

Pruning Wound Protection Products Induce Alterations in the Wood Mycobiome Profile of Grapevines
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Vineyards
2.2. Experimental Setup and Sample Processing
2.2.1. Experimental Setup
2.2.2. Sample Processing
2.3. DNA Extraction, Amplification, Library Preparation and Sequencing
2.4. Bioinformatics
2.5. Data Analysis
3. Result
3.1. Sequencing Dataset Description
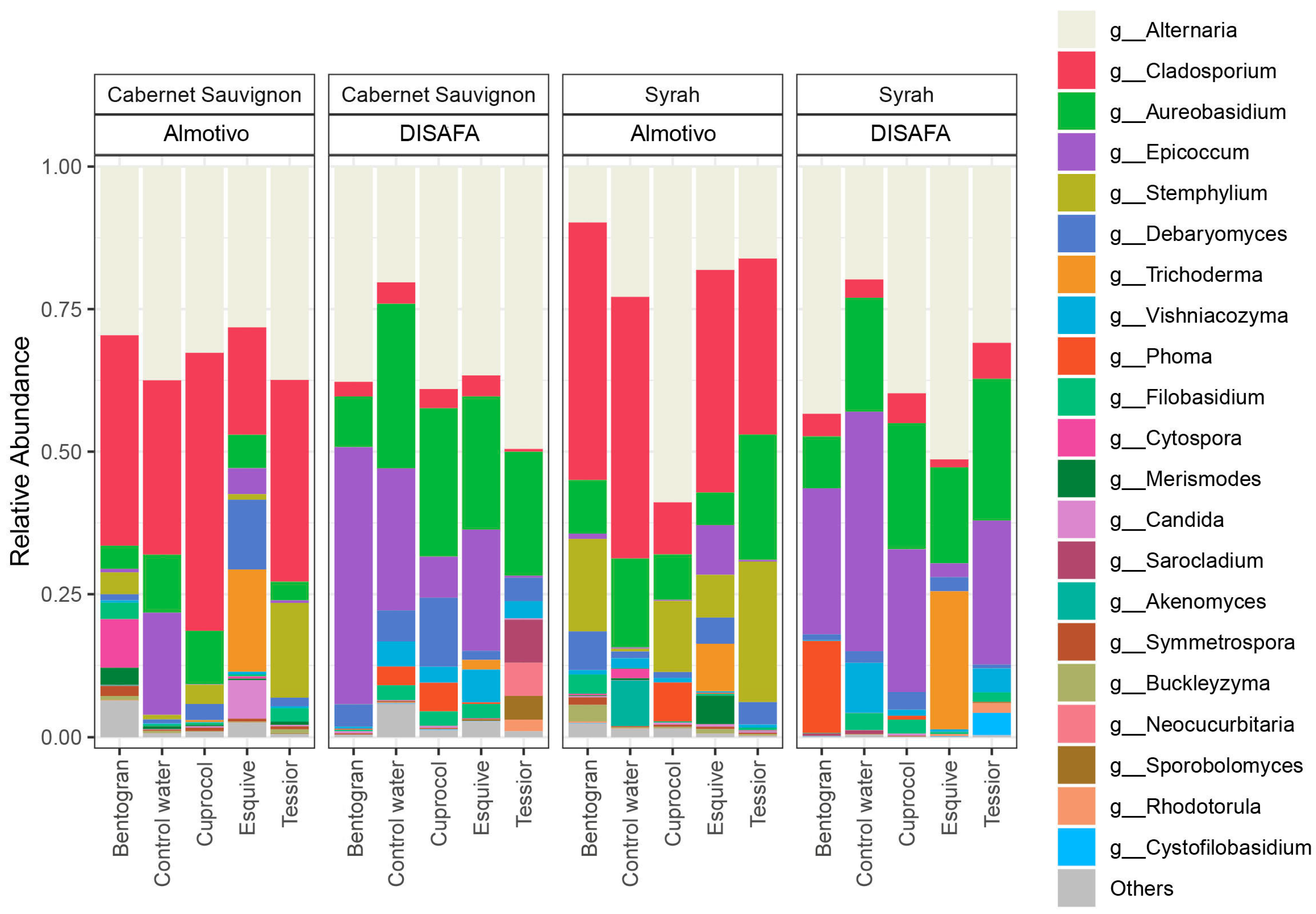
3.2. The Wood Mycobiome
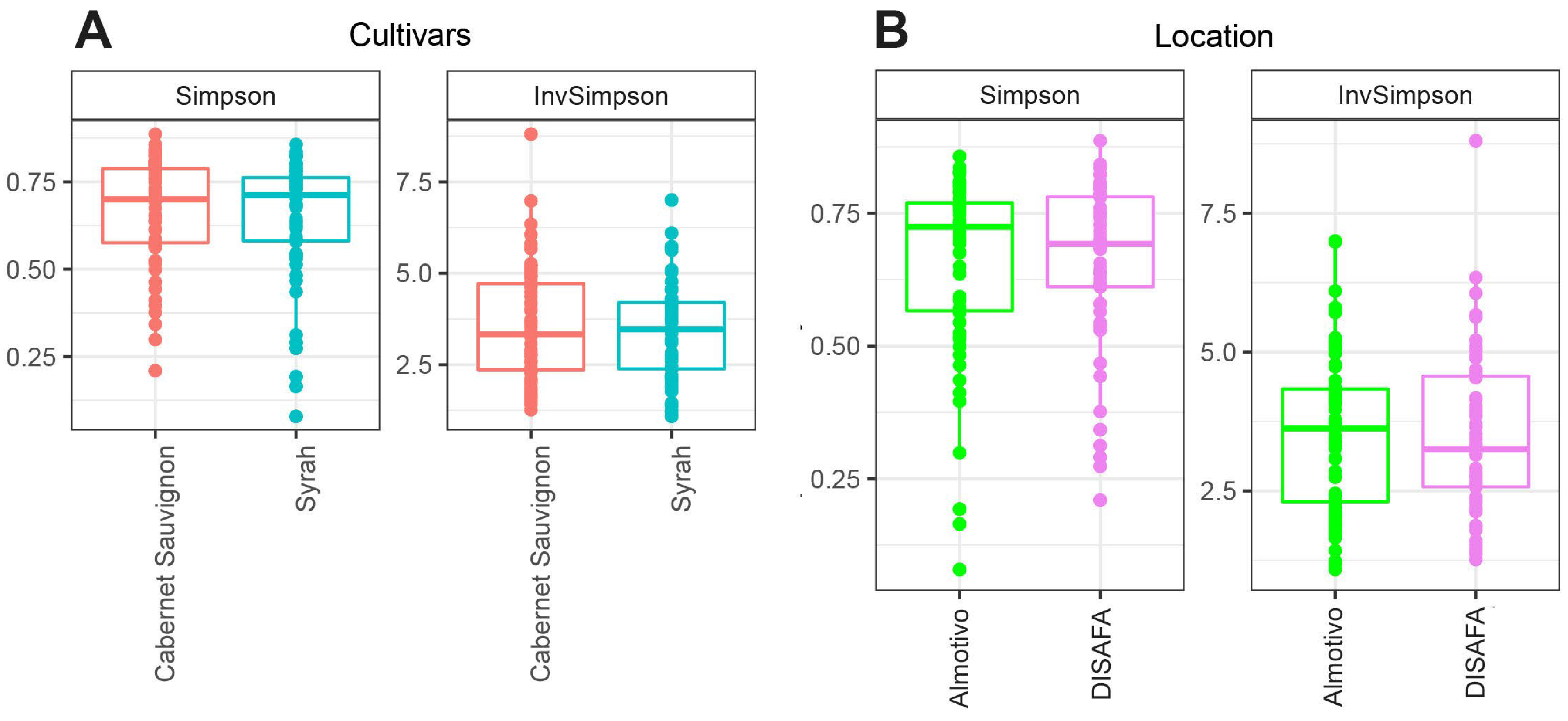
