
Phenotypic and Genotypic Characterization and Antifungal Susceptibility of Sporothrix schenckii sensu stricto Isolated from a Feline Sporotrichosis Outbreak in Bangkok, Thailand
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolates and Data Collection
2.2. Phenotypic Characterization
2.3. Genotypic Characterization
2.4. Antifungal Susceptibility Test
3. Results
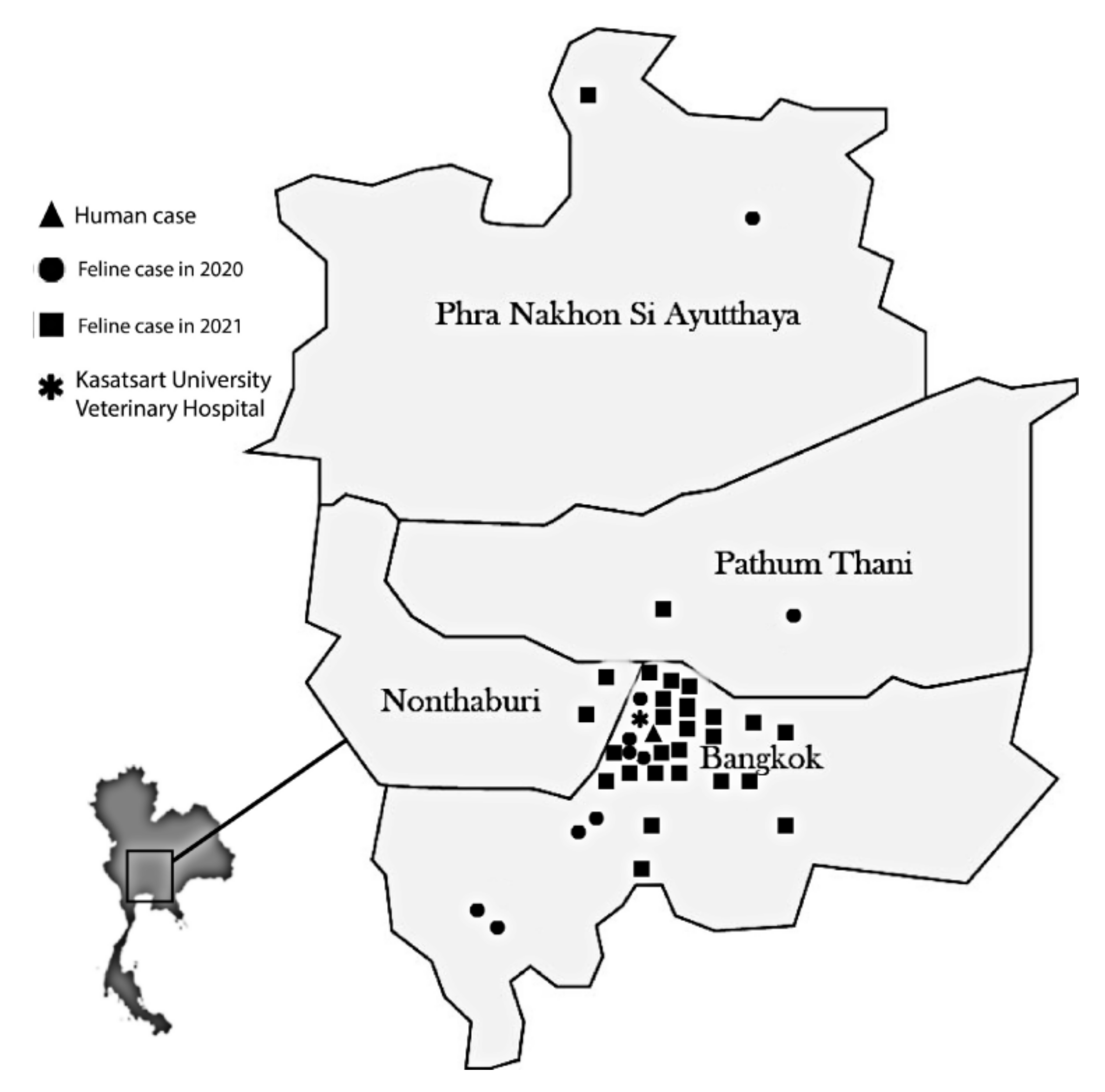
3.1. Geographic Distribution, Demographic Characteristics, and Clinical Data
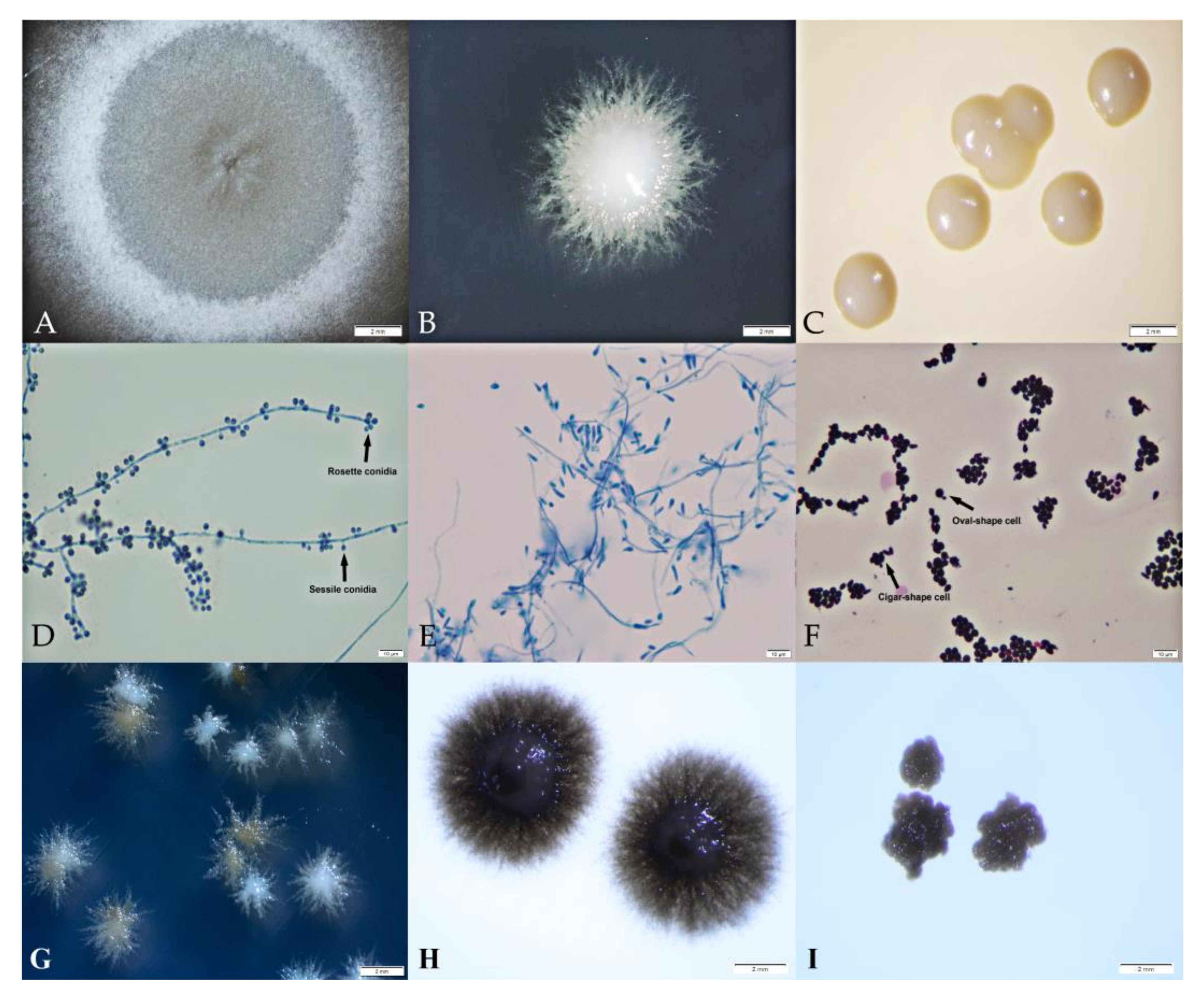
3.2. Phenotypic Characteristics of S. schenckii Thai Isolates
3.3. Phylogenetic Analysis of S. schenckii Thai Isolates
3.4. Antifungal Susceptibility
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Téllez, M.D.; Batista-Duharte, A.; Portuondo, D.; Quinello, C.; Bonne-Hernandez, R.; Carlos, I. Sporothrix schenckii complex biology: Environment and fungal pathogenicity. Microbiology 2014, 160, 2352–2365. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.M.; de Melo Teixeira, M.; de Hoog, G.S.; Schubach, T.M.P.; Pereira, S.A.; Fernandes, G.F.; Bezerra, L.M.L.; Felipe, M.S.; de Camargo, Z.P. Phylogenetic analysis reveals a high prevalence of Sporothrix brasiliensis in feline sporotrichosis outbreaks. PLoS Negl. Trop. Dis. 2013, 7, e2281. [Google Scholar] [CrossRef] [PubMed]
- Schubach, T.M.; Schubach, A.; Okamoto, T.; Barros, M.B.; Figueiredo, F.B.; Cuzzi, T.; Fialho-Monteiro, P.C.; Reis, R.S.; Perez, M.A.; Wanke, B. Evaluation of an epidemic of sporotrichosis in cats: 347 cases (1998–2001). J. Am. Vet. Med. Assoc. 2004, 224, 1623–1629. [Google Scholar] [CrossRef] [PubMed]
- Barros, M.B.d.L.; de Almeida Paes, R.; Schubach, A.O. Sporothrix schenckii and sporotrichosis. Clin. Microbiol. Rev. 2011, 24, 633–654. [Google Scholar] [CrossRef]
- Barros, M.B.; Schubach, T.P.; Coll, J.O.; Gremião, I.D.; Wanke, B.; Schubach, A. Sporotrichosis: Development and challenges of an epidemic. Rev. Panam. Salud. Publica. 2010, 27, 455–460. [Google Scholar]
- Madrid, H.; Cano, J.; Gené, J.; Bonifaz, A.; Toriello, C.; Guarro, J. Sporothrix globosa, a pathogenic fungus with widespread geographical distribution. Rev. Iberoam. Micol. 2009, 26, 218–222. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; Cruz Choappa, R.; Fernandes, G.F.; de Hoog, G.S.; de Camargo, Z.P. Sporothrix chilensis sp. nov. (Ascomycota: Ophiostomatales), a soil-borne agent of human sporotrichosis with mild-pathogenic potential to mammals. Fungal. Biol. 2016, 120, 246–264. [Google Scholar] [CrossRef]
- Marimon, R.; Cano, J.; Gené, J.; Sutton, D.A.; Kawasaki, M.; Guarro, J. Sporothrix brasiliensis, S. globosa, and S. mexicana, three new Sporothrix species of clinical interest. J. Clin. Microbiol. 2007, 45, 3198–3206. [Google Scholar] [CrossRef]
- Marimon, R.; Gené, J.; Cano, J.; Guarro, J. Sporothrix luriei: A rare fungus from clinical origin. Med. Mycol. 2008, 46, 621–625. [Google Scholar] [CrossRef]
- Chakrabarti, A.; Bonifaz, A.; Gutierrez-Galhardo, M.C.; Mochizuki, T.; Li, S. Global epidemiology of sporotrichosis. Med. Mycol. 2015, 53, 3–14. [Google Scholar] [CrossRef]
- Siew, H. The current status of feline sporotrichosis in Malaysia. Med. Mycol. J. 2017, 58, E107–E113. [Google Scholar] [CrossRef] [PubMed]
- Almeida-Paes, R.; de Oliveira, L.C.; Oliveira, M.M.E.; Gutierrez-Galhardo, M.C.; Nosanchuk, J.D.; Zancopé-Oliveira, R.M. Phenotypic characteristics associated with virulence of clinical isolates from the Sporothrix complex. BioMed. Res. Int. 2015, 2015, 212308. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.P.; Nery, J.A.; de Almeida, A.C.; Ferreira, L.C.; Corte-Real, S.; Conceição-Silva, F. Parinaud’s oculoglandular syndrome associated with Sporothrix schenckii. ID Cases 2014, 1, 38–39. [Google Scholar] [CrossRef]
- Duangkaew, L.; Yurayart, C.; Limsivilai, O.; Chen, C.; Kasorndorkbua, C. Cutaneous sporotrichosis in a stray cat from Thailand. Med. Mycol. Case Rep. 2018, 23, 46–49. [Google Scholar] [CrossRef]
- O’Donnell, K.; Nirenberg, H.I.; Aoki, T.; Cigelnik, E. A Multigene phylogeny of the Gibberella fujikuroi species complex: Detection of additional phylogenetically distinct species. Mycoscience 2000, 41, 61–78. [Google Scholar] [CrossRef]
