi, Japan
2
Department of Botany, National Museum of Nature and Science, Tsukuba 305-0005, Ibaraki, Japan
*
Authors to whom correspondence should be addressed.
J. Fungi 2023, 9(6), 615; https://doi.org/10.3390/jof9060615
Submission received: 30 April 2023
/
Revised: 17 May 2023
/
Accepted: 24 May 2023
/
Published: 26 May 2023
(This article belongs to the Special Issue New Perspectives on Fungal Bioluminescence)
Abstract
:Luminous fungi have long attracted public attention in Japan, from old folklore and fiction to current tourism, children’s toys, games, and picture books. At present, 25 species of luminous fungi have been discovered in Japan, which correspond to approximately one-fourth of the globally recognized species. This species richness is arguably due to the abundant presence of mycophiles looking to find new mushroom species and a tradition of night-time activities, such as firefly watching, in Japan. Bioluminescence, a field of bioscience focused on luminous organisms, has long been studied by many Japanese researchers, including the biochemistry and chemistry of luminous fungi. A Japanese Nobel Prize winner, Osamu Shimomura (1928–2018), primarily focused on the bioluminescence system of luminous fungi in the latter part of his life, and total elucidation of the mechanism was finally accomplished by an international research team with representatives from Russia, Brazil, and Japan in 2018. In this review, we focused on multiple aspects related to luminous fungi of Japan, including myth, taxonomy, and modern sciences.
1. Introduction
The occurrence of dim-glowing mycelia on fallen twigs, which was first recognized as unknown ‘shining wood’, and luminous mushrooms on rotten trees has fascinated people around the world (Figure 1A), and Japan is no exception. The special interest in bioluminescent fungi in Japan is probably related to the richness of fungal diversity, which has led to a love of mushrooms and mushroom consumption in this country. In addition, a tradition of night-time activities could also be a factor.
The climate of mainland Japan (Hokkaido, Honshu, Shikoku, and Kyushu) ranges from subarctic in the northern part to temperate in the southern part; it is typically characterized by a cold winter and humid summer. The peripheral Izu Islands, Bonin Islands (Ogasawara Islands), and Ryukyu Archipelago (including Amami and Okinawa Islands) have subtropical oceanic climates with mild winters and hot humid summers. Because of wide-ranging climates with high humidity and species richness in woods and mountainous areas, Japan has a high biodiversity of fungi. Currently, approximately 13,000 described species of fungi have been reported from Japan, with possibly even more undescribed species [1]. Mycophagy has been popular in Japanese food culture, probably since Japan’s Jomon Period (–10th BCE) [2]. Various species of mushrooms have been cultivated and are always available on the market, such as Lentinula edodes (Shii-také, in Japanese), Flammulina filiformis (Enoki-také, formerly recognized as F. velutipes), Hypsizygus marmoreus (Buna-shimeji, also known as H. tessulatus), and Pholiota microspora (Nameko), which are indispensable for everyday Japanese cuisine [2,3]. Many amateur mycologists have been devoted to the understanding of Japanese fungal diversity, and several new mushroom species are found and described every year all across Japan by amateur and professional Japanese mycologists.
Since the old days, especially after the Edo Period, when people had more free time to enjoy their lives, there have been Japanese traditions of enjoying nature at night-time, including watching the moon (Tsukimi) and fireflies (Hotaru-gari), listening to insect calls (Mushi-kiki), and night hiking (Kōchū-tozan) [4]. Thus, it is not surprising that people accidentally witnessed bioluminescent mushrooms or glowing mycelia in the dark, and sometimes this glowing was thought to be caused by Yōkai: creatures, presences, or phenomena that could be described as mysterious or eerie [5]. In Japan, many terrestrial bioluminescent organisms have often been discovered by ordinary people or amateur naturalists based on sporadic observations. Examples of such organisms include the bioluminescent earthworms Microscolex phosphoreus [6] and Pontodrilus litoralis [7], the millipede Paraspirobolus lucifugus [8], the springtail Lobella sp. [9], the tiny mushroom Marasmiellus lucidus [10], and the scarlet mushroom Cruentomycena orientalis [11].
In this paper, we attempt to review bioluminescent mushrooms in Japan and related topics, such as folklore and taxonomy, and some recent research results on bioluminescent fungi in Japan. For this purpose, we intentionally cite many studies written in Japanese to share hidden achievements from Japan with the world.
2. Folklores
There are many folkloric stories about strange glows around the world [12], including Japan. One example is Kitsune-bi, meaning ‘fox’s fire’; it is very interesting coincidence that old Japanese called an unknown glow on the ground ‘fox’s fire’, using the same metaphor as Europeans calling glows ‘foxfire’. Another is Mino-bi, meaning ‘raincoat fire’; Mino is a Japanese traditional raincoat made out of rice straw, bi is fire or glow, and a folklore about the mysterious glow of wet Mino was called Mino-bi. Both Kitsune-bi and Mino-bi are partly considered to be responsible for the luminous mycelia growing on rotten wood or the straw of Mino [13,14]. Mino-bi was sometimes believed to be the work of foxes [15]. Figure 1B is a masterpiece Ukiyo-é woodblock by Hiroshige Utagawa at the end of Japan’s Edo Period, showing foxes bearing fire in their mouths under a large Japanese hackberry tree. Of note, Sakyo Kanda (1874–1939), the author of the book “Shiranui, Hitodama, and Kitsune-bi”, which sheds light on the mysterious luminescence, was a biologist of luminous organisms, and he considered the major cause of the Japanese foxfire to be a misinterpretation of people’s lanterns [14].
The glowing of old pine trees was known as the mysterious phenomena of Hikari-matsu (luminescent pine tree [16]; the reference originally written in 1765) or Ryū-tō (dragon’s lantern [17]), and these supernatural phenomena can most likely be explained by the luminescence of Armillaria’s mycelia [18]. Armillaria are known to be one of the common pathogens that cause wood rot in Pinus pine trees (Armillaria root rot, Narataké-byō in Japanese), which does occur in Japan [19].
Until recently (approximately 50 years ago), people on Hachijo Island (an island located in the Izu Islands) called the glow of the luminous mushrooms in the woods Hato-no-hi, meaning the fire of pigeons [20]. On this subtropical island, more than seven bioluminescent mushroom species are distributed, of which Mycena lux-coeli and Mycena chlorophos exhibit especially strong glows [21]. The Japanese wood pigeon Columba janthina is also distributed in the island woods. Most likely, people made a connection between the unidentifiable glow of luminous mushrooms in the night-time forest and the unsettling night call of the wood pigeon [22,23].
3. The Tale of the Bamboo Cutter
It is believed that luminescence of mycelia appeared as early as in a description by Aristotle (4th BCE) [12,24]. This roughly corresponds to the end of the Jomon Period in Japan, when people in mainland Japan were primarily hunter–gatherers and no written records were available. The first appearance of luminous fungi in Japanese literature was in ancient tales in Japan’s Heian Period (6–12th century).
The Tale of the Bamboo Cutter, “Taketori Monogatari”, was written by an unknown Japanese author in the late 9th or early 10th century and is recognized as the oldest Japanese work of fiction [25,26]. In the first part of this tale, there is an impressive scene in which a man called the Old Bamboo Cutter finds a glowing bamboo in the field.
“One day he noticed among the bamboos a stalk that glowed at the base”(translated by Keene, 1998 [25])
When he examines it, he finds a lovely little girl approximately three inches tall, named Shining Princess (Kaguya-himé in Japanese). She grows to be an adult, and she refuses several proposals of marriage from noble men, including the Emperor, and finally returns to the moon. As it turns out, she was a princess from the Moon.
What was causing the bamboo to glow? A Japanologist, Katsumi Masuda (1923–2010), hypothesized that the cause of the glowing was the luminous fungus Panellus pusillus (Suzume-také in Japanese) [27], and a historian, Michihisa Hotate, agreed with that idea [26]. This species also grows on the rotten culm (ringed stem) of Japanese bamboo [28,29]. The distribution records of P. pusillus were mainly in the southern part of Japan: Bonin Isls., Matsuyama (Ehime Prefecture) [28], and Hachijo Isl. [22,30], but also in central Japan [28]. Shidei reported the growth of P. pusillus on bamboo in Kyoto [29]. As the story is probably set in Kyoto, and at that time many bamboo craftworkers originating from the southern Kyushu region worked there [26], it can be speculated that the author of the tale had learned of the phenomenon of bamboo glowing from the craftworkers’ experiences, inspiring the famous opening scene [27]. Mycena chlorophos (Yakō-také in Japanese) is another candidate; it is distributed mainly in the southern islands, such as Hachijo Isl. and Bonin Isls. [30,31], but is also found in various localities in mainland Japan (Honshu: from Kanto west to Kyushu, including Kyoto) [31,32], and it sometimes grows on bamboo. Of course, we do not exclude the possibility of the glow being caused by luminous mycelia of unknown identity on rotten bamboo [14] (Figure 2) or nonfungal organisms, such as the princess firefly Luciola parvula (Himé-botaru), which sometimes appears in bamboo groves [33,34].
4. Current Commercialization
Currently, the phenomenon of fungi bioluminescence is familiar to many people in Japan. For example, there are the characters of the Pokémon Card Game (a card game that appeared in Japan in 1996 and is now a worldwide success) (Figure 1C), capsule toys (small toys in vending machines packaged in a plastic capsule) (Figure 1D), and picture books that focus on luminous mushrooms [22,35,36] (Figure 1E). TV programs featuring mushrooms often include the topics of luminescence of some mushroom species. Artificial cultivation methods of M. chlorophos have been established [37,38], and the species has been used for special exhibitions in several local museums and botanical gardens (e.g., Hachijo Visitor Center, Tokyo; Yumenoshima Tropical Greenhouse Dome, Tokyo; and Nagoya City Science Museum, Aichi). A culture kit is available online [39]. Night-time hiking ecotours to watch glowing M. chlorophos are one of the most economically significant tour activities in Hachijo Isl. and Bonin Isls. [22,40], and trips to watch glowing M. lux-coeli have occasionally been held at various localities in southern Japan, such as the Amami Islands (Kagoshima Prefecture), Mt. Yokokura (Kochi Prefecture), and Ukui Peninsula (Wakayama Prefecture).
5. Taxonomy
5.1. Bioluminescent Species in Japan
In Japan, scientific surveys of fungi started around the 18th century when Japanese scholars were emancipated from Chinese herbalism Honzō-gaku and began genuine native studies on Japanese flora. For example, a Japanese herbalism scholar, Tomohiro Ichioka (1739–1808), compiled a monograph of local fungi, “Shin-you Kinpu,” in 1799 and mentioned (probably) Omphalotus japonicus as Kumahira with illustration and the remarks “luminescence at night and poisonous.” Another Japanese herbalism scholar, Konen Sakamoto (1800–1853), compiled a monograph of the Japanese fungi “Kinpu” in 1835 and described O. japonicus as “Tsukiyo-také” with illustration and the remarks “this mushroom is called Tsukiyo-také because of luminescence at night”. Though based on pre-Linnean classification, these are probably the earliest scientific references about the bioluminescent mushroom in Japan. However, other luminous species were not described until the 20th century. This is partially because of the climate diversity of Japan. Mainland Japan, where most Japanese people including scholars are located, is characterized by a subarctic to temperate climate, while many luminous mushroom species, especially of the Mycena group, are distributed in tropical and subtropical regions. In other words, O. japonicus is the only bioluminescent mushroom species commonly (frequently) observed in Japan.
