

An Anatomy of Fungal Eye: Fungal Photoreceptors and Signalling Mechanisms

{kind=link}
{kind=link}
{kind=link}
{kind=link}
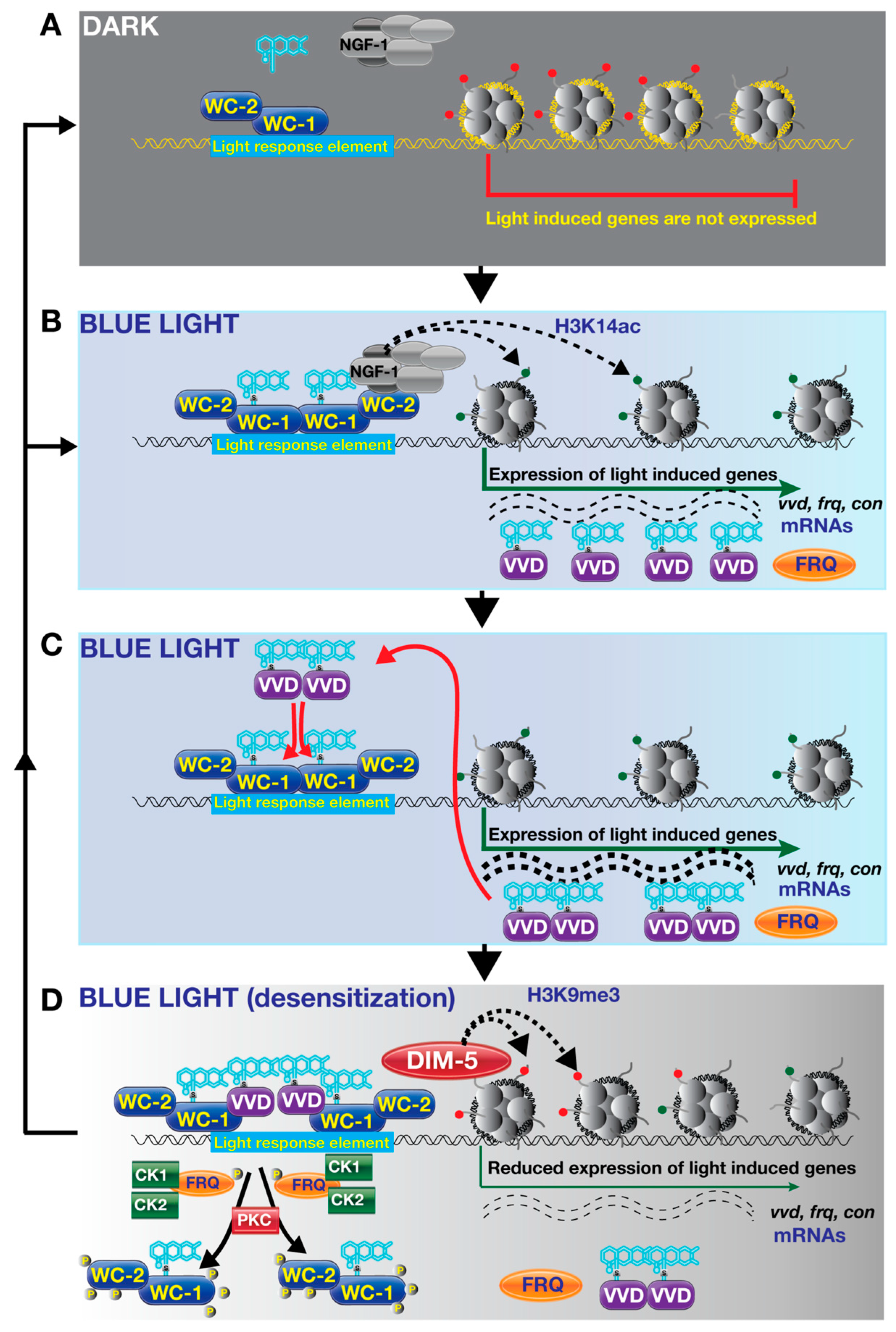{kind=link}
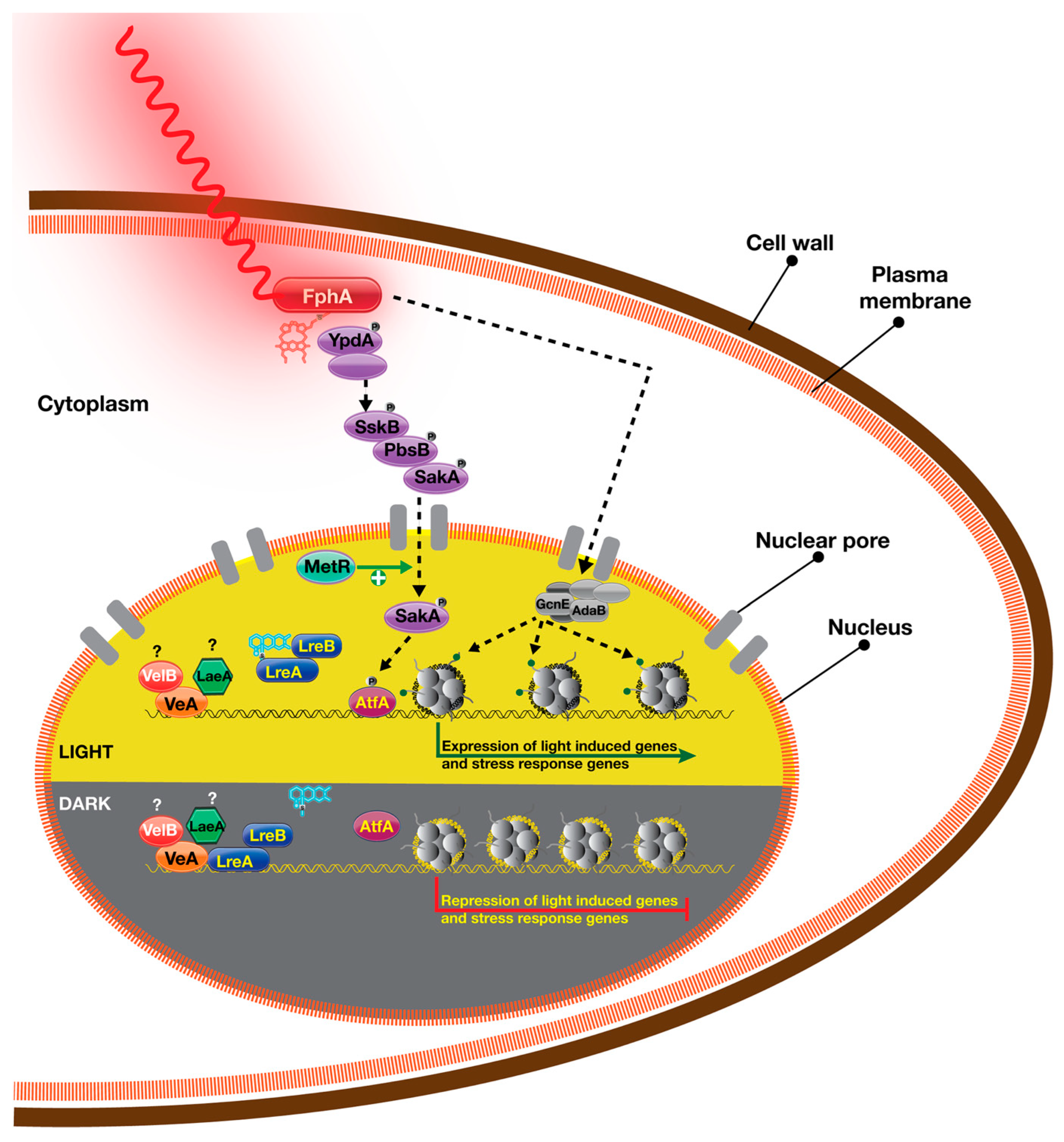{kind=link}
Abstract
:1. Introduction
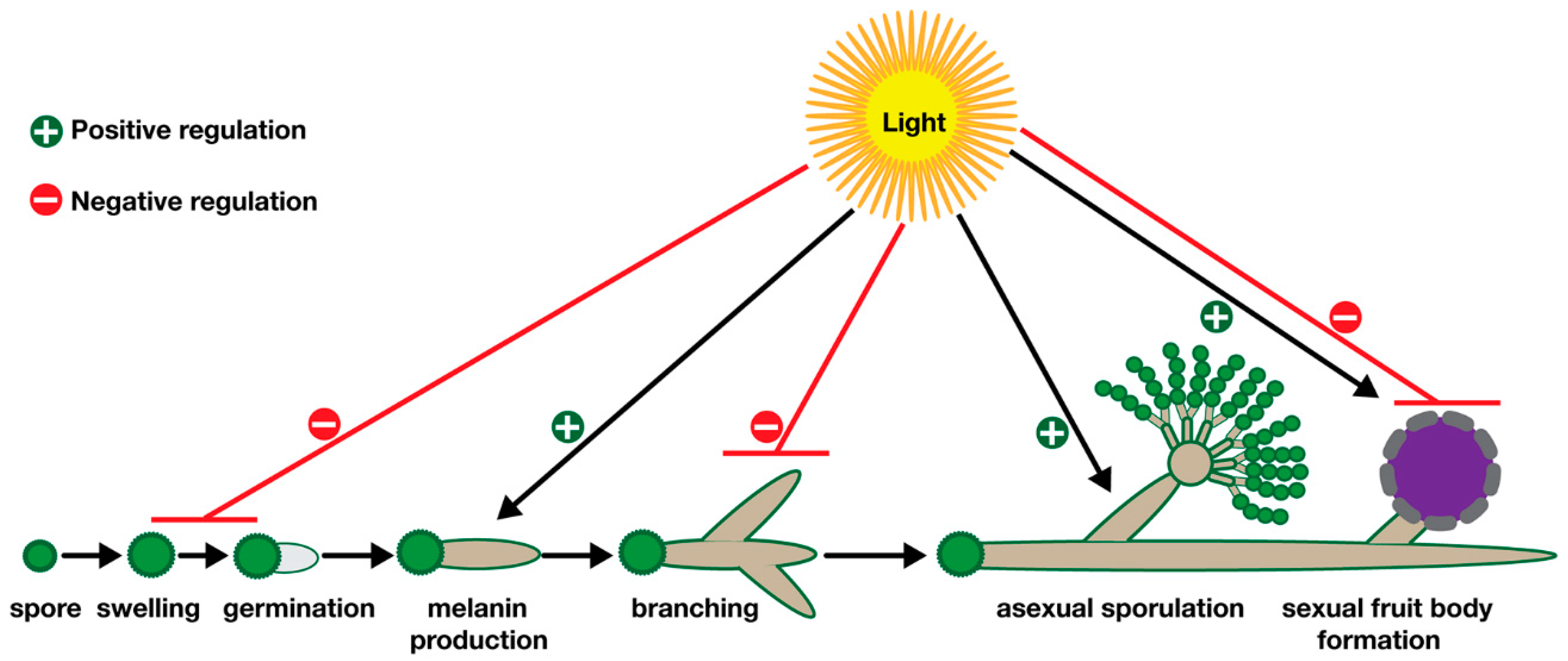
2. Fungal Light Responses
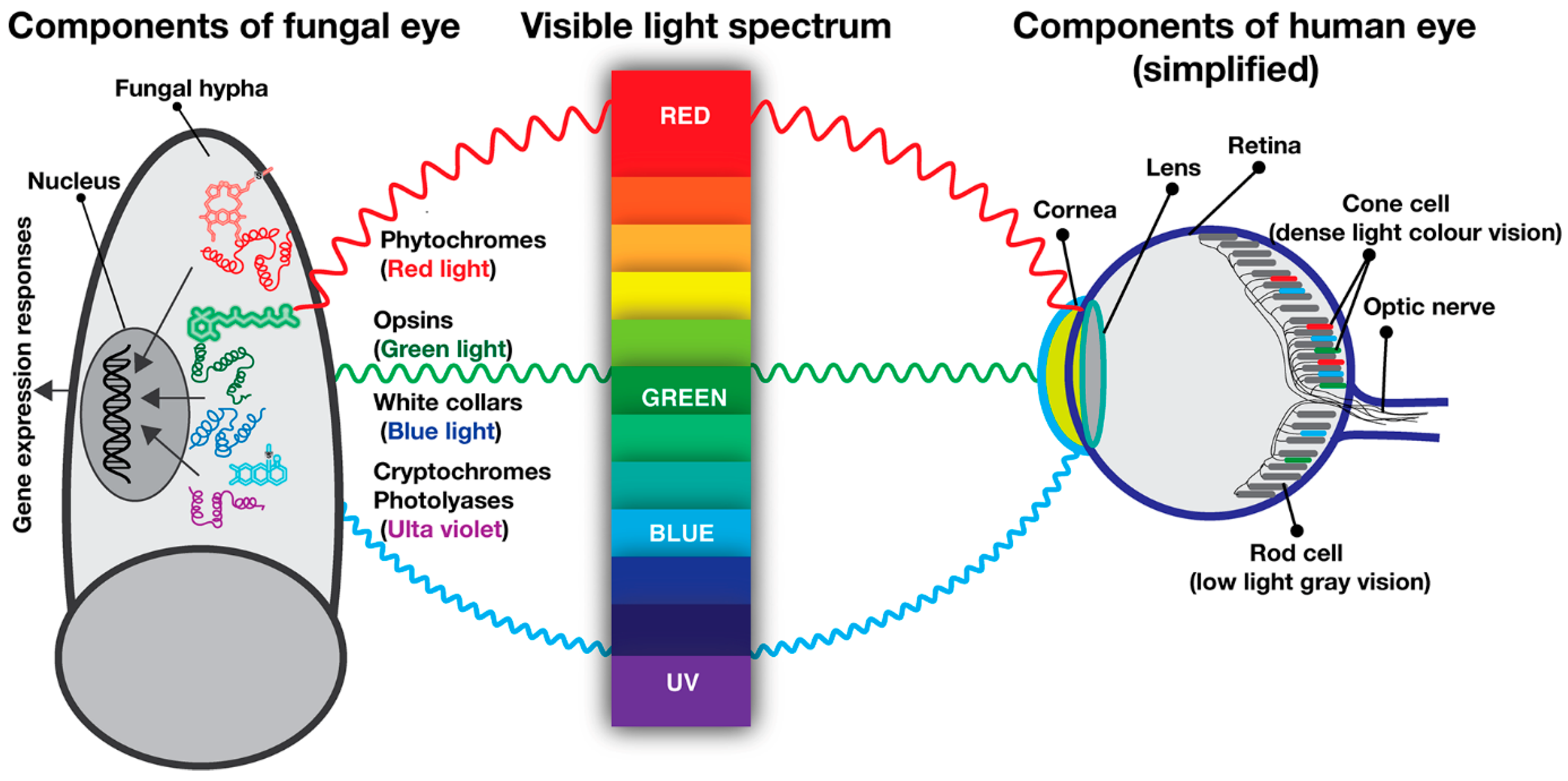
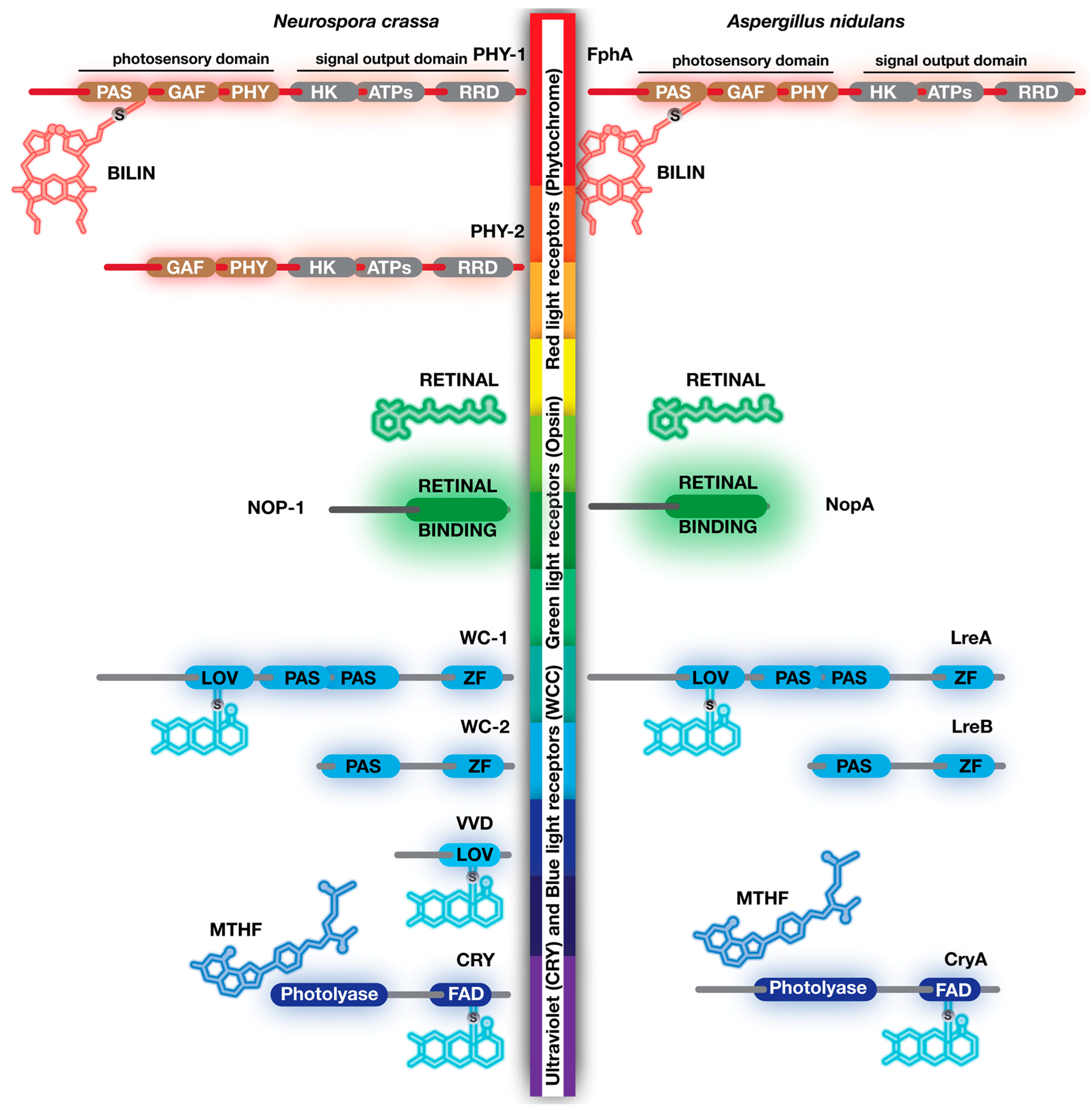
3. Fungal Photoreceptors
4. Blue Light Receptors, White Collars, Vivid, Envoy, Cryptochrome/Photolyase Family
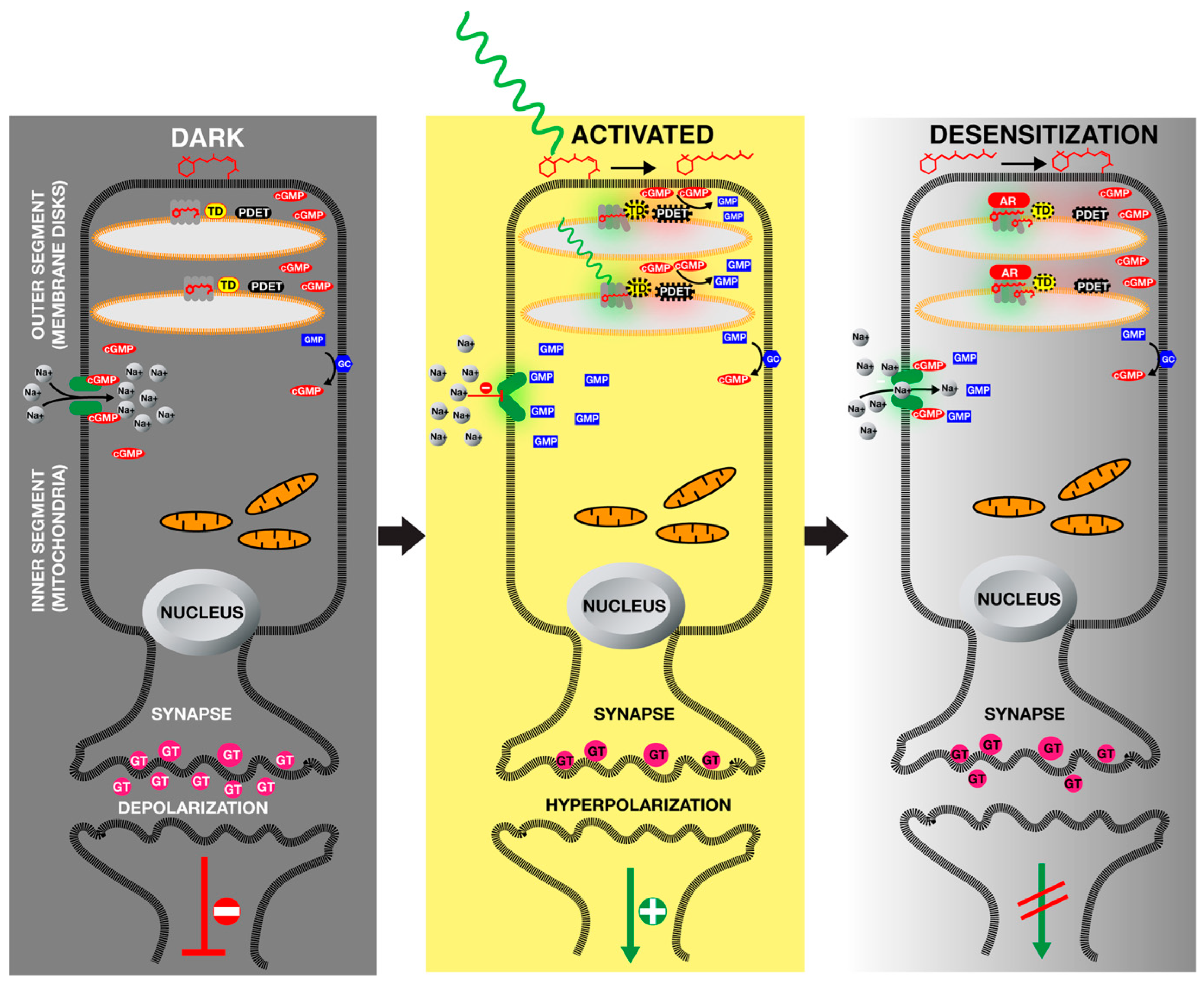
5. Green Light Receptors, Opsin Family
6. Red Light Photoreceptors, Phytochromes
7. Molecular Mechanism of Light Signal Transduction
8. Blue Light Signalling, Regulation of Light-Responsive Genes by White Collar Complexes
9. Red Light Signalling, Control of Light-Responsive Genes by Phytochrome Mediated Signalling Mechanisms

10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baslam, M.; Mitsui, T.; Hodges, M.; Priesack, E.; Herritt, M.T.; Aranjuelo, I.; Sanz-Saez, A. Photosynthesis in a Changing Global Climate: Scaling Up and Scaling Down in Crops. Front. Plant Sci. 2020, 11, 882. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.L.; Baxter, B.K. DNA Repair and Photoprotection: Mechanisms of Overcoming Environmental Ultraviolet Radiation Exposure in Halophilic Archaea. Front. Microbiol. 2017, 8, 1882. [Google Scholar] [CrossRef] [PubMed]
- Meyer, V.; Basenko, E.Y.; Benz, J.P.; Braus, G.H.; Caddick, M.X.; Csukai, M.; de Vries, R.P.; Endy, D.; Frisvad, J.C.; Gunde-Cimerman, N.; et al. Growing a circular economy with fungal biotechnology: A white paper. Fungal. Biol. Biotechnol. 2020, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Corrochano, L.M. Fungal photobiology: A synopsis. IMA Fungus. 2011, 2, 25–28. [Google Scholar] [CrossRef]
- Corrochano, L.M. Light in the Fungal World: From Photoreception to Gene Transcription and Beyond. Annu. Rev. Genet. 2019, 53, 149–170. [Google Scholar] [CrossRef]
- Corrochano, L.M.; Cerda-Olmedo, E. Photomorphogenesis in Phycomyces: Dependence on environmental conditions. Planta 1988, 174, 309–314. [Google Scholar] [CrossRef]
- Corrochano, L.M.; Galland, P.; Lipson, E.D.; Cerda-Olmedo, E. Photomorphogenesis in Phycomyces: Fluence-response curves and action spectra. Planta 1988, 174, 315–320. [Google Scholar] [CrossRef]
- Corrochano, L.M.; Lauter, F.R.; Ebbole, D.J.; Yanofsky, C. Light and developmental regulation of the gene con-10 of Neurospora crassa. Dev Biol 1995, 167, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.; Aguirre, J.; Herrera-Estrella, A.; Corrochano, L.M. The Complexity of Fungal Vision. Microbiol Spectr 2016, 4, 4–6. [Google Scholar] [CrossRef] [PubMed]
- Idnurm, A.; Verma, S.; Corrochano, L.M. A glimpse into the basis of vision in the kingdom Mycota. Fungal. Genet. Biol. 2010, 47, 881–892. [Google Scholar] [CrossRef]
- Bayram, O.; Braus, G.H. Coordination of secondary metabolism and development in fungi: The velvet family of regulatory proteins. FEMS Microbiol. Rev. 2012, 36, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, J. How light affects the life of Botrytis. Fungal. Genet. Biol. 2017, 106, 26–41. [Google Scholar] [CrossRef]
- Corrochano, L.M.; Garre, V. Photobiology in the Zygomycota: Multiple photoreceptor genes for complex responses to light. Fungal. Genet. Biol. 2010, 47, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, T.; Masuda, S. Light-induced chromophore and protein responses and mechanical signal transduction of BLUF proteins. Biophys. Rev. 2018, 10, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Palczewski, K.; Kiser, P.D. Shedding new light on the generation of the visual chromophore. Proc. Natl. Acad. Sci. USA 2020, 117, 19629–19638. [Google Scholar] [CrossRef]
