
Deciphering Plant-Induced Responses toward Botrytis cinerea and Plasmopara viticola Attacks in Two Grapevine Cultivars Colonized by the Root Biocontrol Oomycete, Pythium oligandrum
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Microorganisms
2.3. Experimental Design and Plant Inoculations
2.4. RNA Extraction and RT-qPCR
2.5. Quantitative Polymerase Chain Reaction
2.6. Statistical Analyses
3. Results
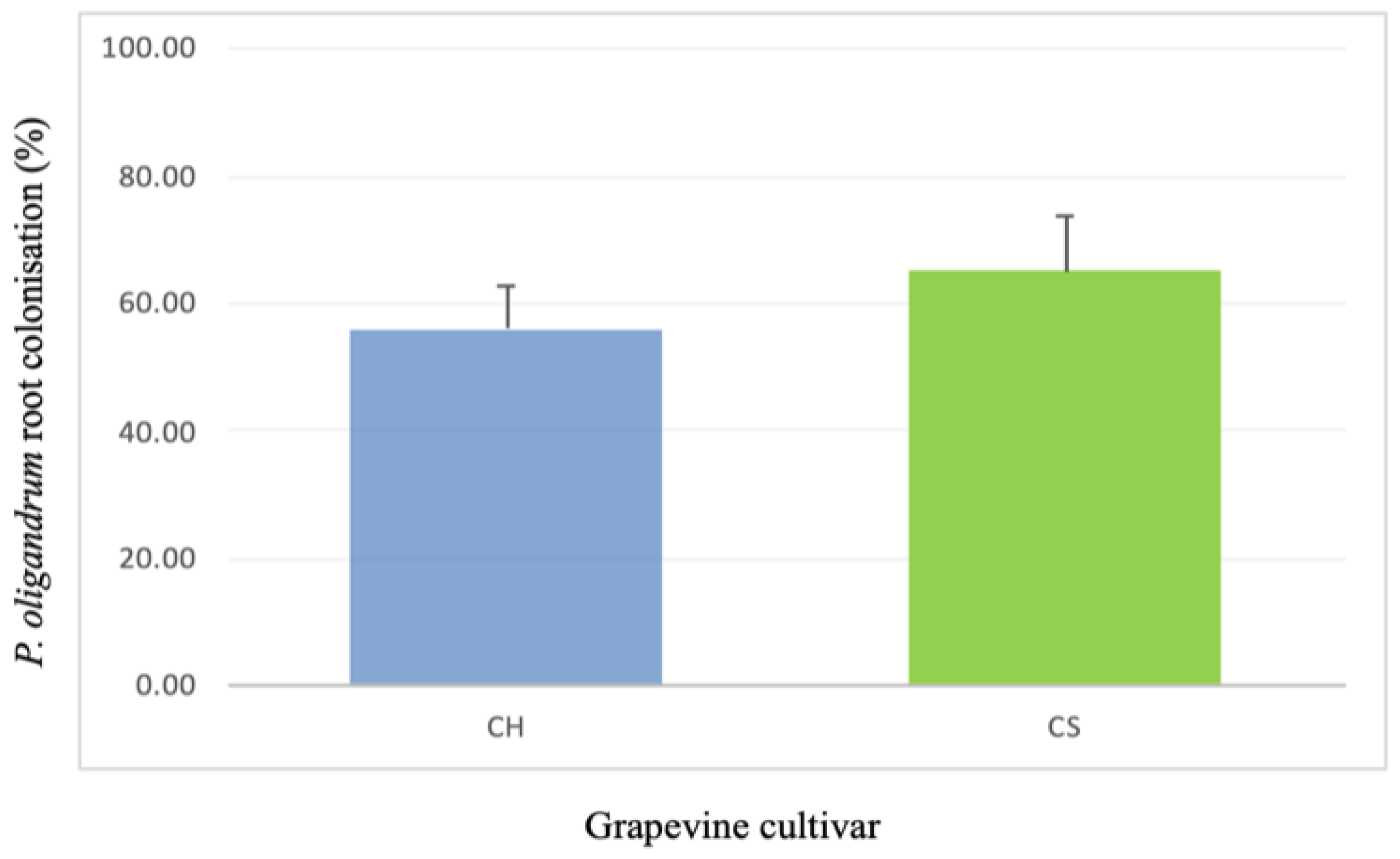
3.1. Assessment of Grapevine Root Colonization by P. oligandrum
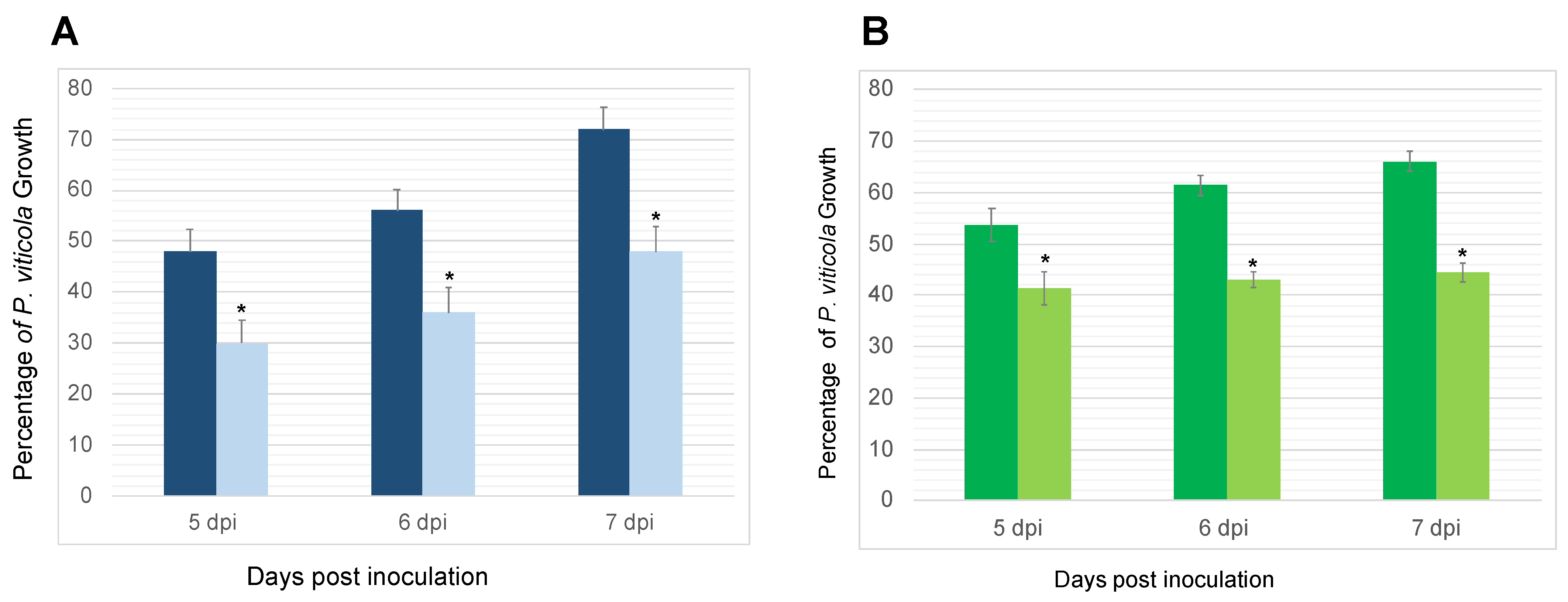
3.2. Protection of Grapevine Leaves against B. cinerea and P. viticola
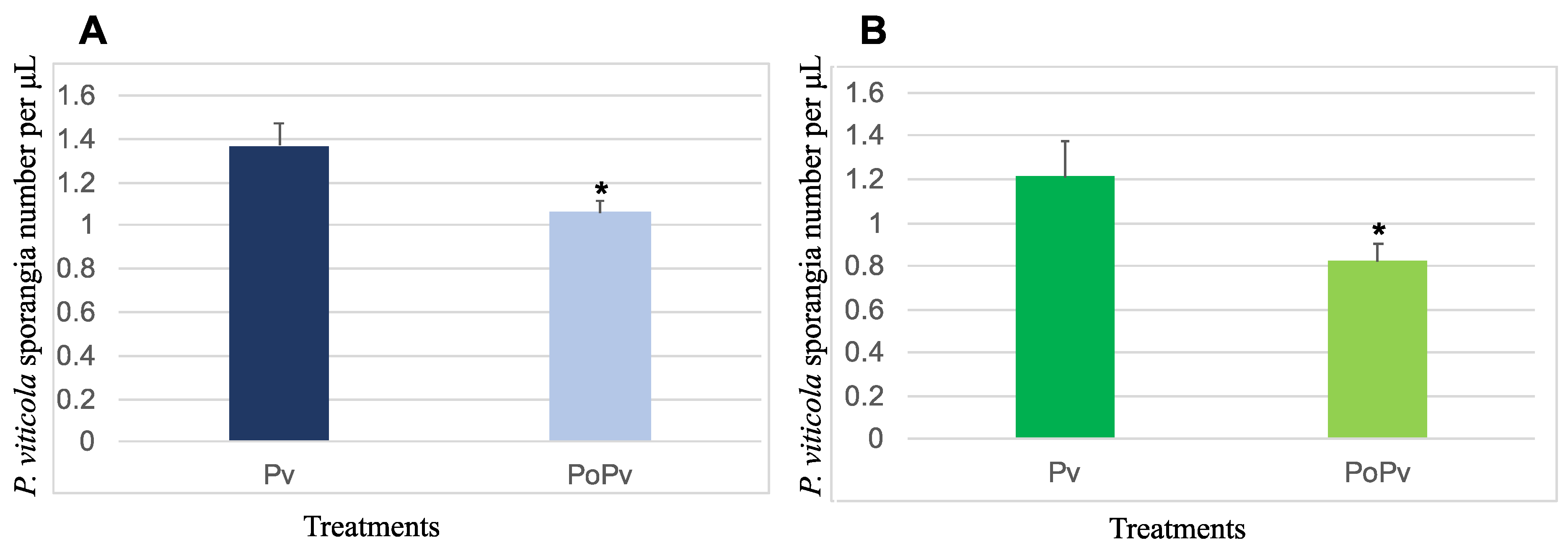
3.2.1. Reduction of P. viticola’s Sporulation on P. oligandrum-Inoculated Plants
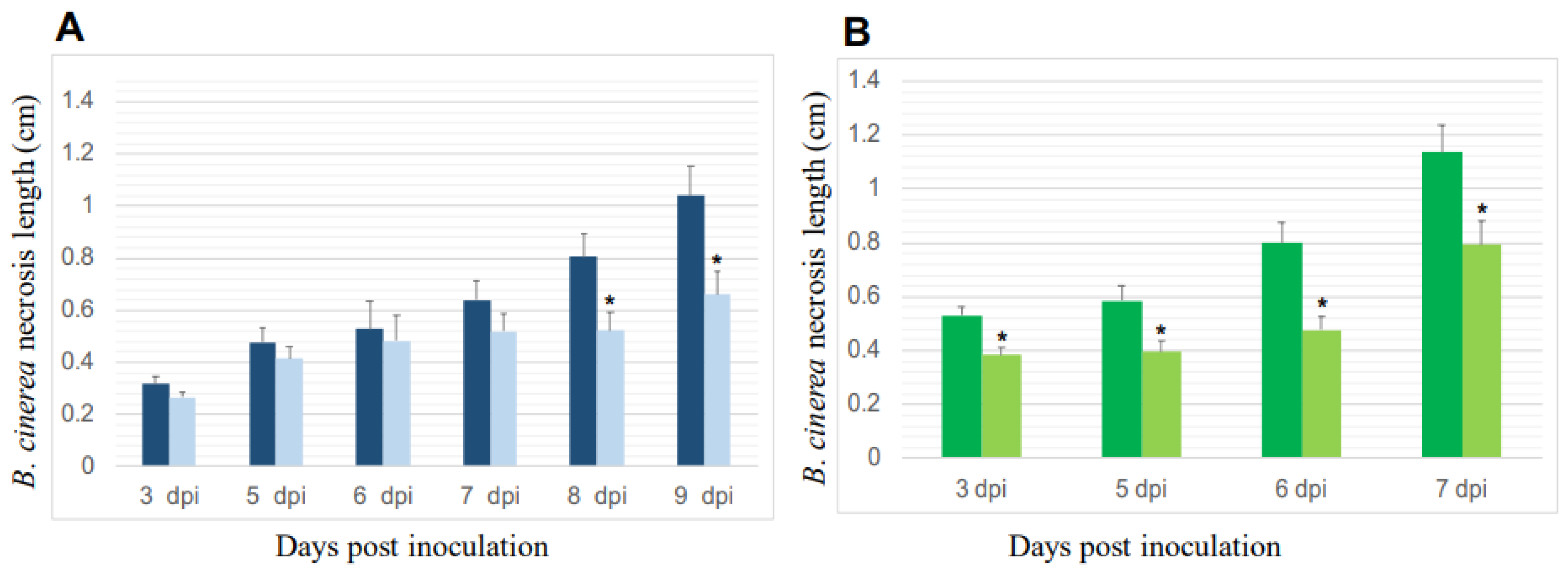
3.2.2. Reduction of B. cinerea’s Necroses on P. oligandrum-Inoculated Plants
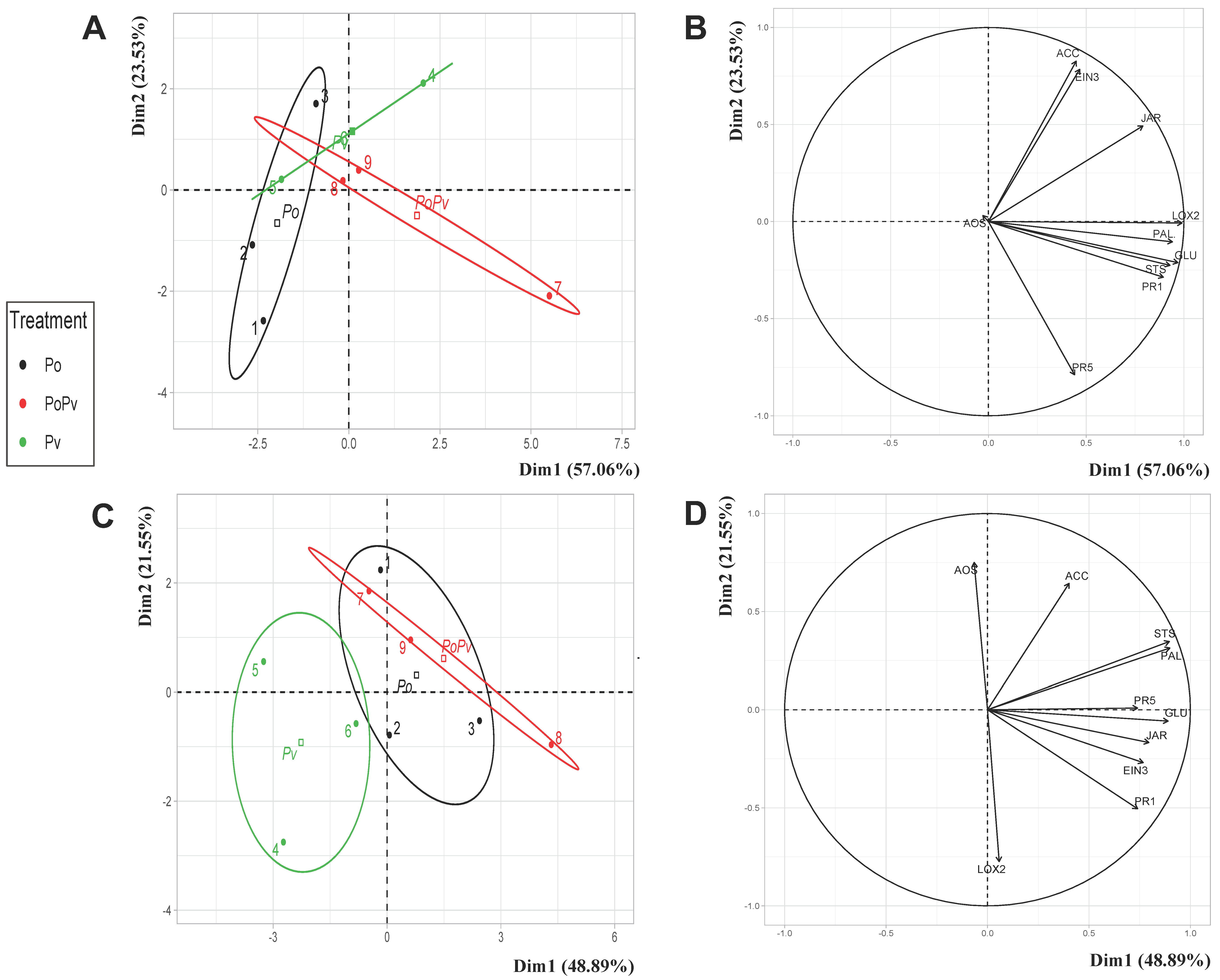
3.3. Evaluation, in the Two Cultivars, of Specific Foliar Grapevine Gene Defense Induction by P. oligandrum in Response to P. viticola and B. cinerea Infection
3.3.1. P. oligandrum Induction of Grapevine Gene Defenses against P. viticola Attack
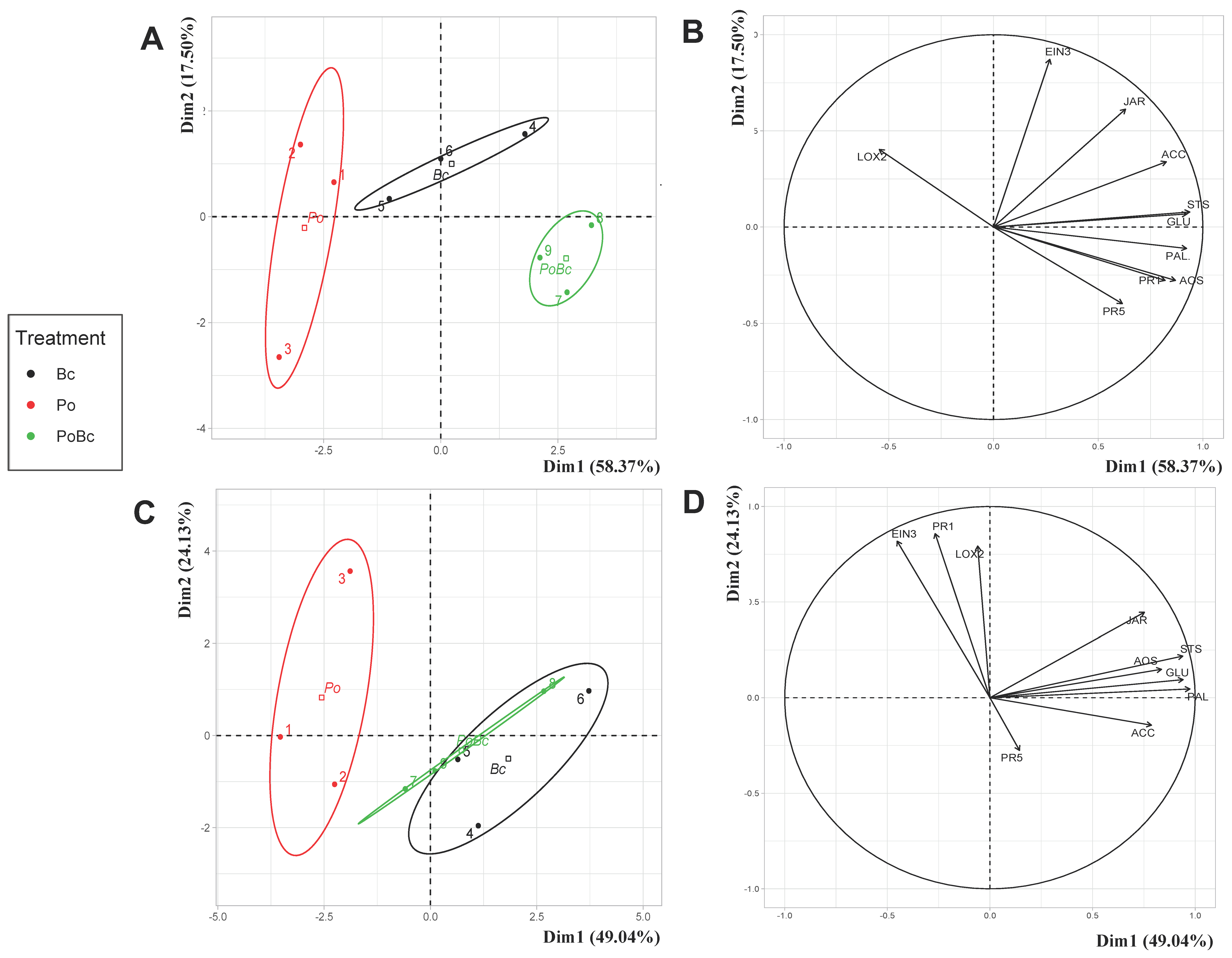
3.3.2. P. oligandrum Induction of Grapevine Defenses against B. cinerea
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gouadec, D.; Blouin, J. Les Parasites de la Vigne: Stratégies de Protection Raisonnée Stratégies de Protection Raisonnée; Dunod: Paris, France, 2007; p. 421. [Google Scholar]
- Massi, F.; Torriani, S.F.F.; Borghi, L.; Toffolatti, S.L. Fungicide resistance evolution and detection in plant pathogens: Plasmopara Viticola as a case study. Microorganisms 2021, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Doulia, D.S.; Anagnos, E.K.; Liapis, K.S.; Klimentzos, D.A. Removal of pesticides from white and red wines by microfiltration. J. Hazard. Mater. 2016, 317, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J.; Barquero, M.; González-Andrés, F. Insight into the Microbiological Control Strategies against Botrytis Cinerea Using Systemic Plant Resistance Activation. Agronomy 2020, 10, 1822. [Google Scholar] [CrossRef]
