
Only One of Three Bcs1 Homologs in Aspergillus fumigatus Confers Respiratory Growth


Abstract
:1. Introduction
2. Material and Methods
2.1. Strains, Culture Conditions, and Chemicals

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Genotype | Relevant Genetic Modification | Parental Strain | Reference |
|---|---|---|---|
| AfS35 (wt) | akuA::loxP | D141 | [14] |
| Δbcs1A | bcs1A::loxP-hygroR/blaster | AfS35 (wt) | This work |
| bcs1 tetOn | (p)bcs1A::ptrA-(p)pkiA-tetOn | AfS35 (wt) | This work |
| Δbcs1B | bcs1B::loxP-hygroR/blaster | AfS35 (wt) | This work |
| bcs1BtetOn | (p)bcs1B::ptrA-(p)pkiA-tetOn | AfS35 (wt) | This work |
| Δbcs1C | bcs1C::loxP-hygroR/blaster | AfS35 (wt) | This work |
| bcs1CtetOn | (p)bcs1C::ptrA-(p)pkiA-tetOn | AfS35 (wt) | This work |
| Rip1tetOn | (p)Rip1::ptrA-(p)pkiA-tetOn | AfS35 (wt) | [23] |
| wt + Bcs1B-GFP | pIK002 | AfS35 (wt) | This work |
| wt + Bcs1C-GFP | pIK003 | AfS35 (wt) | This work |
| wt + mtRFP | pYZ012 | AfS35 (wt) | This work |
| wt + mtRFP + Bcs1B-GFP | pIK002 | wt + mtRFP | This work |
| wt + mtRFP + Bcs1C-GFP | pIK003 | wt + mtRFP | This work |
2.2. Microscopy
2.3. Galleria mellonella Infection Experiments
2.4. Bioinformatics and Databases
3. Results
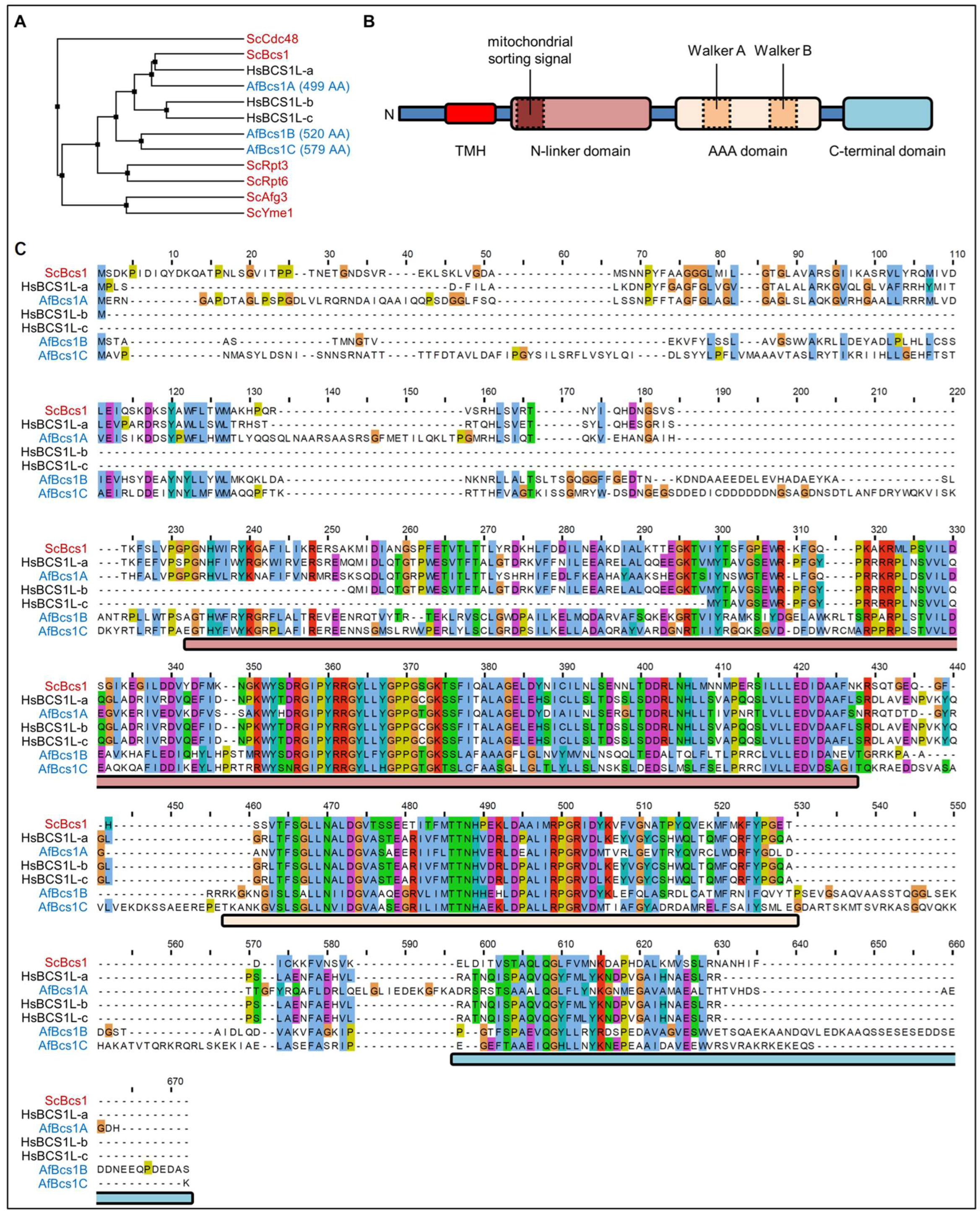
3.1. A. fumigatus Encodes Three Bcs1 Homologs
3.2. Construction and Phenotypic Characterization of bcs1A, bcs1B, and bcs1C Mutants
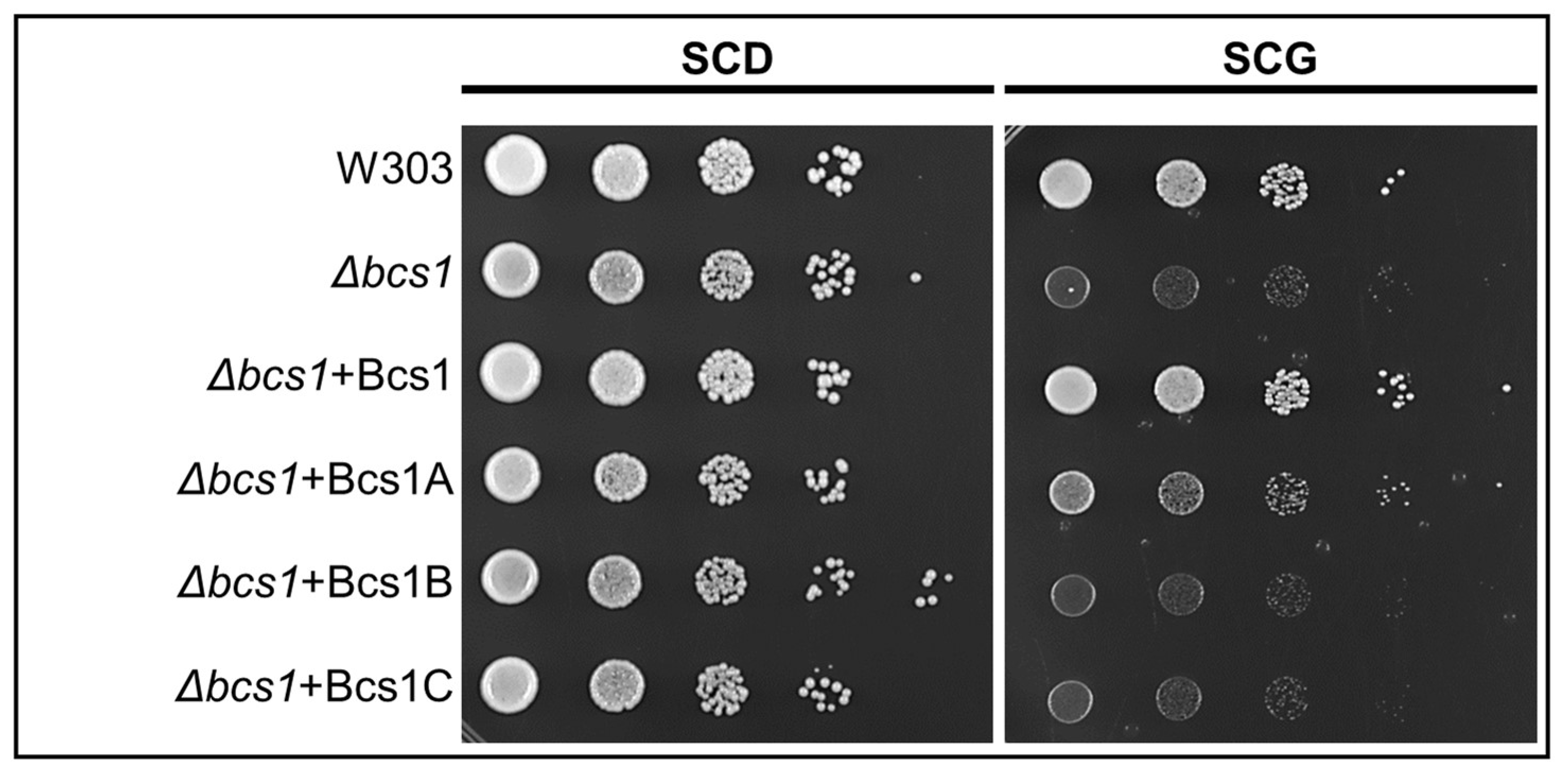
3.3. Expression of A. fumigatus bcs1A Partially Complements the BCS1Deletion in S. cerevisiae
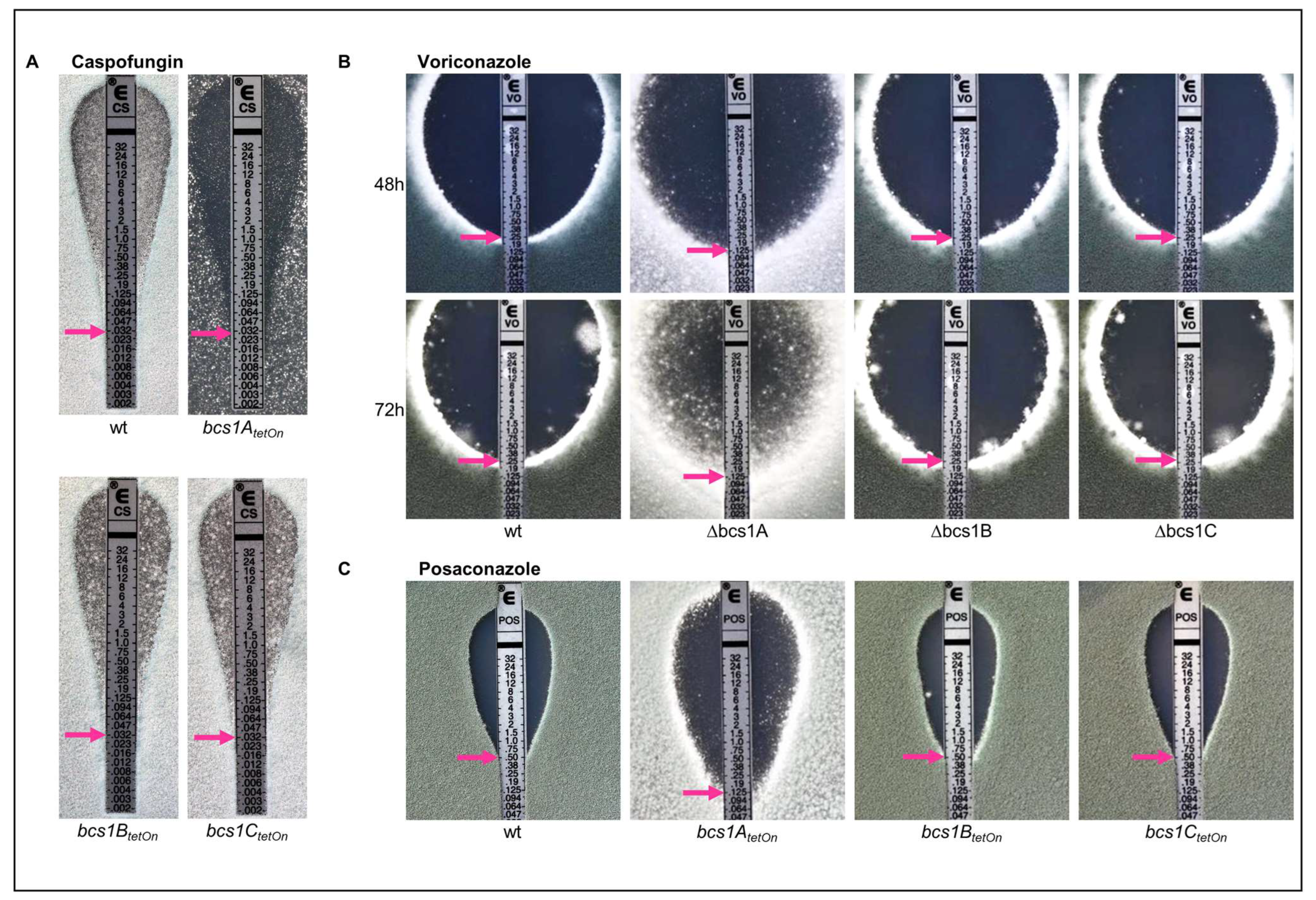
3.4. Deletion of bcs1 Leads to an Increased Growth-Inhibiting but Decreased Fungicidal Activity of Azole Antifungals in A. fumigatus
