
Adding Metal Ions to the Bacillus mojavensis D50 Promotes Biofilm Formation and Improves Ability of Biocontrol

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Medium, Culture Conditions and Plant Materials
2.2. Preparation of the B. mojavensis D50 Suspension
2.3. Semiquantitative Evaluation of the B. mojavensis D50 Biofilm
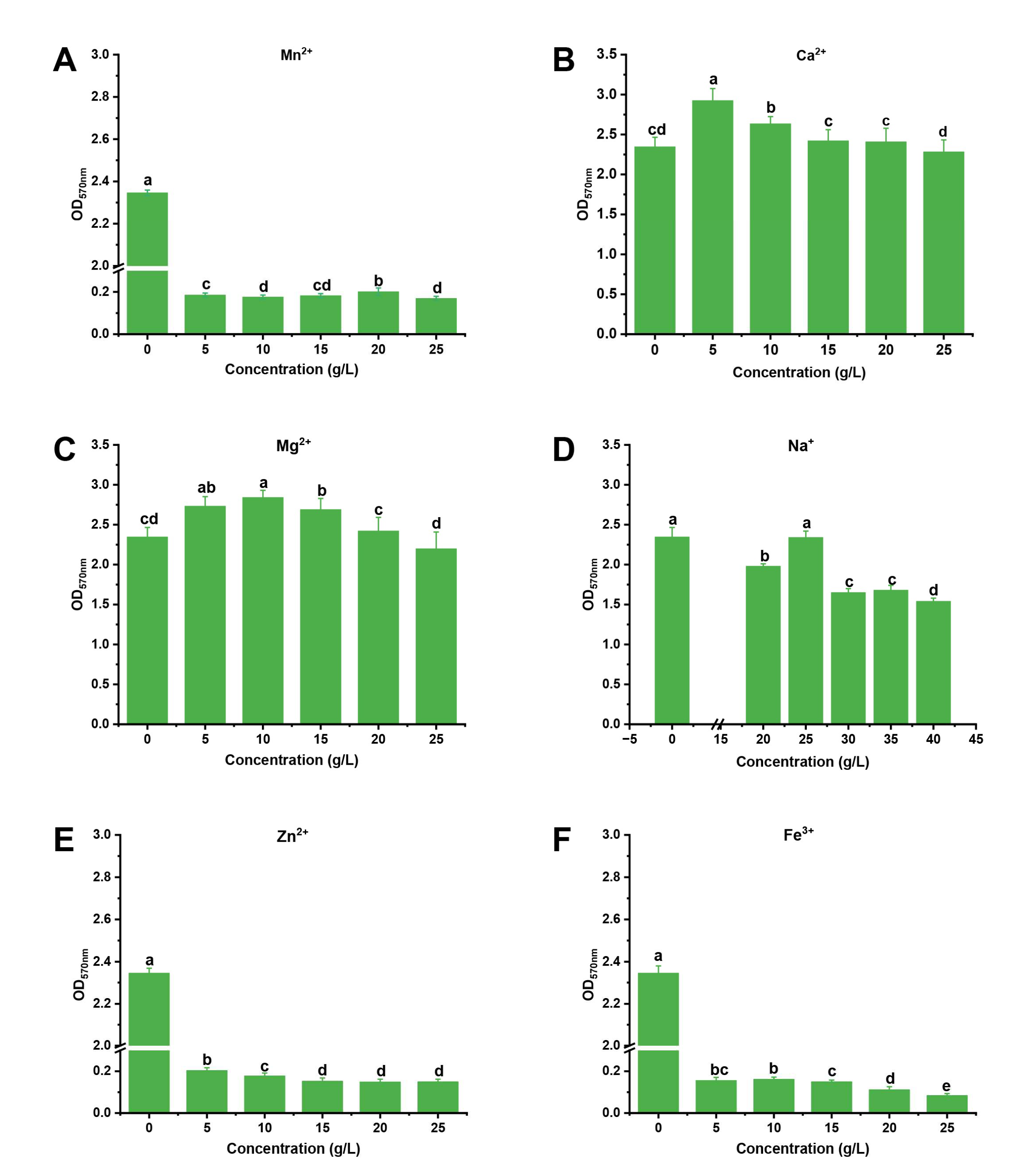
2.4. Screening of the Optimal Metal Ions and Conditions for Biofilm Formation
2.4.1. Determination of the Effect of Metal Ions on Biofilm Formation by a Single Factor Experiment
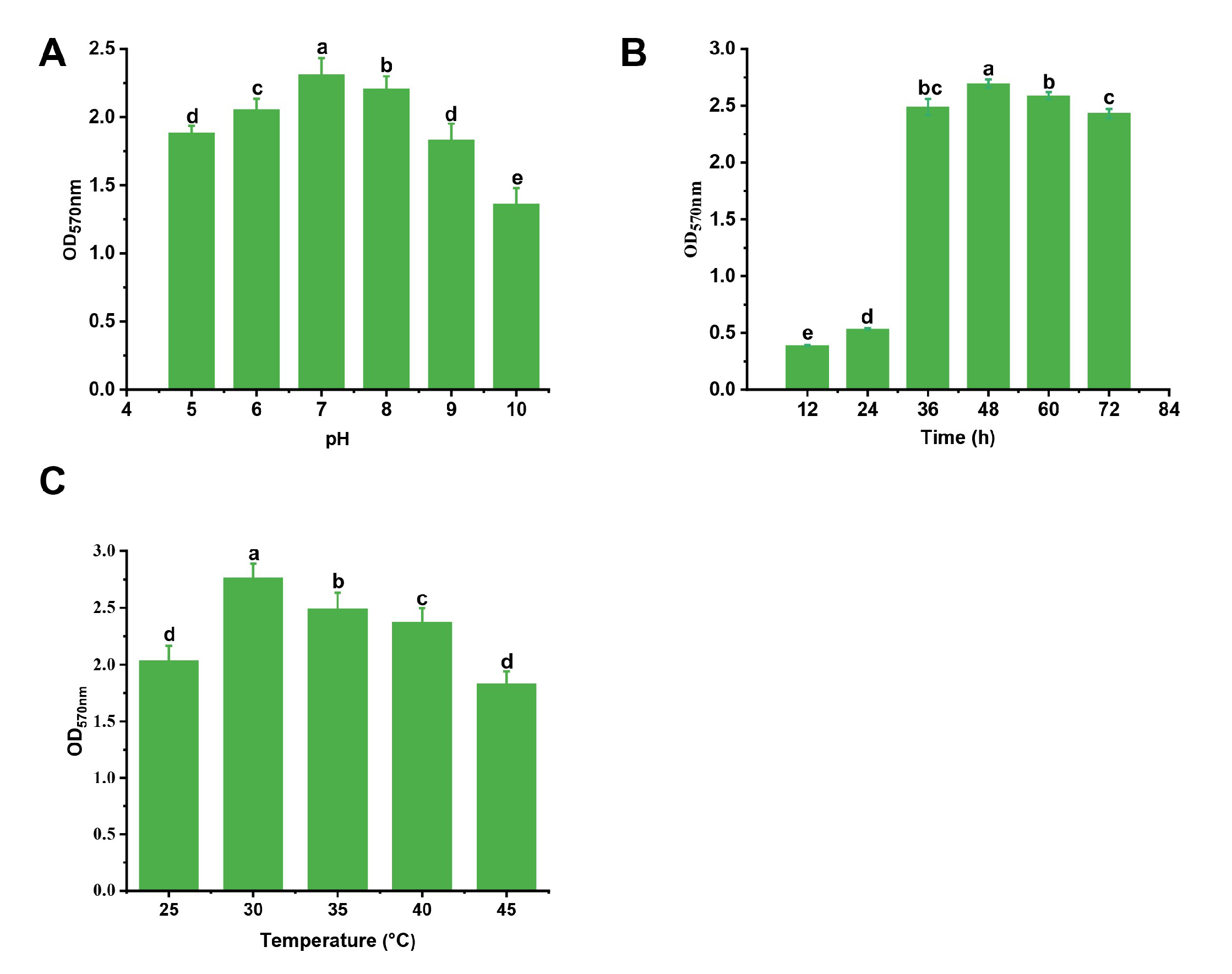
2.4.2. Determination of the Effect of Culture Conditions on Biofilm Formation by a Single Factor Experiment
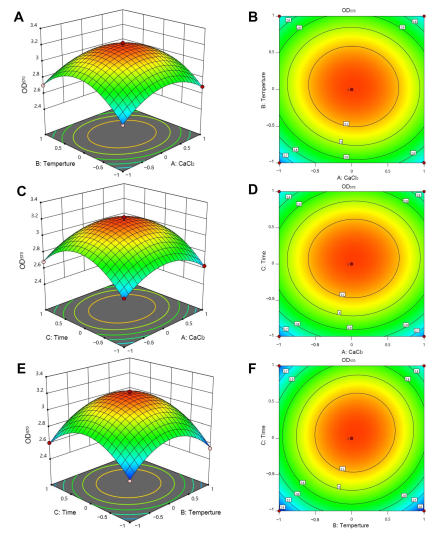
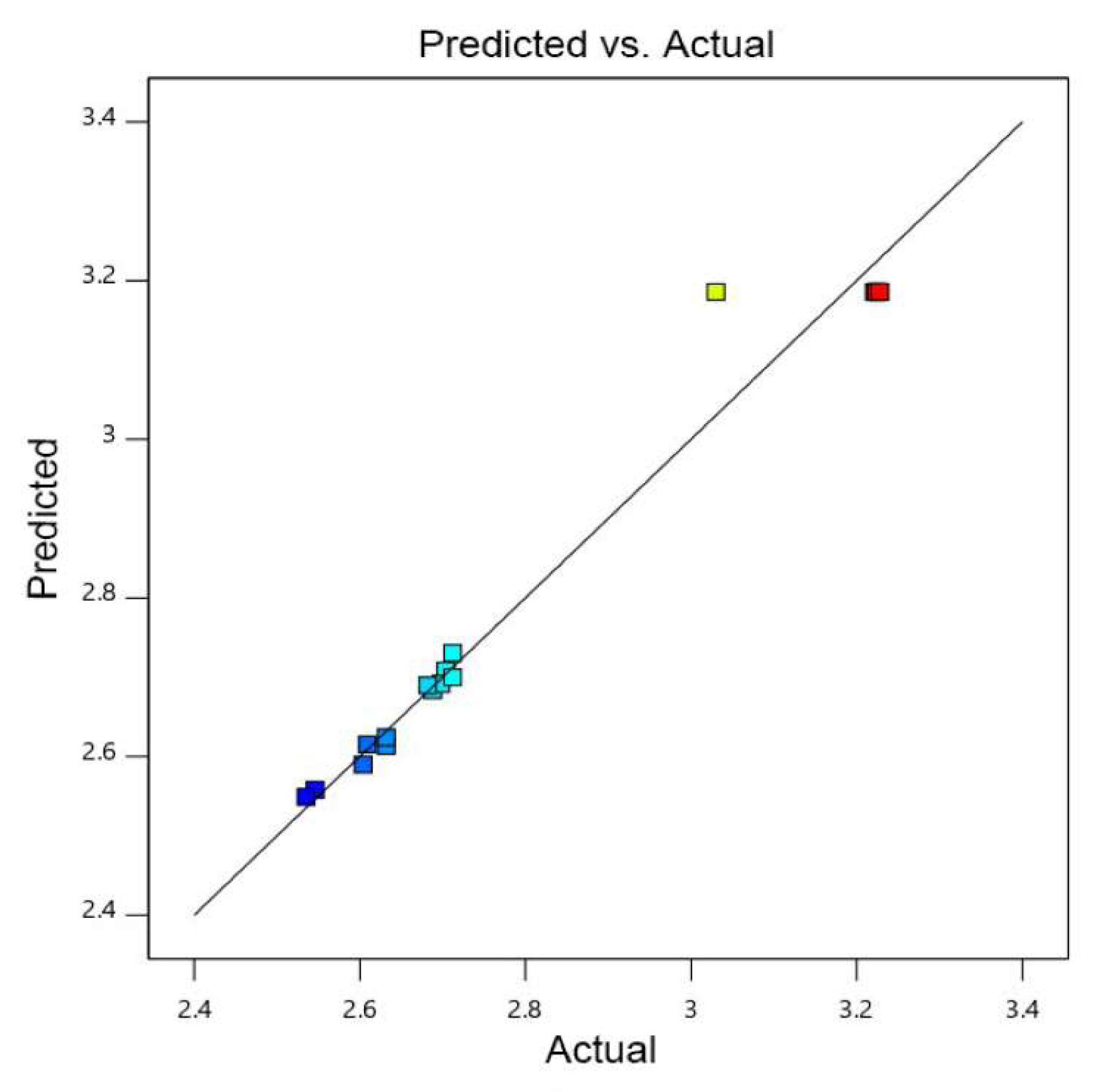
2.4.3. Determination of the Optimal Amount of Metal Ions and Culture Conditions for Biofilm Formation Using a Box-Behnken Design
2.5. Effect of Metal Ions and Culture Conditions on the Biocontrol Characteristics of B. mojavensis D50 against B. cinerea In Vitro
2.6. B. mojavensis D50 Biofilm-Related Analysis of Differentially Expressed Genes
2.7. Effect of Metal Ions and Culture Conditions on the Biocontrol Characteristics of B. mojavensis D50 against B. cinerea In Vivo
2.8. Effect of B. mojavensis D50 on Soil Enzyme Activities before and after Optimization
2.9. Statistical Analysis
3. Results
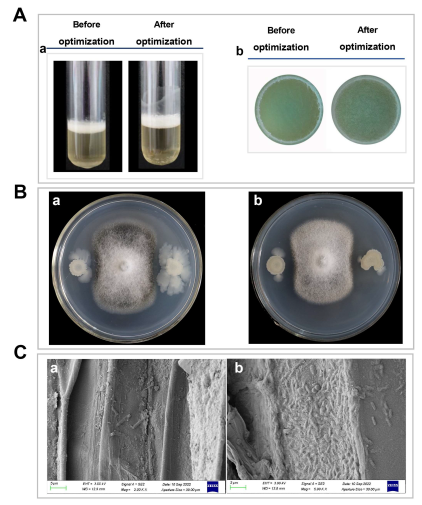
3.1. Screening of the Optimal Metal Ions and Culture Conditions for Biofilm Formation
3.2. Determination of Antagonistic Abilities and Colonization Capacity Differences
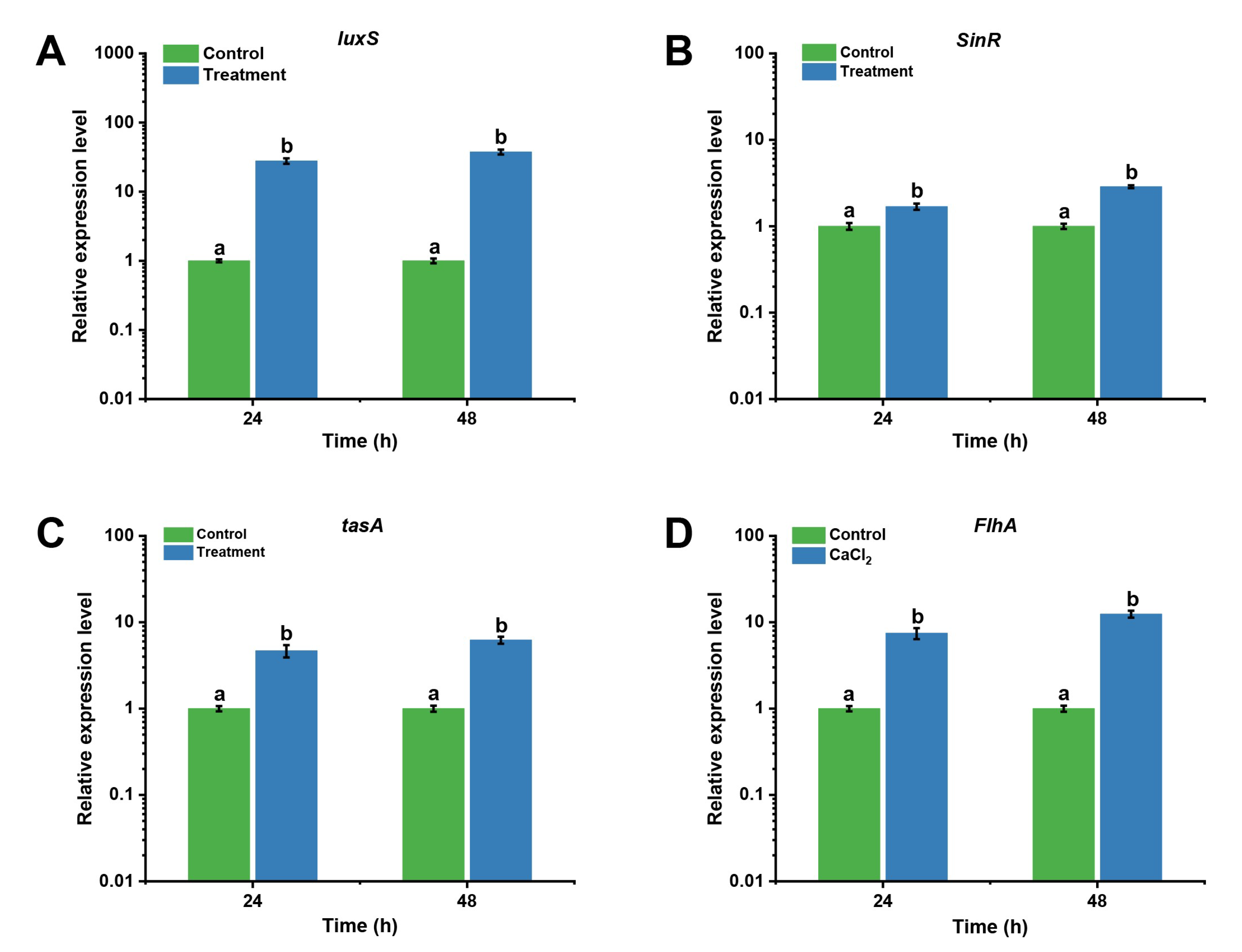
3.3. B. mojavensis D50 Biofilm-Related Gene Differential Expression Analysis
3.4. Effect of Metal Ions and Culture Conditions on the Biocontrol Characteristics of B. mojavensis D50 against B. cinerea In Vivo
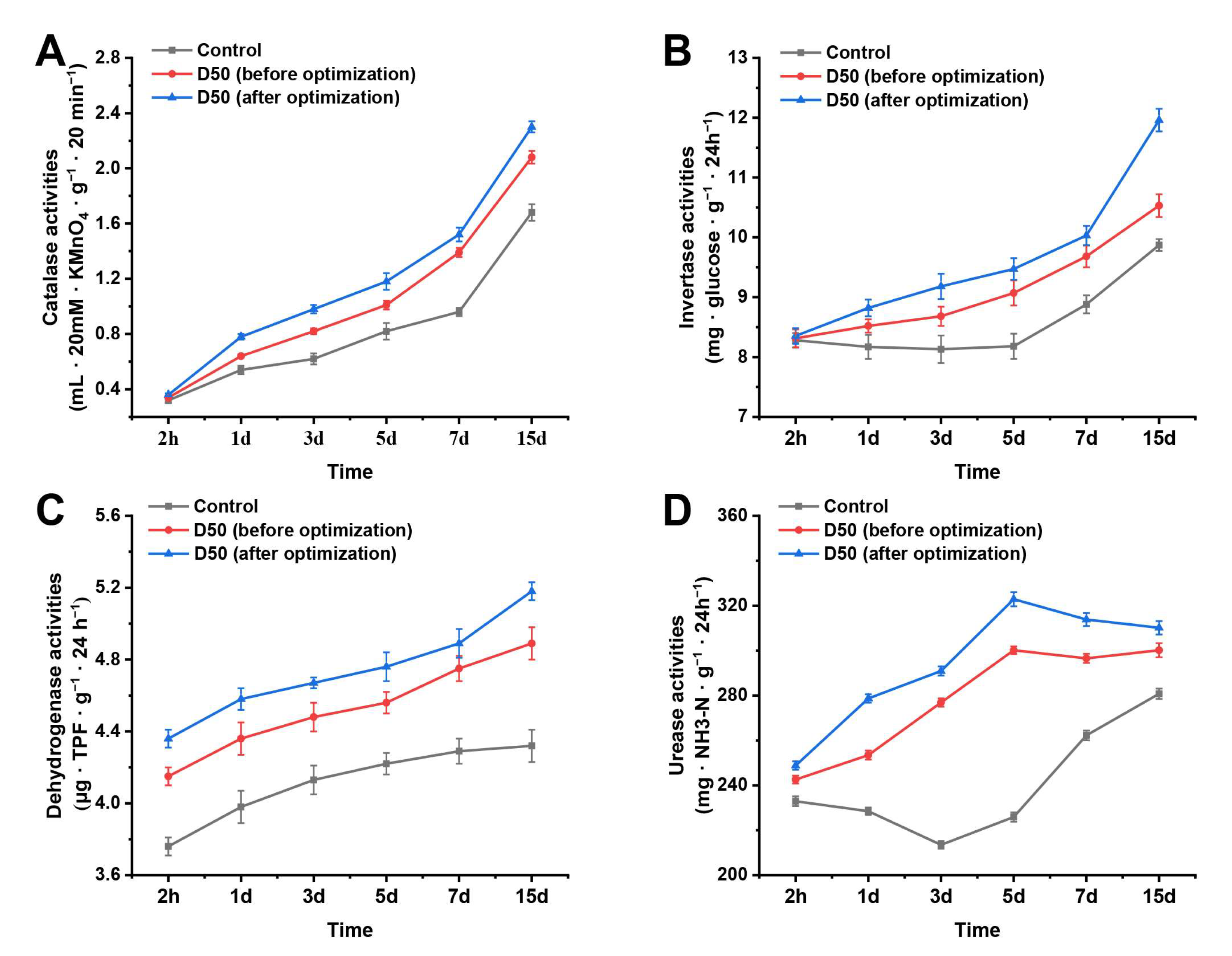