- Stielow, J.B.; Lévesque, C.A.; Seifert, K.A.; Meyer, W.; Iriny, L.; Smits, D.; Renfurm, R.; Verkley, G.J.; Groenewald, M.; Chaduli, D.; et al. One fungus, which genes? Development and assessment of universal primers for potential secondary fungal DNA barcodes. Persoonia 2015, 35, 242–263. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Kwangsukstith, C.; Vanittanakom, N.; Khanjanasthiti, P.; Uthammachai, C. Cutaneous sporotrichosis in Thailand: First reported case. Mycoses 1990, 33, 513–518. [Google Scholar] [CrossRef]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Zhou, X.; Rodrigues, A.; Feng, P.; Hoog, S. Global ITS diversity in the Sporothrix schenckii complex. Fungal Divers. 2013, 66, 153–165. [Google Scholar] [CrossRef]
- Kamal Azam, N.K.; Selvarajah, G.T.; Santhanam, J.; Abdul Razak, M.F.; Ginsapu, S.J.; James, J.E.; Suetrong, S. Molecular epidemiology of Sporothrix schenkii isolates in Malaysia. Med. Mycol. 2020, 58, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Indoung, S.; Chanchayanon, B.; Chaisut, M.; Buapeth, K.-O.; Morteh, R.; Jantrakajorn, S. Feline sporotrichosis caused by Sporothrix schenckii sensu stricto in Southern Thailand: Phenotypic characterization, molecular identification, and antifungal susceptibility. Med. Mycol. 2022, 60, myac075. [Google Scholar] [CrossRef]
- Matos, A.M.F.; Moreira, L.M.; Barczewski, B.F.; de Matos, L.X.; de Oliveira, J.B.V.; Pimentel, M.I.F.; Almeida-Paes, R.; Oliveira, M.G.; Pinto, T.C.A.; Lima, N.; et al. Identification by MALDI-TOF MS of Sporothrix brasiliensis isolated from a subconjunctival infiltrative lesion in an immunocompetent patient. Microorganisms 2020, 8, 22. [Google Scholar]
- Kano, R.; Okubo, M.; Siew, H.H.; Kamata, H.; Hasegawa, A. Molecular typing of Sporothrix schenckii isolates from cats in Malaysia. Mycoses 2015, 58, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Marimon, R.; Gené, J.; Cano, J.; Trilles, L.; Dos Santos Lazéra, M.; Guarro, J. Molecular phylogeny of Sporothrix schenckii. J. Clin. Microbiol. 2006, 44, 3251–3256. [Google Scholar] [CrossRef] [PubMed]
- Indira, G. In Vitro antifungal susceptibility testing of 5 antifungal agents against dermatophytic species by CLSI (M38-A) micro dilution method. Clin. Microbiol. Open Access 2014, 3, 1–5. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; Della Terra, P.P.; Gremião, I.D.; Pereira, S.A.; Orofino-Costa, R.; de Camargo, Z.P. The threat of emerging and re-emerging pathogenic Sporothrix species. Mycopathologia 2020, 185, 813–842. [Google Scholar] [CrossRef]
- Tovikkai, D.; Maitrisathit, W.; Srisuttiyakorn, C.; Vanichanan, J.; Thammahong, A.; Suankratay, C. Sporotrichosis: The case series in Thailand and literature review in Southeast Asia. Med. Mycol. Case Rep. 2020, 27, 59–63. [Google Scholar] [CrossRef]
- Jessica, N.; Sonia, R.L.; Rodrigo, C.; Isabella, D.F.; Tânia, M.P.; Jeferson, C.; Anna, B.F.; Sandro, A. Diagnostic accuracy assessment of cytopathological examination of feline sporotrichosis. Med. Mycol. 2015, 53, 880–884. [Google Scholar] [CrossRef] [PubMed]
- Schubach, T.M.; de Oliveira Schubach, A.; dos Reis, R.S.; Cuzzi-Maya, T.; Blanco, T.C.; Monteiro, D.F.; Barros, B.M.; Brustein, R.; Zancopé-Oliveira, R.M.; Fialho Monteiro, P.C.; et al. Sporothrix schenckii isolated from domestic cats with and without sporotrichosis in Rio de Janeiro, Brazil. Mycopathologia 2002, 153, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Andrade, E.H.P.; Moreira, S.M.; Paiva, M.T.; Zibaoui, H.M.; Salvato, L.A.; Azevedo, M.I.; Oliveira, C.S.F.; Soares, D.F.M.; Keller, K.M.; Magalhães, S.L.; et al. Characterization of animal sporotrichosis in a highly urbanized area. Comp. Immunol. Microbiol. Infect. Dis. 2021, 76, 101651. [Google Scholar] [CrossRef]
- Macêdo-Sales, P.A.; Souto, S.; Destefani, C.A.; Lucena, R.P.; Machado, R.L.D.; Pinto, M.R.; Rodrigues, A.M.; Lopes-Bezerra, L.M.; Rocha, E.M.S.; Baptista, A.R.S. Domestic feline contribution in the transmission of Sporothrix in Rio de Janeiro State, Brazil: A comparison between infected and non-infected populations. BMC Vet. Res. 2018, 14, 19. [Google Scholar] [CrossRef] [PubMed]