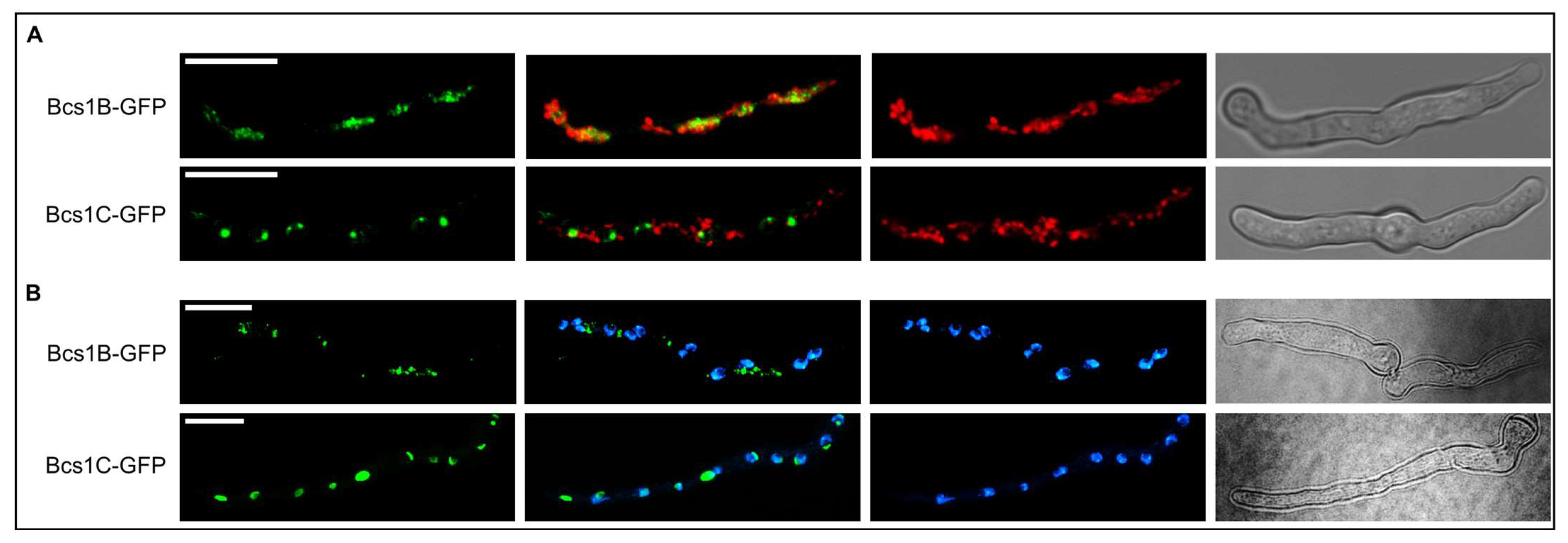
3.5. Subcellular Localizations of Bcs1B and Bcs1C Suggest Functional Roles Distinct from That of Bcs1A
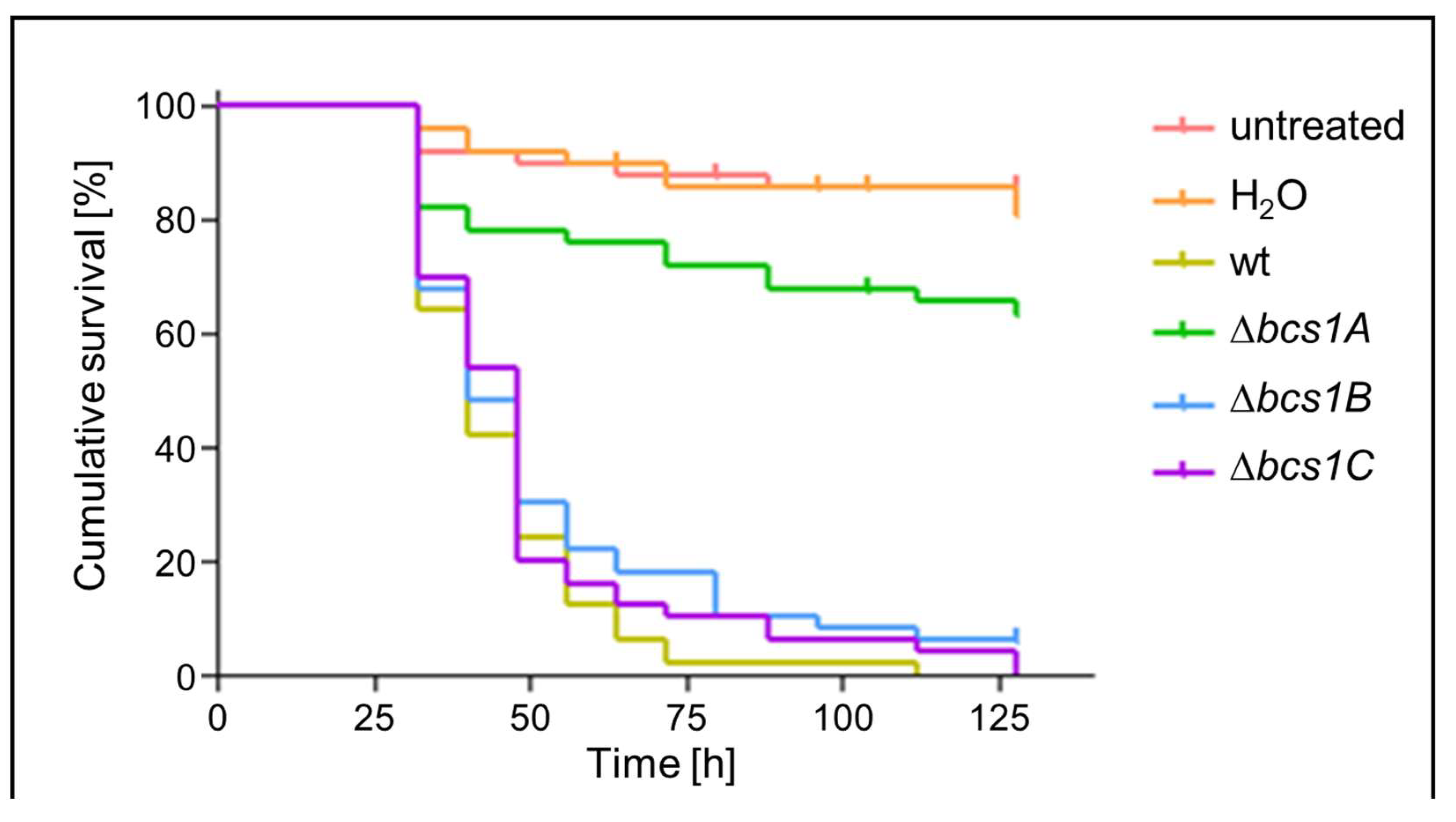
3.6. Bcs1A Is Required for Full Virulence of A. fumigatus in a Galleria mellonella Infection Model
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sousa, J.S.; D’Imprima, E.; Vonck, J. Mitochondrial Respiratory Chain Complexes. Subcell. Biochem. 2018, 87, 167–227. [Google Scholar] [CrossRef] [PubMed]
- Stan, T.; Brix, J.; Schneider-Mergener, J.; Pfanner, N.; Neupert, W.; Rapaport, D. Mitochondrial protein import: Recognition of internal import signals of BCS1 by the TOM complex. Mol. Cell. Biol. 2003, 23, 2239–2250. [Google Scholar] [CrossRef] [PubMed]
- Lange, C.; Hunte, C. Crystal structure of the yeast cytochrome bc1 complex with its bound substrate cytochrome c. Proc. Natl. Acad. Sci. USA 2002, 99, 2800–2805. [Google Scholar] [CrossRef] [PubMed]
- Wagener, N.; Ackermann, M.; Funes, S.; Neupert, W. A pathway of protein translocation in mitochondria mediated by the AAA-ATPase Bcs1. Mol. Cell 2011, 44, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Zara, V.; Palmisano, I.; Conte, L.; Trumpower, B.L. Further insights into the assembly of the yeast cytochrome bc1 complex based on analysis of single and double deletion mutants lacking supernumerary subunits and cytochrome b. Eur. J. Biochem. 2004, 271, 1209–1218. [Google Scholar] [CrossRef]
- Jackson, C.B.; Bauer, M.F.; Schaller, A.; Kotzaeridou, U.; Ferrarini, A.; Hahn, D.; Chehade, H.; Barbey, F.; Tran, C.; Gallati, S.; et al. A novel mutation in BCS1L associated with deafness, tubulopathy, growth retardation and microcephaly. Eur. J. Pediatr. 2016, 175, 517–525. [Google Scholar] [CrossRef]