3.5. Effect of B. mojavensis D50 (before and after Optimization) on Soil Enzyme Activities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bu, S.; Munir, S.; He, P.; Li, Y.; Wu, Y.; Li, X.; Kong, B.; He, P.; He, Y. Bacillus subtilis L1-21 as a biocontrol agent for postharvest gray mold of tomato caused by Botrytis cinerea. Biol. Control 2021, 157, 104568. [Google Scholar] [CrossRef]
- Hong, J.K.; Sook Jo, Y.; Jeong, D.H.; Woo, S.M.; Park, J.Y.; Yoon, D.J.; Lee, Y.H.; Choi, S.H.; Park, C.J. Vapours from plant essential oils to manage tomato grey mould caused by Botrytis cinerea. Fungal Biol. 2023, 127, 985–996. [Google Scholar] [CrossRef]
- Suresh, P.; Shanmugaiah, V.; Rajakrishnan, R.; Muthusamy, K.; Ramamoorthy, V. Pseudomonas fluorescens VSMKU3054 mediated induced systemic resistance in tomato against Ralstonia solanacearum. Physiol. Mol. Plant Pathol. 2022, 199, 101836. [Google Scholar] [CrossRef]
- Guo, J.; Xu, Y.; Liang, S.; Zhou, Z.; Zhang, C.; Li, K.; Peng, X.; Qin, S.; Xing, K. Antifungal activity of volatile compounds from Bacillus tequilensis XK29 against Botrytis cinerea causing gray mold on cherry tomatoes. Postharvest Biol. Technol. 2023, 198, 112239. [Google Scholar] [CrossRef]
- Zheng, L.; Gu, X.; Xiao, Y.; Wang, S.; Liu, L.; Pan, H.; Zhang, H. Antifungal activity of Bacillus mojavensis D50 against Botrytis cinerea causing postharvest gray mold of tomato. Sci. Hortic. 2023, 312, 111841. [Google Scholar] [CrossRef]
- Mali, H.; Shah, C.; Raghunandan, B.H.; Prajapati, A.S.; Patel, D.H.; Trivedi, U.; Subramanian, R.B. Organophosphate pesticides an emerging environmental contaminant: Pollution, toxicity, bioremediation progress, and remaining challenges. J. Environ. Sci. 2023, 127, 234–250. [Google Scholar] [CrossRef]
- Yang, X.; Wang, Y.; Jiang, H.; Song, R.; Liu, Y.; Guo, H.; Meng, D. Antimicrobial peptide CB-M exhibits direct antifungal activity against Botrytis cinerea and induces disease resistance to gray mold in cherry tomato fruit. Postharvest Biol. Technol. 2023, 196, 112184. [Google Scholar] [CrossRef]
- Chen, J.; Wei, X.; Lu, X.; Ming, R.; Huang, D.; Yao, Y.; Li, L.; Huang, R. Burkholderia cenocepacia ETR-B22 volatile organic compounds suppress postharvest grey mould infection and maintain aroma quality of tomato fruit. LWT 2022, 165, 113715. [Google Scholar] [CrossRef]