- Schubach, T.M.; Valle, A.C.; Gutierrez-Galhardo, M.C.; Monteiro, P.C.; Reis, R.S.; Zancopé-Oliveira, R.M.; Marzochi, K.B.; Schubach, A. Isolation of Sporothrix schenckii from the nails of domestic cats (Felis catus). Med. Mycol. 2001, 39, 147–149. [Google Scholar] [CrossRef] [PubMed]
- Gremião, I.D.F.; Martins da Silva da Rocha, E.; Montenegro, H.; Carneiro, A.J.B.; Xavier, M.O.; de Farias, M.R.; Monti, F.; Mansho, W.; de Macedo Assunção Pereira, R.H.; Pereira, S.A.; et al. Guideline for the management of feline sporotrichosis caused by Sporothrix brasiliensis and literature revision. Braz. J. Microbiol. 2021, 52, 107–124. [Google Scholar] [CrossRef]
- Freitas, D.F.S.; Valle, A.C.F.d.; da Silva, M.B.T.; Campos, D.P.; Lyra, M.R.; de Souza, R.V.; Veloso, V.G.; Zancopé-Oliveira, R.M.; Bastos, F.I.; Galhardo, M.C.G. Sporotrichosis: An emerging neglected opportunistic infection in HIV-infected patients in Rio de Janeiro, Brazil. PLoS Negl. Trop. Dis. 2014, 8, e3110. [Google Scholar] [CrossRef] [PubMed]
- Souza, E.; Borba, C.; Pereira, S.; Gremião, I.; Langohr, I.; Oliveira, M.; Oliveira, R.; Cunha, C.; Zancope-Oliveira, R.; Monteiro de Miranda, L.; et al. Clinical features, fungal load, coinfections, histological skin changes, and itraconazole treatment response of cats with sporotrichosis caused by Sporothrix brasiliensis. Sci. Rep. 2018, 8, 9074. [Google Scholar] [CrossRef]
- de Miranda, L.H.M.; Silva, J.N.; Gremião, I.D.F.; Menezes, R.C.; Almeida-Paes, R.; Dos Reis, É.G.; de Oliveira, R.V.C.; de Araujo, D.; Ferreiro, L.; Pereira, S.A. Monitoring fungal burden and viability of Sporothrix spp. in skin lesions of cats for predicting antifungal treatment response. J. Fungi 2018, 4, 92. [Google Scholar] [CrossRef]
- Mesa-Arango, A.C.; Del Rocío Reyes-Montes, M.; Pérez-Mejía, A.; Navarro-Barranco, H.; Souza, V.; Zúñiga, G.; Toriello, C. Phenotyping and genotyping of Sporothrix schenckii isolates according to geographic origin and clinical form of Sporotrichosis. J. Clin. Microbiol. 2002, 40, 3004–3011. [Google Scholar] [CrossRef]
- Tang, M.M.; Tang, J.J.; Gill, P.; Chang, C.C.; Baba, R. Cutaneous sporotrichosis: A six-year review of 19 cases in a tertiary referral center in Malaysia. Int. J. Derm. 2012, 51, 702–708. [Google Scholar] [CrossRef]
- Han, H.S.; Kano, R.; Chen, C.; Noli, C. Comparison of two in vitro antifungal sensitivity tests and monitoring during therapy of Sporothrix schenckii sensu stricto in Malaysian cats. Vet. Derm. 2017, 28, 156–e132. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, E.S. Pathogenic roles for fungal melanins. Clin. Microbiol. Rev. 2000, 13, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Rudramurthy, S.M.; Shankarnarayan, S.A.; Hemashetter, B.M.; Verma, S.; Chauhan, S.; Nath, R.; Savio, J.; Capoor, M.; Kaur, H.; Ghosh, A.K.; et al. Phenotypic and molecular characterisation of Sporothrix globosa of diverse origin from India. Braz. J. Microbiol. 2021, 52, 91–100. [Google Scholar] [CrossRef] [PubMed]
- De Carolis, E.; Posteraro, B.; Sanguinetti, M. Old and new insights into Sporothrix schenckii complex biology and identification. Pathogens 2022, 11, 297. [Google Scholar] [CrossRef]
- de Carvalho, J.A.; Pinheiro, B.G.; Hagen, F.; Gonçalves, S.S.; Negroni, R.; Kano, R.; Bonifaz, A.; de Camargo, Z.P.; Rodrigues, A.M. A new duplex PCR assay for the rapid screening of mating-type idiomorphs of pathogenic Sporothrix species. Fungal Biol. 2021, 125, 834–843. [Google Scholar] [CrossRef]
- Kawasaki, M.; Anzawa, K.; Mochizuki, T.; Ishizaki, H. New strain typing method with Sporothrix schenckii using mitochondrial DNA and polymerase chain reaction restriction fragment length polymorphism (PCR–RFLP) technique. J. Dermatol. 2012, 39, 362–365. [Google Scholar] [CrossRef]
- Zhang, Y.; Hagen, F.; Stielow, B.; Rodrigues, A.M.; Samerpitak, K.; Zhou, X.; Feng, P.; Yang, L.; Chen, M.; Deng, S.; et al. Phylogeography and evolutionary patterns in Sporothrix spanning more than 14 000 human and animal case reports. Persoonia 2015, 35, 1–20. [Google Scholar] [CrossRef]