- Koledenkova, K.; Esmaeel, Q.; Jacquard, C.; Nowak, J.; Clément, C.; Barka, E.A. Plasmopara Viticola the Causal Agent of Downy Mildew of Grapevine: From Its Taxonomy to Disease Management. Front. Microbiol. 2022, 13, 889472. [Google Scholar] [CrossRef]
- Veloso, J.F.C.; Díaz, J. Fusarium Oxysporum Fo47 Confers Protection to Pepper Plants against Verticillium Dahliae and Phytophthora Capsici, and Induces the Expression of Defence Genes. Plant Pathol. 2012, 61, 281–288. [Google Scholar] [CrossRef]
- Ghule, M.R.; Sawant, I.S. Potential of Fusarium spp. for Biocontrol of Downy Mildew of Grapes. Pest. Manag. Hortic. Ecsyst. 2017, 23, 147–152. [Google Scholar]
- Perazzolli, M.; Roatti, B.; Bozza, E.; Pertot, I. Trichoderma Harzianum T39 Induces Resistance against Downy Mildew by Priming for Defense without Costs for Grapevine. Biol. Control. 2011, 58, 74–82. [Google Scholar] [CrossRef]
- Aziz, A.; Trotel-Aziz, P.; Dhuicq, L.; Jeandet, P.; Couderchet, M.; Vernet, G. Chitosan Oligomers and Copper Sulfate Induce Grapevine Defense Reactions and Resistance to Gray Mold and Downy Mildew. Phytopathology 2006, 96, 1188–1194. [Google Scholar] [CrossRef] [PubMed]
- Trouvelot, S.; Varnier, A.-L.; Allègre, M.; Mercier, L.; Baillieul, F.; Arnould, C.; Gianinazzi-Pearson, V.; Klarzynski, O.; Joubert, J.-M.; Pugin, A.; et al. A β-1,3 Glucan Sulfate Induces Resistance in Grapevine against Plasmopara Viticola through Priming of Defense Responses, Including HR-like Cell Death. Mol. Plant Microbe Interact. 2008, 21, 232–243. [Google Scholar] [CrossRef]
- Dubreuil-Maurizi, C.; Trouvelot, S.; Frettinger, P.; Pugin, A.; Wendehenne, D.; Poinssot, B. β-Aminobutyric Acid Primes an NADPH Oxidase-Dependent Reactive Oxygen Species Production during Grapevine-Triggered Immunity. Mol. Plant Microbe Interact. 2010, 23, 1012–1021. [Google Scholar] [CrossRef]