- Li, S.; Xiao, Q.; Yang, H.; Huang, J.; Li, Y. Characterization of a new Bacillus velezensis as a powerful biocontrol agent against tomato gray mold. Pestic. Biochem. Physiol. 2022, 187, 105199. [Google Scholar] [CrossRef]
- Zhao, Q.; Shi, Y.; Xu, C.; Jiang, Z.; Liu, J.; Sui, Y.; Zhang, H. Control of postharvest blue and gray mold in kiwifruit by Wickerhamomyces anomalus and its mechanism of antifungal activity. Postharvest Biol. Technol. 2023, 201, 112345. [Google Scholar] [CrossRef]
- Geng, L.; Fu, Y.; Peng, X.; Yang, Z.; Zhang, M.; Song, Z.; Guo, N.; Chen, S.; Chen, J.; Bai, B.; et al. Biocontrol potential of Trichoderma harzianum against Botrytis cinerea in tomato plants. Biol. Control. 2022, 174, 105019. [Google Scholar] [CrossRef]
- Wang, B.; Wang, C.; Hu, Y. Sorption behavior of Pb(II) onto polyvinyl chloride microplastics affects the formation and ecological functions of microbial biofilms. Sci. Total Environ. 2022, 832, 155026. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Shi, M.; Chen, L.; Zhou, J.; Zhang, L.; Li, Y.; Xue, Q.; Lai, H. The biocontrol agent Streptomyces pactum increases Pseudomonas koreensis populations in the rhizosphere by enhancing chemotaxis and biofilm formation. Soil Biol. Biochem. 2020, 144, 107755. [Google Scholar] [CrossRef]
- Yang, W.; Yan, H.; Dong, G.; Li, Z.; Jiang, C.; Gu, D.; Niu, D.; Zhou, D.; Luo, Y. Comparative transcriptomics reveal different genetic adaptations of biofilm formation in Bacillus subtilis isolate 1JN2 in response to Cd2+ treatment. Front. Microbiol. 2022, 13, 1002482. [Google Scholar] [CrossRef]
- Fu, H.; Chen, F.; Liu, W.; Kong, W.; Wang, C.; Fang, X.; Ye, J. Adding nutrients to the biocontrol strain JK-SH007 promotes biofilm formation and improves resistance to stress. AMB Express 2020, 10, 32. [Google Scholar] [CrossRef] [Green Version]