Indeed, O. japonicus is the first luminous mushroom species described scientifically under the Linnean system, which was formulated in 1915 by a mycologist, Seiichi Kawamura (1881–1946) [41]. The second piece of scientific evidence of luminescent fungi from Japan was reported by Yosio Kobayasi (1907–1993), who reported the luminescence of four known (currently three) species: Favolaschia peziziformis, Panellus pusillus, Mycena chlorophos (from Bonin Isl.), and Mycena cyanophos (=M. chlorophos) (Bonin Isl., and also from Hachijo Isl.) [42].
Before and during the Second World War, a Japanese researcher of bioluminescent organisms, Yata Haneda (1907–1995), extensively surveyed luminous mushrooms when he stayed at Palao Tropical Biological Station in Palau as a researcher under the mandate of Japan (during 1937–1942) and as Army Civil Administrator of Shonan Museum (the present National Museum of Singapore) in Singapore (during 1942–1945). After the Second World War, he returned to Japan and continued his luminous mushroom survey at Hachijo Island, Japan, and described several luminous mushrooms from Japan with assistance from the British mycologist/botanist Edred John Henry Corner (1906–1996) [43]. Of note, at the end of the Second World War, Corner was a captive prisoner of Japan. Thus, the official relationship between Haneda and Corner was that of enemies, but they struck up a scientific friendship during and after the war [44]. Although many of these species names described by Haneda and Corner were invalidly published, which unfortunately caused taxonomic confusion [45,46], their contributions paved the way for understanding the diversity of luminous mushrooms in Japan after the Second World War; Mycena lux-coeli (Shiino-tomoshibi-také) was collected by Haneda on Hachijo Island and described by Corner, and the species name remains valid.
Even recently, many new localities of luminescent mushroom species have been recorded, and Terashima and her colleagues described eight new luminescent species from southwestern Japan in their book [47]. Currently, approximately 100 species of luminous fungi have been recognized [48,49], of which 25 species are distributed in Japan [22,46,47].
In this section, all luminescent fungal species recognized in Japan are listed with remarks. Phylogenetic positions of these species are not presented in this paper, but some previous studies based on genome-scale DNA data have clearly demonstrated the relative positions of major bioluminescent genera and the polyphyly of bioluminescent taxa among mushroom-forming fungi [50,51]. Species that were “excluded, doubtful or insufficiently known” [46] were not included. Of note, Nothopanus noctilucens is sometimes listed as a luminescent species distributed in Japan [46]. However, the Japanese Pleurotus noctilucens (=Nothopanus noctilicens) sensu Inoko is an invalid name [52], and the true Pleurotus noctilucens Lév. (Syn. Nothopanus noctilucens) has not been reported in Japan [53]. Mycena illuminans has been reported as a luminescent species distributed in Japan [46]. However, this species is often considered a synonym of M. chlorophos [54] and thus is not included in the following list. The Japanese names were adopted from the list by Katumoto, 2010 [55], unless they have a more recent name.
5.1.1. Family Mycenaceae
Cruentomycena orientalis Har. Takah. & Taneyama
Japanese name: Gahnetto-ochiba-také [47]
Remarks: ‘Gahnetto’ means garnet in Japanese. The suffix ‘-také’ means mushroom. This species was described from Ishigaki Island, southern Japan [47]. The luminescence of the mycelium and fruitbody of this species and other similar species was reported in Fukuoka, Miyazaki, and Miyagi Prefectures [11,56,57]. The luminescence of the fruitbody was weak compared to that of the mycelium and detected only by a long-exposure CCD camera.
Dictyopanus foliicola Kobayasi
Japanese name: Konoha-suzume-také [55]
Remarks: The Japanese konoha and suzume mean leaf and sparrow, respectively (‘sparrow’ represents a small creature in Japanese [58]). The mycelia and fruitbodies are luminous. This species has not been officially reported since the original description by Kobayasi from Miyazaki Prefecture [59]. The taxonomic status of this species warrants further study.
Favolaschia peziziformis (Berk. & M. A. Curtis) Kuntze (Figure 3)
Japanese name: Enashi-rasshi-také [55]
Remarks: This species was originally described from the Bonin Islands but is also known on Hachijo Island, Okinawa and other countries in Australasia [60]. Whole fruitbodies are reported to be bioluminescent. Enashi means the lack of a stipe. Rasshi derives from the genus name Laschia in honor of German mycologist Wilhelm Gottfried Lasch (1787–1863) [58].
Mycena chlorophos (Berk. & M. A. Curtis) Sacc. (Figure 4)
Japanese name: Yakō-také [55]
Remarks: The Japanese Yakō-také means ‘night-illuminating mushroom’. This species is distributed in Honshu (probably from Aomori, the northernmost prefecture [61]), Shikoku, Kyushu, Izu Islands, and Bonin Islands [62]. In addition, the species is widely recorded in the Southern Pacific islands, e.g., Polynesia and Micronesia [62]. This species is listed in the Japanese Red Data as endangered in Fukushima, Chiba, and Miyazaki Prefectures [63]. The bioluminescence of the fruitbody is considered brighter than many other known luminous mushrooms, but some strains, such as a strain in Miyazaki and Aomori Prefectures, seem darker compared to those in Hachijo and Bonin Islands [10,36]. The draft genome sequence of this species (Hachijo Isl. strain) has been assembled [64]. We consider Mycena cyanophos (Berk. & M.A. Curtis) Sacc. to be a synonym.
Mycena daisyogunensis Kobayasi
Japanese name: Hyūga-yakō-také [55]
Remarks: This species was collected from Daisyogun Cave in Miyazaki Prefecture in Kyushu (Hyūga is an old name of Miyazaki Prefecture) [59], but no further collections have been made since the original description. The taxonomic status of this species warrants further study.
Mycena flammifera Har. Takah. & Taneyama
Japanese name: Mori-no-ayashi-bi [47]
Remarks: The Japanese Morino-ayashi-bi means ‘forest ghost-fire’. This species was described from Ishigaki Island, southern Japan [47]. The morphological differences from the better-known bioluminescent species, M. manipularis (Berk.) Sacc. are subtle, and the taxonomic status of this species warrants further study.
Mycena lazulina Har. Takah., Taneyama, Terashima & Oba (Figure 5)
Japanese name: Konruri-kyūban-také [47]
Remarks: The Japanese Konruri-kyūban-také means ‘ultramarine-colored sucker mushroom’ because of the presence of a vivid blue (Konruri) disk-like (Kyūban) base. This species was described from the Yaeyama Islands, southern Japan [47]. Although its morphological characteristics seem to indicate that it belongs to the genus Mycena, the phylogenetic tree presented by Terashima et al. [47] has a very long branch leading to this species. The quality of DNA sequence data warrants further investigation.
Mycena lux-coeli Corner (Figure 6)
Remarks: The Japanese Shiino-tomoshibi-také means ‘Castanopsis tree’s lantern mushroom’. This species was originally described from Hachijo Island, but it is currently known from wider areas of central to southern Japan, mostly along the Pacific Ocean [60]. This species is listed in the Red Data as endangered in Mie Prefecture and as vulnerable in Miyazaki Prefecture [63].
Mycena luxfoliata Har. Takah., Taneyama & Terashima
Japanese name: Kareha-yakō-také [47]
Remarks: This species was described from the Ishigaki and Iriomote Islands, southern Japan [47]. Its bioluminescence was observed from mycelia on fallen leaves (Kareha means fallen leaves).
Mycena manipularis (Berk.) Sacc. (Figure 7)
Syn. Filoboletus manipularis (Berk.) Singer, Polyporus hanedae Kawam.
Japanese name: Ami-hikari-také [55]
Remarks: The Japanese Ami-hikari-také means ‘reticulated luminous mushroom’. This species is known from central to southern Japan and has also been recorded in many other countries, including Indonesia and Australia [60]. It is listed in the Red Data as endangered in Chiba Prefecture and as near threatened in Miyazaki Prefecture [63]. The unique feature of this species is that its stems, rather than caps, are brightly luminous (Figure 7). The bioluminescent property seems erratic; it has been reported for the strain on Okinawa Island that nonluminescent and weak-luminescent fruitbodies sometimes appeared when cultivated in the laboratory [65]. The bioluminescence of the local strain in Miyazaki Prefecture seemed weaker [10]. Currently, the species is often called Filoboletus manipularis (Berk.) Singer.
Mycena pseudostylobates Kobayasi
Japanese name: Kyūbantaké-modoki [55]
Remarks: The Japanese Kyūbantaké-modoki means ‘pseudo sucker-mushroom’. This species was recorded from Miyazaki Prefecture, but no definitive collections have been made since the original description by Kobayasi, 1951 [59]. The taxonomic status of this species warrants further study. The mycelium is bioluminescent, but the luminosity of the fruitbody is unknown [59].
Mycena stellaris Har. Takah., Taneyama & A. Hadano (Figure 8)
Japanese name: Hoshino-hikari-také [47]
Remarks: The Japanese Hoshino-hikari-také means ‘starlight mushroom’. This species was described from the Ishigaki and Okinawa Islands, southern Japan [47]. The bioluminescence of the whole fruitbodies was recorded.
Panellus pusillus (Pers. ex Lév.) Burds. & O. K. Mill. (Figure 9)
Remarks: The Japanese name Hinano-uchiwa means ‘princess fan’. This species is known from central to southern Japan but is also widely reported from North and South America and Australasia [60,62]. It often grows on bamboo.
Resiomycena fulgens Har. Takah., Taneyama & Oba (Figure 10)
Japanese name: Ginga-také [47]
Remarks: This species is known from Yaku Isl. (Kagoshima Prefecture), Hachijo Isl., and Kochi Prefecture [47]. The fruitbodies are small (up to ca. 3 mm), but they often grow in large numbers on the standing timber of Castanopsis, visually evoking an image of the Milky Way (Ginga means the Galaxy or Milky Way). Whole fruitbodies were reported to be bioluminescent [47].
Roridomyces sp.
Japanese name: Aya-hikari-také
Remarks: Its taxonomic status has not been thoroughly studied, but it presumably represents a new species of the genus based on several morphological characteristics. Bioluminescence of Japanese samples (spores) was reported by Kurogi, 2015 [10]. The Japanese name Aya is derived from the fact that the species was discovered from Aya, Miyazaki Prefecture [10]. This species is listed in the Red Data as endangered in Miyazaki Prefecture [63].
5.1.2. Family Omphalotaceae
Marasmiellus lucidus Har. Takah., Taneyama & S. Kurogi
Japanese name: Himé-hotaru-také [47]
Remarks: Hotaru means firefly in Japanese. This species was discovered in Miyazaki Prefecture [47] during a survey of the Himé-botaru firefly (L. parvula) [10]. The whole fruitbodies were reported to be bioluminescent.
Marasmiellus venosus Har. Takah., Taneyama & A. Hadano
Japanese name: Himé-hikari-také [47]
Remarks: The Japanese Himé-hikari-také means ‘princess luminous mushroom’. This species was described from Oita Prefecture in Kyushu [47]. The whole fruitbodies and mycelia are both reported to be bioluminescent. This and the previous species belong to the genus Marasmiellus, but their taxonomic treatment warrants further investigation. Currently, no other species are known to be bioluminescent in the genus Marasmiellus, and their accurate phylogenetic relationship to other bioluminescent species will give important insights into the evolution of bioluminescence in fungi.
Omphalotus japonicus (Kawam.) Kirchm. & O. K. Mill. (Figure 11)
Syn. Lampteromyces japonicus (Kawam.) Sing.
Japanese name: Tsukiyo-také (old names: Watari, Bunano-kataha, Kumahira, Hikari-goke, and Hotaru-také) [55].
Remarks: The Japanese Tsukiyo-také means ‘moonlit-night mushroom’. This species is distributed widely in mainland Japan and is thus arguably one of the most well-known bioluminescent mushrooms in Japan. An anecdote in the mid-Edo Period (ca. 1800s), “Zoku Sanshū Kidan”, introduced a story called “Nanao Kōrin” where there was a bright luminescent mushroom called Yamiyo-také, meaning black-night mushroom, in Nanao (the current Nanao City in Ishikawa Prefecture); it claimed that the luminescence was strong enough to illuminate 1 m square when holding 2–3 pieces “like noon” [66].