- Levéen, P.; Kotarsky, H.; Mörgelin, M.; Karikoski, R.; Elmér, E.; Fellman, V. The GRACILE mutation introduced into Bcs1l causes postnatal complex III deficiency: A viable mouse model for mitochondrial hepatopathy. Hepatology 2011, 53, 437–447. [Google Scholar] [CrossRef]
- Siddiqi, S.; Siddiq, S.; Mansoor, A.; Oostrik, J.; Ahmad, N.; Kazmi, S.A.R.; Kremer, H.; Qamar, R.; Schraders, M. Novel mutation in AAA domain of BCS1L causing Bjornstad syndrome. J. Hum. Genet. 2013, 58, 819–821. [Google Scholar] [CrossRef]
- Kater, L.; Wagener, N.; Berninghausen, O.; Becker, T.; Neupert, W.; Beckmann, R. Structure of the Bcs1 AAA-ATPase suggests an airlock-like translocation mechanism for folded proteins. Nat. Struct. Mol. Biol. 2020, 27, 142–149. [Google Scholar] [CrossRef]
- Tang, W.K.; Borgnia, M.J.; Hsu, A.L.; Esser, L.; Fox, T.; de Val, N.; Xia, D. Structures of AAA protein translocase Bcs1 suggest translocation mechanism of a folded protein. Nat. Struct. Mol. Biol. 2020, 27, 202–209. [Google Scholar] [CrossRef]
- NM_001374085.1 BCS1L BCS1 Homolog, Ubiquinol-Cytochrome c Reductase Complex Chaperone [Homo Sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/?term=NM_001374085.1 (accessed on 21 September 2021).
- Zhan, J.; Xia, D. Bcs1, a novel target for fungicide. Front. Chem. 2023, 11, 1146753. [Google Scholar] [CrossRef] [PubMed]
- Thompson, G.R.; Young, J.-A.H. Aspergillus Infections. N. Engl. J. Med. 2021, 385, 1496–1509. [Google Scholar] [CrossRef] [PubMed]