- Fessia, A.; Sartori, M.; Garcia, D.; Fernandez, L.; Ponzio, R.; Barros, G.; Nesci, A. In vitro studies of biofilm-forming Bacillus strains, biocontrol agents isolated from the maize phyllosphere. Biofilm 2022, 4, 100097. [Google Scholar] [CrossRef]
- Deng, Q.; Lei, X.; Zhang, H.; Deng, L.; Yi, L.; Zeng, K. Phenylalanine Promotes Biofilm Formation of Meyerozyma caribbica to Improve Biocontrol Efficacy against Jujube Black Spot Rot. J. Fungi 2022, 8, 1313. [Google Scholar] [CrossRef]
- Zhang, Y.; Gu, Y.; Zheng, Y.; Wang, Y.; Nie, L.; Qiao, R.; He, Y. Deletion of luxS gene mediated by λRed gene recombination technology reduces biofilm formation and stress resistance of Lactobacillus fermentum. Food Biosci. 2022, 49, 101892. [Google Scholar] [CrossRef]
- Liu, J.; Liu, K.; Zhao, Z.; Wang, Z.; Wang, F.; Xin, Y.; Qu, J.; Song, F.; Li, Z. The LuxS/AI-2 Quorum-Sensing System Regulates the Algicidal Activity of Shewanella xiamenensis Lzh-2. Front. Microbiol. 2021, 12, 814929. [Google Scholar] [CrossRef]
- Meng, F.; Zhang, F.; Chen, Q.; Yang, M.; Yang, Y.; Li, X.; Gu, W.; Yu, J. Virtual screening and in vitro experimental verification of LuxS inhibitors from natural products for Lactobacillus reuteri. Biomed. Pharmacother. 2022, 147, 112521. [Google Scholar] [CrossRef]
- Meng, F.; Zhao, M.; Lu, Z. The LuxS/AI-2 system regulates the probiotic activities of lactic acid bacteria. Trends Food Sci. Technol. 2022, 127, 272–279. [Google Scholar] [CrossRef]
- Colledge, V.L.; Fogg, M.J.; Levdikov, V.M.; Leech, A.; Dodson, E.J.; Wilkinson, A.J. Structure and organisation of SinR, the master regulator of biofilm formation in Bacillus subtilis. J. Mol. Biol. 2011, 411, 597–613. [Google Scholar] [CrossRef] [Green Version]
- Minamino, T.; Kinoshita, M.; Inoue, Y.; Kitao, A.; Namba, K. Conserved GYXLI Motif of FlhA Is Involved in Dynamic Domain Motions of FlhA Required for Flagellar Protein Export. Microbiol. Spectr. 2022, 10, e0111022. [Google Scholar] [CrossRef] [PubMed]