- Yu, Z.; Fischer, R. Light sensing and responses in fungi. Nat. Rev. Microbiol. 2019, 17, 25–36. [Google Scholar] [CrossRef]
- Dasgupta, A.; Fuller, K.K.; Dunlap, J.C.; Loros, J.J. Seeing the world differently: Variability in the photosensory mechanisms of two model fungi. Environ. Microbiol. 2016, 18, 5–20. [Google Scholar] [CrossRef]
- Bayram, O.; Braus, G.H.; Fischer, R.; Rodriguez-Romero, J. Spotlight on Aspergillus nidulans photosensory systems. Fungal. Genet. Biol. 2010, 47, 900–908. [Google Scholar] [CrossRef]
- Pardo-Medina, J.; Limón, M.C.; Avalos, J. Fusarium Photoreceptors. J. Fungi 2023, 9, 319. [Google Scholar] [CrossRef]
- Sarikaya Bayram, O.; Bayram, O.; Karahoda, B.; Meister, C.; Kohler, A.M.; Thieme, S.; Elramli, N.; Frawley, D.; McGowan, J.; Fitzpatrick, D.A.; et al. F-box receptor mediated control of substrate stability and subcellular location organizes cellular development of Aspergillus nidulans. PLoS Genet. 2022, 18, e1010502. [Google Scholar] [CrossRef]
- Karahoda, B.; Pardeshi, L.; Ulas, M.; Dong, Z.; Shirgaonkar, N.; Guo, S.; Wang, F.; Tan, K.; Sarikaya-Bayram, O.; Bauer, I.; et al. The KdmB-EcoA-RpdA-SntB chromatin complex binds regulatory genes and coordinates fungal development with mycotoxin synthesis. Nucleic Acids Res. 2022, 50, 9797–9813. [Google Scholar] [CrossRef] [PubMed]
- Irniger, S.; Sarikaya-Bayram, Ö.; Bayram, Ö. 6 Fungal MAP-Kinase-Mediated Regulatory Pathways. In Biochemistry and Molecular Biology; Hoffmeister, D., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 97–117. [Google Scholar]
- Garcia-Martinez, J.; Brunk, M.; Avalos, J.; Terpitz, U. The CarO rhodopsin of the fungus Fusarium fujikuroi is a light-driven proton pump that retards spore germination. Sci. Rep. 2015, 5, 7798. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Qin, H.; Zhang, X.; Li, W.; Liu, M. Inhibition of Aspergillus oryzae Mycelium Growth and Conidium Production by Irradiation with Light at Different Wavelengths and Intensities. Microbiol. Spectr. 2021, 9, e0021321. [Google Scholar] [CrossRef]
- Rohrig, J.; Kastner, C.; Fischer, R. Light inhibits spore germination through phytochrome in Aspergillus nidulans. Curr. Genet. 2013, 59, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Casas-Flores, S.; Rios-Momberg, M.; Bibbins, M.; Ponce-Noyola, P.; Herrera-Estrella, A. BLR-1 and BLR-2, key regulatory elements of photoconidiation and mycelial growth in Trichoderma atroviride. Microbiology 2004, 150, 3561–3569. [Google Scholar] [CrossRef]
- Avalos, J.; Estrada, A.F. Regulation by light in Fusarium. Fungal. Genet. Biol. 2010, 47, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Fuller, K.K.; Loros, J.J.; Dunlap, J.C. Fungal photobiology: Visible light as a signal for stress, space and time. Curr. Genet. 2015, 61, 275–288. [Google Scholar] [CrossRef] [PubMed]
- Kamada, T.; Sano, H.; Nakazawa, T.; Nakahori, K. Regulation of fruiting body photomorphogenesis in Coprinopsis cinerea. Fungal. Genet. Biol. 2010, 47, 917–921. [Google Scholar] [CrossRef]
- Elramli, N.; Karahoda, B.; Sarikaya-Bayram, O.; Frawley, D.; Ulas, M.; Oakley, C.E.; Oakley, B.R.; Seiler, S.; Bayram, O. Assembly of a heptameric STRIPAK complex is required for coordination of light-dependent multicellular fungal development with secondary metabolism in Aspergillus nidulans. PLoS Genet. 2019, 15, e1008053. [Google Scholar] [CrossRef]