- Bělonožníková, K.; Hýsková, V.; Chmelík, J.; Kavan, D.; Čeřovská, N.; Ryšlavá, H. Pythium Oligandrum in Plant Protection and Growth Promotion: Secretion of Hydrolytic Enzymes, Elicitors and Tryptamine as Auxin Precursor. Microbiol. Res. 2022, 258, 126976. [Google Scholar] [CrossRef]
- Gerbore, J.; Vallance, J.; Yacoub, A.; Delmotte, F.; Grizard, D.; Regnault-Roger, C.; Rey, P. Characterization of Pythium Oligandrum Populations That Colonize the Rhizosphere of Vines from the Bordeaux Region. FEMS Microbiol. Ecol. 2014, 90, 153–167. [Google Scholar] [CrossRef]
- Rey, P.; Floch, G.; Benhamou, N.; Tirilly, Y. Pythium Oligandrum Biocontrol: Its Relationships with Fungi and Plants. Plant Microbe. 2008, 43–67. [Google Scholar]
- Benhamou, N.; le Floch, G.; Vallance, J.; Gerbore, J.; Grizard, D.; Rey, P. Pythium Oligandrum: An Example of Opportunistic Success. Microbiology 2012, 158, 2679–2694. [Google Scholar] [CrossRef] [PubMed]
- Gerbore, J.; Benhamou, N.; Vallance, J.; Le Floch, G.; Grizard, D.; Regnault-Roger, C.; Rey, P. Biological Control of Plant Pathogens: Advantages and Limitations Seen through the Case Study of Pythium Oligandrum. Environ. Sci. Pollut. Res. 2014, 21, 4847–4860. [Google Scholar] [CrossRef]
- Yacoub, A.; Haidar, R.; Gerbore, J.; Masson, C.; Dufour, M.-C.; Guyoneaud, R.; Rey, P. Pythium Oligandrum Induces Grapevine Defence Mechanisms against the Trunk Pathogen Neofusicoccum Parvum. Phytopathol. Mediterr. 2020, 59, 565–580. [Google Scholar] [CrossRef]
- Yacoub, A.; Gerbore, J.; Magnin, N.; Chambon, P.; Dufour, M.-C.; Corio-Costet, M.-F.; Guyoneaud, R.; Rey, P. Ability of Pythium Oligandrum Strains to Protect Vitis Vinifera L., by Inducing Plant Resistance against Phaeomoniella Chlamydospora, a Pathogen Involved in Esca, a Grapevine Trunk Disease. Biol. Control 2016, 92, 7–16. [Google Scholar] [CrossRef]
- Daraignes, L.; Gerbore, J.; Yacoub, A.; Dubois, L.; Romand, C.; Zekri, O.; Roudet, J.; Chambon, P.; Fermaud, M. Efficacy of P. Oligandrum Affected by Its Association with Bacterial BCAs and Rootstock Effect in Controlling Grapevine Trunk Diseases. Biol. Control. 2018, 119, 59–67. [Google Scholar] [CrossRef]