3.3. Alpha Diversity
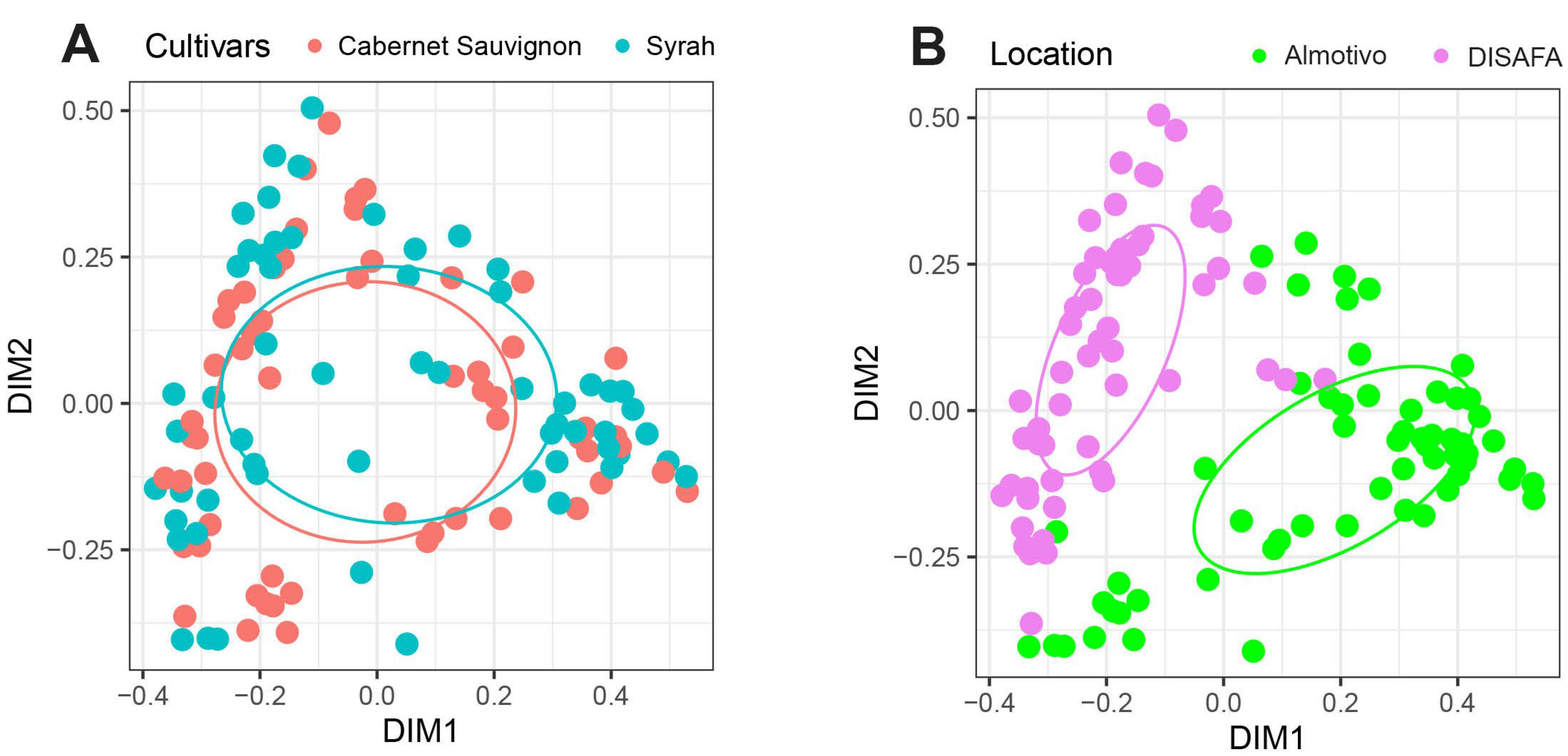
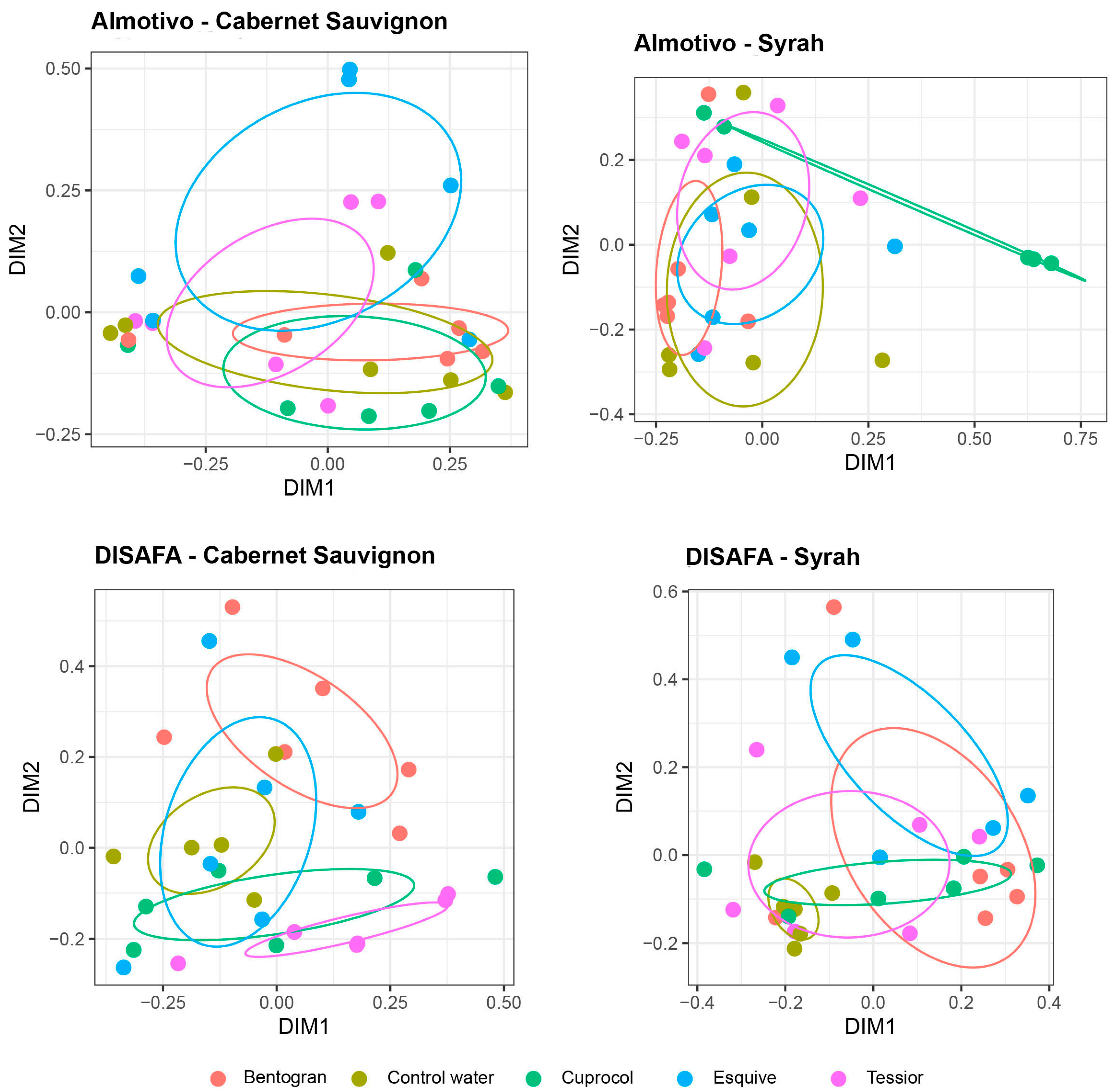
3.4. Beta Diversity
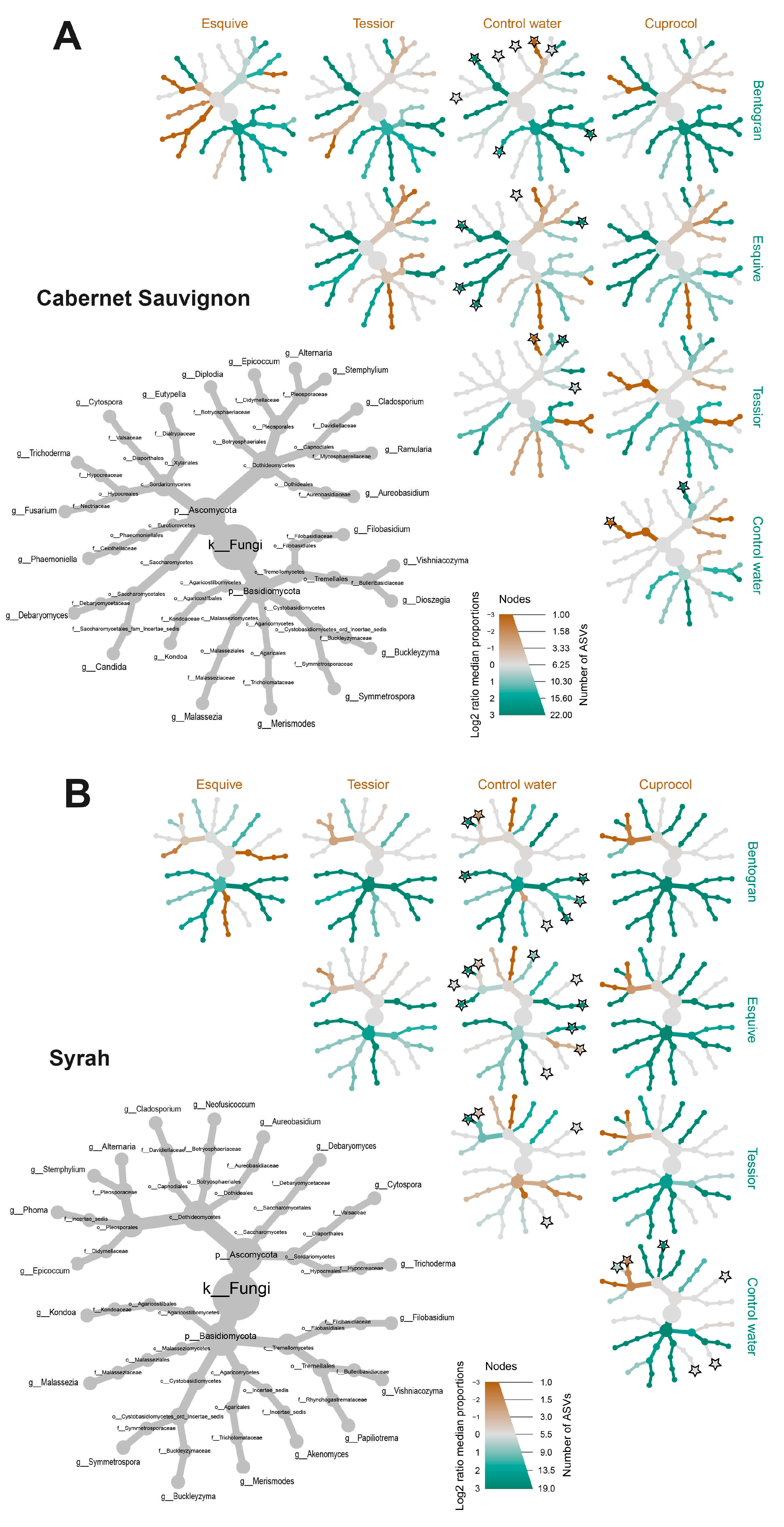
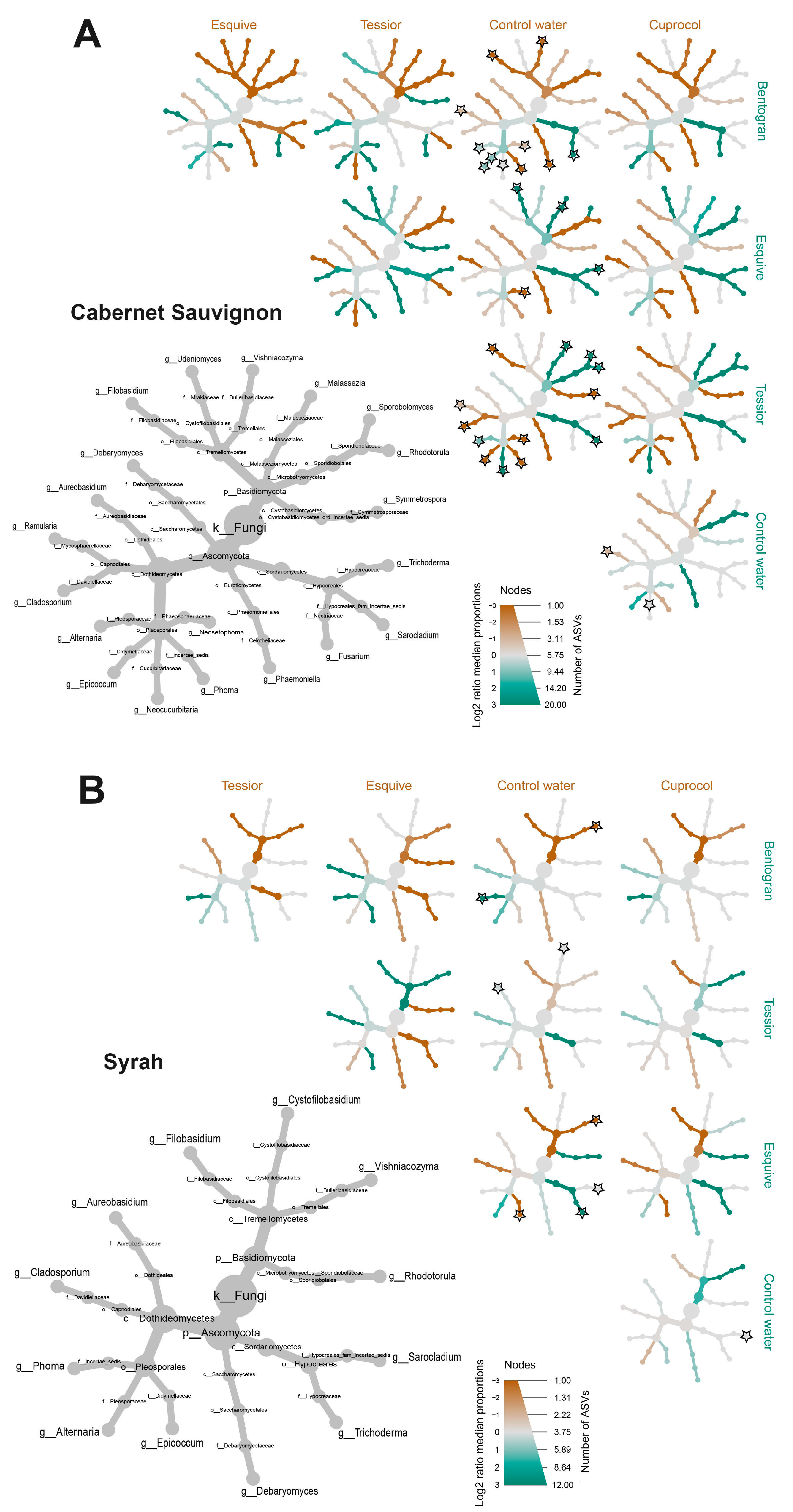
3.5. Taxa over/under-Representation
4. Discussion
4.1. Mycobiome Composition
4.2. Effects of Pruning Wound Protection Products on Mycobiome Profile
4.2.1. Cuprocol®
4.2.2. Tessior®
4.2.3. Esquive®
4.2.4. Bentogran®
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gramaje, D.; Urbez-Torres, J.R.; Sosnowski, M.R. Managing