- Suzuki, S.; Sarikaya Bayram, O.; Bayram, O.; Braus, G.H. conF and conJ contribute to conidia germination and stress response in the filamentous fungus Aspergillus nidulans. Fungal Genet. Biol. 2013, 56, 42–53. [Google Scholar] [CrossRef]
- Valsecchi, I.; Sarikaya-Bayram, O.; Wong Sak Hoi, J.; Muszkieta, L.; Gibbons, J.; Prevost, M.C.; Mallet, A.; Krijnse-Locker, J.; Ibrahim-Granet, O.; Mouyna, I.; et al. MybA, a transcription factor involved in conidiation and conidial viability of the human pathogen Aspergillus fumigatus. Mol. Microbiol. 2017, 105, 880–900. [Google Scholar] [CrossRef] [PubMed]
- Sarikaya Bayram, O.; Latge, J.P.; Bayram, O. MybA, a new player driving survival of the conidium of the human pathogen Aspergillus fumigatus. Curr. Genet 2018, 64, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Blachowicz, A.; Raffa, N.; Bok, J.W.; Choera, T.; Knox, B.; Lim, F.Y.; Huttenlocher, A.; Wang, C.C.C.; Venkateswaran, K.; Keller, N.P. Contributions of Spore Secondary Metabolites to UV-C Protection and Virulence Vary in Different Aspergillus fumigatus Strains. mBio 2020, 11, e03415–e03419. [Google Scholar] [CrossRef]
- Rodriguez-Romero, J.; Hedtke, M.; Kastner, C.; Muller, S.; Fischer, R. Fungi, hidden in soil or up in the air: Light makes a difference. Annu Rev Microbiol 2010, 64, 585–610. [Google Scholar] [CrossRef] [PubMed]
- Ni, M.; Feretzaki, M.; Sun, S.; Wang, X.; Heitman, J. Sex in fungi. Annu. Rev. Genet. 2011, 45, 405–430. [Google Scholar] [CrossRef]
- Molday, R.S.; Moritz, O.L. Photoreceptors at a glance. J. Cell Sci. 2015, 128, 4039–4045. [Google Scholar] [CrossRef] [PubMed]
- Hussey, K.A.; Hadyniak, S.E.; Johnston, R.J., Jr. Patterning and Development of Photoreceptors in the Human Retina. Front. Cell Dev. Biol. 2022, 10, 878350. [Google Scholar] [CrossRef]
- Mure, L.S. Intrinsically Photosensitive Retinal Ganglion Cells of the Human Retina. Front Neurol. 2021, 12, 636330. [Google Scholar] [CrossRef]
- Ballario, P.; Vittorioso, P.; Magrelli, A.; Talora, C.; Cabibbo, A.; Macino, G. White collar-1, a central regulator of blue light responses in Neurospora, is a zinc finger protein. EMBO J. 1996, 15, 1650–1657. [Google Scholar] [CrossRef]
- Perkins, D.D.; Newmeyer, D.; Taylor, C.W.; Bennett, D.C. New markers and map sequences in Neurospora crassa, with a description of mapping by duplication coverage, and of multiple translocation stocks for testing linkage. Genetica 1969, 40, 247–278. [Google Scholar] [CrossRef]
- Idnurm, A.; Rodriguez-Romero, J.; Corrochano, L.M.; Sanz, C.; Iturriaga, E.A.; Eslava, A.P.; Heitman, J. The Phycomyces madA gene encodes a blue-light photoreceptor for phototropism and other light responses. Proc. Natl. Acad. Sci. USA 2006, 103, 4546–4551. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.L.; Zhulin, I.B. PAS domains: Internal sensors of oxygen, redox potential, and light. Microbiol. Mol. Biol. Rev. 1999, 63, 479–506. [Google Scholar] [CrossRef] [PubMed]
- Purschwitz, J.; Muller, S.; Kastner, C.; Schoser, M.; Haas, H.; Espeso, E.A.; Atoui, A.; Calvo, A.M.; Fischer, R. Functional and physical interaction of blue- and red-light sensors in Aspergillus nidulans. Curr. Biol. 2008, 18, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Ballario, P.; Macino, G. White collar proteins: PASsing the light signal in Neurospora crassa. Trends Microbiol. 1997, 5, 458–462. [Google Scholar] [CrossRef] [PubMed]
- Schleicher, E.; Kowalczyk, R.M.; Kay, C.W.; Hegemann, P.; Bacher, A.; Fischer, M.; Bittl, R.; Richter, G.; Weber, S. On the reaction mechanism of adduct formation in LOV domains of the plant blue-light receptor phototropin. J. Am. Chem. Soc. 2004, 126, 11067–11076. [Google Scholar] [CrossRef]