Of course, this story most likely contains some hearsay exaggeration (the true luminescence of O. japonicus is such that “the fungi of different sizes could be easily recognized at a distance of thirty meters” in pitch dark, and of course not like noon, [41]), but this mushroom could possibly be O. japonicus because the story also introduces its gastrointestinal toxicity for humans, which is a characteristic property of this species [67]. The poisonousness of this species is well recognized in Japan because the fruitbody is similar to several Japanese edible species, including Pleurotus ostreatus (Hira-také, in Japanese), Pleurotus pulmonarius (Usu-hirataké), Lentinula edodes (Shii-také), and Sarcomyxa edulis (Muki-také), such that it is often consumed mistakenly [40,68]. In the 12th century tale “Konjaku Monogatarishū”, there is a story where a priest in Nara planned to kill his old supervisor to obtain the supervisor’s position by serving cooked O. japonicus (old Japanese name, Watari) under the guise of the edible mushroom P. ostreatus. Eventually, the old supervisor ate all of the mushroom dishes and said, “For years, this old priest has never had such deliciously cooked watari”; the old supervisor knew all along, but he was of a special constitution such that he never got affected by the toxin [69].
Currently, Japan experiences approximately 30 cases of mushroom poisoning annually, and the cases of O. japonicus are among the highest every year, accounting for approximately 50% of the cases [70]. The primary toxic substance was isolated and identified as illudin S (lampterol) by two Japanese organic chemists, Koji Nakanishi (1925–2019) and Takeshi Matsumoto (1923–2014) [71,72]. The major symptoms of the toxin are vomiting, diarrhea, and stomachache. In one case, curiously, “They felt dizzy and everything around them appeared blue to their eyes. Moreover, they experienced a feeling as if a number of fire-flies were flying around them” [41]. The draft genome sequence of this species (Korean cultivar) was assembled, and bioluminescence-related genes were identified [73]. Haneda reported weak luminescence of the spore mass on moist paper based on specimens collected from Akita Prefecture [30]. This species has an essential role in beech log decomposition in cool temperate forest floors in Japan [74], and because of the recent decline in natural beech forests, it is listed in some prefectural Red Data as a threatened species (e.g., Mie, Osaka, and Shimane Prefectures) [63]. Tsukiyo-také is one of the seasonal terms of the Japanese short poetry Haiku for mid-autumn [75].
“Wolves wander along/mountain trails, their ways lit by/moonlit-night mushrooms”, Kansuke Naka (1885–1965, a Japanese novelist, essayist, and poet) (translated by Nathaniel Guy [3], and his personal communication).
5.1.3. Family Physalacriaceae (Figure 12)
Armillaria cepistipes Velen.
Japanese name: Kuroge-narataké [55]
Remarks: Bioluminescence of Japanese samples (mycelium) was reported by Hiroi, 2006 [18]. Japanese Kuroge means black hair.
Armillaria gallica Marxmuller & Romagn.
Remarks: Bioluminescence of Japanese samples (mycelium) was reported by Hiroi, 2006 [18]. Luminescence of the rhizomorphs has been reported elsewhere [76] but not from the Japanese samples. The fruitbodies of several Armillaria species, including A. gallica and A. mellea, are popular in Japan as a tasty edible mushroom species [62]. Japanese Narataké means ‘oak mushroom’, although the Armillaria species also grow on other varieties of tree. Watage means fluff because the veil of this mushroom is covered by a fluff-like structure [21]. Yawa means soft.
Armillaria mellea (Vahl) P. Kumm.
Remarks: Bioluminescence of Japanese samples (mycelium), which are sometimes called Armillaria mellea subsp. nipponica J.Y. Cha & Igarashi, was reported by Hiroi, 2006 [18]. Luminescence of young rhizomorphs is also reported [62]. Japanese Harigane and Kuri mean wire and chestnut tree, respectively.
Armillaria nabsnona T. J. Volk & Burds.
Japanese name: Yachi-narataké [55]
Remarks: Bioluminescence of Japanese samples (mycelium) was reported by Hiroi, 2006 [18]. Japanese Yachi means marsh land because this species appears in marsh areas [68].
Armillaria ostoyae (Romagn.) Herink
Remarks: Bioluminescence of Japanese samples (mycelium) was reported by Hiroi, 2006 [18]. Japanese Oni and Tsuba mean a Yōkai ogre and mushroom ring (annulus) [58]. The mushroom possesses an obvious veil. Rough scales on the cap evoke the image of violent Oni [21].
Armillaria sp.
Japanese name: Kitsubu-narataké [77]
Remarks: Its taxonomic status has not been thoroughly studied, but it presumably represents a new species of the genus based on several morphological characteristics. Bioluminescence of Japanese samples (mycelium) was reported by Hiroi, 2006 [18]. In Japanese, kitsubu means yellow dots, referring to this characteristic of the cap surface.
Desarmillaria tabescens (Scop.) R. A. Koch & Aime
Syn. Armillaria tabescens (Scop.) Emel
Japanese name: Narataké-modoki [55]
Remarks: Luminescence intensities of the fruitbody measured by a chemiluminescence detector largely depend on the specimens, but even in the most luminescent specimen, the light was too weak to be observed by the human eye [18,78]. Luminescent intensities of the mycelia also vary, but some could be clearly observed by the human eye [18,78]. The luminescence intensities are correlated with the strains of fruitbody and mycelium, suggesting that the luminescence characteristics are hereditary [18,78]. The species has long been known as Armillaria tabescens but was recently transferred to a newly established genus, Desarmillaria [79]. Japanese -modoki means pseudo, because this mushroom is similar to that of Narataké (A. mellea), but it possesses no veil [21]. This mushroom is regarded as edible but can cause gastrointestinal disorders when consumed in large quantities [10].
5.1.4. Family Pleurotaceae
Pleurotus nitidus Har. Takah. & Taneyama (Figure 13)
Japanese name: Shiro-hikari-také [47]
Remarks: The Japanese Shiro-hikari-také means ‘white luminescent mushroom’. This species from Ishigaki and Iriomote Islands, southern Japan, was described as being new [47]. However, it probably needs to be transferred to other genera containing bioluminescent species, such as Neonothopanus or Nothopanus, based on its morphological characteristics. Currently, no bioluminescent species are known from the genus Pleurotus and closely related genera. The only exception can be seen in Pleurotus eugrammus [46], but it is now treated as Nothopanus eugrammus, a species more closely related to Omphalotus and only distantly related to Pleurotus [80].
5.2. Nonbioluminescent Species Based on Samples Collected from Japan
Panellus stipticus (Bull.) P. Karst.
Remarks: The Japanese name is Wasabi-také because of the strong pungent taste of the fruitbody, similar to ‘wasabi’, a spicy green paste served with sushi [31,62]. It is widely distributed in the world [60,62]. The North American population of this species is luminescent, but the European and Japanese strains are nonluminescent. The North American and European strains are interfertile, and luminosity is dominant over nonluminosity [81]. Samples from Turkey (nonbioluminescent) lack the genes related to bioluminescence (luciferase, hispidin-3-hydroxylase, and hispidin synthetase) in the genome [50]. Fruitbodies are frequently attacked by slugs, which may be important agents in the dispersal of their spores [82], but their involvement in bioluminescence for the attraction of dispersers is unknown (see Section 7). Japanese Kawaki- means ‘dried-’.
5.3. Potentially Bioluminescent Species in Japan
There are several fungal species that have been reported to be bioluminescent elsewhere but not in Japan. Some (probably most) of these species are bioluminescent at least in mycelial stages. According to Desjardin et al., “mycelium of most (if not all) Armillaria species is luminescent” [46], and thus the Armillaria species, in which bioluminescence has not been reported in Japan, might also be bioluminescent: for example, A. jezoensis Cha & Igarashi (Japanese name, Kobari-narataké [55]), A. singula J. Y. Cha & Igarashi (Hitori-narataké [55]), and A. tympanitica (Berk. & M. A. Curtis) Sacc. (which has no Japanese name, but was collected once from Bonin Isl. [83], although Ito suggested its species identification was doubtful [84]). Since bioluminescence of Gerronema viridilucens mycelia and fruitbodies has recently been reported from Brazil [85], the congeneric species recorded from Japan (such as G. holochlorum and G. nemorale) may also be bioluminescent. Table 1 summarizes the species reported to be bioluminescent elsewhere but not in Japan.
6. Bioluminescence
6.1. Before the Meiji Period
Chemical research on luminous fungi began in the 17th century, coinciding with the Scientific Revolution. Sir Robert Boyle (1627–1691), a founder of modern chemistry, examined luminous mycelia on rotten wood, called ‘shining wood’, using his newly invented vacuum pump and showed the involvement of air in luminescence [98]. Although Boyle did not know that the luminescence of ‘shining wood’ was caused by the mycelia of luminous fungi, his elegant experiments were recognized as a pioneering study leading the chemical understanding of bioluminescence. Jean-Henri Fabre (1823–1915), a French entomologist, demonstrated the requirement of oxygen for the luminescence of the ‘jack-o’-lantern’ mushroom growing on the olive tree, Omphalotus olearius [99]. In contrast, Japan had a late start in modern science, which is also true for the chemistry of fungal bioluminescence. In the days of Boyle and Fabre, during Japan’s Edo Period, Japanese people still believed in the story of Yamiyo-také (see above) or simply composed Haiku poems on the luminescence of mushrooms [100].
6.2. After the Meiji Revolution
In Japan, modern science started after the Meiji Revolution in 1868. Most likely, S. Kawamura was the first Japanese individual who studied the bioluminescence of fungi. In his descriptive paper of O. japonicus, he also reported the effects of nitrogen, hydrogen, and oxygen gasses on the luminosity of O. japonicus fruitbodies, showing that the luminescence did not fade when it was treated with oxygen [41,101]. These results are basically the same as those using O. olearius by Fabre [99], as described above. Kawamura also observed that the juice squeezed out from the luminous gills had no luminosity [41,67].
6.3. Airth’s Achievement
Bioluminescence reactions in vitro using extracts of luminous fungi had not yet been achieved despite decades of trials (Harvey, 1952 [24], and references therein), but Airth and McElroy finally succeeded using the extracts of ‘luminous fungi’ species [102]. They detected luminescence when a hot-water extract from the mycelia of the luminous fungus Armillaria mellea and a cold-water extract from the mycelia of the luminous fungus ‘Collybia velutipes’ were mixed in the presence of NADH or NADPH [102,103]. Airth and Foerster showed that this reaction consists of at least two steps, involving the reduction of unidentified dehydro- or oxyluciferin (luciferin precursor) by NADH or NADPH with a soluble enzyme (approximately 25 kDa [104]) and light-emitting oxidation of luciferin by molecular oxygen with an insoluble membrane-bound enzyme [103,105]. Airth and Foerster also showed that a cold-water extract from Panellus stipticus (Panus stipticus, in their paper) mycelia (luminescent strain) exhibited luminescence activity for a hot-water extract from A. mellea mycelia with NADH [106]. Based on these results, they suggested that fungal luminescence is explained as a luciferin–luciferase reaction, and the presence of reduced pyridine nucleotides in the reaction mixture is key to reproducing the luminescence in vitro [104]. Regarding the species ‘Collybia velutipes’, the authors of this paper wrote that the mycelia of ‘C. velutipes’ used for cold-water extraction was “luminous” [106], but this species is currently recognized as nonluminous Flammulina velutipes [46,76]. We confirmed that a cold-water extract of the F. velutipes (or F. filiformis) fruitbody has no luminescence activity by mixing a hot-water extract of the M. chlorophos fruitbody and NADPH. Interestingly, on the other hand, strong luminescence activity was detected in a hot-water extract of the F. velutipes fruitbody by mixing a cold-water extract of the M. chlorophos fruitbody and NADPH [107]. Accordingly, we conclude that Airth’s research group misidentified unknown luminous mycelia as ‘C. velutipes’, and true F. velutipes has no enzymatic activity but unexpectedly contains a luciferin precursor. Later, we realized that several nonluminous fungi contain a large amount of the luciferin precursor hispidin [108], as described below.
