
Morphological and Phylogenetic Analyses Reveal Three New Species of Distoseptispora (Distoseptisporaceae, Distoseptisporales) from Yunnan, China

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Isolation, and Morphological Examination
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Sequence Alignment and Phylogenetic Analyses
3. Results
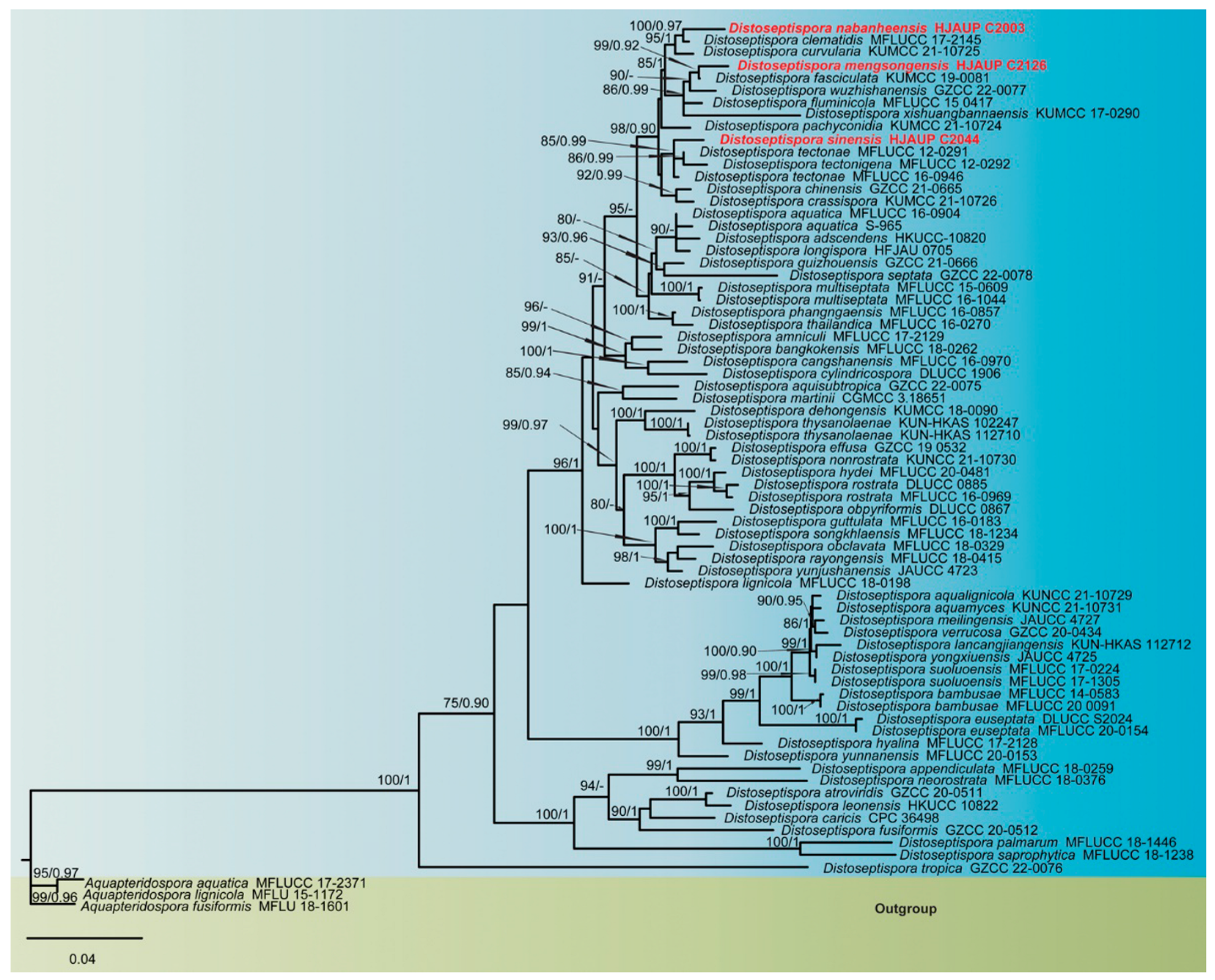
3.1. Molecular Phylogeny
3.2. Taxonomy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Su, H.Y.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Ariyawansa, H.A.; Luo, Z.L.; Promputtha, I.; Tian, Q.; Lin, C.G.; Shang, Q.J.; Zhao, Y.C.; et al. The families Distoseptisporaceae fam. nov., Kirschsteiniotheliaceae, Sporormiaceae and Torulaceae, with new species from freshwater in Yunnan Province, China. Fungal Divers. 2016, 80, 375–409. [Google Scholar] [CrossRef]
- Yang, J.; Maharachchikumbura, S.S.N.; Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Al-Sadi, A.M.; Liu, Z.Y. Pseudostanjehughesia aquitropica gen. et sp. nov. and Sporidesmium sensu lato species from freshwater habitats. Mycol. Prog. 2018, 17, 591–616. [Google Scholar] [CrossRef]
- Hyde, K.D.; Tennakoon, D.S.; Jeewon, R.; Bhat, D.J.; Maharachchikumbura, S.S.N.; Rossi, W.; Leonardi, M.; Lee, H.B.; Mun, H.Y.; Houbraken, J.; et al. Fungal diversity notes 1036–1150: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2019, 96, 1–242. [Google Scholar] [CrossRef]
- Index Fungorum. Available online: http://www.indexfungorum.org/Names/Names.asp (accessed on 1 March 2023).