- Yacoub, A.; Magnin, N.; Gerbore, J.; Haidar, R.; Bruez, E.; Compant, S.; Guyoneaud, R.; Rey, P. The Biocontrol Root-Oomycete, Pythium Oligandrum, Triggers Grapevine Resistance and Shifts in the Transcriptome of the Trunk Pathogenic Fungus, Phaeomoniella Chlamydospora. Int. J. Mol. Sci. 2020, 21, 6876. [Google Scholar] [CrossRef]
- Mohamed, N.; Lherminier, J.; Farmer, M.-J.; Fromentin, J.; Béno, N.; Houot, V.; Milat, M.-L.; Blein, J.-P. Defense Responses in Grapevine Leaves Against Botrytis Cinerea Induced by Application of a Pythium Oligandrum Strain or Its Elicitin, Oligandrin, to Roots. Phytopathology 2007, 97, 611–620. [Google Scholar] [CrossRef] [PubMed]
- QiuYan, B.; XiuYing, H.; ZhiQiang, M.; JianJiang, Z.; HaiMin, J.; WenQiao, W. Inhibitory effect of Pythium oligandrum interaction with dimethomorph and the application of chemical decrement on grape downy mildew. Acta Phytopathol. Sin 2018, 48, 675–681. [Google Scholar]
- Krawiec, P.; Krawiec, M. The Use of Phytium Oligandrum to Control Grey Mold in Raspberry. Acta Hortic. 2020, 1277, 413–416. [Google Scholar] [CrossRef]
- Taibi, O.; Bardelloni, V.; Bove, F.; Scaglia, F.; Caffi, T.; Rossi, V. Activity of Resistance Inducers against Plasmopara Viticola in Vineyard. BIO Web Conf. 2022, 50, 03003. [Google Scholar] [CrossRef]
- Haidar, R.; Deschamps, A.; Roudet, J.; Calvo-Garrido, C.; Bruez, E.; Rey, P.; Fermaud, M. Multi-Organ Screening of Efficient Bacterial Control Agents against Two Major Pathogens of Grapevine. Biol. Control. 2016, 92, 55–65. [Google Scholar] [CrossRef]
- Berger, H.; Yacoub, A.; Gerbore, J.; Grizard, D.; Rey, P.; Sessitsch, A.; Compant, S. Draft Genome Sequence of Biocontrol Agent Pythium Oligandrum Strain Po37, an Oomycota. Genome Announc. 2016, 4, e00215-16. [Google Scholar] [CrossRef] [PubMed]