- Froehlich, A.C.; Liu, Y.; Loros, J.J.; Dunlap, J.C. White Collar-1, a circadian blue light photoreceptor, binding to the frequency promoter. Science 2002, 297, 815–819. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.; Torres-Martinez, S.; Garre, V. Distinct white collar-1 genes control specific light responses in Mucor circinelloides. Mol. Microbiol. 2006, 61, 1023–1037. [Google Scholar] [CrossRef]
- Rodriguez-Romero, J.; Corrochano, L.M. Regulation by blue light and heat shock of gene transcription in the fungus Phycomyces: Proteins required for photoinduction and mechanism for adaptation to light. Mol. Microbiol. 2006, 61, 1049–1059. [Google Scholar] [CrossRef]
- Schmoll, M.; Franchi, L.; Kubicek, C.P. Envoy, a PAS/LOV domain protein of Hypocrea jecorina (Anamorph Trichoderma reesei), modulates cellulase gene transcription in response to light. Eukaryot. Cell 2005, 4, 1998–2007. [Google Scholar] [CrossRef]
- Shrode, L.B.; Lewis, Z.A.; White, L.D.; Bell-Pedersen, D.; Ebbole, D.J. vvd is required for light adaptation of conidiation-specific genes of Neurospora crassa, but not circadian conidiation. Fungal Genet. Biol. 2001, 32, 169–181. [Google Scholar] [CrossRef]
- Schmoll, M. Regulation of plant cell wall degradation by light in Trichoderma. Fungal Biol. Biotechnol. 2018, 5, 10. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, A.T.; Chen, C.H.; Dunlap, J.C.; Loros, J.J.; Crane, B.R. Structure of a light-activated LOV protein dimer that regulates transcription. Sci. Signal 2011, 4, ra50. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.T.; Malzahn, E.; Brunner, M.; Mayer, M.P. Light-induced differences in conformational dynamics of the circadian clock regulator VIVID. J. Mol. Biol. 2014, 426, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, L.; Zhong, D. Dynamics and mechanisms of DNA repair by photolyase. Phys. Chem. Chem. Phys. 2015, 17, 11933–11949. [Google Scholar] [CrossRef]
- Michael, A.K.; Fribourgh, J.L.; Van Gelder, R.N.; Partch, C.L. Animal Cryptochromes: Divergent Roles in Light Perception, Circadian Timekeeping and Beyond. Photochem. Photobiol. 2017, 93, 128–140. [Google Scholar] [CrossRef]
- Kiontke, S.; Gobel, T.; Brych, A.; Batschauer, A. DASH-type cryptochromes—Solved and open questions. Biol. Chem. 2020, 401, 1487–1493. [Google Scholar] [CrossRef]
- Bayram, O.; Biesemann, C.; Krappmann, S.; Galland, P.; Braus, G.H. More than a repair enzyme: Aspergillus nidulans photolyase-like CryA is a regulator of sexual development. Mol. Biol. Cell 2008, 19, 3254–3262. [Google Scholar] [CrossRef]
- Yu, Z.; Streng, C.; Seibeld, R.F.; Igbalajobi, O.A.; Leister, K.; Ingelfinger, J.; Fischer, R. Genome-wide analyses of light-regulated genes in Aspergillus nidulans reveal a complex interplay between different photoreceptors and novel photoreceptor functions. PLoS Genet. 2021, 17, e1009845. [Google Scholar] [CrossRef]
- Froehlich, A.C.; Chen, C.H.; Belden, W.J.; Madeti, C.; Roenneberg, T.; Merrow, M.; Loros, J.J.; Dunlap, J.C. Genetic and molecular characterization of a cryptochrome from the filamentous fungus Neurospora crassa. Eukaryot. Cell 2010, 9, 738–750. [Google Scholar] [CrossRef]
- Nsa, I.Y.; Karunarathna, N.; Liu, X.; Huang, H.; Boetteger, B.; Bell-Pedersen, D. A novel cryptochrome-dependent oscillator in Neurospora crassa. Genetics 2015, 199, 233–245. [Google Scholar] [CrossRef]
- Bieszke, J.A.; Spudich, E.N.; Scott, K.L.; Borkovich, K.A.; Spudich, J.L. A eukaryotic protein, NOP-1, binds retinal to form an archaeal rhodopsin-like photochemically reactive pigment. Biochemistry 1999, 38, 14138–14145. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.S.; Dioumaev, A.K.; Lanyi, J.K.; Spudich, E.N.; Spudich, J.L. Photochemical reaction cycle and proton transfers in Neurospora rhodopsin. J. Biol. Chem. 2001, 276, 32495–32505. [Google Scholar] [CrossRef] [PubMed]