- Monkai, J.; Boonmee, S.; Ren, G.C.; Wei, D.P.; Phookamsak, R.; Mortimer, P.E. Distoseptispora hydei sp. nov. (Distoseptisporaceae), a novel lignicolous fungus on decaying bamboo in Thailand. Phytotaxa 2020, 459, 93–107. [Google Scholar] [CrossRef]
- Zhai, Z.-J.; Yan, J.-Q.; Li, W.-W.; Gao, Y.; Hu, H.-J.; Zhou, J.-P.; Song, H.-Y.; Hu, D.-M. Three novel species of Distoseptispora (Distoseptisporaceae) isolated from bamboo in Jiangxi Province, China. Mycokeys 2022, 88, 35–54. [Google Scholar] [CrossRef]
- Dong, W.; Hyde, K.D.; Jeewon, R.; Doilom, M.; Yu, X.-D.; Wang, G.-N.; Liu, N.G.; Hu, D.M.; Nalumpang, S.; Zhang, H. Towards a natural classification of annulatascaceae-like taxa II: Introducing five new genera and eighteen new species from freshwater. Mycosphere 2021, 12, 1–88. [Google Scholar] [CrossRef]
- Xia, J.W.; Ma, Y.R.; Li, Z.; Zhang, X.G. Acrodictys-like wood decay fungi from southern China, with two new families Acrodictyaceae and Junewangiaceae. Sci. Rep. 2017, 7, 7888. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Hyde, K.; Liu, J.; Bhat, D.; Bao, D.; Li, W.; Su, H. Lignicolous freshwater fungi from China II: Novel Distoseptispora (Distoseptisporaceae) species from northwestern Yunnan Province and a suggested unified method for studying lignicolous freshwater fungi. Mycosphere 2018, 9, 444–461. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.C.; Bhat, D.J.; Phillips, A.J.L.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.S.; Dissanayake, A.J.; Tennakoon, D.S.; et al. Fungal diversity notes 840–928: Micro-fungi associated with Pandanaceae. Fungal Divers. 2018, 93, 1–160. [Google Scholar] [CrossRef]
- Yang, J.; Liu, L.-L.; Jones, E.B.G.; Li, W.-L.; Hyde, K.D.; Liu, Z.-Y. Morphological variety in Distoseptispora and introduction of six novel species. J. Fungi 2021, 7, 945. [Google Scholar] [CrossRef]
- Hyde, K.D.; Suwannarach, N.; Jayawardena, R.S.; Manawasinghe, I.S.; Liao, C.F.; Doilom, M.; Cai, L.; Zhao, P.; Buyck, B.; Phukhamsakda, C.; et al. Mycosphere notes 325–344—Novel species and records of fungal taxa from around the world. Mycosphere 2021, 12, 1101–1156. [Google Scholar] [CrossRef]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspé, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef] [Green Version]
- Song, H.Y.; El Sheikha, A.F.; Zhai, Z.J.; Zhou, J.P.; Chen, M.H.; Huo, G.H.; Huang, X.G.; Hu, D.M. Distoseptispora longispora sp. nov. from freshwater habitats in China. Mycotaxon 2020, 135, 513–523. [Google Scholar] [CrossRef]
- Sun, Y.; Goonasekara, I.D.; Thambugala, K.M.; Jayawardena, R.S.; Wang, Y.; Hyde, K.D. Distoseptispora bambusae sp. nov. (Distoseptisporaceae) on bamboo from China and Thailand. Biodivers. Data J. 2020, 8, e53678. [Google Scholar] [CrossRef]
- Li, W.-L.; Liu, Z.-P.; Zhang, T.; Dissanayake, A.J.; Luo, Z.-L.; Su, H.-Y.; Liu, J.-K. Additions to Distoseptispora (Distoseptisporaceae) associated with submerged decaying wood in China. Phytotaxa 2021, 520, 75–86. [Google Scholar] [CrossRef]