grapevine trunk diseases with respect to etiology and epidemiology: Current strategies and future prospects. Plant Dis. 2018, 102, 12–39. [Google Scholar] [CrossRef] [PubMed]
- Hrycan, J.; Hart, M.; Bowen, P.; Forge, T.; Urbez-Torres, J.R. Grapevine trunk disease fungi: Their roles as latent pathogens and stress factors that favour disease development and symptom expression. Phytopathol. Mediterr. 2020, 59, 395–424. [Google Scholar]
- Kenfaoui, J.; Radouane, N.; Mennani, M.; Tahiri, A.; El Ghadraoui, L.; Belabess, Z.; Fontaine, F.; El Hamss, H.; Amiri, S.; Lahlali, R.; et al. A panoramic view on grapevine trunk diseases threats: Case of Eutypa dieback, Botryosphaeria dieback, and esca disease. J. Fungi. 2022, 8, 595. [Google Scholar] [CrossRef]
- Fontaine, F.; Gramaje, D.; Armengol, J.; Smart, R.; Nagy, Z.A.; Borgo, M.; Rego, C.; Corio-Costet, M.-F. Grapevine Trunk Diseases. A Review; OIV Publications: Paris, France, 2016; p. 25. Available online: http://www.oiv.int/public/medias/4650/trunk-diseases-oiv-2016.pdf (accessed on 15 December 2022).