- Krappmann, S.; Sasse, C.; Braus, G.H. Gene targeting in Aspergillus fumigatus by homologous recombination is facilitated in a nonhomologous end- joining-deficient genetic background. Eukaryot. Cell 2006, 5, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Wagener, J.; Echtenacher, B.; Rohde, M.; Kotz, A.; Krappmann, S.; Heesemann, J.; Ebel, F. The putative alpha-1,2-mannosyltransferase AfMnt1 of the opportunistic fungal pathogen Aspergillus fumigatus is required for cell wall stability and full virulence. Eukaryot. Cell 2008, 7, 1661–1673. [Google Scholar] [CrossRef]
- Neubauer, M.; Zhu, Z.; Penka, M.; Helmschrott, C.; Wagener, N.; Wagener, J. Mitochondrial dynamics in the pathogenic mold Aspergillus fumigatus: Therapeutic and evolutionary implications. Mol. Microbiol. 2015, 98, 930–945. [Google Scholar] [CrossRef]
- Hartmann, T.; Dümig, M.; Jaber, B.M.; Szewczyk, E.; Olbermann, P.; Morschhäuser, J.; Krappmann, S. Validation of a self-excising marker in the human pathogen Aspergillus fumigatus by employing the beta-rec/six site-specific recombination system. Appl. Environ. Microbiol. 2010, 76, 6313–6317. [Google Scholar] [CrossRef]
- Helmschrott, C.; Sasse, A.; Samantaray, S.; Krappmann, S.; Wagener, J. Upgrading fungal gene expression on demand: Improved systems for doxycycline-dependent silencing in Aspergillus fumigatus. Appl. Environ. Microbiol. 2013, 79, 1751–1754. [Google Scholar] [CrossRef]
- Dichtl, K.; Ebel, F.; Dirr, F.; Routier, F.H.; Heesemann, J.; Wagener, J. Farnesol misplaces tip-localized Rho proteins and inhibits cell wall integrity signalling in Aspergillus fumigatus. Mol. Microbiol. 2010, 76, 1191–1204. [Google Scholar] [CrossRef]