- Ernst, O.P.; Lodowski, D.T.; Elstner, M.; Hegemann, P.; Brown, L.S.; Kandori, H. Microbial and animal rhodopsins: Structures, functions, and molecular mechanisms. Chem. Rev. 2014, 114, 126–163. [Google Scholar] [CrossRef] [PubMed]
- Galindo, L.J.; Milner, D.S.; Gomes, S.L.; Richards, T.A. A light-sensing system in the common ancestor of the fungi. Curr. Biol. 2022, 32, 3146–3153.e3. [Google Scholar] [CrossRef]
- Blaalid, R.; Khomich, M. Current knowledge of Chytridiomycota diversity in Northern Europe and future research needs. Fungal Biol. Rev. 2021, 36, 42–51. [Google Scholar] [CrossRef]
- Navarro, E.; Penaranda, A.; Hansberg, W.; Torres-Martinez, S.; Garre, V. A white collar 1-like protein mediates opposite regulatory functions in Mucor circinelloides. Fungal Genet. Biol. 2013, 52, 42–52. [Google Scholar] [CrossRef]
- Blumenstein, A.; Vienken, K.; Tasler, R.; Purschwitz, J.; Veith, D.; Frankenberg-Dinkel, N.; Fischer, R. The Aspergillus nidulans phytochrome FphA represses sexual development in red light. Curr. Biol. 2005, 15, 1833–1838. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, N.; Li, J.; Dunlap, J.C.; Trail, F.; Townsend, J.P. The Fast-Evolving phy-2 Gene Modulates Sexual Development in Response to Light in the Model Fungus Neurospora crassa. mBio 2016, 7, e02148. [Google Scholar] [CrossRef]
- Dombrovsky, A.; Arthaud, L.; Ledger, T.N.; Tares, S.; Robichon, A. Profiling the repertoire of phenotypes influenced by environmental cues that occur during asexual reproduction. Genome Res. 2009, 19, 2052–2063. [Google Scholar] [CrossRef]
- Li, F.W.; Melkonian, M.; Rothfels, C.J.; Villarreal, J.C.; Stevenson, D.W.; Graham, S.W.; Wong, G.K.; Pryer, K.M.; Mathews, S. Phytochrome diversity in green plants and the origin of canonical plant phytochromes. Nat. Commun. 2015, 6, 7852. [Google Scholar] [CrossRef]
- Hirakawa, H.; Kurushima, J.; Hashimoto, Y.; Tomita, H. Progress Overview of Bacterial Two-Component Regulatory Systems as Potential Targets for Antimicrobial Chemotherapy. Antibiotics 2020, 9, 635. [Google Scholar] [CrossRef] [PubMed]
- Lokhandwala, J.; Hopkins, H.C.; Rodriguez-Iglesias, A.; Dattenbock, C.; Schmoll, M.; Zoltowski, B.D. Structural biochemistry of a fungal LOV domain photoreceptor reveals an evolutionarily conserved pathway integrating light and oxidative stress. Structure 2015, 23, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Streng, C.; Hartmann, J.; Leister, K.; Krauss, N.; Lamparter, T.; Frankenberg-Dinkel, N.; Weth, F.; Bastmeyer, M.; Yu, Z.; Fischer, R. Fungal phytochrome chromophore biosynthesis at mitochondria. EMBO J. 2021, 40, e108083. [Google Scholar] [CrossRef] [PubMed]
- Wensel, T. Molecular Biology of Vision. In Basic Neurochemistry; Academic Press: Cambridge, MA, USA, 2012; pp. 889–903. [Google Scholar] [CrossRef]
- Fain, G.L.; Hardie, R.; Laughlin, S.B. Phototransduction and the evolution of photoreceptors. Curr. Biol. 2010, 20, R114–R124. [Google Scholar] [CrossRef] [PubMed]
- Tyler, J.; Lu, Y.; Dunlap, J.; Forger, D.B. Evolution of the repression mechanisms in circadian clocks. Genome Biol. 2022, 23, 17. [Google Scholar] [CrossRef]
- Chen, C.H.; Loros, J.J. Neurospora sees the light: Light signaling components in a model system. Commun. Integr. Biol. 2009, 2, 448–451. [Google Scholar] [CrossRef]
- Dunlap, J.C.; Loros, J.J. Making Time: Conservation of Biological Clocks from Fungi to Animals. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef]