- Sasaki, A.A.; Fernandes, G.F.; Rodrigues, A.M.; Lima, F.M.; Marini, M.M.; dos S. Feitosa, L.; de Melo Teixeira, M.; Felipe, M.S.S.; da Silveira, J.F.; de Camargo, Z.P. Chromosomal polymorphism in the Sporothrix schenckii complex. PLoS ONE 2014, 9, e86819. [Google Scholar]
- de Carvalho, J.A.; Monteiro, R.C.; Hagen, F.; Camargo, Z.P.d.; Rodrigues, A.M. Trends in molecular diagnostics and genotyping tools applied for emerging Sporothrix species. J. Fungi 2022, 8, 809. [Google Scholar] [CrossRef]
- Irinyi, L.; Serena, C.; Garcia-Hermoso, D.; Arabatzis, M.; Desnos-Ollivier, M.; Vu, D.; Cardinali, G.; Arthur, I.; Normand, A.C.; Giraldo, A.; et al. International society of human and animal mycology (ISHAM)-ITS reference DNA barcoding database--the quality controlled standard tool for routine identification of human and animal pathogenic fungi. Med. Mycol. 2015, 53, 313–337. [Google Scholar] [CrossRef] [PubMed]
- Córdoba, S.; Isla, G.; Szusz, W.; Vivot, W.; Hevia, A.; Davel, G.; Canteros, C.E. Molecular identification and susceptibility profile of Sporothrix schenckii sensu lato isolated in Argentina. Mycoses 2018, 61, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, S.; Zaini, F.; Kordbacheh, P.; Safara, M.; Heidari, M. Sporothrix schenckii complex in Iran: Molecular identification and antifungal susceptibility. Med. Mycol. 2016, 54, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Trilles, L.; Fernández-Torres, B.; dos Santos Lazéra, M.; Wanke, B.; de Oliveira Schubach, A.; de Almeida Paes, R.; Inza, I.; Guarro, J. In vitro antifungal susceptibilities of Sporothrix schenckii in two growth phases. Antimicrob. Agents Chemother. 2005, 49, 3952–3954. [Google Scholar] [CrossRef] [PubMed]
- Espinel-Ingroff, A.; Abreu, D.P.B.; Almeida-Paes, R.; Brilhante, R.S.N.; Chakrabarti, A.; Chowdhary, A.; Hagen, F.; Córdoba, S.; Gonzalez, G.M.; Govender, N.P.; et al. Multicenter, international study of MIC/MEC distributions for definition of epidemiological cutoff values for Sporothrix species identified by molecular methods. Antimicrob. Agents Chemother. 2017, 61, e01057-17. [Google Scholar] [CrossRef]
- Espinel-Ingroff, A.; Turnidge, J. The role of epidemiological cutoff values (ECVs/ECOFFs) in antifungal susceptibility testing and interpretation for uncommon yeasts and moulds. Rev. Iberoam. Micol. 2016, 33, 63–75. [Google Scholar] [CrossRef]
- Waller, S.B.; Dalla Lana, D.F.; Quatrin, P.M.; Ferreira, M.R.A.; Fuentefria, A.M.; Mezzari, A. Antifungal resistance on Sporothrix species: An overview. Braz. J. Microbiol. 2021, 52, 73–80. [Google Scholar] [CrossRef]
- Rodrigues, A.; Hoog, S.; Pires, D.; Brihante, R.; Sidrim, J.; Gadelha, M.; Colombo, A.; Camargo, Z. Genetic diversity and antifungal susceptibility profiles in causative agents of sporotrichosis. BMC Infect. Dis. 2014, 14, 219. [Google Scholar] [CrossRef]
- Liang, C.; Shan, Q.; Zhong, J.; Li, W.; Zhang, X.; Wang, J.; Cao, C.; Zeng, Z. Pharmacokinetics and bioavailability of itraconazole oral solution in cats. J. Feline Med. Surg. 2016, 18, 310–314. [Google Scholar] [CrossRef]
- Farowski, F.; Vehreschild, J.J.; Cornely, O.A. Posaconazole: A next-generation triazole antifungal. Future Microbiol. 2007, 2, 231–243. [Google Scholar] [CrossRef]
- Pereira Silveira, C.; Torres-Rodríguez, J.M.; Alvarado-Ramírez, E.; Murciano-Gonzalo, F.; Dolande, M.; Panizo, M.; Reviakina, V. MICs and minimum fungicidal concentrations of amphotericin B, itraconazole, posaconazole and terbinafine in Sporothrix schenckii. J. Med. Microbiol. 2009, 58, 1607–1610. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.C.; Lopes, P.G.; Spader, T.B.; Mahl, C.D.; Tronco-Alves, G.R.; Lara, V.M.; Santurio, J.M.; Alves, S.H. Antifungal susceptibilities of Sporothrix albicans, S. brasiliensis, and S. luriei of the S. schenckii complex identified in Brazil. J. Clin. Microbiol. 2011, 49, 3047–3049. [Google Scholar] [CrossRef] [PubMed]
- Marimon, R.; Serena, C.; Gené, J.; Cano, J.; Guarro, J. In vitro antifungal susceptibilities of five species of Sporothrix. Antimicrob. Agents Chemother. 2008, 52, 732–734. [Google Scholar] [CrossRef] [PubMed]