- Verma, N.; Srivastava, S.; Malik, R.; Goyal, P.; Pandey, J. Inhibition and disintegration of Bacillus subtilis biofilm with small molecule inhibitors identified through virtual screening for targeting TasA(28-261), the major protein component of ECM. J. Biomol. Struct. Dyn. 2022, 41, 2431–2447. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Zhang, J.; Wu, X.; Gu, X.; Wang, S.; Zhang, H. A novel biocontrol strain Pantoea jilinensis D25 for effective biocontrol of tomato gray mold (causative agent Botrytis cinerea). Biol. Control 2021, 164, 104766. [Google Scholar] [CrossRef]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. 2011, 47, 2437. [Google Scholar] [CrossRef]
- Camara-Almiron, J.; Navarro, Y.; Diaz-Martinez, L.; Magno-Perez-Bryan, M.C.; Molina-Santiago, C.; Pearson, J.R.; de Vicente, A.; Perez-Garcia, A.; Romero, D. Dual functionality of the amyloid protein TasA in Bacillus physiology and fitness on the phylloplane. Nat. Commun. 2020, 11, 1859. [Google Scholar] [CrossRef] [Green Version]
- El-Naggar, N.E.; Saber WI, A.; Zweil, A.M.; Bashir, S.I. An innovative green synthesis approach of chitosan nanoparticles and their inhibitory activity against phytopathogenic Botrytis cinerea on strawberry leaves. Sci. Rep. 2022, 12, 3515. [Google Scholar] [CrossRef]
- Gao, N.; Zhang, J.; Pan, Z.; Zhao, X.; Ma, X.; Zhang, H. Biodegradation of Atrazine by Mixed Bacteria of Klebsiella variicola Strain FH-1 and Arthrobacter sp. NJ-1. Bull. Environ. Contam. Toxicol. 2020, 105, 481–489. [Google Scholar] [CrossRef]
- Xi, X.; Fan, J.; Yang, X.; Liang, Y.; Zhao, X.; Wu, Y. Evaluation of the anti-oomycete bioactivity of rhizosphere soil-borne isolates and the biocontrol of soybean root rot caused by Phytophthora sojae. Biol. Control 2022, 166, 104818. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, C.; Zhang, S.; Yu, H.; Pan, H.; Zhang, H. Klebsiella jilinsis 2N3 promotes maize growth and induces resistance to northern corn leaf blight. Biol. Control 2021, 156, 104554. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, T.; Xia, J.; Yu, D.; Wang, J.; Zheng, X. Biocontrol of postharvest gray mold of cherry tomatoes with the marine yeast Rhodosporidium paludigenum. Biol. Control 2010, 53, 178–182. [Google Scholar] [CrossRef]