- Berbegal, M.; Ramón-albalat, A.; León, M.; Armengol, J. Evaluation of long-term protection from nursery to vineyard provided by Trichoderma atroviride SC1 against fungal grapevine trunk pathogens. Pest Manag. Sci. 2019, 76, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Battiston, E.; Compant, S.; Antonielli, L.; Mondello, V.; Clément, C.; Simoni, A.; Di Marco, S.; Mugnai, L.; Fontaine, F. In planta activity of novel copper (II)-based formulations to inhibit the esca-associated fungus Phaeoacremonium minimum in grapevine propagation material. Front. Plant Sci. 2021, 12, 649694. [Google Scholar] [CrossRef]
- Lade, S.B.; Štraus, D.; Buñol, A.; Oliva, J. Hot water treatment causes lasting alteration to the grapevine (Vitis vinifera L.) mycobiome and reduces pathogenic species causing grapevine trunk diseases. J. Fungi 2022, 8, 485. [Google Scholar] [CrossRef]
- Claverie, M.; Notaro, M.; Fontaine, F. Current knowledge on grapevine trunk diseases with complex etiology: A systemic approach. Phytopathol. Mediterr. 2020, 59, 29–53. [Google Scholar] [CrossRef]
- Reis, P.; Gaspar, A.; Alves, A.; Fontaine, F.; Rego, C. Combining an HA + Cu (II) site-targeted copper-based product with a pruning wound protection program to prevent infection with Lasiodiplodia spp. in grapevine. Plants 2021, 10, 2376. [Google Scholar] [CrossRef]
- Martínez-Diz, M.d.P.; Díaz-Losada, E.; Díaz-Fernandez, A.; Bouzas-Cid, Y.; Gramaje, D. Protection of grapevine pruning wounds against Phaeomoniella chlamydospora and Diplodia seriata by commercial biological and chemical methods. Crop Prot. 2021, 143, 105465. [Google Scholar] [CrossRef]
- Lecomte, P.; Diarra, B.; Carbonneau, A.; Rey, P.; Chevrier, C. Esca of grapevine and training practices in France: Results of a 10-year survey. Phytopathol. Mediterr. 2018, 57, 472–487. [Google Scholar]
- Cobos, R.; Mateos, R.M.; Álvarez-pérez, J.M.; Olego, M.A.; Sevillano, S.; González-García, S.; Garzón-Jimeno, E.; Coque, J.J.R. Effectiveness of natural antifungal compounds in controlling infection by grapevine trunk disease pathogens through pruning wounds. Appl. Environ. Microbiol. 2015, 81, 6474–6483. [Google Scholar] [CrossRef] [PubMed]
- Halleen, F.; Fourie, P.H.; Lombard, P.J. Protection of grapevine pruning wounds against Eutypa lata by biological and chemical methods. S. Afr. J. Enol. Vitic. 2010, 31, 125–132. [Google Scholar] [CrossRef]
- Sosnowski, M.R.; Creaser, M.L.; Wicks, T.J.; Lardner, R.; Scott, E.S. Protection of grapevine pruning wounds from infection by Eutypa lata. Aust. J. Grape Wine Res. 2008, 14, 134–142. [Google Scholar] [CrossRef]
- Díaz, G.A.; Latorre, B.A. Efficacy of paste and liquid fungicide formulations to protect pruning wounds against pathogens associated with grapevine trunk diseases in Chile. Crop Prot. 2013, 46, 106–112. [Google Scholar] [CrossRef]
- Mondello, V.; Songy, A.; Battiston, E.; Pinto, C.; Coppin, C.; Trotel-Aziz, P.; Clément, C.; Mugnai, L.; Fontaine, F. Grapevine trunk diseases: A review of fifteen years of trials for their control with chemicals and biocontrol agents. Plant Dis. 2018, 102, 1189–1217. [Google Scholar] [CrossRef]
- La Torre, A.; Iovino, V.; Caradonia, F. Copper in plant protection: Current situation and prospects. Phytopathol. Mediterr. 2018, 57, 201–236. [Google Scholar]
- Mondello, V.; Lemaître-Guillier, C.; Trotel-Aziz, P.; Gougeon, R.; Acedo, A.; Schmitt-Kopplin, P.; Adrian, M.; Pinto, C.; Fernandez, O.; Fontaine, F. Assessment of a new copper-based formulation to control esca disease in field and study of its impact on the vine microbiome, vine physiology and enological parameters of the juice. J. Fungi 2022, 8, 151. [Google Scholar] [CrossRef]
- Pinto, C.; Custódio, V.; Nunes, M.; Songy, A.; Rabenoelina, F.; Courteaux, B.; Clément, C.; Gomes, A.C.; Fontaine, F. Understand the potential Role of Aureobasidium pullulans, a resident microorganism from grapevine, to prevent the infection caused by Diplodia seriata. Front. Microbiol. 2018, 9, 3047. [Google Scholar] [CrossRef]
- Del Frari, G.; Cabral, A.; Nascimento, T.; Boavida Ferreira, R.; Oliveira, H. Epicoccum layuense a potential biological control agent of esca-associated fungi in grapevine. PLoS ONE 2019, 14, e0213273. [Google Scholar] [CrossRef]
- Di Marco, S.; Osti, F.; Cesari, A. Experiments on the control of esca by Trichoderma. Phytopathol. Mediterr. 2004, 43, 108–115. [Google Scholar]
- Yacoub, A.; Gerbore, J.; Magnin, N.; Chambon, P.; Dufour, M.C.; Corio-Costet, M.F.; Guyoneaud, R.; Rey, P. Ability of Pythium oligandrum strains to protect Vitis vinifera L., by inducing plant resistance against Phaeomoniella chlamydospora, a pathogen involved in esca, a grapevine trunk disease. Biol. Control 2016, 92, 7–16. [Google Scholar] [CrossRef]
- Di Marco, S.; Metruccio, E.G.; Moretti, S.; Nocentini, M.; Carella, G.; Pacetti, A.; Battiston, E.; Osti, F.; Mugnai, L. Activity of Trichoderma asperellum strain ICC 012 and Trichoderma gamsii strain ICC 080 toward diseases of esca complex and associated pathogens. Front. Microbiol. 2022, 12, 813410. [Google Scholar] [CrossRef] [PubMed]
- Langa-Lomba, N.; Martín-Ramos, P.; Casanova-Gascón, J.; Julián-Lagunas, C.; González-García, V. Potential of native Trichoderma strains as antagonists for the control of fungal wood pathologies in young grapevine plants. Agronomy 2022, 12, 336. [Google Scholar] [CrossRef]