- Sanchotene, K.O.; Brandolt, T.M.; Klafke, G.B.; Poester, V.R.; Xavier, M.O. In vitro susceptibility of Sporothrix brasiliensis: Comparison of yeast and mycelial phases. Med. Mycol. 2017, 55, 869–876. [Google Scholar] [CrossRef] [PubMed]
- Adams, H.R. Veterinary Pharmacology and Therapeutics; Iowa State University Press: Ames, IA, USA, 2001. [Google Scholar]
- Rossow, J.A.; Queiroz-Telles, F.; Caceres, D.H.; Beer, K.D.; Jackson, B.R.; Pereira, J.G.; Ferreira Gremião, I.D.; Pereira, S.A. A one health approach to combatting Sporothrix brasiliensis: Narrative review of an emerging zoonotic fungal pathogen in South America. J. Fungi 2020, 6, 247. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Source | Origin | Accession Number | ||
|---|---|---|---|---|---|---|
| ITS | CAL | b-TUB | ||||
| S. schenckii | KU20017.9 | Cat | Thailand | MW819850 | MZ215901 | MZ215921 |
| S. schenckii | KU2001810 | Cat | Thailand | MW819852 | MZ215902 | MZ215922 |
| S. schenckii | KU20019.3 | Cat | Thailand | MW819851 | MZ215903 | MZ215923 |
| S. schenckii | KU20020.45 | Cat | Thailand | MW857103 | MZ215906 | MZ215926 |
| S. schenckii | KU20020.376 | Cat | Thailand | MW857105 | MZ215907 | MZ215927 |
| S. schenckii | KU20020.377 | Cat | Thailand | MW857106 | MZ215908 | MZ215928 |
| S. schenckii | KU20020.378 | Cat | Thailand | MW857108 | MZ215909 | MZ215929 |
| S. schenckii | KU20020.379 | Cat | Thailand | MW857109 | MZ215910 | MZ215930 |
| S. schenckii | KU20020.380 | Cat | Thailand | MW857111 | MZ215911 | MZ215931 |
| S. schenckii | KU20020.381 | Cat | Thailand | MW857112 | MZ215912 | MZ215932 |
| S. schenckii | KU20020.382 | Cat | Thailand | MW857113 | MZ215913 | MZ215933 |
| S. schenckii | KU20020.383 | Human | Thailand | MW857114 | MZ215914 | MZ215934 |
| S. schenckii | KU20020.384 | Cat | Thailand | MW857115 | MZ215915 | MZ215935 |
| S. schenckii | KU20020.385 | Cat | Thailand | MW857126 | MZ215916 | MZ215936 |
| S. schenckii | KU20020.386 | Cat | Thailand | MW857128 | MZ215917 | MZ215937 |
| S. schenckii | KU20021.02 | Cat | Thailand | MW857129 | MZ215919 | MZ215939 |
| S. schenckii | KU20021.05 | Cat | Thailand | MZ882155 | OK065908 | OK065918 |
| S. schenckii | KU20021.06 | Cat | Thailand | MZ882257 | OK065909 | OK065919 |
| S. schenckii | KU20021.07 | Cat | Thailand | MZ882288 | OK065910 | OK065920 |
| S. schenckii | KU20021.08 | Cat | Thailand | MZ882289 | OK065911 | OK065921 |
| S. schenckii | KU20021.10 | Cat | Thailand | MZ882290 | OK065912 | OK065922 |
| S. schenckii | KU20021.11 | Cat | Thailand | MZ882291 | OK065913 | OK065923 |
| S. schenckii | KU20021.12 | Cat | Thailand | MZ882292 | OK065914 | OK065924 |
| S. schenckii | KU20021.13 | Cat | Thailand | MZ882293 | OK065915 | OK065925 |
| S. schenckii | KU20021.40 | Cat | Thailand | MZ882294 | OK065916 | OK065926 |
| S. schenckii | KU20021.42 | Cat | Thailand | MZ882295 | OK065917 | OK065927 |
| S. schenckii | UTHSC 04-1064 | Human | USA | - | AM399014 | AM747285 |
| S. schenckii | UTHSC 04-771 | Human | USA | - | AM398985 | AM747287 |
| S. schenckii | UdeA_0001 | Human | Columbia | - | ON060845 | MG953922 |
| S. schenckii | UdeA_0007 | ND | Columbia | - | ON060848 | ON023043 |
| S. schenckii | UdeA_0008 | ND | Columbia | - | ON060849 | ON023044 |
| S. schenckii | UdeA_22 | ND | Columbia | - | ON060860 | MG953936 |
| S. schenckii | UdeA_220 | Human | Columbia | - | ON060861 | MG953935 |
| S. schenckii | UdeA 1023 | Human | Columbia | - | ON060850 | MG953933 |
| S. schenckii | UdeA 1027 | ND | Columbia | - | ON060851 | MG953934 |
| S. schenckii | UdeA_3017 | Human | Columbia | - | ON060863 | MG953916 |
| S. schenckii | UdeA_4015 | ND | Columbia | - | ON060864 | ON023050 |
| S. schenckii | UdeA_4023 | ND | Columbia | - | ON060865 | MG953917 |
| S. schenckii | UdeA_4047 | ND | Columbia | - | ON060866 | ON023051 |
| S. schenckii | UdeA_6450 | Human | Columbia | - | ON060870 | MG953918 |
| S. schenckii | UdeA_6514 | ND | Columbia | - | ON060871 | ON023052 |
| S. schenckii | UdeA_7011 | ND | Columbia | - | ON060872 | ON023053 |
| S. schenckii | UdeA_7027 | Human | Columbia | - | ON060873 | MH174665 |