6.4. Various Candidate Compounds
Candidates of luciferin or compounds involved in fungal luminescence have been proposed by several research groups (Figure 14). Airth et al. [103] showed that the luciferin precursor is soluble in phosphate buffer (pH 7.5) with 2% Tween 80 rather than in organic solvents, including acetone, chloroform, benzene, and diethyl ether [103]. In 1966, Kuwabara and Wassink reported the purification and crystallization of an ‘active substance’ (luciferin or dehydroluciferin) from the mycelia of the luminous fungi Mycena citricolor (Omphalia flavida, in the paper); this substance had enzymatic luminescent activity for “Dr. Airth’s fungal luciferase system” and nonenzymatic chemiluminescent activity in the presence of H2O2 [109]. The crystal was a brownish-orange ‘microcrystalline solid’ (needle-like crystalline, see Kuwabara & Wassink, 1966 [109]). The UV, IR, and fluorescence of this ‘active substance’ were measured, but the chemical structure was not determined. Later, Airth himself examined the enzymatic activity of this crystal shipped from Kuwabara using a cold-water extract of ‘C. velutipes’ with NADH or NADPH. However, the result was negative: “Since the possibility of inactivation during transport has not been disproved, the question as to whether Kuwabara and Wassink (1966) did crystallize fungal luciferin remains unanswered” [104]. Wassink suggested that there remains some doubt whether this crystallized compound was indeed fungal luciferin [76]. From the current viewpoint, it is believed that the basic optical properties (UV and fluorescent spectra) of the ‘active substance’ correspond with neither of the candidate precursors of luciferin (hispidin and caffeic acid); both have luminescence activities to the crude buffer extract of the luminescent fungi [108,110]. Of note, Seishi Kuwabara, the first author of the paper about crystallization of the ‘active substance’, is a Japanese biologist. His biography is uncertain, but he also worked on bacterial luciferase with a bioluminescence researcher, Milton Cormier, in the 1960s (see Kuwabara et al., 1965 [111]).
Nakanishi and his colleague isolated fluorescent compounds illudin S (Figure 14A) and ergosta-4,6,8(14),22-tetraen-3-on (Figure 14B), which showed emission maxima close to those of fungal luminescence, from the mycelia and fruitbody of O. japonicus as potential substances involved in the bioluminescence of fungi [112]. Illudin S is a compound responsible for the toxicity of O. japonicus, as shown above. The bioluminescence and chemiluminescence activities of these compounds have not been examined.
Lampteroflavin (Figure 14C) is a pentofuranosyl riboflavin compound isolated from O. japonicus that possesses green fluorescence (λmax = 524 nm) identical to the bioluminescence spectrum [113,114]. Minoru Isobe, a Japanese organic chemist, and his colleagues suggested that this compound is the light emitter of O. japonicus luminescence because it was the only fluorescent compound that showed an identical fluorescence spectrum to the in vivo bioluminescence detected in the lamellae of this mushroom [114]. Several chemicals, including flavins, exhibit chemiluminescence in Fenton’s reagent (Fe2+ and H2O2) [115], and Isobe showed that the chemiluminescence of lampteroflavin in Fenton’s reagent was significantly stronger than that of other flavins (lumiflavin, riboflavin, FMN, and FAD). Based on this result, Isobe expected that lampteroflavin would also be involved in the luminescence reaction itself [116]. On the other hand, O’Kane et al. suggested that flavin could not be the light emitter because luminous mushrooms are pigmented, and the ‘corrected’ bioluminescence spectrum of O. japonicus mycelia, which do not have the filter effect by pigmentation, did not match the fluorescence spectrum of flavin [117].
Nobel Prize winner Osamu Shimomura (1929–2018), a Japanese chemist/biochemist living in the USA, also contributed to research on fungal bioluminescence. He won the prize by finding the green fluorescent protein from luminous jellyfish Aequorea aequorea, and he also studied fungal bioluminescence using a Panellus stipticus; he was fascinated by this luminous mushroom, which appeared on an oak stump cut for building his new house at Falmouth in the 1980s [118]. He cultivated the fruitbody himself by applying his Shii-také (L. edodes) cultivation experience [119,120]. Using those cultivated P. stipticus mushrooms, Shimomura isolated panal, a sesquiterpene aldehyde, and its derivatives (Figure 14D), PS-A and PS-B, as possible candidates for luciferin precursors from the fruitbody [121,122,123]. After activation with primary amine, these panal derivatives exhibited chemiluminescence in the presence of Fe2+ and H2O2. The emission spectral peak of the chemiluminescence depends on the reaction conditions, some of which were close to the luminescence spectra in vivo. The same chemiluminescence was also observed for various other luminous fungi, such as Armillaria mycelia and Mycena fruitbodies [124]. Based on these results, Shimomura suggested that the bioluminescence of fungi might be a nonenzymatic reaction, which is inconsistent with Airth’s suggestion.
Other candidate compounds for luciferin or its precursor of fungal luminescence were isolated from the mycelia of M. citricolor by O. Shimomura (see Shimomura, 2006 [119]), and the chemical structure was analyzed by a Japanese organic chemist, Hideshi Nakamura (1952–2000). He suggested that all luciferin precursors contain a common catechol-derived group (4-(3,4-dihydroxyphenyl)-3-buten-2-carbonyl group) (Figure 14E) in their structures (personal communication from Dr. H. Nakamura, 1998, as referred to in Shimomura, 2006 [119]). To our surprise, this partial structure and UV absorption peak of the luciferin precursor (369 nm; see Shimomura, 2006 [119]) match those of hispidin, which we currently think is a true luciferin precursor of fungal bioluminescence [108].
6.5. Current Hispidin-Recycling Mechanism
The enzymatic reaction proposed by Airth’s group has been verified using several species of luminous fungal fruitbodies and mycelia, including the genera representing all four bioluminescent fungal lineages, Gerronema, Mycena, Armillaria, and Neonothopanus [125,126,127,128]. Oliveira et al. also examined cross-reactions between species, including the nonluminous fungi Filoboletus gracilis and Mycena singeri, and the results were all negative when nonluminous species were used for the assay [128]. Based on these results, they suggested that all luminous fungi share the same or similar luciferin and luciferase, whereas these components are absent in nonluminous species [128].
The luminescence mechanism of Japanese M. chlorophos was recently investigated by two independent Japanese research groups [129,130]. Mori et al. showed that Dubois’ classical luciferin–luciferase test was negative, bioluminescent components are insoluble in various surfactants, and the luminescence is heat unstable and thus will be an enzymatic reaction [129]. Hayashi et al. isolated an unknown compound that exhibited a UV absorption spectrum similar to that of flavins from the fruitbody, and the fluorescence peak almost matched both riboflavin and the bioluminescence of M. chlorophos [130]. Based on these findings, the authors concluded that this flavin-like compound is a factor in fungal bioluminescence. The bioluminescence or chemiluminescence activity of this compound was not examined in this report.
In 2015, we eventually identified the chemical structure of fungal luciferin and its precursor as 3-hydroxyhispidin and hispidin (Figure 14F), respectively, using the mycelium of the Vietnamese luminous fungus Neonothopanus nambi [108]. In 2017, we reported the whole mechanism of fungal bioluminescence by determining the chemical structure of the reaction product as caffeylpyruvic acid as a light emitter (Figure 14F) [131]. In 2017, we also determined the presence of hispidin as an active compound for the NADPH-dependent bioluminescence reaction in the fruitbodies of Japanese M. chlorophos, O. japonicus, and Brazilian N. gardneri [110]. The unopened ‘young’ fruitbody of M. chlorophos is nonluminescent, but it starts luminescence within seconds to the visible level when soaking with hispidin solution (Figure 15). This can be explained by the presence of luciferase but no luciferin precursor, hispidin, in the young fruitbody [110].
Katsunori Teranishi, a Japanese organic chemist, proposed a conflicting suggestion of light emitter compounds of luminescence in the M. chlorophos mushroom as riboflavin, riboflavin 5′-monophosphate, and/or flavin adenine dinucleotide [132]. He stated that 3-hydroxyhisidin can produce light by partially purified luciferase from M. chlorophos, but it remains unclear whether the compound actually produces light in the natural tissue of luminous fungi [133].
In 2018, we identified the luciferase gene, as well as luciferin-regenerating enzymes from N. nambi (Figure 14F); these genes were clustered on the fungal genome, and the molecular size of the luciferase (Luz) was approximately 28.5 kDa [50]. When these genes were transferred to yeast, the strain emitted the same green light. Currently, bioluminescent gene clusters have been identified in various luminous fungal species [50,51,73]. Furthermore, this fungal bioluminescence system is completely different from all known bioluminescent systems in other luminous organisms [134], suggesting that basic bioluminescence mechanisms are common in all luminous fungi; thus, bioluminescence in fungi has a single evolutionary origin [51]. In 2020, a self-luminescent plant was genetically produced using fungal luminescence genes [135]. Indeed, this bioengineering was realized because a common compound produced by all plants, caffeic acid (Figure 14F), is coincidentally a precursor of fungal luciferin. This finding has the potential to advance plant science as a novel tool for bioimaging technology. To date, no higher plant species with natural bioluminescence has ever been reported [23], but in the future, fungal luminescent genes may illuminate the city as glowing street trees [136].
7. Biological Function
For the biological function of fungal bioluminescence, many hypotheses have been proposed [137] but are not yet conclusive [138]. Arguably the most plausible hypothesis is that light attracts insects as a spore vector [139]. In Japan, we observed various animals visiting the fruitbodies of M. chlorophos (Figure 16) at night. Undescribed Japanese species Roridomyces sp. (Aya-hikari-také) has luminescent spores, suggesting the presence of a spore dispersal function by light-attracted animals [10]. Aposematism is another strong hypothesis [137]. However, for example, O. japonicus is toxic to humans, but many insect species visit and consume the fruitbody [140] (Figure 16).
8. Conclusions
In this review, we demonstrated the enthusiasm for luminous fungi in Japan, from old folklore and taxonomic surveys to current popular entertainment and life sciences. Luminous mushrooms activate ecotourism, and the biotechnology born from the science of luminous fungi has the potential to change our urban life.
Author Contributions
Conceptualization, Y.O.; writing—original draft preparation, Y.O. and K.H.; writing—review and editing, Y.O. and K.H.; visualization, Y.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially funded by JSPS KAKENHI grant number 20K06805 to KH.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank Nathaniel Guy for his help with editing and Japanese–English translations of specific terms; Masashi Naito and Yumiko Oba for their advice about the knowledge on bioluminescent mushrooms in Japan; and Takehito Miyatake, Yoshinori Nishino, and So Yamashita for kindly providing the photographs.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hosoya, T. History and the current status of fungal inventory and databasing in Japan. In Proceedings of the 7th and 8th Symposia on Collection Building and Natural History Studies in Asia and the Pacific Rim; Tomida, Y., Kubodera, T., Akiyama, S., Kitayama, T., Eds.; National Science Museum Monographs: Ibaraki, Japan, 2006; pp. 51–61. [Google Scholar]
- Sugahara, T. Japanese and mushrooms. J. Cook. Sci. Jpn. 2001, 34, 313–320. [Google Scholar]