- Blundell, R.; Eskalen, A. Evaluation of biological and chemical pruning wound protectants to control grapevine trunk disease pathogens Eutypa lata and Neofusicoccum parvum. Plant Health Prog. 2022, 23, 197–205. [Google Scholar] [CrossRef]
- Pollard-flamand, J.; Boul, J.; Hart, M.; Úrbez-Torres, J.R. Biocontrol activity of Trichoderma species isolated from grapevines in British Columbia against Botryosphaeria Dieback fungal pathogens. J. Fungi 2022, 8, 409. [Google Scholar] [CrossRef]
- Pacifico, D.; Squartini, A.; Crucitti, D.; Barizza, E.; Lo Schiavo, F.; Muresu, R.; Carimi, F.; Zottini, M. The role of the endophytic microbiome in the grapevine response to environmental triggers. Front. Plant. Sci. 2019, 10, 1256. [Google Scholar] [CrossRef]
- Bettenfeld, P.; Cadena, J.; Jacquens, L.; Fernandez, O.; Fontaine, F.; van Schaik, E.; Courty, P.-E.; Trouvelot, S. The microbiota of the grapevine holobiont: A key component of plant health. J. Adv. Res. 2021, 40, 1–15. [Google Scholar] [CrossRef]
- Fournier, P.; Pellan, L.; Barroso-bergadà, D.; Labarthe, S.; Vacher, C. The functional microbiome of grapevine throughout plant evolutionary history and lifetime. Adv. Ecol. Res. 2022, 67, 27–99. [Google Scholar]
- Del Frari, G.; Gobbi, A.; Aggerbeck, M.R.; Oliveira, H.; Hansen, L.H.; Ferreira, R.B. Fungicides and the grapevine wood mycobiome: A case study on tracheomycotic ascomycete Phaeomoniella chlamydospora reveals potential for two novel control strategies. Front. Plant. Sci. 2019, 10, 1405. [Google Scholar] [CrossRef]
- Noel, Z.A.; Longley, R.; Maria, G.; Benucci, N. Non-target impacts of fungicide disturbance on phyllosphere yeasts in conventional and no-till management. ISME Commun. 2022, 2, 19. [Google Scholar] [CrossRef]
- Gobbi, A.; Kyrkou, I.; Filippi, E.; Ellegaard-Jensen, L.; Hansen, L.H. Seasonal epiphytic microbial dynamics on grapevine leaves under biocontrol and copper fungicide treatments. Sci. Rep. 2020, 10, 681. [Google Scholar] [CrossRef] [PubMed]
- Bettenfeld, P.; Fontaine, F.; Trouvelot, S.; Fernandez, O.; Courty, P. Woody plant declines. What’s wrong with the microbiome? Trends Plant Sci. 2020, 25, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Del Frari, G.; Gobbi, A.; Aggerbeck, M.R.; Oliveira, H.; Hansen, L.H.; Ferreira, R.B. Characterization of the wood mycobiome of Vitis vinifera in a vineyard affected by esca. Spatial distribution of fungal communities and their putative relation with leaf symptoms. Front. Plant. Sci. 2019, 10, 910. [Google Scholar] [CrossRef] [PubMed]
- Vaz, A.T.; Del Frari, G.; Chagas, R.; Oliveira, H.; Boavida Ferreira, R. Precise nondestructive location of defective woody tissue in grapevines affected by wood diseases. Phytopathol. Mediterr. 2020, 59, 441–451. [Google Scholar]
- Catoni, M.; Falsone, G.; Bonifacio, E. Assessing the origin of carbonates in a complex soil with a suite of analytical methods. Geoderma 2012, 175–176, 47–57. [Google Scholar] [CrossRef]
- Cenis, J.L. Rapid extraction of fungal DNA for PCR amplification. Nucleic Acids Res. 1992, 20, 2380. [Google Scholar] [CrossRef]
- Scibetta, S.; Schena, L.; Abdelfattah, A.; Pangallo, S.; Cacciola, S.O. Selection and experimental evaluation of universal primers to study the fungal microbiome of higher plants. Phytobiomes J. 2018, 2, 225–236. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Gobbi, A.; Santini, R.; Filippi, E.; Ellegaard-Jensen, L.; Jacobsen, C.S.; Hansen, L.H. Quantitative and qualitative evaluation of the impact of the G2 enhancer, bead sizes and lysing tubes on the bacterial community composition during DNA extraction from recalcitrant soil core samples based on community sequencing and qPCR. PLoS ONE 2019, 14, e0200979. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Holmes, S.P. Exact sequence variants should replace operational taxonomic units in marker-gene data analysis. ISME J. 2017, 11, 2639–2643. [Google Scholar] [CrossRef]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Caporaso, G.J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 1–17. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Paulson, J.N. Biomformat: An Interface Package for the BIOM File Format; Bioconductor, Bioconductor.org: 2016; p. 19.
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.0-10. 2013. Available online: http://CRAN.R-project.org/package=vegan (accessed on 1 October 2022).
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Wang, A.Y.; Naumann, U.; Wright, S.; Eddelbuettel, D.; Warton, D.; Wright, S. Mvabund: Statistical Methods for Analysing Multivariate Abundance Data; 2017. pp. 1–95. Available online: https://cran.r-project.org/package=mvabund (accessed on 1 October 2022).
- Foster, Z.S.L.; Sharpton, T.; Grunwald, N.J. MetacodeR: An R package for manipulation and heat tree visualization of community taxonomic data from metabarcoding. bioRxiv 2017, 13, 071019. [Google Scholar]
- Foster, Z.S.L.; Chamberlain, S.; Grunwald, N.J. Taxa: An R package implementing data standards and methods for taxonomic data. F1000Research 2018, 7, 272. [Google Scholar] [CrossRef]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Lahti, L.; Sudarshan, S. Tools for Microbiome Analysis in R. In Microbiome Package Version Bioconductor, 1.8.0.
- Valero-Mora, P.M. ggplot2: Elegant Graphics for Data Analysis. J. Stat. Softw. Book Rev. 2010, 35, 1–3. [Google Scholar] [CrossRef]
- Martinez Arbizu, P. pairwiseAdonis: Pairwise multilevel comparison using adonis. In R package version 0.4; 2020; p. 1.