- Raynaldo, F.A.; Dhanasekaran, S.; Ngea GL, N.; Yang, Q.; Zhang, X.; Zhang, H. Investigating the biocontrol potentiality of Wickerhamomyces anomalus against postharvest gray mold decay in cherry tomatoes. Sci. Hortic. 2021, 285, 110137. [Google Scholar] [CrossRef]
- Chen, N.; Li, X.; Shi, H.; Hu, Q.; Zhang, Y.; Leng, X. Effect of biodegradable film mulching on crop yield, soil microbial and enzymatic activities, and optimal levels of irrigation and nitrogen fertilizer for the Zea mays crops in arid region. Sci. Total Environ. 2021, 776, 145970. [Google Scholar] [CrossRef]
- Ghiloufi, W.; Seo, J.; Kim, J.; Chaieb, M.; Kang, H. Effects of Biological Soil Crusts on Enzyme Activities and Microbial Community in Soils of an Arid Ecosystem. Microb. Ecol. 2019, 77, 201–216. [Google Scholar] [CrossRef] [PubMed]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Zhang, P.; Xin, H.; van der Lee, T. Tree pathogens Armillaria solidipes influence the biocontrol activity of Bacillus velezensis BY6. Biol. Control 2023, 179, 105176. [Google Scholar] [CrossRef]
- Gao, T.; Wang, X.; Qin, Y.; Ren, Z.; Zhao, X. Watermelon Root Exudates Enhance Root Colonization of Bacillus amyloliquefaciens TR2. Curr. Microbiol. 2023, 80, 110. [Google Scholar] [CrossRef]
- Mauritzen, J.J.; Sondberg, E.; Kalatzis, P.G.; Roager, L.; Gram, L.; Svenningsen, S.L.; Middelboe, M. Strain-specific quorum-sensing responses determine virulence properties in Vibrio anguillarum. Environ. Microbiol. 2023. [Google Scholar] [CrossRef]
- Sadanandan, B.; Vaniyamparambath, V.; Lokesh, K.N.; Shetty, K.; Joglekar, A.P.; Ashrit, P.; Hemanth, B. Candida albicans biofilm formation and growth optimization for functional studies using response surface methodology. J. Appl. Microbiol. 2022, 132, 3277–3292. [Google Scholar] [CrossRef]
- Wang, X.; Xie, S.; Mu, X.; Guan, B.; Hu, Y.; Ni, Y. Investigating the resistance responses to Alternaria brassicicola in ‘Korla’ fragrant pear fruit induced by a biocontrol strain Bacillus subtilis Y2. Postharvest Biol. Technol. 2023, 199, 112293. [Google Scholar] [CrossRef]