- Shen, H.W.; Bao, D.F.; Hyde, K.D.; Su, H.Y.; Bhat, D.J.; Luo, Z.L. Two novel species and two new records of Distoseptispora from freshwater habitats in China and Thailand. MycoKeys 2021, 84, 79–101. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Hyde, K.D.; Wang, S.; Sun, Y.R.; Suwannarach, N.; Sysouphanthong, P.; Abdel-Wahab, M.A.; Abdel-Aziz, F.A.; Abeywickrama, P.D.; Abreu, V.P.; et al. Fungal diversity notes 1512–1610: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2022, 117, 1–272. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; Nilsson, R.H.; Bhunjun, C.S.; de Farias, A.R.G.; Sun, Y.-R.; Wijesinghe, S.N.; Raza, M.; Bao, D.-F.; Lu, L.; Tibpromma, S.; et al. The numbers of fungi: Contributions from traditional taxonomic studies and challenges of metabarcoding. Fungal Divers. 2022, 114, 327–386. [Google Scholar] [CrossRef]
- Ma, J.; Zhang, J.-Y.; Xiao, X.-J.; Xiao, Y.-P.; Tang, X.; Boonmee, S.; Kang, J.-C.; Lu, Y.-Z. Multi-gene phylogenetic analyses revealed five new species and two new records of Distoseptisporales from China. J. Fungi 2022, 8, 1202. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, R.; Qing, Y.; Yang, H.; Li, C.; Wang, G.; Zhang, D.; Ning, P. Polyphasic identification of Distoseptispora with six new species from fresh water. J. Fungi 2022, 8, 1063. [Google Scholar] [CrossRef]
- Luo, Z.-L.; Hyde, K.D.; Liu, J.-K.J.; Maharachchikumbura, S.S.N.; Jeewon, R.; Bao, D.-F.; Bhat, D.J.; Lin, C.-G.; Li, W.-L.; Yang, J.; et al. Freshwater Sordariomycetes. Fungal Divers. 2019, 99, 451–660. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Maharachchikumbura, S.S.N.; Hyde, K.D.; Bhat, D.J.; McKenzie, E.H.C.; Bahkali, A.H.; Gareth Jones, E.B.G.; Liu, Z.Y. Aquapteridospora lignicola gen. et sp. nov., a new hyphomycetous taxon (Sordariomycetes) from wood submerged in a freshwater stream. Cryptogam. Mycol. 2015, 36, 469–478. [Google Scholar] [CrossRef]
- Hyde, K.D.; Bao, D.F.; Hongsanan, S.; Chethana, K.; Yang, J.; Suwannarach, N. Evolution of freshwater Diaporthomycetidae (Sordariomycetes) provides evidence for five new orders and six new families. Fungal Divers. 2021, 107, 71–105. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Crous, P.; Wingfield, M.; Lombard, L.; Roets, F.; Swart, W.; Alvarado, P.; Carnegie, A.; Moreno, G.; Luangsaard, J.; Thangavel, R.; et al. Fungal Planet description sheets: 951–1041. Persoonia 2019, 43, 223–425. [Google Scholar] [CrossRef]
- Wong, K.M.K.; Goh, T.K.; Hodgkiss, I.J.; Hyde, K.D.; Ranghoo, V.M.; Tsui, C.K.M.; Ho, W.H.; Wong, S.W.; Yuen, T.K. Role of fungi in freshwater ecosystems. Biodivers. Conserv. 1998, 7, 1187–1206. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Ba, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Ma, J.; Wang, Y.; Ma, L.G.; Zhang, Y.D.; Castañeda-Ruíz, R.F.; Zhang, X.G. Three new species of Neosporidesmium from Hainan, China. Mycol. Prog. 2011, 10, 157–162. [Google Scholar] [CrossRef]
- Goh, T.K. Single-spore isolation using a hand-made glass needle. Fungal Divers. 1999, 2, 47–63. [Google Scholar]
- Rayner, R.W. A Mycological Colour Chart; Commonwealth Mycological Institute, Kew, Surrey & British Mycological Society: Kew, UK, 1970. [Google Scholar]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Rehner, S.A.; Buckley, E. A beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Gao, F.L.; Jakovli’c, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2017, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Shenoy, B.D.; Jeewon, R.; Wu, W.P.; Bhat, D.J.; Hyde, K.D. Ribosomal and RPB2 DNA sequence analyses suggest that Sporidesmium and morphologically similar genera are polyphyletic. Mycol. Res. 2006, 110, 916–928. [Google Scholar] [CrossRef] [PubMed]