- Hurley, J.; Loros, J.J.; Dunlap, J.C. Dissecting the mechanisms of the clock in Neurospora. Methods Enzymol. 2015, 551, 29–52. [Google Scholar] [CrossRef]
- Fuller, K.K.; Dunlap, J.C.; Loros, J.J. Fungal Light Sensing at the Bench and Beyond. Adv. Genet. 2016, 96, 1–51. [Google Scholar] [CrossRef]
- Goldsmith, C.S.; Bell-Pedersen, D. Diverse roles for MAPK signaling in circadian clocks. Adv. Genet. 2013, 84, 1–39. [Google Scholar] [CrossRef]
- Lakin-Thomas, P.L.; Bell-Pedersen, D.; Brody, S. The genetics of circadian rhythms in Neurospora. Adv. Genet. 2011, 74, 55–103. [Google Scholar] [CrossRef]
- Schwerdtfeger, C.; Linden, H. Blue light adaptation and desensitization of light signal transduction in Neurospora crassa. Mol. Microbiol. 2001, 39, 1080–1087. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Liu, Y. Molecular mechanism of light responses in Neurospora: From light-induced transcription to photoadaptation. Genes Dev. 2005, 19, 2888–2899. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Cheng, P.; Yang, Y.; Wang, L.; Gardner, K.H.; Liu, Y. White collar-1, a DNA binding transcription factor and a light sensor. Science 2002, 297, 840–843. [Google Scholar] [CrossRef]
- Ma, H.; Li, L.; Yan, J.; Zhang, Y.; Ma, X.; Li, Y.; Yuan, Y.; Yang, X.; Yang, L.; Guo, J. The Resonance and Adaptation of Neurospora crassa Circadian and Conidiation Rhyth ms to Short Light-Dark Cycles. J. Fungi 2021, 8, 27. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, B.; Coiro, P.; Filetici, P.; Berge, E.; Dobosy, J.R.; Freitag, M.; Selker, E.U.; Ballario, P. The Neurospora crassa White Collar-1 dependent blue light response requires acetylation of histone H3 lysine 14 by NGF-1. Mol. Biol. Cell 2006, 17, 4576–4583. [Google Scholar] [CrossRef] [PubMed]
- Hedtke, M.; Rauscher, S.; Rohrig, J.; Rodriguez-Romero, J.; Yu, Z.; Fischer, R. Light-dependent gene activation in Aspergillus nidulans is strictly dependent on phytochrome and involves the interplay of phytochrome and white collar-regulated histone H3 acetylation. Mol. Microbiol. 2015, 97, 733–745. [Google Scholar] [CrossRef]
- Purschwitz, J.; Muller, S.; Fischer, R. Mapping the interaction sites of Aspergillus nidulans phytochrome FphA with the global regulator VeA and the White Collar protein LreB. Mol. Genet. Genomics 2009, 281, 35–42. [Google Scholar] [CrossRef]
- Sancar, C.; Ha, N.; Yilmaz, R.; Tesorero, R.; Fisher, T.; Brunner, M.; Sancar, G. Combinatorial control of light induced chromatin remodeling and gene activation in Neurospora. PLoS Genet. 2015, 11, e1005105. [Google Scholar] [CrossRef]
- Kaldi, K.; Gonzalez, B.H.; Brunner, M. Transcriptional regulation of the Neurospora circadian clock gene wc-1 affects the phase of circadian output. EMBO Rep. 2006, 7, 199–204. [Google Scholar] [CrossRef]
- Wu, C.; Yang, F.; Smith, K.M.; Peterson, M.; Dekhang, R.; Zhang, Y.; Zucker, J.; Bredeweg, E.L.; Mallappa, C.; Zhou, X.; et al. Genome-wide characterization of light-regulated genes in Neurospora crassa. G3 (Bethesda) 2014, 4, 1731–1745. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.M.; Sancar, G.; Dekhang, R.; Sullivan, C.M.; Li, S.; Tag, A.G.; Sancar, C.; Bredeweg, E.L.; Priest, H.D.; McCormick, R.F.; et al. Transcription factors in light and circadian clock signaling networks revealed by genomewide mapping of direct targets for neurospora white collar complex. Eukaryot. Cell 2010, 9, 1549–1556. [Google Scholar] [CrossRef] [PubMed]