- Guy, N. Kinoko: A Window into the Mystical World of Japanese Mushrooms; Amazon on Demand: Seattle, WA, USA, 2023; ISBN 979-8987537671. [Google Scholar]
- Nakano, J. Primer of Yamigaku (Study of Dark); Shueisha: Tokyo, Japan, 2014. (In Japanese) [Google Scholar]
- Komatsu, K. Introduction to Yokai Culture: Monsters, Ghosts, and Outsiders in Japanese History; Japan Publishing Industry Foundation for Culture: Tokyo, Japan, 2018. [Google Scholar]
- Yamaguchi, H. Occurrence of the luminous oligochæte Microscolex phosphoreus (Dug.). Annot. Zool. Japon 1935, 15, 200–202. [Google Scholar]
- Kanda, S. On the luminous earthworm Hikari-umi-mimizu. Rigakukai 1938, 36, 1–7. (In Japanese) [Google Scholar]
- Shinohara, K.; Higa, Y. New record of the luminous millipede from Okinawa. Edaphologia 1997, 59, 61–62, (In Japanese with English Abstract). [Google Scholar]
- Kashiwabara, S. An origin of firefly luminescence? Luminescence of Collembola. SCIaS 1997, 2, 10. (In Japanese) [Google Scholar]
- Kurogi, S. Mushrooms in Miyazaki Prefecture (Miyazaki no Kinoko); Komyakusha: Miyazaki, Japan, 2015. (In Japanese) [Google Scholar]
- Iwama, A. An unexpected finding of the luminescent Cruentomycena. Mycol. Circ. Jpn. 2020, 213, 2–3. (In Japanese) [Google Scholar]
- Harvey, E.N. A History of Luminescence. From the Earliest Times Until 1900; American Philosophical Society: Philadelphia, PA, USA, 1957. [Google Scholar]
- Watasé, S. On the Fireflies (Hotaru no Hanashi); Kaiseikan: Tokyo, Japan, 1902. (In Japanese) [Google Scholar]
- Kanda, S. Shiranui, Hitodama, and Kitsune-bi; Shunyodo: Tokyo, Japan, 1931. (In Japanese) [Google Scholar]
- Togawa, A. Haguro-san Yobanashi (or Haguro-san Yawa). Tabi Densetsu 1944, 193, 13–24. (In Japanese) [Google Scholar]
- Sakai, T. Nankou Chawa. In Nihon Zuihitsu Taisei; Yoshikawakobunkan: Tokyo, Japan, 1977; pp. 413–448. (In Japanese) [Google Scholar]
- Takahashi, T. Yamagata-ken ni okeru Torimono no Keifu. Seikou Minzoku 1991, 135, 36–37. (In Japanese) [Google Scholar]
- Hiroi, M. On the luminescence of mushrooms. Kinoko-ken Dayori 2006, 28, 10–20. (In Japanese) [Google Scholar]
- Hasegawa, E.; Ota, Y.; Hattori, T.; Sahashi, N.; Kikuchi, T. Ecology of Armillaria species on conifers in Japan. For. Path. 2011, 41, 429–437. [Google Scholar] [CrossRef]
- Dan, I. The Smoke of a Pipe, Still; Asahi Shimbun Co.: Tokyo, Japan, 1972. (In Japanese) [Google Scholar]
- Nakajima, A. Kinoko Zukan; Natsume Pub.: Tokyo, Japan, 2017. (In Japanese) [Google Scholar]
- Nishino, Y.; Oba, Y. Luminous Mushrooms with Night Forests; Iwanami Shoten Pub.: Tokyo, Japan, 2013. (In Japanese) [Google Scholar]
- Oba, Y. Luminous Organisms of the World: Diversity, Ecology, and Biochemistry; The University of Nagoya Press: Nagoya, Japan, 2022. (In Japanese) [Google Scholar]
- Harvey, E.N. Bioluminescence; Academic Press: New York, NY, USA, 1952. [Google Scholar]
- Keene, D. The Tale of the Bamboo Cutter; Kodansha International: Tokyo, Japan, 1998. [Google Scholar]
- Hotate, M. Shining Princess and the Sovereign Myth (Kaguya-himé to Ouken-shinwa); Yosensha: Tokyo, Japan, 2010. (In Japanese) [Google Scholar]
- Masuda, K.; Suzuki, H. The origins of fiction. Kokubungaku 1993, 38, 6–24. (In Japanese) [Google Scholar]
- Yasuda, A. Miscellaneous notes on fungi. Bot. Mag. Tokyo 1920, 34, 194–196. (In Japanese) [Google Scholar]
- Shidei, T. Report of fungal flora in bamboo forest. Trans. Mycol. Soc. Jpn. 1974, 15, 251–257. (In Japanese) [Google Scholar]
- Haneda, Y. Luminous Organisms; Kouseisha-Kouseikaku: Tokyo, Japan, 1985. (In Japanese) [Google Scholar]
- Imazeki, R.; Otani, Y.; Hongo, T. Mushrooms in Japan; Yama-Kei Pub.: Tokyo, Japan, 1988. (In Japanese) [Google Scholar]
- Kyoto Prefecture. The List of Biological and Geological Components of the Natural Environment 2015, Red Data Book of Kyoto Prefecture 2015, Supplementary; Kyoto Prefecture: Kyoto, Japan, 2015. (In Japanese) [Google Scholar]
- Kanda, S. Fireflies; Nippon Hakkō Seibutsu Kenkyu Kai: Tokyo, Japan, 1935. (In Japanese) [Google Scholar]
- Ohba, N. Mystery of Fireflies; Yokosuka City Mus.: Yokosuka, Japan, 2004. (In Japanese) [Google Scholar]
- Oba, Y.; Miyatake, T. Luminous Creatures, Mushrooms; Kumon Pub: Tokyo, Japan, 2015. (In Japanese) [Google Scholar]
- Miyatake, T. Luminous mushrooms (World of Wonders); Fukuinkan Shoten: Tokyo, Japan, 2023. (In Japanese) [Google Scholar]
- Niitsu, H.; Hanyuda, N.; Sugiyama, Y. Cultural properties of a luminous mushroom, Mycena chlorophos. Mycoscience 2000, 41, 551–558. [Google Scholar] [CrossRef]
- Niitsu, H.; Hanyuda, N. Fruit-body production of a luminous mushroom, Mycena chlorophos. Mycoscience 2000, 41, 559–564. [Google Scholar] [CrossRef]
- Ise Shimbun. Enjoy Luminous Mushroom. Iwade Research Institute of Mycology Group in Tsu City Sells the Culture Kit; The Ise Shimbun Chusei Local: Tsu, Japan, 24 May 2021. (In Japanese) [Google Scholar]
- Neda, H. Kinoko Museum; Yasaka Shobo: Tokyo, Japan, 2014. (In Japanese) [Google Scholar]
- Kawamura, S. Studies on the luminous fungus, Pleurotus japonicus sp. nov. J. Coll. Sci. Tokyo Imp. Univ. 1915, 35, 1–29. [Google Scholar]
- Kobayasi, Y. Seven luminous mycomycetes from Bonin Islands. Bull. Biogeographic. Soc. Jpn. 1937, 7, 1–7. [Google Scholar]
- Corner, E.J.H. Further description of luminous agarics. Trans. Br. Mycol. Soc. 1954, 37, 256–271. [Google Scholar] [CrossRef]
- Corner, E.J.H. The Marquis: A Tale of Syonan-to; Heinemann Asia: Kuala Lumpur, Singapore, 1981. [Google Scholar]
- Wassink, E.C. On Fungus Luminescence; Mededelingen Landbouwhogeschool; Wageningen, H. Veenman & Zonen: Wageningen, The Netherlands, 1979. [Google Scholar]
- Desjardin, D.E.; Oliveira, A.G.; Stevani, C.V. Fungi bioluminescence revisited. Photochem. Photobiol. Sci. 2008, 7, 170–182. [Google Scholar] [CrossRef]
- Terashima, Y.; Takahashi, H.; Taneyama, Y. The Fungal Flora in Southwestern Japan: Agarics and Boletes; Tokai University Press: Kanagawa, Japan, 2016. [Google Scholar]
- Cortés-Pérez, A.; Desjardin, D.E.; Perry, B.A.; Ramírez-Cruz, V.; Ramírez-Guillén, F.; Villalobos-Arámbula, A.R.; Rockefeller, A. New species and records of bioluminescent Mycena from Mexico. Mycologia 2019, 111, 319–338. [Google Scholar] [CrossRef]
- Chang, C.-C.; Chen, C.-Y.; Lin, W.-W.; Kao, H.-W. Mycena jingyinga, Mycena luguensis, and Mycena venus: Three new species of bioluminescent fungi from Taiwan. Taiwania 2020, 65, 396–406. [Google Scholar]
- Kotlobay, A.A.; Sarkisyan, K.S.; Mokrushina, Y.A.; Marcet-Houben, M.; Serebrovskaya, E.O.; Markina, N.M.; Somermeyer, L.G.; Gorokhovatsky, A.Y.; Vvedensky, A.; Purtov, K.V.; et al. Genetically encodable bioluminescent system from fungi. Proc. Natl. Acad. Sci. USA 2018, 115, 12728–12732. [Google Scholar] [CrossRef] [PubMed]
- Ke, H.–M.; Lee, H.–H.; Lin, C.–Y.I.; Liu, Y.–C.; Lu, M.R.; Hsieh, J.–W.H.; Chang, C.–C.; Wu, P.–H.; Lu, M.J.; Li, J.–Y.; et al. Mycena genomes resolve the evolution of fungal bioluminescence. Proc. Natl. Acad. Sci. USA 2020, 117, 31267–31277. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, N. On the report about Japanese poisonous mushrooms part 1 by Kichindo Inoko. J. Plant Res. 1889, 3, 47–51. (In Japanese) [Google Scholar]
- Haneda, Y. Luminous Organisms of Japan and the Far East. In The Luminescence of Biological Systems; Johnson, F.H., Ed.; American Association for the Advancement of Science: Washington, DC, USA, 1955; pp. 335–385. [Google Scholar]
- Chew, A.L.C.; Tan, Y.-S.; Desjardin, D.E.; Musa, M.Y.; Sabaratnam, V. Taxonomic and phylogenetic re-evaluation of Mycena illuminans. Mycologia 2013, 105, 1325–1335. [Google Scholar] [CrossRef] [PubMed]
- Katumoto, K. List of Fungi Recorded in Japan; The Kanto Branch of the Mycological Society of Japan: Tokyo, Japan, 2010. (In Japanese) [Google Scholar]
- Takahashi, H. An impact on the finding of the luminescent Cruentomycena by Ami Iwama. Mycol. Circ. Jpn. 2020, 213, 4. (In Japanese) [Google Scholar]
- Taneyama, Y. Cruentomycena was bioluminescent! Mycol. Circ. Jpn. 2020, 213, 5. (In Japanese) [Google Scholar]
- Okuzawa, Y.; Okuzawa, M. Mushroom Etymology and Dialect Dictionary; Yama-Kei Pub.: Tokyo, Japan, 1998. (In Japanese) [Google Scholar]
- Kobayasi, Y. Contributions to the luminous fungi from Japan. J. Hattori Bot. Lab. 1951, 5, 1–6. [Google Scholar]
- GBIF.org. GBIF Home Page. 2023. Available online: https://www.gbif.org (accessed on 19 April 2023).
- Kawai, D. Handbook of Mushrooms in Oirase Stream (Oirase Keiryu, Kinoko Handbook), Spring—Early Summer; NPO Oiken: Aomori, Japan, 2022. (In Japanese) [Google Scholar]
- Imazeki, R.; Hongo, T. Colored Illustrations of Mushrooms of Japan, Vol. I.; Hoikusha: Osaka, Japan, 1987. (In Japanese) [Google Scholar]
- Search System of Japanese Red Data; Association of Wildlife Research and EnVision. Available online: http://jpnrdb.com/index.html (accessed on 15 April 2023).
- Tanaka, Y.; Kasuga, Y.; Oba, Y.; Hase, S.; Sakakibara, Y. Genome sequence of the luminous mushroom Mycena chlorophos for searching fungal bioluminescence genes. Luminescence 2014, 29, 47–48. [Google Scholar]
- Miyagi, G. Notes on luminous fungi, Filoboletus manipularis, on Okinawa. Bull. Coll. Sci. Univ. Ryukyus 1960, 4, 77–87. (In Japanese) [Google Scholar]
- Hioki, K. (Ed.) Sanshu Kidan; Ishikawa Prefecture Library Association: Ishikawa, Japan, 1933. (In Japanese) [Google Scholar]
- Kawamura, S. Studies on a luminous fungus, Pleurotus japonicus sp. nov. Bot. Mag. Tokyo 1910, 24, 165–177. [Google Scholar] [CrossRef]
- Kudo, S. Macrofungi of Aomori; Access 21 Pub.: Aomori, Japan, 2017. (In Japanese) [Google Scholar]
- Dykstra, Y.K. (Translator). The Konjaku Tales. Japanese section (Honcho-Hen)(III) from a Medieval Japanese Collection; Kansai Gaidai Univ. Pub.: Osaka, Japan, 2003. [Google Scholar]
- Watari, T.; Tachibana, T.; Okada, A.; Nishikawa, K.; Otsuki, K.; Nagai, N.; Abe, H.; Nakano, Y.; Takagi, S.; Amano, Y. A review of food poisoning caused by local food in Japan. J. Gen. Fam. Med. 2021, 22, 15–23. [Google Scholar] [CrossRef]
- Nakanishi, K.; Ohashi, M.; Tada, M.; Yamada, Y. Illudin S (lampterol). Tetrahedron 1965, 21, 1231–1246. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Shirahama, H.; Ichihara, A.; Fukuoka, Y.; Takahashi, Y.; Mori, Y.; Watanabe, M. Structure of lampterol (illudin S). Tetrahedron 1965, 21, 2671–2676. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Park, M.-J.; Shim, D.; Ryoo, R. De novo genome assembly of the bioluminescent mushroom Omphalotus guepiniiformis reveals an Omphalotus-species lineage of the luciferase. Genomics 2022, 114, 110514. [Google Scholar] [CrossRef] [PubMed]
- Fukasawa, Y.; Osono, T.; Takeda, H. Beech log decomposition by wood-inhabiting fungi in a cool temperate forest floor: A quantitative analysis focused on the decay activity of a dominant basidiomycete Omphalotus guepiniformis. Ecol. Res. 2010, 25, 959–966. [Google Scholar] [CrossRef]
- Ooka, M. Easy-Access Index of Haiku Seasonal Words (Autumn); Yushikan: Tokyo, Japan, 2005. (In Japanese) [Google Scholar]
- Wassink, E.C. Luminescence in Fungi. In Bioluminescence in Action; Herring, P.J., Ed.; Academic Press: New York, NY, USA, 1978; pp. 171–197. [Google Scholar]
- Kudo, S.; Tezuka, Y.; Yonaiyama, H. Fungi of Aomori; Graph Aomori: Aomori, Japan, 1998. (In Japanese) [Google Scholar]
- Hiroi, M. Light emission of Armillaria tabescens (Scop.:Fr.) Emel–Measurement of light emission of mushrooms using chemiluminescence detector–. Bull. Koriyama Women’s Univ. 2013, 49, 179–189. [Google Scholar]
- Koch, R.A.; Wilson, A.W.; Séne, O.; Henkel, T.W.; Aime, M.C. Resolved phylogeny and biogeography of the root pathogen Armillaria and its gasteroid relative, Gyanogaster. BMC Evol. Biol. 2017, 17, 33. [Google Scholar] [CrossRef]
- Moncalvo, J.-M.; Lutzoni, F.M.; Rehner, S.A.; Johnson, J.; Vilgalys, R. Phylogenetic relationships of agaric fungi based on nuclear large subunit ribosomal DNA sequences. Syst. Biol. 2000, 49, 278–305. [Google Scholar] [CrossRef]
- Macrae, R. Interfertility studies and inheritance of luminosity in Panus stypticus. Can. J. Res. 1942, 20c, 411–434. [Google Scholar] [CrossRef]
- Dickinson, C.; Lucas, J. (Eds.) . The Encyclopedia of Mushrooms; G. P. Putnam’s Sons: New York, NY, USA, 1979. [Google Scholar]
- Berkeley, M.J.; Curtis, M.A. Characters of new fungi, collected in the North Pacific Exploring Expedition by Charles Wright. Proc. Amer. Acad. Arts Sci. 1860, 4, 111–130. [Google Scholar]
- Ito, S. Mycological Flora of Japan. Vol. II. Basidiomycetes No. 5. Agaricales, Gasteromycetales; Yokendo: Tokyo, Japan, 1959. (In Japanese) [Google Scholar]