- Le Floch, G.L.; Rey, P.; Déniel, F.; Benhamou, N.; Picard, K.; Tirilly, Y. Enhancement of Development and Induction of Resistance in Tomato Plants by the Antagonist, Pythium Oligandrum. Agron. 2003, 23, 455–460. [Google Scholar] [CrossRef]
- Dussert, Y.; Gouzy, J.; Richart-Cervera, S.; Mazet, I.D.; Delière, L.; Couture, C.; Legrand, L.; Piron, M.-C.; Mestre, P.; Delmotte, F. Draft Genome Sequence of Plasmopara Viticola, the Grapevine Downy Mildew Pathogen. Genome Announc. 2016, 4, e00987-16. [Google Scholar] [CrossRef] [PubMed]
- Ciliberti, N.; Fermaud, M.; Roudet, J.; Languasco, L.; Rossi, V. Environmental Effects on the Production of Botrytis Cinerea Conidia on Different Media, Grape Bunch Trash, and Mature Berries. Aust. J. Grape Wine Res. 2016, 22, 262–270. [Google Scholar] [CrossRef]
- Aziz, A.; Poinssot, B.; Daire, X.; Adrian, M.; Bézier, A.; Lambert, B.; Joubert, J.-M.; Pugin, A. Laminarin Elicits Defense Responses in Grapevine and Induces Protection against Botrytis Cinerea and Plasmopara Viticola. Mol. Plant Microbe. Interact. 2003, 16, 1118–1128. [Google Scholar] [CrossRef]
- Dufour, M.-C.; Magnin, N.; Dumas, B.; Vergnes, S.; Corio-Costet, M.-F. High-Throughput Gene-Expression Quantification of Grapevine Defense Responses in the Field Using Microfluidic Dynamic Arrays. BMC Genom. 2016, 17, 957. [Google Scholar] [CrossRef] [PubMed]
- Jeffers, S.N.; Martin, S.B. Comparison of Two Media Selective for Phytophthora and Pythium Species. Plant Dis. 1986, 70, 1038–1043. [Google Scholar] [CrossRef]
- Plaats-Niterink, A.J.V. der Monograph of the Genus Pythium. Stud. Mycol. 1981, 21, 242. [Google Scholar]
- Corio-Costet, M.-F.; Dufour, M.-C.; Cigna, J.; Abadie, P.; Chen, W.-J. Diversity and Fitness of Plasmopara Viticola Isolates Resistant to QoI Fungicides. Eur. J. Plant Pathol. 2011, 129, 315–329. [Google Scholar] [CrossRef]
- Dufour, M.C.; Lambert, C.; Bouscaut, J.; Mérillon, J.M.; Corio-Costet, M.F. Benzothiadiazole-Primed Defence Responses and Enhanced Differential Expression of Defence Genes in Vitis Vinifera Infected with Biotrophic Pathogens Erysiphe Necator and Plasmopara Viticola. Plant Pathol. 2013, 62, 370–382. [Google Scholar] [CrossRef]