- Baptista, J.P.; Teixeira, G.M.; de Jesus ML, A.; Berte, R.; Higashi, A.; Mosela, M.; da Silva, D.V.; de Oliveira, J.P.; Sanches, D.S.; Brancher, J.D.; et al. Antifungal activity and genomic characterization of the biocontrol agent Bacillus velezensis CMRP 4489. Sci. Rep. 2022, 12, 17401. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Zhang, C.; Zhang, M.; Wu, Y.; Zhang, Z.; Zhang, H. Degradation characteristics and soil remediation of thifensulfuron-methyl by immobilized Serratia marcecens N80 beads. Environ. Technol. Innov. 2021, 24, 102059. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Design Variable (Factors) | Unit | Code | Real Values of the Coded Levels | ||
|---|---|---|---|---|---|
| −1 | 0 | +1 | |||
| CaCl2 | g/L | A | 0 | 5 | 10 |
| Temperature | ℃ | B | 25 | 30 | 35 |
| Time | h | C | 36 | 48 | 60 |
| Runs | A | B | C | OD570 |
|---|---|---|---|---|
| 1 | 0 | −1 | −1 | 2.546 |
| 2 | 0 | 0 | 0 | 3.224 |
| 3 | 0 | 1 | −1 | 2.535 |
| 4 | −1 | 0 | −1 | 2.632 |
| 5 | 0 | −1 | 1 | 2.604 |
| 6 | 0 | 0 | 0 | 3.226 |
| 7 | −1 | −1 | 0 | 2.609 |
| 8 | −1 | 1 | 0 | 2.704 |
| 9 | 0 | 0 | 0 | 3.228 |
| 10 | 1 | 1 | 0 | 2.698 |
| 11 | 0 | 1 | 1 | 2.712 |
| 12 | 1 | 0 | 1 | 2.712 |
| 13 | 1 | −1 | 0 | 2.688 |
| 14 | 0 | 0 | 0 | 3.221 |
| 15 | 0 | 0 | 0 | 3.03 |
| 16 | −1 | 0 | 1 | 2.682 |
| 17 | 1 | 0 | −1 | 2.632 |
| Source | Sum of Squares | df | Mean Square | F-Value | p-Value | |
|---|---|---|---|---|---|---|
| Model | 1.07 | 9 | 0.1189 | 26.01 | 0.0001 | Significant |
| A-CaCl2 | 0.0013 | 1 | 0.0013 | 0.2902 | 0.6068 | |
| B-Temperature | 0.0051 | 1 | 0.0051 | 1.12 | 0.03258 | |
| C-Time | 0.0167 | 1 | 0.0167 | 3.64 | 0.0979 | |
| AB | 0.0018 | 1 | 0.0018 | 0.3953 | 0.5495 | |
| AC | 0.0002 | 1 | 0.0002 | 0.0492 | 0.8307 | |
| BC | 0.0035 | 1 | 0.0035 | 0.7748 | 0.4079 | |
| A2 | 0.2092 | 1 | 0.2092 | 45.78 | 0.0003 | |
| B2 | 0.3496 | 1 | 0.3496 | 76.51 | <0.0001 | |