| S. schenckii | UdeA_8004 | Human | Columbia | - | ON060874 | MG953919 |
| S. schenckii | UdeA_9043 | Human | Columbia | - | ON060876 | MH174668 |
| S. schenckii | UdeA 10937 | Human | Columbia | - | ON060852 | MG953927 |
| S. schenckii | UdeA 11024 | Human | Columbia | - | ON060853 | MG953925 |
| S. schenckii | UdeA 15565 | Human | Columbia | - | ON060857 | MG953924 |
| S. schenckii | UdeA 15741 | Human | Columbia | - | ON060858 | MG953920 |
| S. schenckii | UdeA 20008 | Human | Columbia | - | ON060859 | MG953921 |
| S. schenckii | PSUV_SS18-1 | Cat | Thailand | - | ON211523 | ON211485 |
| S. schenckii | PSUV_SS18-2 | Cat | Thailand | - | ON211527 | ON211489 |
| S. schenckii | PSUV_SS18-3 | Cat | Thailand | - | ON211510 | ON211472 |
| S. schenckii | PSUV_SS18-4 | Cat | Thailand | - | ON211508 | ON211470 |
| S. schenckii | PSUV_SS18-6 | Cat | Thailand | - | ON211517 | ON211479 |
| S. schenckii | PSUV_SS18-7 | Cat | Thailand | - | ON211511 | ON211473 |
| S. schenckii | PSUV_SS18-8 | Cat | Thailand | - | ON211520 | ON211482 |
| S. schenckii | PSUV_SS18-9 | Cat | Thailand | - | ON211519 | ON211481 |
| S. schenckii | PSUV_SS18-10 | Cat | Thailand | - | ON211529 | ON211491 |
| S. schenckii | PSUV_SS19-1 | Cat | Thailand | - | ON211515 | ON211477 |
| S. schenckii | PSUV_SS19-2 | Cat | Thailand | - | ON211513 | ON211475 |
| S. schenckii | PSUV_SS19-3 | Cat | Thailand | - | ON211514 | ON211476 |
| S. schenckii | PSUV_SS19-5 | Cat | Thailand | - | ON211542 | ON211504 |
| S. schenckii | PSUV_SS19-6 | Cat | Thailand | - | ON211530 | ON211492 |
| S. schenckii | PSUV_SS19-7 | Cat | Thailand | - | ON211540 | ON211502 |
| S. schenckii | PSUV_SS19-8 | Cat | Thailand | - | ON211524 | ON211486 |
| S. schenckii | PSUV_SS19-9 | Cat | Thailand | - | ON211518 | ON211480 |
| S. schenckii | PSUV_SS19-10 | Cat | Thailand | - | ON211529 | ON211491 |
| S. schenckii | PSUV_SS20-3 | Cat | Thailand | - | ON211516 | ON211478 |
| S. schenckii | PSUV_SS20-4 | Cat | Thailand | - | ON211543 | ON211505 |
| S. schenckii | PSUV_SS20-5 | Cat | Thailand | - | ON211525 | ON211487 |
| S. schenckii | PSUV_SS20-6 | Cat | Thailand | - | ON211534 | ON211496 |
| S. schenckii | PSUV_SS20-7 | Cat | Thailand | - | ON211539 | ON211501 |
| S. schenckii | PSUV_SS20-8 | Cat | Thailand | - | ON211521 | ON211483 |
| S. schenckii | PSUV_SS20-9 | Cat | Thailand | - | ON211522 | ON211484 |
| S. schenckii | PSUV_SS21-1 | Cat | Thailand | - | ON211535 | ON211497 |
| S. schenckii | PSUV_SS21-2 | Cat | Thailand | - | ON211537 | ON211499 |
| S. schenckii | PSUV_SS21-3 | Cat | Thailand | - | ON211538 | ON211500 |
| S. schenckii | PSUV_SS21-5 | Cat | Thailand | - | ON211544 | ON211506 |
| S. schenckii | PSUV_SS21-6 | Cat | Thailand | - | ON211545 | ON211507 |
| S. schenckii | PSUV_SS20-10 | Cat | Thailand | - | ON211541 | ON211503 |
| S. schenckii | PSUV_SS20-11 | Cat | Thailand | - | ON211526 | ON211488 |
| S. schenckii | PSUV_SS20-12 | Cat | Thailand | - | ON211528 | ON211490 |
| S. schenckii | PSUV_SS20-13 | Cat | Thailand | - | ON211512 | ON211474 |
| S. schenckii | PSUV_SS20-14 | Cat | Thailand | - | ON211531 | ON211493 |
| S. schenckii | PSUV_SS20-15 | Cat | Thailand | - | ON211533 | ON211495 |
| S. schenckii | PSUV_SS20-16 | Cat | Thailand | - | ON211532 | ON211494 |
| S. schenckii | PSUV_SS20-17 | Cat | Thailand | - | ON211536 | ON211498 |
| S. schenckii | NBRC_8158 | ND | Japan | - | AM117438 | AM116910 |
| S. schenckii | IHEM_3774 | Human | Colombia | - | AM117447 | AM116921 |
| S. schenckii | IHEM_3787 | ND | South Africa | - | AM117435 | AM116925 |
| S. schenckii | IHEM_15502 | Human | Peru | - | AM117427 | AM116927 |
| S. schenckii | IHEM_15503 | Human | Peru | - | AM117433 | AM116930 |
| S. schenckii | IHEM_15508 | Human | Peru | - | AM117443 | AM116924 |
| S. schenckii | IHEM_15511 | Human | Peru | - | AM117440 | AM116917 |
| S. schenckii | IHEM_15477 | Human | Bolivia | - | AM117444 | AM116916 |
| S. schenckii | IHEM_15486 | Human | Peru | - | AM117432 | AM116929 |
| S. schenckii | IHEM_15489 | Human | Peru | - | AM117430 | AM116926 |