- Delmas, C.E.L.; Mazet, I.D.; Jolivet, J.; Delière, L.; Delmotte, F. Simultaneous Quantification of Sporangia and Zoospores in a Biotrophic Oomycete with an Automatic Particle Analyzer: Disentangling Dispersal and Infection Potentials. J. Microbiol. Methods 2014, 107, 169–175. [Google Scholar] [CrossRef]
- Lakkis, S.; Trotel-Aziz, P.; Rabenoelina, F.; Schwarzenberg, A.; Nguema-Ona, E.; Clément, C.; Aziz, A. Strengthening Grapevine Resistance by Pseudomonas Fluorescens PTA-CT2 Relies on Distinct Defense Pathways in Susceptible and Partially Resistant Genotypes to Downy Mildew and Gray Mold Diseases. Front. Plant Sci. 2019, 10, 1112. [Google Scholar] [CrossRef]
- Reid, K.E.; Olsson, N.; Schlosser, J.; Peng, F.; Lund, S.T. An Optimized Grapevine RNA Isolation Procedure and Statistical Determination of Reference Genes for Real-Time RT-PCR during Berry Development. BMC Plant Biol. 2006, 6, 27. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef]
- Sachdev, S.; Singh, R.P. Isolation, Characterisation and Screening of Native Microbial Isolates for Biocontrol of Fungal Pathogens of Tomato. Clim. Chang. Environ. Sustain. 2018, 6, 46. [Google Scholar] [CrossRef]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “Secrets” of a Multitalented Biocontrol Agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef] [PubMed]
- Galet, P. Sensibilité Des Cépages à La Pourriture Grise et Incidences Économiques. OENO One 1970, 4, 235–242. [Google Scholar] [CrossRef]
- Sluyter, S.V.; Durako, M.J.; Halkides, C.J. Comparison of Grape Chitinase Activities in Chardonnay and Cabernet Sauvignon with Vitis Rotundifolia Cv. Fry. Am. J. Enol. Vitic. 2005, 56, 81–85. [Google Scholar] [CrossRef]