- Bustamante, M.I.; Elfar, K.; Smith, R.J.; Bettiga, L.J.; Tian, T.; Torres, G.A.; Eskalen, A. First report of Fusarium annulatum associated with young vine decline in California. Plant Dis. 2022, 106, 2752. [Google Scholar] [CrossRef]
- Lawrence, D.P.; Travadon, R.; Pouzoulet, J.; Rolshausen, P.E.; Wilcox, W.F. Characterization of Cytospora isolates from wood cankers of declining grapevine in North America, with the descriptions of two new Cytospora species. Plant Pathol. 2017, 66, 713–725. [Google Scholar] [CrossRef]
- Bruez, E.; Vallance, J.; Gautier, A.; Laval, V.; Compant, S.; Maurer, W.; Sessitsch, A.; Lebrun, M.-H.; Rey, P. Major changes in grapevine wood microbiota are associated with the onset of esca, a devastating trunk disease. Environ. Microbiol. 2020, 22, 5189–5206. [Google Scholar] [CrossRef] [PubMed]
- Cobos, R.; Ibañez, A.; Diez-gal, A.; Calvo-peña, C.; Ghoreshizadeh, S.; Coque, J.J.R. The grapevine microbiome to the rescue: Implications for the biocontrol of trunk diseases. Plants 2022, 11, 840. [Google Scholar] [CrossRef] [PubMed]
- Morgan, H.H.; du Toit, M.; Setati, M.E. The grapevine and wine microbiome: Insights from high-throughput amplicon sequencing. Front. Microbiol. 2017, 8, 820. [Google Scholar] [CrossRef]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Laurent, B.; Marchand, M.; Chancerel, E.; Saint-jean, G.; Capdevielle, X.; Poeydebat, C.; Bellee, A.; Comont, G.; Villate, L.; Desprez-Loustau, M.-L. A richer community of Botryosphaeriaceae within a less diverse community of fungal endophytes in grapevines than in adjacent forest trees revealed by a mixed metabarcoding strategy. Phytobiomes J. 2020, 4, 252–267. [Google Scholar] [CrossRef]
- Deyett, E.; Rolshausen, P.E. Endophytic microbial assemblage in grapevine. FEMS Microbiol. Ecol. 2020, 96, fiaa053. [Google Scholar] [CrossRef]
- Martínez-Diz, P.; Eichmeier, A.; Spetik, M.; Bujanda, R.; Díaz-Losada, E.; Gramaje, D. Grapevine pruning time affects natural wound colonization by wood-invading fungi. Fungal Ecol. 2020, 48, 100994. [Google Scholar] [CrossRef]
- González, V.; Tello, M.L. The endophytic mycota associated with Vitis vinifera in central Spain. Fungal Divers. 2011, 47, 29–42. [Google Scholar] [CrossRef]
- Kraus, C.; Voegele, R.T.; Fischer, M. Temporal development of the culturable, endophytic fungal community in healthy grapevine branches and occurrence of GTD-associated fungi. Microb. Ecol. 2019, 77, 866–876. [Google Scholar] [CrossRef]
- Pancher, M.; Ceol, M.; Corneo, P.E.; Longa, C.M.O.; Yousaf, S.; Pertot, I.; Campisano, A. Fungal endophytic communities in grapevines (Vitis vinifera L.) respond to crop management. Appl. Environ. Microbiol. 2012, 78, 4308–4317. [Google Scholar] [CrossRef]
- Hardoim, P.R.; Van Overbeek, L.S.; Berg, G.; Pirttilä, M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed]
- Milanovi, V.; Cardinali, F.; Ferrocino, I.; Boban, A.; Franciosa, I.; Mucalo, A.; Osimani, A.; Aquilanti, L.; Garofalo, C.; Budic-Leto, I. Croatian white grape variety Marastina: First taste of its indigenous mycobiota. Food Res. Int. 2022, 162, 111917. [Google Scholar] [CrossRef] [PubMed]
- Pinto, C.; Pinho, D.; Sousa, S.; Pinheiro, M.; Egas, C.; Gomes, A.C. Unravelling the diversity of grapevine microbiome. PLoS ONE 2014, 9, e85622. [Google Scholar] [CrossRef] [PubMed]
- Cureau, N.; Threlfall, R.; Savin, M.; Marasini, D.; Lavefve, L.; Carbonero, F. Year, location, and variety impact on grape-, soil-, and leaf-associated fungal microbiota of Arkansas-grown table grapes. Microb. Ecol. 2021, 82, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, E.; Bekris, F.; Vasileiadis, S.; Karpouzas, D.G.; Papadopoulou, K.K. Different factors are operative in shaping the epiphytic grapevine microbiome across different geographical scales: Biogeography, cultivar or vintage ? J. Sustain. Agric. Environ. 2022, 1, 287–301. [Google Scholar] [CrossRef]
- Singh, P.; Gobbi, A.; Santoni, S.; Hansen, L.H.; This, P.; Péros, J. Assessing the impact of plant genetic diversity in shaping the microbial community structure of Vitis vinifera phyllosphere in the Mediterranean. Front. Life Sci. 2018, 11, 35–46. [Google Scholar] [CrossRef]
- Bekris, F.; Vasileiadis, S.; Papadopoulou, E.; Samaras, A.; Testempasis, S.; Gkizi, D.; Tavlaki, G.; Tzima, A.; Paplomatas, E.; Markakis, E.; et al. Grapevine wood microbiome analysis identifies key fungal pathogens and potential interactions with the bacterial community implicated in grapevine trunk disease appearance. Environ. Microbiome 2021, 16, 23. [Google Scholar] [CrossRef]
- Berlanas, C.; Berbegal, M.; Elena, G.; Laidani, M.; Cibriain, J.F.; Sagües, A.; Gramaje, D. The fungal and bacterial rhizosphere microbiome associated with grapevine rootstock genotypes in mature and young vineyards. Front. Microbiol. 2019, 10, 1142. [Google Scholar] [CrossRef]
- Awad, M.; Giannopoulos, G.; Mylona, P.V.; Polidoros, A.N. Comparative analysis of grapevine epiphytic microbiomes among different varieties, tissues, and developmental stages in the same terroir. Appl. Sci. 2023, 13, 102. [Google Scholar] [CrossRef]