| S. schenckii | IHEM_15499 | Human | Peru | - | AM117434 | AM116928 |
| S. schenckii | FMR_8604 | Human | Peru | - | AM117429 | AM116914 |
| S. schenckii | FMR_8605 | Human | Peru | - | AM117442 | AM116923 |
| S. schenckii | FMR_8606 | Human | Peru | - | AM117431 | AM116913 |
| S. schenckii | FMR_8608 | Human | Peru | - | AM117441 | AM116919 |
| S. schenckii | FMR_8609 | Human | Peru | - | AM117439 | AM116918 |
| S. schenckii | FMR_8677 | Human | Argentina | - | AM117436 | AM116915 |
| S. schenckii | FMR_8678 | Human | Argentina | - | AM117446 | AM116920 |
| S. schenckii | FMR_8679 | Human | Argentina | - | AM117445 | AM116922 |
| S. schenckii | EH-195 | ENV | Mexico | - | ON060840 | ON023042 |
| S. schenckii | EH-197 | Human | Mexico | - | ON060841 | MH174664 |
| S. schenckii | EH-230 | Human | Guatemala | - | ON060842 | MG953930 |
| S. schenckii | EH-251 | ENV | Mexico | - | ON060843 | MG953938 |
| S. schenckii | EH-254 | ENV | Mexico | - | ON060844 | MG953937 |
| S. schenckii | 1099-18 | Human | USA | - | JF313360 | AXCR01000005 |
| S. schenckii | VSMU-SS1702 | Human | Thailand | MG270182 | - | - |
| S. schenckii | NUBS14005 | Cat | Malaysia | LC012513 | - | - |
| S. schenckii | NUBS14007 | Cat | Malaysia | LC012518 | - | - |
| S. schenckii | NUBS14009 | Cat | Malaysia | LC012520 | - | - |
| S. schenckii | NUBS14013 | Cat | Malaysia | LC012599 | - | - |
| S. schenckii | NUBS14016 | Cat | Malaysia | LC012602 | - | - |
| S. schenckii | NUBS14018 | Cat | Malaysia | LC012604 | - | - |
| S. schenckii | NUBS14019 | Cat | Malaysia | LC012605 | - | - |
| S. schenckii | FMR 8609 | Human | Peru | - | AM117439 | AM116918 |
| S. schenckii | CBS 359.36T | Human | USA | KP017100 | AM117437 | AM116911 |
| S. schenckii | CBS 444.67 | ND | Mozambique | KP017099 | - | - |
| S. schenckii | CBS 130097 | Human | Bolivia | KC113218 | - | - |
| S. schenckii | CBS 130098 | Human | Peru | KP017091 | - | - |
| S. schenckii | ATCC 10268 | Human | USA | AB122038 | - | - |
| S. schenckii | CBS 130111 | Human | Colombia | KC113219 | - | - |
| S. schenckii | CBS 130103 | Human | Argentina | KC113222 | - | - |
| S. schenckii | CBS 130114 | Human | Peru | KC113221 | - | - |
| S. schenckii | CBS 130101 | Human | Peru | KP017095 | - | - |
| S. schenckii | CBS 130112 | Human | Peru | KC113223 | - | - |
| S. schenckii | CBS 211.61 | ND | South Africa | KP017093 | - | - |
| S. schenckii | CBS 938.72 | Human | France | KP017094 | - | - |
| S. schenckii | CBS 130100 | Human | Argentina | KC113217 | - | - |
| S. globosa | CBS 130117 | Human | Japan | KC113229 | - | - |
| S. globosa | CBS 120340 T | Human | Spain | KC113229 | KP101459. | WOEH00000000 |
| S. globosa | CBS 129720 | ND | China | - | KP101463 | KC113237 |
| S. globosa | CBS 129722 | Human | China | - | KP101464 | KC113236 |
| S. globosa | CBS 129724 | Human | China | - | KP101466 | KC113238 |
| S. globosa | UdeA 0003 | ND | Columbia | - | ON060846 | MG953948 |
| S. globosa | UdeA 0004 | ND | Columbia | - | ON060847 | MG953945 |
| S. globosa | UdeA 64 | ND | Columbia | - | ON060869 | MG953946 |
| S. globosa | UdeA 5048 | ND | Columbia | - | ON060867 | MG953939 |
| S. globosa | UdeA 6004 | ND | Columbia | - | ON060868 | MG953940 |
| S. globosa | UdeA 8029 | ND | Columbia | - | ON060875 | MH174666 |
| S. globosa | UdeA 9051 | ND | Columbia | - | ON060877 | MG953941 |
| S. globosa | UdeA 12926 | ND | Columbia | - | ON060855 | MH174667 |
| S. globosa | NCCPF 220126 | Human | India | - | MN243809 | KX881724 |
| S. globosa | NCCPF 220127 | Human | India | - | MN243810 | KX881725 |
| S. globosa | NCCPF 220129 | Human | India | - | MN243811 | KX881701 |
| S. globosa | NCCPF 220135 | Human | India | - | MN243812 | KX881721 |
| S. globosa | NCCPF 220136 | Human | India | - | MN243813 | KX881722 |
| S. globosa | NCCPF 220146 | Human | India | - | MN243822 | KX881719 |
| S. globosa | NCCPF 220149 | Human | India | - | MN243823 | MN257636 |
| S. luriei | CBS 937.72 T | Human | South Africa | AB128012 | - | - |
| S. brasiliensis | CBS 120339 T | Human | Brazil | KP017087 | - | - |
| S. brasiliensis | CBS 130106 | Human | Brazil | KC113212 | - | - |