- Hill, T.W.; Kafer, E. Improved protocols for Aspergillus salt stock solutions. Fungal Genet Newsl 2001, 48, 20–21. [Google Scholar]
- Nobrega, F.G.; Nobrega, M.P.; Tzagoloff, A. BCS1, a novel gene required for the expression of functional Rieske iron-sulfur protein in Saccharomyces cerevisiae. EMBO J. 1992, 11, 3821–3829. [Google Scholar] [CrossRef]
- Sturm, L.; Geißel, B.; Martin, R.; Wagener, J. Differentially Regulated Transcription Factors and ABC Transporters in a Mitochondrial Dynamics Mutant Can Alter Azole Susceptibility of Aspergillus fumigatus. Front. Microbiol. 2020, 11, 1017. [Google Scholar] [CrossRef] [PubMed]
- Geißel, B.; Loiko, V.; Klugherz, I.; Zhu, Z.; Wagener, N.; Kurzai, O.; van den Hondel, C.A.M.J.J.; Wagener, J. Azole-induced cell wall carbohydrate patches kill Aspergillus fumigatus. Nat. Commun. 2018, 9, 3098. [Google Scholar] [CrossRef]
- Cerqueira, G.C.; Arnaud, M.B.; Inglis, D.O.; Skrzypek, M.S.; Binkley, G.; Simison, M.; Miyasato, S.R.; Binkley, J.; Orvis, J.; Shah, P.; et al. The Aspergillus Genome Database: Multispecies curation and incorporation of RNA-Seq data to improve structural gene annotations. Nucleic Acids Res. 2014, 42, D705–D710. [Google Scholar] [CrossRef]
- Amos, B.; Aurrecoechea, C.; Barba, M.; Barreto, A.; Basenko, E.Y.; Bażant, W.; Belnap, R.; Blevins, A.S.; Böhme, U.; Brestelli, J.; et al. VEuPathDB: The eukaryotic pathogen, vector and host bioinformatics resource center. Nucleic Acids Res. 2022, 50, D898–D911. [Google Scholar] [CrossRef] [PubMed]
- Cherry, J.M.; Hong, E.L.; Amundsen, C.; Balakrishnan, R.; Binkley, G.; Chan, E.T.; Christie, K.R.; Costanzo, M.C.; Dwight, S.S.; Engel, S.R.; et al. Saccharomyces Genome Database: The genomics resource of budding yeast. Nucleic Acids Res. 2012, 40, D700–D705. [Google Scholar] [CrossRef] [PubMed]