- Desjardin, D.E.; Capelari, M.; Stevani, C.V. A new bioluminescent agaric form São Paulo, Brazil. Fung. Div. 2005, 18, 9–14. [Google Scholar]
- Bothe, F. Über das Leuchten verwesender Blätter und seine Erreger. Planta 1931, 14, 752–765. (In Germany) [Google Scholar] [CrossRef]
- Berliner, M.D. Studies in fungal luminescence. Mycologia 1961, 53, 84–90. [Google Scholar] [CrossRef]
- Treu, R.; Agerer, R. Culture characteristics of some Mycena species. Mycotaxon 1990, 38, 279–309. [Google Scholar]
- Bermudes, D.; Petersen, R.H.; Nealson, K.H. Low-level bioluminescence detected in Mycena haematopus basidocarps. Mycologia 1992, 84, 799–802. [Google Scholar] [CrossRef]
- Josserand, M. Sur la luminescence de “Mycena rorida” en Europe occidentale. Bull. Mens. Soc. Linn. Lyon 1953, 22, 99–102. [Google Scholar] [CrossRef]
- Neda, H.; Sato, H. New record of Armillaria fuscipes Petch from Japan. In Abstract of Papers Presented at the 53th Annual Meeting of the Mycological Society of Japan; The Mycological Society of Japan: Tokyo, Japan, 2009; p. 48. (In Japanese) [Google Scholar]
- Mihail, J.D. Bioluminescence patterns among North American Armillaria species. Fungal Biol. 2015, 119, 528–537. [Google Scholar] [CrossRef]
- Ainsworth, M. Searching for luminous mushrooms of the marsh fungus Armillaria ectypa. Field Mycol. 2004, 5, 142–144. [Google Scholar] [CrossRef]
- Wassink, E.C. Observations on the luminescence in fungi, I, including a critical review of the species mentioned as luminescent in literature. Recueil Trav. Bot. Neerl. 1948, 41, 150–212. [Google Scholar]
- Hongo, T. New taxa of the Agaricales published by T. Hongo from 1973 to 1988. Trans. Mycol. Soc. Jpn. 1989, 30, 499–504. [Google Scholar]
- Harder, C.B.; Laæssøe, T.; Frøslev, T.G.; Ekelund, F.; Rosendahl, S.; Kjøller, R. A three-gene phylogeny of the Mycena pura complex reveals 11 phylogenetic species and shows ITS to be unreliable for species identification. Fungal Biol. 2013, 117, 764–775. [Google Scholar] [CrossRef] [PubMed]
- Ota, Y.; Kim, M.-S.; Neda, H.; Klopfenstein, N.B.; Hasegawa, E. The phylogenetic position of an Armillaria species from Amami-Oshima, a subtropical island of Japan, based on elongation factor and ITS sequences. Mycoscience 2011, 52, 53–58. [Google Scholar] [CrossRef]
- Boyle, R. Observations and tryals about the resemblances and differences between a burning coal and shining wood. Philos. Trans. Royal Soc. 1668, 2, 605–612. [Google Scholar]
- Fabre, J. –H. Recherches sur la cause de la phosphorescence de l’agaric de l’olivier. Ann. Sci. Nat. Bot. 1855, 4, 179–197. (In French) [Google Scholar]
- Goto, Y. Luminous organisms appeared in Edo-period literatures. Bull. Firefly Mus. Toyota Town 2022, 14, 81–86. (In Japanese) [Google Scholar]
- Kawamura, S. Studies on a Luminous fungus, Pleurotus japonicus sp. nov. (Continued from p. 213). Bot. Mag. Tokyo 1910, 24, 249–260. [Google Scholar] [CrossRef]
- Airth, R.L.; McElroy, W.D. Light emission from extracts of luminous fungi. J. Bacteriol. 1959, 77, 249–250. [Google Scholar] [CrossRef]
- Airth, R.L.; Foerster, G.E.; Behrens, P.Q. The luminous fungi. In Bioluminescence in Progress; Johnson, F.H., Haneda, Y., Eds.; Princeton University Press: Princeton, NJ, USA, 1966; pp. 203–223. [Google Scholar]
- Airth, R.L.; Foerster, G.E.; Hinde, R. Bioluminescence. In Photobiology of Microorganisms; Halldal, P., Ed.; John Wiley & Sons: London, UK, 1970; pp. 417–446. [Google Scholar]
- Airth, R.L.; Foerster, G.E. The isolation of catalytic components required for cell-free fungal bioluminescence. Arch. Biochem. Biophys. 1962, 97, 567–573. [Google Scholar] [CrossRef]
- Airth, R.L.; Foerster, G.E. Enzymes associated with bioluminescence in Panus stypticus luminescence and Panus stypticus non-luminescence. J. Bacteriol. 1964, 88, 1372–1379. [Google Scholar] [CrossRef]
- Oba, Y. Bioluminescent mechanism of luminous mushroom. Kinoko-ken Dayori 2017, 40, 18–25. (In Japanese) [Google Scholar]
- Purtov, K.V.; Petushkov, V.N.; Baranov, M.S.; Mineev, K.S.; Rodionova, N.S.; Kaskova, Z.M.; Tsarkova, A.S.; Petunin, A.I.; Bondar, V.S.; Rodicheva, E.K.; et al. The chemical basis of fungal bioluminescence. Angew. Chem. Int. Ed. 2015, 54, 8124–8128. [Google Scholar] [CrossRef] [PubMed]
- Kuwabara, S.; Wassink, E.C. Purification and properties of the active substance of fungal luminescence. In Bioluminescence in Progress; Johnson, F.H., Haneda, Y., Eds.; Princeton University Press: Princeton, NJ, USA, 1966; pp. 233–245. [Google Scholar]
- Oba, Y.; Suzuki, Y.; Martins, G.N.R.; Carvalho, R.P.; Pereira, T.A.; Waldenmaier, H.E.; Kanie, S.; Naito, M.; Oliveira, A.G.; Dörr, F.A.; et al. Identification of hispidin as a bioluminescent active compound and its recycling biosynthesis in the luminous fungal fruiting body. Photochem. Photobiol. Sci. 2017, 16, 1435–1440. [Google Scholar] [CrossRef]
- Kuwabara, S.; Cormier, M.J.; Dure, L.S.; Kreiss, P.; Pfuderer, P. Crystalline bacterial luciferase from Photobacterium fischeri. Proc. Natl. Acad. Sci. USA 1965, 53, 822–828. [Google Scholar] [CrossRef]
- Endo, M.; Kajiwara, M.; Nakanishi, K. Fluorescent constituents and cultivation of Lampteromyces japonicus. Chem. Commun. 1970, 309–310. [Google Scholar] [CrossRef]
- Isobe, M.; Uyakul, D.; Goto, T. Lampteromyces bioluminescence-2. Lampteroflavin, a light emitter in the luminous mushroom, L. japonicus. Tetrahedron Lett. 1988, 29, 1169–1172. [Google Scholar] [CrossRef]
- Uyakul, D.; Isobe, M.; Goto, T. Lampteromyces bioluminescence 3. Structure of lampteroflavin, the light emitter in the luminous mushroom, L. japonicus. Bioorg. Chem. 1989, 17, 454–460. [Google Scholar] [CrossRef]
- Zeng, J.; Jewsbury, R.A. Chemiluminescence of flavins in the presence of Fe(II). J. Photochem. Photobiol. 1995, A91, 117–120. [Google Scholar] [CrossRef]
- Isobe, M. New bioluminescent systems in fungi, plant and animals. Nippon. Nōgeikagaku Kaishi 1992, 66, 736–741. (In Japanese) [Google Scholar] [CrossRef]
- O’Kane, D.J.; Fuhrer, B.; Lingle, W.L. Spectral studies on fungal bioluminescence. In Bioluminescence and Chemiluminescence: Fundamentals and Applied Aspects; Campbell, A.K., Ed.; John Wiley & Sons: Chichester, UK, 1994; pp. 552–555. [Google Scholar]
- Shimomura, O. Learning from Jellyfish (Kurage ni Manabu); Nagasaki Bunken Sha: Nagasaki, Japan, 2010. (In Japanese) [Google Scholar]
- Shimomura, O. Bioluminescence: Chemical Principles and Methods; World Scientific: Toh Tuck Link, Singapore, 2006. [Google Scholar]
- Shimomura, O.; Shimomura, S.; Bringer, J.H. Luminous Pursuit: Jellyfish, GFP, and the Unforeseen Path to the Nobel Prize; World Scientific: Toh Tuck Link, Singapore, 2017. [Google Scholar]
- Nakamura, H.; Kishi, Y.; Shimomura, O. Panal: A possible precursor of fungal luciferin. Tetrahedron 1988, 44, 1597–1602. [Google Scholar] [CrossRef]
- Shimomura, O. Superoxide-triggered chemiluminescence of the extract of luminous mushroom Panellus stipticus after treatment with methylamine. J. Exp. Bot. 1991, 42, 555–560. [Google Scholar] [CrossRef]
- Shimomura, O. Structure and non-enzymatic light emission of two luciferin precursors isolated from the luminous mushroom Panellus stipticus. J. Biolumin. Chemilumin. 1993, 8, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, O. The role of superoxide dismutase in regulating the light emission of luminescent fungi. J. Exp. Bot. 1992, 43, 1519–1525. [Google Scholar] [CrossRef]
- Kamzolkina, O.V.; Danilov, V.S.; Egorov, N.S. On the nature of luciferase from the bioluminescent fungus Armillariella mellea. Dokl. Akad. Nauk. SSSR 1983, 3, 750–752. (In Russian) [Google Scholar]
- Kamzolkina, O.V.; Bekker, Z.E.; Egorov, N.S. Luciferin-luciferase system of the mushroom Armillariella mellea. Biol. Nauk. 1984, 1, 73–77. (In Russian) [Google Scholar]
- Oliveira, A.G.; Stevani, C.V. The enzymatic nature of fungal bioluminescence. Photochem. Photobiol. Sci. 2009, 8, 1416–1421. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.G.; Desjardin, D.E.; Perry, B.A.; Stevani, C.V. Evidence that a single bioluminescent system is shared by all known bioluminescent fugal lineages. Photochem. Photobiol. Sci. 2012, 11, 848–852. [Google Scholar] [CrossRef]
- Mori, K.; Kojima, S.; Maki, S.; Hirano, T.; Niwa, H. Bioluminescence characteristics of the fruiting body of Mycena chlorophos. Luminescence 2011, 26, 604–610. [Google Scholar] [CrossRef]
- Hayashi, S.; Fukushima, R.; Wada, N. Extraction and purification of a luminiferous substance from the luminous mushroom Mycena chlorophos. Biophysics 2012, 8, 111–114. [Google Scholar] [CrossRef]
- Kaskova, Z.M.; Dörr, F.A.; Petushkov, V.N.; Purtov, K.V.; Tsarkova, A.S.; Rodionova, N.S.; Mineev, K.S.; Guglya, E.B.; Kotlobay, A.; Baleeva, N.S.; et al. Mechanism and color modulation of fungal bioluminescence. Sci. Adv. 2017, 3, e1602847. [Google Scholar] [CrossRef]
- Teranishi, K. Identification of possible light emitters in the gills of a bioluminescent fungus Mycena chlorophos. Luminescence 2016, 31, 1407–1413. [Google Scholar] [CrossRef] [PubMed]
- Teranishi, K. Bioluminescence and chemiluminescence abilities of trans-3-hydroxyhispidin on the luminous fungus Mycena chlorophos. Luminescence 2018, 33, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Wilson, T.; Hastings, J.W. Bioluminescence. Annu. Rev. Cell Dev. Biol. 1998, 14, 197–230. [Google Scholar] [CrossRef]
- Mitiouchkina, T.; Mishin, A.S.; Somermeyer, L.G.; Markina, N.M.; Chepurnyh, T.V.; Guglya, E.B.; Karataeva, T.A.; Palkina, K.A.; Shakhova, E.S.; Fakhranurova, L.I.; et al. Plants with genetically encoded autoluminescence. Nat. Biotechnol. 2020, 38, 944–946. [Google Scholar] [CrossRef]
- Strack, R. Harnessing fungal bioluminescence (research highlights). Nat. Methods 2019, 16, 139–145. [Google Scholar]
- Sivinski, J. Arthropods attracted to luminous fungi. Psyche 1981, 88, 383–390. [Google Scholar] [CrossRef]
- Weinstein, P.; Delean, S.; Wood, T.; Austin, D. Bioluminescence in the ghost fungus Omphalotus nidiformis does not attract potential spore dispersing insects. IMA Fungus 2016, 7, 229–234. [Google Scholar] [CrossRef]
- Oliveira, A.G.; Stevani, C.V.; Waldenmaier, H.E.; Viviani, V.; Emerson, J.M.; Loros, J.J.; Dunlap, J.C. Circadian control sheds light on fungal bioluminescence. Curr. Biol. 2015, 25, 964–968. [Google Scholar] [CrossRef]
- Makihara, H.; Takeno, K.; Chûjô, M. Check list of the insects collected from Lampteromyces japonicus (Kawam.) Sing. on Mt. Hiko. Sci. Bull. Fac. Agr. Kyushu Univ. 1972, 26, 595–600, (In Japanese with English Title). [Google Scholar]
Figure 1.
(A) Luminous mushroom stamps. (From left to right) Mycena chlorophos/West Samoa, 1985; Mycena manipularis/Việt Nam, 1996; Mycena lucentipes/USA, 2018. There are no luminous mushroom stamps in other countries, including Japan. (B) Foxfire at Ōji (Ōji Shōzoku Enoki Ōmisoka no Kitsune-bi) by Hiroshige Utagawa (recarved edition, original print in 1857). (C) Pokémon cards. English names (from left to right): Morelull (basic), Shinotic (stage 1), and Glimwood Tangle (stadium), produced by The Pokémon Company (Tokyo, Japan). (D) Capsule toys: “luminous mushroom magnet” series (2015-) of eight Japanese luminous mushroom species, produced by Ikimon Co. (Tokyo, Japan). The diameter of the model of Mycena chlorophos (center) is approximately 30 mm, which is close in size to the largest specimens found in the wild. (E) Picture books of luminous mushrooms. (From left to right) Nishino & Oba, 2013; Oba & Miyatake, 2015; Miyatake, 2023. All are Yuichi Oba’s personal collections.
Figure 1.
(A) Luminous mushroom stamps. (From left to right) Mycena chlorophos/West Samoa, 1985; Mycena manipularis/Việt Nam, 1996; Mycena lucentipes/USA, 2018. There are no luminous mushroom stamps in other countries, including Japan. (B) Foxfire at Ōji (Ōji Shōzoku Enoki Ōmisoka no Kitsune-bi) by Hiroshige Utagawa (recarved edition, original print in 1857). (C) Pokémon cards. English names (from left to right): Morelull (basic), Shinotic (stage 1), and Glimwood Tangle (stadium), produced by The Pokémon Company (Tokyo, Japan). (D) Capsule toys: “luminous mushroom magnet” series (2015-) of eight Japanese luminous mushroom species, produced by Ikimon Co. (Tokyo, Japan). The diameter of the model of Mycena chlorophos (center) is approximately 30 mm, which is close in size to the largest specimens found in the wild. (E) Picture books of luminous mushrooms. (From left to right) Nishino & Oba, 2013; Oba & Miyatake, 2015; Miyatake, 2023. All are Yuichi Oba’s personal collections.