- Swift, J.F.; Hall, M.E.; Harris, Z.N.; Kwasniewski, M.T.; Miller, A.J. Grapevine microbiota reflect diversity among compartments and complex interactions within and among root and shoot systems. Microorganisms 2021, 9, 92. [Google Scholar] [CrossRef]
- Lade, S.B.; Štraus, D.; Oliva, J. Variation in fungal community in grapevine (Vitis vinifera) nursery stock depends on nursery, variety and rootstock. J. Fungi 2022, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Gramaje, D.; Eichmeier, A.; Spetik, M.; Bujanda, R.; Vallance, J.; Rey, P. Exploring the temporal dynamics of the fungal microbiome in rootstocks, the lesser-known half of the grapevine crop. J. Fungi 2022, 8, 421. [Google Scholar] [CrossRef] [PubMed]
- Travadon, R.; Lecomte, P.; Diarra, B.; Lawrence, D.P.; Renault, D.; Ojeda, H.; Rey, P.; Baumgartner, K. Grapevine pruning systems and cultivars influence the diversity of wood-colonizing fungi. Fungal Ecol. 2016, 24, 82–93. [Google Scholar] [CrossRef]
- Costa, J. Evaluation of Chemicals against Esca Related Pathogens, In Vitro and As Pruning Wound Protectants. Master’s Thesis, University of Lisbon, Lisbon, Portugal, 2018. [Google Scholar]
- Colombo, A. Protection of Grapevine Pruning Wounds. Effect on Esca Progression and on Wood Secondary Metabolites. Master’s Thesis, University of Lisbon, Lisbon, Portugal, 2020. [Google Scholar]
- Cascavilla, V. Evaluation of Non-Conventional Strategies for Pruning Wound Protection in Grapevine. Master’s Thesis, University of Lisbon, Lisbon, Portugal, 2022. [Google Scholar]
- McCann, K.S. The diversity–stability debate. Nature 2000, 405, 228–233. [Google Scholar] [CrossRef]
- Deyett, E.; Rolshausen, P. Temporal dynamics of the sap microbiome of grapevine under high Pierce’s disease pressure. Front Plant Sci 2019, 10, 1246. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trade Name | Manufacturer | Active Ingredient/BCA | Tested Concentration |
|---|---|---|---|
| Cuprocol® | Syngenta (Basel, Switzerland) | Copper oxychloride | 36.5 g L−1 |
| Esquive® | Agrauxine (Marcq-en-Barœul, France) | Trichoderma atroviride I-1237 (1 × 108 CFU g−1) | 100.0 g L−1 |
| Tessior® | BASF (Ludwigshafen, Germany) | Boscalid 10 g L−1 + Pyraclostrobin 5 g L−1 | 10.0 g L−1 boscalid + 5.0 g L−1 pyraclostrobin |
| Bentogran® | AEB Bioq. Port. (Singapore) | Sodium bentonite | 80% (v/v) sodium bentonite in water |
| Phylum | Family | Genus | RA (%) | Vineyard | Cultivar | GTD | ||
|---|---|---|---|---|---|---|---|---|
| Almotivo | DISAFA | Cabernet Sauvignon | Syrah | |||||
| Ascomycetes | Pleosporaceae | Alternaria | 30.0 | + | + | + | + | − |
| Stemphylium | 4.0 | + | − | + | + | − | ||
| Davidiellaceae | Cladosporium | 17.0 | + | + | + | + | − | |
| Aureobasidiaceae | Aureobasidium | 13.4 | + | + | + | + | − | |
| Didymellaceae | Epicoccum | 11.6 | + | + | + | + | − | |
| Phoma | 1.4 | + | + | + | + | − | ||
| Debaryomycetaceae | Debaryomyces | 3.3 | + | + | + | + | − | |
| Hypocreaceae | Trichoderma | 2.3 | + | + | + | + | − | |
| Saccharomycetaceae | Candida | 0.7 | + | + | + | + | − | |
| Valsaceae | Cytospora | 0.5 | + | - | + | + | + | |
| Hypocreales | Sarocladium | 0.5 | − | + | + | + | − | |
| Cucurbitariaceae | Neocucurbitaria | 0.3 | − | + | + | - | − | |
| Phaeosphaeriaceae | Neosetophoma | 0.2 | + | + | + | + | − | |
| Phaeomoniellaceae | Phaemoniella | 0.2 | + | + | + | + | + | |
| Mycosphaerellaceae | Ramularia | 0.1 | + | + | + | + | − | |
| Diatrypaceae | Eutypella | 0.1 | + | − | + | − | + | |
| Nectriaceae | Fusarium | 0.1 | + | + | + | + | + * | |
| Basidiomycetes | Bulleribasidiaceae | Vishniacozyma | 1.8 | + | + | + | + | − |
| Dioszegia † | 0.1 | + | + | + | + | − | ||
| Filobasidiaceae | Filobasidium | 1.3 | + | + | + | + | − | |
| Tricholomataceae | Merismodes | 0.5 | + | - | + | + | − | |
| Buckleyzymaceae | Buckleyzyma † | 0.4 | + | + | + | + | − | |
| Malasseziaceae | Malassezia | 0.4 | + | + | + | + | − | |
| Symmetrosporaceae | Symmetrospora † | 0.4 | + | + | + | + | − | |
| Sporidiobolaceae | Sporobolomyces | 0.3 | + | + | + | + | − | |
| Rhodotorula | 0.2 | + | + | + | + | − | ||
| n/a | Akenomyces † | 0.3 | + | + | − | + | − | |
| Cystofilobasidiaceae | Cystofilobasidium | 0.3 | − | + | − | + | − | |
| Rhynchogastremataceae | Papiliotrema † | 0.2 | + | + | + | + | − | |
| Kondoaceae | Kondoa † | 0.2 | + | − | + | + | − | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Del Frari, G.; Aggerbeck, M.R.; Gobbi, A.; Ingrà, C.; Volpi, L.; Nascimento, T.; Ferrandino, A.; Hansen, L.H.; Ferreira, R.B. Pruning Wound Protection Products Induce Alterations in the Wood Mycobiome Profile of Grapevines. J. Fungi 2023, 9, 488. https://doi.org/10.3390/jof9040488
Del Frari G, Aggerbeck MR, Gobbi A, Ingrà C, Volpi L, Nascimento T, Ferrandino A, Hansen LH, Ferreira RB. Pruning Wound Protection Products Induce Alterations in the Wood Mycobiome Profile of Grapevines. Journal of Fungi. 2023; 9(4):488. https://doi.org/10.3390/jof9040488
Chicago/Turabian StyleDel Frari, Giovanni, Marie Rønne Aggerbeck, Alex Gobbi, Chiara Ingrà, Lorenzo Volpi, Teresa Nascimento, Alessandra Ferrandino, Lars Hestbjerg Hansen, and Ricardo Boavida Ferreira. 2023. "Pruning Wound Protection Products Induce Alterations in the Wood Mycobiome Profile of Grapevines" Journal of Fungi 9, no. 4: 488. https://doi.org/10.3390/jof9040488