- Gramates, L.S.; Agapite, J.; Attrill, H.; Calvi, B.R.; Crosby, M.A.; Dos Santos, G.; Goodman, J.L.; Goutte-Gattat, D.; Jenkins, V.K.; Kaufman, T.; et al. FlyBase: A Guided Tour of Highlighted Features. Genetics 2022, 220, iyac035. [Google Scholar] [CrossRef]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2022, 50, D20–D26. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—A Multiple Sequence Alignment Editor and Analysis Workbench. Bioinforma. Oxf. Engl. 2009, 25, 1189–1191. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Fölsch, H.; Guiard, B.; Neupert, W.; Stuart, R.A. Internal Targeting Signal of the BCS1 Protein: A Novel Mechanism of Import into Mitochondria. EMBO J. 1996, 15, 479–487. [Google Scholar] [CrossRef]
- Frickey, T.; Lupas, A.N. Phylogenetic Analysis of AAA Proteins. J. Struct. Biol. 2004, 146, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Hinson, J.T.; Fantin, V.R.; Schönberger, J.; Breivik, N.; Siem, G.; McDonough, B.; Sharma, P.; Keogh, I.; Godinho, R.; Santos, F.; et al. Missense Mutations in the BCS1L Gene as a Cause of the Björnstad Syndrome. N. Engl. J. Med. 2007, 356, 809–819. [Google Scholar] [CrossRef]
- Morán, M.; Marín-Buera, L.; Gil-Borlado, M.C.; Rivera, H.; Blázquez, A.; Seneca, S.; Vázquez-López, M.; Arenas, J.; Martín, M.A.; Ugalde, C. Cellular Pathophysiological Consequences of BCS1L Mutations in Mitochondrial Complex III Enzyme Deficiency. Hum. Mutat. 2010, 31, 930–941. [Google Scholar] [CrossRef] [PubMed]