Figure 2.
Mycelia of Mycena stellaris growing on bamboo. Photo by Yoshinori Nishino on Ishigaki Isl., Okinawa Prefecture.
Figure 2.
Mycelia of Mycena stellaris growing on bamboo. Photo by Yoshinori Nishino on Ishigaki Isl., Okinawa Prefecture.

Figure 3.
Fruitbody of Favolaschia peziziformis. Photo by So Yamashita on Hachijo Island, Tokyo.

Figure 4.
Fruitbody of Mycena chlorophos. Photo by So Yamashita on Hachijo Island, Tokyo.

Figure 5.
Fruitbody of Mycena lazulina. Photo by Yoshinori Nishino on Iriomote Island, Okinawa Prefecture.
Figure 5.
Fruitbody of Mycena lazulina. Photo by Yoshinori Nishino on Iriomote Island, Okinawa Prefecture.

Figure 6.
Fruitbody of Mycena lux-coeli. Photo by So Yamashita on Hachijo Island, Tokyo.

Figure 7.
Fruitbody of Mycena manipularis. Photo by Yoshinori Nishino on Ishigaki Island, Okinawa Prefecture.
Figure 7.
Fruitbody of Mycena manipularis. Photo by Yoshinori Nishino on Ishigaki Island, Okinawa Prefecture.

Figure 8.
Fruitbody of Mycena stellaris. Photo by Yoshinori Nishino at Kunigami, Okinawa Isl., Okinawa Prefecture.
Figure 8.
Fruitbody of Mycena stellaris. Photo by Yoshinori Nishino at Kunigami, Okinawa Isl., Okinawa Prefecture.

Figure 9.
Fruitbody of Panellus pusillus. Photo by So Yamashita on Hachijo Island, Tokyo.

Figure 10.
Fruitbody of Resiomycena fulgens. Photo by Takehito Miyatake on Hachijo Island, Tokyo.

Figure 11.
Fruitbody of Omphalotus japonicus. Photo by Yoshinori Nishino at Mt. Odaigahara, Nara Prefecture.
Figure 11.
Fruitbody of Omphalotus japonicus. Photo by Yoshinori Nishino at Mt. Odaigahara, Nara Prefecture.

Figure 12.
Rhizomorphs of Armillaria sp. Photo by So Yamashita on Hachijo Island, Tokyo.

Figure 13.
Pleurotus nitidus. Photo by Yoshinori Nishino on Ishigaki Island, Okinawa Prefecture.

Figure 14.
Chemical structure of luciferin candidates. (A) Illudin S, (B) ergosta-4,6,8(14),22-tetraen-3-on, and (C) lampteroflavin from Omphalotus japonicus. (D) Panal from Panellus stipticus. (E) A common partial structure of the luciferin precursors from Mycena citricolor. (F) Scheme of hispidin-recycling reaction determined in Neonothopanus nambi. Four enzymes involved in the cycle are shown in bold: H3H, hispidin-3-hydroxylase; Luz, fungal luciferase; CPH, caffeylpyruvate hydrolase; and HispS, hispidin synthase.
Figure 14.
Chemical structure of luciferin candidates. (A) Illudin S, (B) ergosta-4,6,8(14),22-tetraen-3-on, and (C) lampteroflavin from Omphalotus japonicus. (D) Panal from Panellus stipticus. (E) A common partial structure of the luciferin precursors from Mycena citricolor. (F) Scheme of hispidin-recycling reaction determined in Neonothopanus nambi. Four enzymes involved in the cycle are shown in bold: H3H, hispidin-3-hydroxylase; Luz, fungal luciferase; CPH, caffeylpyruvate hydrolase; and HispS, hispidin synthase.

Figure 15.
Chemical induction of the luminescence of the unopened young Mycena chlorophos fruitbody (5 mm diameter) by a buffer with and without 220 μM hispidin (left and right). Photos were taken 10 s after treatment.
Figure 15.
Chemical induction of the luminescence of the unopened young Mycena chlorophos fruitbody (5 mm diameter) by a buffer with and without 220 μM hispidin (left and right). Photos were taken 10 s after treatment.