- Herrera-Téllez, V.I.; Cruz-Olmedo, A.K.; Plasencia, J.; Gavilanes-Ruíz, M.; Arce-Cervantes, O.; Hernández-León, S.; Saucedo-García, M. The Protective Effect of Trichoderma Asperellum on Tomato Plants against Fusarium Oxysporum and Botrytis Cinerea Diseases Involves Inhibition of Reactive Oxygen Species Production. Int. J. Mol. Sci. 2019, 20, 2007. [Google Scholar] [CrossRef] [PubMed]
- Bělonožníková, K.; Hýsková, V.; Vašková, M.; Křížek, T.; Čokrtová, K.; Vaněk, T.; Halířová, L.; Chudý, M.; Žufić, A.; Ryšlavá, H. Seed Protection of Solanum Lycopersicum with Pythium Oligandrum against Alternaria Brassicicola and Verticillium Albo-atrum. Microorganisms 2022, 10, 1348. [Google Scholar] [CrossRef] [PubMed]
- Polesani, M.; Bortesi, L.; Ferrarini, A.; Zamboni, A.; Fasoli, M.; Zadra, C.; Lovato, A.; Pezzotti, M.; Delledonne, M.; Polverari, A. General and Species-Specific Transcriptional Responses to Downy Mildew Infection in a Susceptible (Vitis Vinifera) and a Resistant (V. riparia) Grapevine Species. BMC Genom. 2010, 11, 117. [Google Scholar] [CrossRef]
- Malacarne, G.; Vrhovsek, U.; Zulini, L.; Cestaro, A.; Stefanini, M.; Mattivi, F.; Delledonne, M.; Velasco, R.; Moser, C. Resistance to Plasmopara Viticola in a Grapevine Segregating Population Is Associated with Stilbenoid Accumulation and with Specific Host Transcriptional Responses. BMC Plant Biol. 2011, 11, 114. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, A.; Monteiro, F.; Sebastiana, M. First Clues on a Jasmonic Acid Role in Grapevine Resistance against the Biotrophic Fungus Plasmopara Viticola. Eur. J. Plant Pathol. 2015, 142, 645–652. [Google Scholar] [CrossRef]
- Hase, S.; Takahashi, S.; Takenaka, S.; Nakaho, K.; Arie, T.; Seo, S.; Ohashi, Y. Involvement of Jasmonic Acid Signalling in Bacterial Wilt Disease Resistance Induced by Biocontrol Agent Pythium Oligandrum in Tomato. Plant Pathol. 2008, 57, 870–876. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | Cabernet Sauvignon | B | Chardonnay | |||||
|---|---|---|---|---|---|---|---|---|
| Po | Pv | PoPv | Po | Pv | PoPv | |||
| PR1 | 1.42 | 2.24 | 3.01 | PR1 | 0.83 | |||
| GLU | 1.52 | GLU | 0.83 | |||||
| PR5 | 2.81 | PR5 | 0.04 | 0.39 | ||||