- Grahl, N.; Dinamarco, T.M.; Willger, S.D.; Goldman, G.H.; Cramer, R.A. Aspergillus Fumigatus Mitochondrial Electron Transport Chain Mediates Oxidative Stress Homeostasis, Hypoxia Responses and Fungal Pathogenesis. Mol. Microbiol. 2012, 84, 383–399. [Google Scholar] [CrossRef] [PubMed]
- Petruzzella, V.; Tiranti, V.; Fernandez, P.; Ianna, P.; Carrozzo, R.; Zeviani, M. Identification and Characterization of Human cDNAs Specific to BCS1, PET112, SCO1, COX15, and COX11, Five Genes Involved in the Formation and Function of the Mitochondrial Respiratory Chain. Genomics 1998, 54, 494–504. [Google Scholar] [CrossRef]
- Visapää, I.; Fellman, V.; Vesa, J.; Dasvarma, A.; Hutton, J.L.; Kumar, V.; Payne, G.S.; Makarow, M.; Van Coster, R.; Taylor, R.W.; et al. GRACILE Syndrome, a Lethal Metabolic Disorder with Iron Overload, Is Caused by a Point Mutation in BCS1L. Am. J. Hum. Genet. 2002, 71, 863–876. [Google Scholar] [CrossRef]
- Shingu-Vazquez, M.; Traven, A. Mitochondria and Fungal Pathogenesis: Drug Tolerance, Virulence, and Potential for Antifungal Therapy. Eukaryot. Cell 2011, 10, 1376–1383. [Google Scholar] [CrossRef]
- de Lonlay, P.; Valnot, I.; Barrientos, A.; Gorbatyuk, M.; Tzagoloff, A.; Taanman, J.W.; Benayoun, E.; Chrétien, D.; Kadhom, N.; Lombès, A.; et al. A Mutant Mitochondrial Respiratory Chain Assembly Protein Causes Complex III Deficiency in Patients with Tubulopathy, Encephalopathy and Liver Failure. Nat. Genet. 2001, 29, 57–60. [Google Scholar] [CrossRef]
- Vincent, B.M.; Langlois, J.-B.; Srinivas, R.; Lancaster, A.K.; Scherz-Shouval, R.; Whitesell, L.; Tidor, B.; Buchwald, S.L.; Lindquist, S. A Fungal-Selective Cytochrome Bc1 Inhibitor Impairs Virulence and Prevents the Evolution of Drug Resistance. Cell Chem. Biol. 2016, 23, 978–991. [Google Scholar] [CrossRef]