- Phukhamsakda, C.; McKenzie, E.H.C.; Phillips, A.J.L.; Jones, E.B.G.; Bhat, D.J.; Marc, S.; Bhunjun, C.S.; Wanasinghe, D.N.; Thongbai, B.; Camporesi, E.; et al. Microfungi associated with Clematis (Ranunculaceae) with an integrated approach to delimiting species boundaries. Fungal Divers. 2020, 102, 1–203. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Konta, S.; Tibpromma, S.; Karunarathna, S.C.; Samarakoon, M.C.; Steven, L.S.; Mapook, A.; Boonmee, S.; Senwanna, C.; Balasuriya, A.; Eungwanichayapant, P.D.; et al. Morphology and multigene phylogeny reveal ten novel taxa in Ascomycota from terrestrial palm substrates (Arecaceae) in Thailand. Mycosphere 2023, 14, 107–152. [Google Scholar]
- Farr, D.F.; Rossman, A.Y. Fungal Databases. Systematic Mycology and Microbiology Laboratory, ARS, USDA. Available online: http://nt.ars-grin.gov/fungaldatabases/ (accessed on 1 March 2023).
- Wu, W.; Zhuang, W. Sporidesmium, Endophragmiella and Related Genera from China; Fungal Diversity Press: Hong Kong, China, 2005. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Voucher/Strain Number | GenBank Accession Numbers | References | ||
|---|---|---|---|---|---|
| LSU | ITS | TEF1 | |||
| Aquapteridospora aquatica | MFLUCC 17-2371T | MW287767 | MW286493 | — | [7] |
| A. fusiformis | MFLUCC 18-1606T | MK849798 | MK828652 | MN194056 | [22] |
| A. lignicola | MFLUCC 15-0377T | KU221018 | — | — | [23] |
| Distoseptispora adscendens | HKUCC 10820 | DQ408561 | — | — | [40] |
| D. amniculi | MFLUCC 17-2129T | MZ868761 | MZ868770 | — | [11] |
| D. appendiculata | MFLUCC 18-0259T | MN163023 | MN163009 | MN174866 | [22] |
| D. aqualignicola | KUNCC 21-10729T | ON400845 | OK341186 | OP413480 | [21] |
| D. aquamyces | KUNCC 21-10731T | OK341199 | OK341187 | OP413482 | [21] |
| D. aquatica | MFLUCC 16-0904 | MK849794 | MK828649 | — | [22] |
| D. aquatica | S-965 | MK849792 | MK828647 | MN194051 | [22] |
| D. aquisubtropica | GZCC 22-0075T | ON527941 | ON527933 | ON533677 | [20] |
| D. atroviridis | GZCC 20-0511T | MZ868763 | MZ868772 | MZ892978 | [11] |
| D. bambusae | MFLUCC 20-0091T | MT232718 | MT232713 | MT232880 | [15] |
| D. bambusae | MFLUCC 14-0583 | MT232717 | MT232712 | — | [15] |
| D. bangkokensis | MFLUCC 18-0262T | MZ518206 | MZ518205 | — | [17] |
| D. cangshanensis | MFLUCC 16-0970T | MG979761 | MG979754 | MG988419 | [9] |
| D. caricis | CPC 36498T | MN567632 | NR_166325 | — | [26] |
| D. chinensis | GZCC 21-0665T | MZ474867 | MZ474871 | MZ501609 | [12] |
| D. clematidis | MFLUCC 17-2145T | MT214617 | MT310661 | — | [41] |
| D. crassispora | KUMCC 21-10726T | OK341196 | OK310698 | OP413479 | [21] |
| D. curvularia | KUMCC 21-10725T | OK341195 | OK310697 | OP413478 | [21] |
| D. cylindricospora | DLUCC 1906T | OK513523 | OK491122 | OK524220 | [19] |
| D. dehongensis | KUMCC 18-0090T | MK079662 | MK085061 | MK087659 | [3] |