- Schafmeier, T.; Kaldi, K.; Diernfellner, A.; Mohr, C.; Brunner, M. Phosphorylation-dependent maturation of Neurospora circadian clock protein from a nuclear repressor toward a cytoplasmic activator. Genes Dev. 2006, 20, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Talora, C.; Franchi, L.; Linden, H.; Ballario, P.; Macino, G. Role of a white collar-1-white collar-2 complex in blue-light signal transduction. EMBO J. 1999, 18, 4961–4968. [Google Scholar] [CrossRef]
- Marzoll, D.; Serrano, F.E.; Shostak, A.; Schunke, C.; Diernfellner, A.C.R.; Brunner, M. Casein kinase 1 and disordered clock proteins form functionally equivalent, phospho-based circadian modules in fungi and mammals. Proc. Natl. Acad. Sci. USA 2022, 119, e2118286119. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Cha, J.; He, Q.; Lee, H.C.; Yang, Y.; Liu, Y. CKI and CKII mediate the FREQUENCY-dependent phosphorylation of the WHITE COLLAR complex to close the Neurospora circadian negative feedback loop. Genes Dev. 2006, 20, 2552–2565. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.T.; Li, S.; Long, C.; Cha, J.; Huang, G.; Li, L.; Chen, S.; Liu, Y. Setting the pace of the Neurospora circadian clock by multiple independent FRQ phosphorylation events. Proc. Natl. Acad. Sci. USA 2009, 106, 10722–10727. [Google Scholar] [CrossRef]
- Querfurth, C.; Diernfellner, A.C.; Gin, E.; Malzahn, E.; Hofer, T.; Brunner, M. Circadian conformational change of the Neurospora clock protein FREQUENCY triggered by clustered hyperphosphorylation of a basic domain. Mol. Cell 2011, 43, 713–722. [Google Scholar] [CrossRef]
- Shi, M.; Collett, M.; Loros, J.J.; Dunlap, J.C. FRQ-interacting RNA helicase mediates negative and positive feedback in the Neurospora circadian clock. Genetics 2010, 184, 351–361. [Google Scholar] [CrossRef]
- Cha, J.; Chang, S.S.; Huang, G.; Cheng, P.; Liu, Y. Control of WHITE COLLAR localization by phosphorylation is a critical step in the circadian negative feedback process. EMBO J. 2008, 27, 3246–3255. [Google Scholar] [CrossRef]
- Huang, G.; Chen, S.; Li, S.; Cha, J.; Long, C.; Li, L.; He, Q.; Liu, Y. Protein kinase A and casein kinases mediate sequential phosphorylation events in the circadian negative feedback loop. Genes Dev. 2007, 21, 3283–3295. [Google Scholar] [CrossRef]
- Wang, B.; Kettenbach, A.N.; Zhou, X.; Loros, J.J.; Dunlap, J.C. The Phospho-Code Determining Circadian Feedback Loop Closure and Output in Neurospora. Mol. Cell 2019, 74, 771–784.e3. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, V.V.; Hanson, S.M.; Song, X.; Vishnivetskiy, S.A.; Gurevich, E.V. The functional cycle of visual arrestins in photoreceptor cells. Prog. Retin. Eye Res. 2011, 30, 405–430. [Google Scholar] [CrossRef]
- Perkins, D.D.; Margolin, B.S.; Selker, E.U.; Haedo, S.D. Occurrence of repeat induced point mutation in long segmental duplications of Neurospora. Genetics 1997, 147, 125–136. [Google Scholar] [CrossRef]
- Dasgupta, A.; Chen, C.H.; Lee, C.; Gladfelter, A.S.; Dunlap, J.C.; Loros, J.J. Biological Significance of Photoreceptor Photocycle Length: VIVID Photocycle Governs the Dynamic VIVID-White Collar Complex Pool Mediating Photo-adaptation and Response to Changes in Light Intensity. PLoS Genet. 2015, 11, e1005215. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, A.; Caicedo-Casso, A.; Cui, G.; Du, M.; He, Q.; Lim, S.; Kim, H.J.; Hong, C.I.; Liu, Y. FRQ-CK1 interaction determines the period of circadian rhythms in Neurospora. Nat. Commun. 2019, 10, 4352. [Google Scholar] [CrossRef] [PubMed]
- Inglese, J.; Glickman, J.F.; Lorenz, W.; Caron, M.G.; Lefkowitz, R.J. Isoprenylation of a protein kinase. Requirement of farnesylation/alpha-carboxyl methylation for full enzymatic activity of rhodopsin kinase. J. Biol. Chem. 1992, 267, 1422–1425. [Google Scholar] [CrossRef]