- Fisher, N.; Meunier, B.; Biagini, G.A. The Cytochrome Bc1 Complex as an Antipathogenic Target. FEBS Lett. 2020, 594, 2935–2952. [Google Scholar] [CrossRef]
- Gow, N.A.R.; Johnson, C.; Berman, J.; Coste, A.T.; Cuomo, C.A.; Perlin, D.S.; Bicanic, T.; Harrison, T.S.; Wiederhold, N.; Bromley, M.; et al. The Importance of Antimicrobial Resistance in Medical Mycology. Nat. Commun. 2022, 13, 5352. [Google Scholar] [CrossRef] [PubMed]
- Lamoth, F.; Lewis, R.E.; Kontoyiannis, D.P. Investigational Antifungal Agents for Invasive Mycoses: A Clinical Perspective. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2022, 75, 534–544. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, S.; Remuson, F.; Caddoux, L.; Barrès, B. Investigation of the Sensitivity of Plasmopara Viticola to Amisulbrom and Ametoctradin in French Vineyards Using Bioassays and Molecular Tools. Pest Manag. Sci. 2019, 75, 2115–2123. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klugherz, I.; Basch, M.; Ng, N.; Zhu, Z.; Wagener, N.; Wagener, J. Only One of Three Bcs1 Homologs in Aspergillus fumigatus Confers Respiratory Growth. J. Fungi 2023, 9, 1074. https://doi.org/10.3390/jof9111074
Klugherz I, Basch M, Ng N, Zhu Z, Wagener N, Wagener J. Only One of Three Bcs1 Homologs in Aspergillus fumigatus Confers Respiratory Growth. Journal of Fungi. 2023; 9(11):1074. https://doi.org/10.3390/jof9111074
Chicago/Turabian StyleKlugherz, Isabel, Marion Basch, Natanya Ng, Zhaojun Zhu, Nikola Wagener, and Johannes Wagener. 2023. "Only One of Three Bcs1 Homologs in Aspergillus fumigatus Confers Respiratory Growth" Journal of Fungi 9, no. 11: 1074. https://doi.org/10.3390/jof9111074