Figure 16.
Fruitbodies of Mycena chlorophos with collembolas gathering under the gills (A) and Omphalotus japonicus with millipede and land snail grazing (B). Photos by Yoshinori Nishino on Bonin Isls., Tokyo (A) and by Takehito Miyatake at Tokushima Prefecture (B).
Figure 16.
Fruitbodies of Mycena chlorophos with collembolas gathering under the gills (A) and Omphalotus japonicus with millipede and land snail grazing (B). Photos by Yoshinori Nishino on Bonin Isls., Tokyo (A) and by Takehito Miyatake at Tokushima Prefecture (B).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
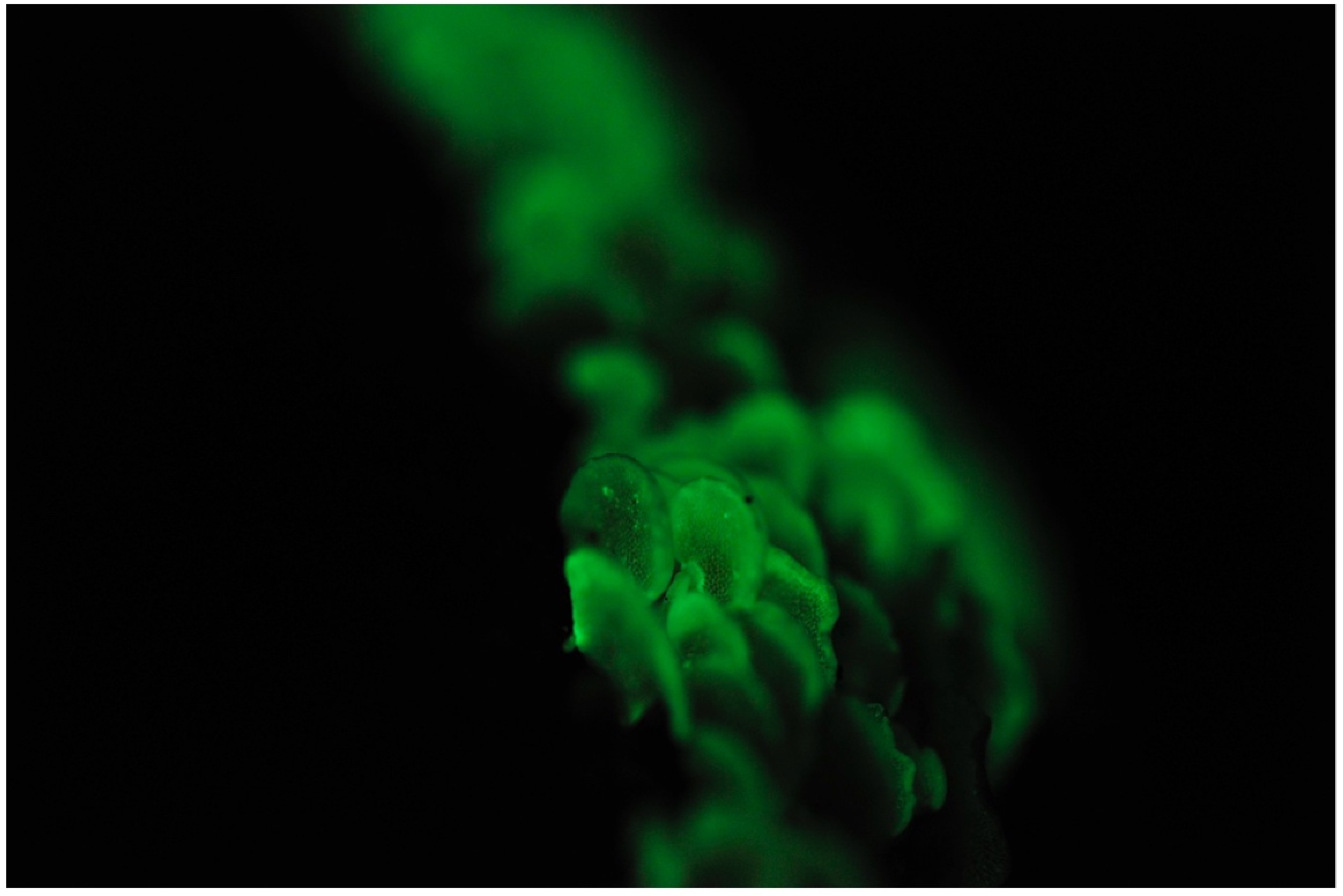{kind=link}
{kind=link}
{kind=link}
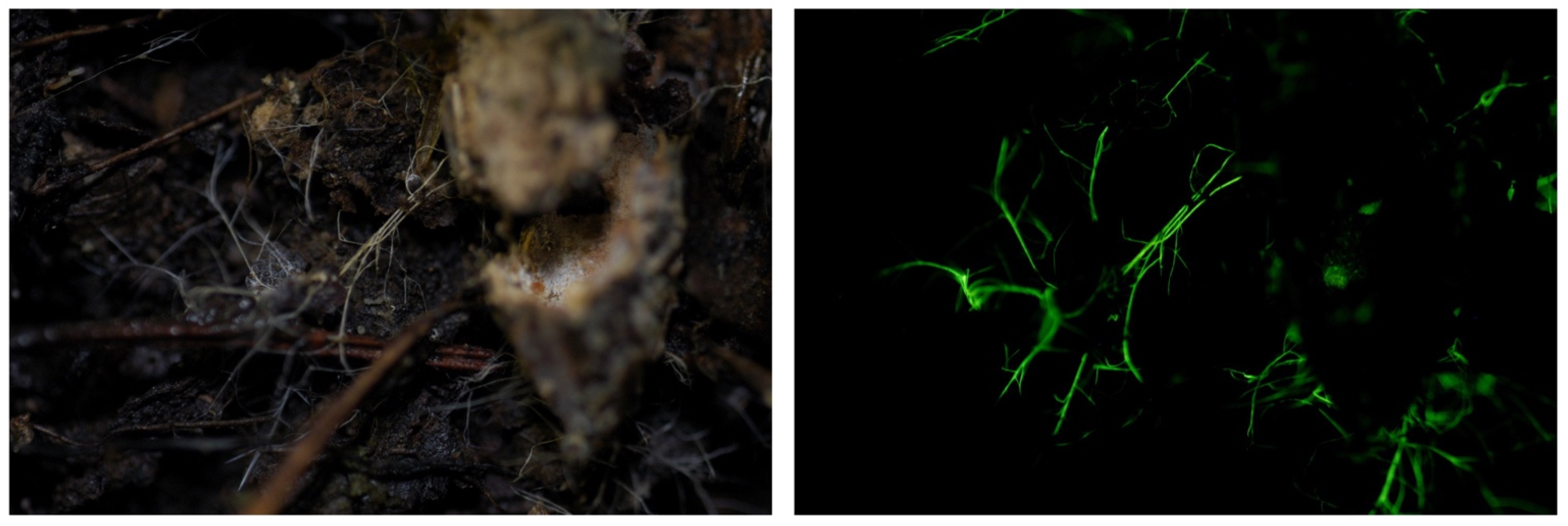{kind=link}
{kind=link}
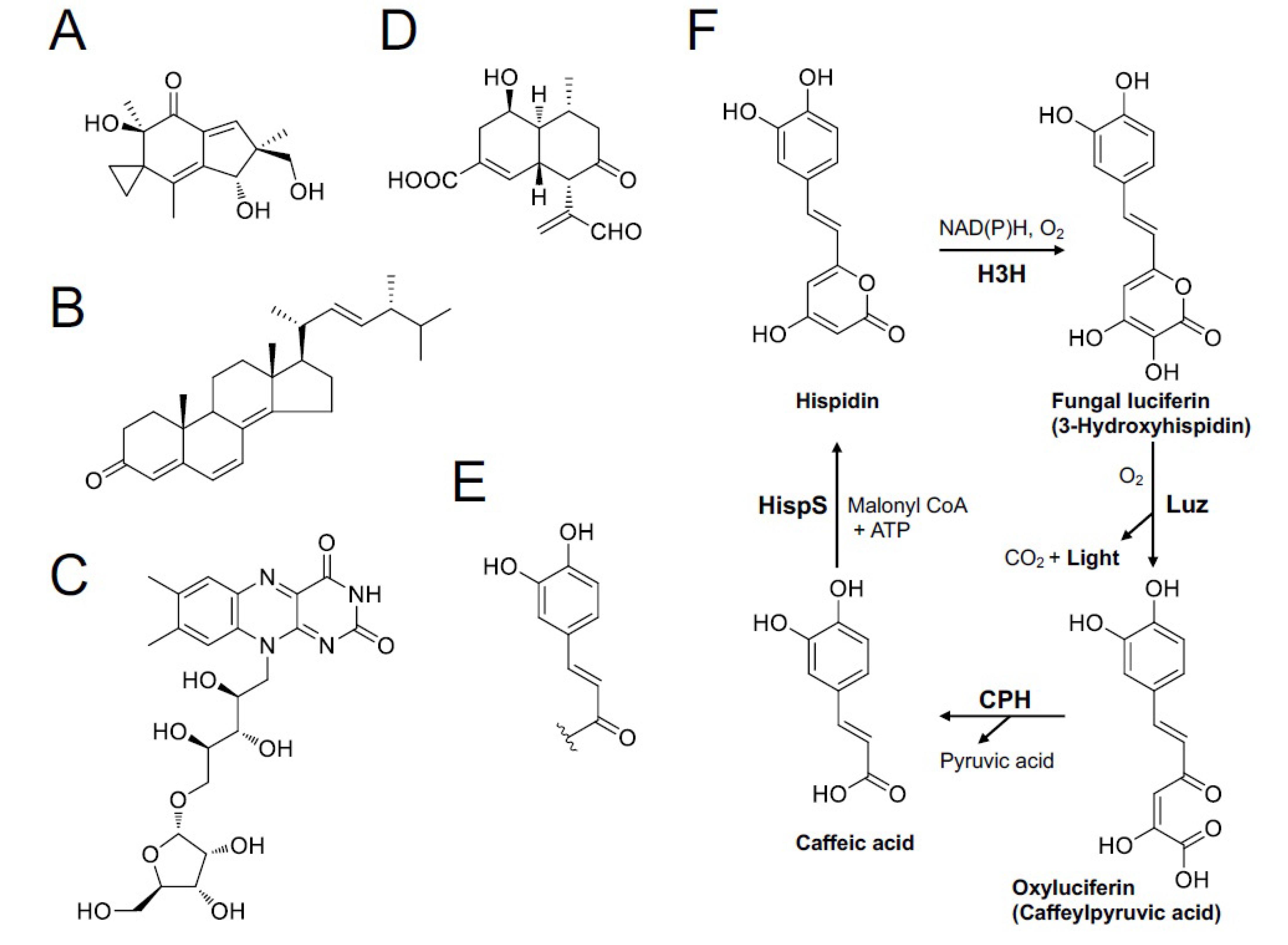{kind=link}
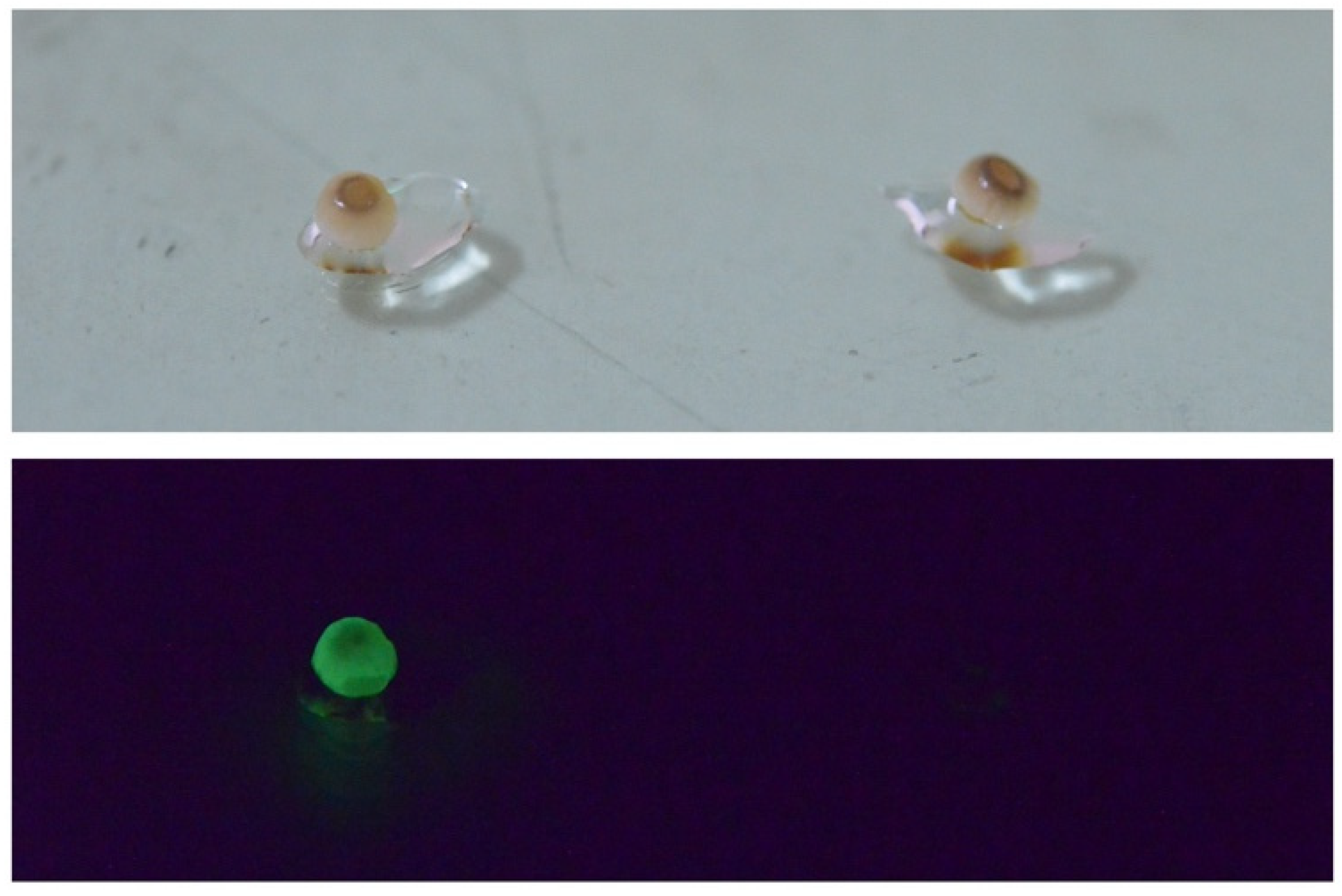{kind=link}
{kind=link}
Table 1.
Potentially bioluminescent species in Japan.
| Taxon | Japanese Name (*1) | Bioluminescence (References) (*2) |
|---|---|---|
| Family Mycenaceae | ||
| Mycena epipterygia (Scop.) S.F. Gray | Nameashi-také | Mycelium (Bothe, 1931 [86]; Wassink, 1978 [76], 1979 [45]; Desjardin et al., 2008 [46]) |
| Mycena galopus (Pers.) P. Kumm. | Nise-chishio-také | Mycelium (Bothe, 1931 [86]; Berliner, 1961 [87]; Wassink, 1978 [76], 1979 [45]; Treu & Agerer, 1990 [88]; Desjardin et al., 2008 [46]) |
| Mycena haematopus (Pers.) P. Kumm. | Chishio-také | Mycelium (Treu & Agerer, 1990 [88]; Bermudes et al., 1992 [89]; Desjardin et al., 2008 [46]): Basidiomes (weak) (Bermudes et al., 1992 [89]; Desjardin et al., 2008 [46]) |
| Mycena inclinata (Fr.) Quél. | Sembon-ashinaga-také | Mycelium (Wassink, 1978 [76], 1979 [45]; Desjardin et al., 2008 [46]) |
| Mycena olivaceomarginata (Massee) Massee | Fuchidori-kunugitaké (*3) | Mycelium (Wassink, 1978 [76], 1979 [45]; Desjardin et al., 2008 [46]) |
| Mycena pura (Pers.) P. Kumm. (*4) | Sakura-také | Mycelium (Treu & Agerer, 1990 [88]; Desjardin et al., 2008 [46]): gill of basidiome (Bothe, 1931 [86]; Wassink, 1978 [76], 1979 [45]) |
| Mycena rosea (Bull.) Gramberg | Sakurairo-také [68] | Mycelium (Treu & Agerer, 1990 [88]; Desjardin et al., 2008 [46]) |
| Mycena sanguinolenta (Alb. & Schwein.) P. Kumm. | Himé-chishio-také | Mycelium (Bothe, 1931 [86]; Wassink, 1978 [76], 1979 [45]; Desjardin et al., 2008 [46]) |
| Mycena stylobates (Pers.) P. Kumm. | Kyūban-také | Mycelium (Bothe, 1931 [86]; Wassink, 1978 [76], 1979 [45]; Desjardin et al., 2008 [46]) |
| Roridomyces roridus (Fr.) Rexer | Nunawa-také | Mycelium (Josserand, 1953 [90]; Wassink, 1978 [76], 1979 [45]; Desjardin et al., 2008 [46]) |
| Family Physalacriaceae | ||
| Armillaria fuscipes Petch (*5) | Ashiguro-narataké [91] | Mycelium (Wassink, 1978 [76], 1979 [45]; Berliner, 1961 [87]; Desjardin et al., 2008 [46]): Rhizomorph (Wassink, 1978 [76]) |
| Armillaria sinapina Berube & Dessur. | Hotei-narataké | Mycelium (Mihail, 2015 [92]) |
| Desarmillaria ectypa (Fr.) R.A. Koch & Aime | Yachihiro-hidataké | Mycelium, rhizomorph, basidiomes (Ainsworth, 2004 [93]) |
*1. The Japanese names were adopted from the list by Katumoto, 2010 [55], unless they have a more recent name. *2. Question marks represent the references showing that luminescence is doubtful or worth further investigation. Wassink (1948) [94] was not referenced in this list because his recent review papers [45,76] are considered updated versions of it. *3. Hongo (1989) [95] suggested that Mycena neoavenacea may be the same species as Mycena olivaceomarginata. *4. Molecular analysis suggests that the current M. pura morphospecies represent the species complex [96], and the bioluminescent ability of each phylospecies is unknown. *5. The morphological characteristics of this mushroom (named Ashiguro-narataké in Japanese, from Amami-Oshima) appeared indistinguishable from those of A. fuscipes, but the species name was not confirmed [97].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Oba, Y.; Hosaka, K. The Luminous Fungi of Japan. J. Fungi 2023, 9, 615. https://doi.org/10.3390/jof9060615
AMA Style
Oba Y, Hosaka K. The Luminous Fungi of Japan. Journal of Fungi. 2023; 9(6):615. https://doi.org/10.3390/jof9060615
Chicago/Turabian StyleOba, Yuichi, and Kentaro Hosaka. 2023. "The Luminous Fungi of Japan" Journal of Fungi 9, no. 6: 615. https://doi.org/10.3390/jof9060615
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.