| LOX2 | 0.62 | 1.6 | LOX2 | 5.08 | ||||
| AOS | 3.48 | AOS | 0.1 | |||||
| JAR | 1.63 | 1.71 | JAR | 1.25 |  | |||
| PAL | 1.45 | 2.08 | PAL | 0.48 | 0.26 | 0.57 | ||
| STS | 2.11 | STS | 0.36 | 0.09 | 0.53 | |||
| ACC | 1.16 | ACC | 0.86 | |||||
| EIN3 | EIN3 | 0.74 | ||||||
| C | Cabernet Sauvignon | D | Chardonnay | |||||
| Po | Bc | PoBc | Po | Bc | PoBc | |||
| PR1 | 0.88 | 1.4 | PR1 | 1.50 | 1.9 | |||
| GLU | 0.71 | 1.34 | 1.43 | GLU | 1.56 | 7.81 | 6.97 | |
| PR5 | 3.56 | 5.36 | 26.33 | PR5 | 1.43 | 5.03 | 7.15 | |
| LOX2 | 0.23 | 0.41 | LOX2 | 1.42 | 1.58 | |||
| AOS | 133.97 | 673.38 | AOS | 2.00 | 5890 | 8009.14 | ||
| JAR | JAR | 1.27 | 1.7 | 1.29 | ||||
| PAL | 2.96 | 6.86 | PAL | 0.72 | 6.62 | 4.98 | ||
| STS | 2.53 | 4.51 | STS | 0.97 | 6.84 | 5.34 | ||
| ACC | 0.49 | 1.69 | 1.51 | ACC | 1.45 | 2.99 | 2.37 | |
| EIN3 | EIN3 | 1.37 | 0.87 | 0.77 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yacoub, A.; Haidar, R.; Mesguida, O.; Gerbore, J.; Hachicha, M.; Attard, E.; Guyoneaud, R.; Rey, P. Deciphering Plant-Induced Responses toward Botrytis cinerea and Plasmopara viticola Attacks in Two Grapevine Cultivars Colonized by the Root Biocontrol Oomycete, Pythium oligandrum. J. Fungi 2023, 9, 511. https://doi.org/10.3390/jof9050511
Yacoub A, Haidar R, Mesguida O, Gerbore J, Hachicha M, Attard E, Guyoneaud R, Rey P. Deciphering Plant-Induced Responses toward Botrytis cinerea and Plasmopara viticola Attacks in Two Grapevine Cultivars Colonized by the Root Biocontrol Oomycete, Pythium oligandrum. Journal of Fungi. 2023; 9(5):511. https://doi.org/10.3390/jof9050511
Chicago/Turabian StyleYacoub, Amira, Rana Haidar, Ouiza Mesguida, Jonathan Gerbore, Maya Hachicha, Eléonore Attard, Rémy Guyoneaud, and Patrice Rey. 2023. "Deciphering Plant-Induced Responses toward Botrytis cinerea and Plasmopara viticola Attacks in Two Grapevine Cultivars Colonized by the Root Biocontrol Oomycete, Pythium oligandrum" Journal of Fungi 9, no. 5: 511. https://doi.org/10.3390/jof9050511