| C2 | 0.3749 | 1 | 0.3749 | 82.05 | <0.0001 | |
| Residual | 0.0320 | 7 | 0.0046 | |||
| Lack of Fit | 0.0016 | 3 | 0.0005 | 0.0710 | 0.9724 | Not significant |
| Pure Error | 0.0304 | 4 | 0.0076 | |||
| Cor Total | 1.10 | 16 |
| Treatment | Inoculants of Treatment | Disease Severity Index (%) | Disease Reduction (%) |
|---|---|---|---|
| BS | B. cinerea + Sterile water | 85.6 | - |
| BSB | B. cinerea + strain D50 (before optimization) | 58.4 | 39.7 |
| BSA | B. cinerea + strain D50 (after optimization) | 45.3 | 46.2 |
| Treatments | Fresh Weight (g) | Root Length (cm) | Shoot Length (cm) | Chl a (mg/mL) | Chl b (mg/mL) | Total Chlorophyll (mg/mL) | Total Phenolic Content (mg/100 g) | Total Soluble Protein (mg/100 g) |
|---|---|---|---|---|---|---|---|---|
| BS | 1.45 ± 0.05 c | 6.15 ± 0.13 c | 17.82 ± 0.32 c | 11.99 ± 0.08 c | 3.75 ± 0.08 c | 15.74 ± 0.15 c | 163.38 ± 0.28 c | 301.76 ± 1.42 c |
| BSB | 2.06 ± 0.07 b | 7.06 ± 0.12 b | 20.23 ± 0.12 b | 12.48 ± 0.13 b | 4.18 ± 0.09 b | 17.27 ± 0.08 b | 225.82 ± 0.68 b | 345.2 ± 2.68 b |
| BSA | 2.73 ± 0.03 a | 8.26 ± 0.24 a | 24.36 ± 0.18 a | 14.89 ± 0.06 a | 4.59 ± 0.05 a | 19.48 ± 0.12 a | 249.17 ± 0.36 a | 390.8 ± 3.87 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, L.; Gu, X.; Sun, L.; Dong, M.; Gao, A.; Han, Z.; Pan, H.; Zhang, H. Adding Metal Ions to the Bacillus mojavensis D50 Promotes Biofilm Formation and Improves Ability of Biocontrol. J. Fungi 2023, 9, 526. https://doi.org/10.3390/jof9050526
Zheng L, Gu X, Sun L, Dong M, Gao A, Han Z, Pan H, Zhang H. Adding Metal Ions to the Bacillus mojavensis D50 Promotes Biofilm Formation and Improves Ability of Biocontrol. Journal of Fungi. 2023; 9(5):526. https://doi.org/10.3390/jof9050526
Chicago/Turabian StyleZheng, Lining, Xuehu Gu, Liangpeng Sun, Meiqi Dong, Ao Gao, Zhe Han, Hongyu Pan, and Hao Zhang. 2023. "Adding Metal Ions to the Bacillus mojavensis D50 Promotes Biofilm Formation and Improves Ability of Biocontrol" Journal of Fungi 9, no. 5: 526. https://doi.org/10.3390/jof9050526