| S. brasiliensis | CBS 130109 | Human | Brazil | KC113213 | - | - |
| S. brasiliensis | CF2018 | ND | Argentina | - | MK850446 | MK850450 |
| S. brasiliensis | CHS2018 | ND | Argentina | - | MK850448 | MK850452 |
| S. brasiliensis | CN2018 | ND | Argentina | - | MK850447 | MK850451 |
| S. brasiliensis | IPEC_16919 | Human | Brazil | - | AM116898 | AM116934 |
| S. pallida | CBS 150.87 T | ENV | Germany | EF127879 | - | - |
| S. chilensis | CBS 139891 T | Human | Chile | KP711811 | - | - |
| Characteristic | Number (%) | |
|---|---|---|
| Case number | 38 | |
| Geographic origin | Bangkok | 32 (85) |
| Nonthaburi | 2 (5) | |
| Pathum Thani | 2 (5) | |
| Ayutthaya | 2 (5) | |
| Age (years) | mean (±standard deviation) | 3.96 (±3.93) |
| Youngest | 6 M | |
| Oldest | 14 Y 10 M | |
| Sex | Male | 23 (61) |
| Female | 15 (39) | |
| Breed | Domestic short hair | 36 (95) |
| Purebred | 2 (5) | |
| Castration | Yes | 22 (58) |
| No | 16 (42) | |
| Cat rearing | Indoor only | 9 (24) |
| Outdoor access | 29 (76) | |
| Possible transmission | Fights with other cats | 19 (50) |
| None | 11 (29) | |
| Unknown | 8 (21) | |
| FeLV/FIV infected | Yes | 2 (5) |
| No | 21 (55) | |
| No data | 15 (40) | |
| Number of lesions | 1 | 9 (24) |
| 2 | 11 (29) | |
| ≥3 | 18 (47) | |
| The form of lesion | Ulcer | 35 (92) |
| Granulomatous nodular | 3 (8) | |
| Other organ involvement | Respiratory system | 8 (21) |
| Eyes lids | 2 (5) | |
| Bone | 1 (3) | |
| None | 27 (71) | |
| Therapeutics | Antifungal drugs | 31 (82) |
| Combined surgy and anti-fungal drugs | 7 (18) | |
| Outcome | Recovery | 20 (53) |
| Stable disease and continue treatment | 4 (10) | |
| death | 14 (37) |
| Growth Condition | Growth (Number, %) and Average Colony Diameter (mm) | |||
|---|---|---|---|---|
| No Growth | Mold | Yeast-Like | Yeast | |
| 30 °C | ||||
| SDA | 0 | 5, 19 (6.99) | 21, 81 (3.41) | 0 |
| PDA | 0 | 5, 19 (9.34) | 21, 81 (4.11) | 0 |
| BHIA | 0 | 0 | 13, 50 (4.11) | 13, 50 (4.15) |
| 35 °C | ||||
| SDA | 0 | 1, 4 (0.74) | 22, 85 (1.74) | 3, 12 (2.24) |
| PDA | 0 | 1, 4 (1.77) | 19, 73 (1.97) | 6, 23 (2.15) |
| BHIA | 0 | 0 | 4, 15 (3.91) | 22, 85 (3.33) |
| 37 °C | ||||
| SDA | 8 (31) | 0 | 6, 23 (0.39) | 12, 46 (0.48) |
| PDA | 0 | 8, 31 (0.18) | 10, 38 (0.38) | |
| BHIA | 0 | 3, 11 (0.46) | 15, 58 (0.98) | |
| Agents | Yeast Phase | Mold Phase | ||||||
|---|---|---|---|---|---|---|---|---|
| MIC Range | Mode | MIC50 | MIC90 | MIC Range | Mode | MIC50 | MIC90 | |
| AMB | 0.25 to 4 | 2 | 1 | 4 | 0.25 to 4 | 0.5 | 0.5 | 4 |
| ITR | 0.25 to >32 | 1 | 1 | 4 | 0.0625 to ≥32 | 0.5 | 0.5 | 4 |
| POS | 0.25 to >32 | 0.5 | 0.5 | 8 | 0.0625 to ≥32 | 0.25 | 0.25 | 8 |
| TERB | 0.0156 to 2 | 1 | 0.5 | 2 | 0.0312 to 4 | 1 | 0.75 | 2 |
| KET | 0.0039 to 1 | 0.5 | 0.25 | 0.5 | 0.0039 to 1 | 0.25 | 0.25 | 0.5 |
| MCZ | 0.0039 to 1 | 0.5 | 0.25 | 0.5 | 0.0039 to 1 | 0.25 | 0.25 | 0.5 |
| FLU | 32 to >256 | >256 | >256 | >256 | 32 to >256 | >256 | >256 | >256 |
| GRI | 0.0625 to ≥32 | 4 | 4 | 32 | 0.0625 to ≥32 | 4 | 4 | 16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yingchanakiat, K.; Limsivilai, O.; Sunpongsri, S.; Niyomtham, W.; Lugsomya, K.; Yurayart, C. Phenotypic and Genotypic Characterization and Antifungal Susceptibility of Sporothrix schenckii sensu stricto Isolated from a Feline Sporotrichosis Outbreak in Bangkok, Thailand. J. Fungi 2023, 9, 590. https://doi.org/10.3390/jof9050590
Yingchanakiat K, Limsivilai O, Sunpongsri S, Niyomtham W, Lugsomya K, Yurayart C. Phenotypic and Genotypic Characterization and Antifungal Susceptibility of Sporothrix schenckii sensu stricto Isolated from a Feline Sporotrichosis Outbreak in Bangkok, Thailand. Journal of Fungi. 2023; 9(5):590. https://doi.org/10.3390/jof9050590
Chicago/Turabian StyleYingchanakiat, Kanokporn, Orawan Limsivilai, Supita Sunpongsri, Waree Niyomtham, Kittitat Lugsomya, and Chompoonek Yurayart. 2023. "Phenotypic and Genotypic Characterization and Antifungal Susceptibility of Sporothrix schenckii sensu stricto Isolated from a Feline Sporotrichosis Outbreak in Bangkok, Thailand" Journal of Fungi 9, no. 5: 590. https://doi.org/10.3390/jof9050590