- Kutuzov, M.A.; Andreeva, A.V.; Bennett, N. Regulation of the methylation status of G protein-coupled receptor kinase 1 (rhodopsin kinase). Cell. Signal 2012, 24, 2259–2267. [Google Scholar] [CrossRef]
- Ruesch, C.E.; Ramakrishnan, M.; Park, J.; Li, N.; Chong, H.S.; Zaman, R.; Joska, T.M.; Belden, W.J. The histone H3 lysine 9 methyltransferase DIM-5 modifies chromatin at frequency and represses light-activated gene expression. G3 (Bethesda) 2014, 5, 93–101. [Google Scholar] [CrossRef]
- Yu, Z.; Armant, O.; Fischer, R. Fungi use the SakA (HogA) pathway for phytochrome-dependent light signalling. Nat. Microbiol. 2016, 1, 16019. [Google Scholar] [CrossRef]
- Pardi, S.A.; Nusinow, D.A. Out of the Dark and Into the Light: A New View of Phytochrome Photobodies. Front. Plant Sci. 2021, 12, 732947. [Google Scholar] [CrossRef] [PubMed]
- Sarikaya Bayram, O.; Bayram, O.; Valerius, O.; Park, H.S.; Irniger, S.; Gerke, J.; Ni, M.; Han, K.H.; Yu, J.H.; Braus, G.H. LaeA control of velvet family regulatory proteins for light-dependent development and fungal cell-type specificity. PLoS Genet. 2010, 6, e1001226. [Google Scholar] [CrossRef] [PubMed]
- Bayram, O.; Bayram, O.S.; Valerius, O.; Johnk, B.; Braus, G.H. Identification of protein complexes from filamentous fungi with tandem affinity purification. Methods Mol. Biol. 2012, 944, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Bayram, O.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.J.; Keller, N.P.; Yu, J.H.; et al. VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef] [PubMed]
- Han, K.H.; Prade, R.A. Osmotic stress-coupled maintenance of polar growth in Aspergillus nidulans. Mol. Microbiol. 2002, 43, 1065–1078. [Google Scholar] [CrossRef]
- Jaimes-Arroyo, R.; Lara-Rojas, F.; Bayram, O.; Valerius, O.; Braus, G.H.; Aguirre, J. The SrkA Kinase Is Part of the SakA Mitogen-Activated Protein Kinase Interactome and Regulates Stress Responses and Development in Aspergillus nidulans. Eukaryot. Cell 2015, 14, 495–510. [Google Scholar] [CrossRef]
- Azuma, N.; Kanamaru, K.; Matsushika, A.; Yamashino, T.; Mizuno, T.; Kato, M.; Kobayashi, T. In vitro analysis of His-Asp phosphorelays in Aspergillus nidulans: The first direct biochemical evidence for the existence of His-Asp phosphotransfer systems in filamentous fungi. Biosci. Biotechnol. Biochem. 2007, 71, 2493–2502. [Google Scholar] [CrossRef]
- Yu, Z.; Gao, J.; Igbalajobi, O.; Skoneczny, M.; Sienko, M.; Maciejewska, A.M.; Brzywczy, J.; Fischer, R. The sulfur metabolism regulator MetR is a global regulator controlling phytochrome-dependent light responses in Aspergillus nidulans. Sci. Bull. 2021, 66, 592–602. [Google Scholar] [CrossRef]
- Cerimi, K.; Akkaya, K.C.; Pohl, C.; Schmidt, B.; Neubauer, P. Fungi as source for new bio-based materials: A patent review. Fungal Biol. Biotechnol. 2019, 6, 17. [Google Scholar] [CrossRef]
- Meyer, V. Connecting materials sciences with fungal biology: A sea of possibilities. Fungal Biol. Biotechnol. 2022, 9, 5. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bayram, Ö.S.; Bayram, Ö. An Anatomy of Fungal Eye: Fungal Photoreceptors and Signalling Mechanisms. J. Fungi 2023, 9, 591. https://doi.org/10.3390/jof9050591
Bayram ÖS, Bayram Ö. An Anatomy of Fungal Eye: Fungal Photoreceptors and Signalling Mechanisms. Journal of Fungi. 2023; 9(5):591. https://doi.org/10.3390/jof9050591
Chicago/Turabian StyleBayram, Özlem Sarikaya, and Özgür Bayram. 2023. "An Anatomy of Fungal Eye: Fungal Photoreceptors and Signalling Mechanisms" Journal of Fungi 9, no. 5: 591. https://doi.org/10.3390/jof9050591