| D. effusa | GZCC 19-0532T | MZ227224 | MW133916 | MZ206156 | [11] |
| D. euseptata | MFLUCC 20–0154T | MW081544 | MW081539 | — | [16] |
| D. euseptata | DLUCC S2024 | MW081545 | MW081540 | MW084994 | [16] |
| D. fasciculata | KUMCC 19-0081T | MW287775 | MW286501 | MW396656 | [7] |
| D. fluminicola | MFLUCC 15-0417T | KU376270 | MF077553 | — | [9] |
| D. fusiformis | GZCC 20-0512T | MZ868764 | MZ868773 | MZ892979 | [11] |
| D. guizhounesis | GZCC 21-0666T | MZ474869 | MZ474868 | MZ501610 | [12] |
| D. guttulata | MFLUCC 16-0183T | MF077554 | MF077543 | MF135651 | [2] |
| D. hyalina | MFLUCC 17-2128T | MZ868760 | MZ868769 | MZ892976 | [11] |
| D. hydei | MFLUCC 20-0481T | MT742830 | MT734661 | — | [5] |
| D. lancangjiangensis | KUN-HKAS 112712T | MW879522 | MW723055 | — | [17] |
| D. leonensis | HKUCC 10822T | DQ408566 | — | — | [40] |
| D. lignicola | MFLUCC 18-0198T | MK849797 | MK828651 | — | [22] |
| D. longispora | HFJAU 0705T | MH555357 | MH555359 | — | [14] |
| D. martinii | CGMCC 3.18651T | KX033566 | KU999975 | — | [8] |
| D. meilingensis | JAUCC 4727T | OK562396 | OK562390 | OK562408 | [6] |
| D. mengsongensis | HJAUP C2126T | OP787874 | OP787876 | OP961937 | This study |
| D. multiseptata | MFLUCC 15-0609T | KX710140 | KX710145 | MF135659 | [42] |
| D. multiseptata | MFLUCC 16-1044 | MF077555 | MF077544 | MF135652 | [2] |
| D. nabanheensis | HJAUP C2003T | OP787877 | OP787873 | OP961935 | This study |
| D. neorostrata | MFLUCC 18-0376T | MN163017 | MN163008 | — | [22] |
| D. nonrostrata | KUNCC 21-10730T | OK341198 | OK310699 | OP413481 | [21] |
| D. obclavata | MFLUCC 18-0329T | MN163010 | MN163012 | — | [22] |
| D. obpyriformis | DLUCC 0867 | MG979765 | MG979757 | MG988423 | [9] |
| D. pachyconidia | KUMCC 21-10724T | OK341194 | OK310696 | OP413477 | [21] |
| D. palmarum | MFLUCC 18-1446T | MK079663 | MK085062 | MK087660 | [3] |
| D. phangngaensis | MFLUCC 16-0857T | MF077556 | MF077545 | MF135653 | [2] |
| D. rayongensis | MFLUCC 18-0415T | MH457137 | MH457172 | MH463253 | [28] |
| D. rostrata | MFLUCC 16-0969T | MG979766 | MG979758 | MG988424 | [9] |
| D. rostrata | DLUCC 0885 | MG979767 | MG979759 | MG988425 | [9] |
| D. saprophytica | MFLUCC 18-1238T | MW287780 | MW286506 | MW396651 | [7] |
| D. septate | GZCC 22-0078T | ON527947 | ON527939 | ON533683 | [20] |
| D. songkhlaensis | MFLUCC 18-1234T | MW287755 | MW286482 | MW396642 | [7] |
| D. sinensis | HJAUP C2044T | OP787875 | OP787878 | OP961936 | This study |
| D. suoluoensis | MFLUCC 17-0224T | MF077557 | MF077546 | MF135654 | [2] |
| D. suoluoensis | MFLUCC 17-1305 | MF077558 | MF077547 | — | [2] |
| D. tectonae | MFLUCC 12-0291T | KX751713 | KX751711 | KX751710 | [42] |
| D. tectonae | MFLUCC 16-0946 | MG979768 | MG979760 | MG988426 | [9] |
| D. tectonigena | MFLUCC 12-0292T | KX751714 | KX751712 | — | [42] |
| D. thailandica | MFLUCC 16-0270T | MH260292 | MH275060 | MH412767 | [10] |
| D. thysanolaenae | KUN-HKAS 112,710 | MW879524 | MW723057 | MW729783 | [17] |
| D. thysanolaenae | KUN-HKAS 102247T | MK064091 | MK045851 | MK086031 | [13] |
| D. tropica | GZCC 22-0076T | ON527943 | ON527935 | ON533679 | [20] |
| D. verrucosa | GZCC 20-0434T | MZ868762 | MZ868771 | MZ892977 | [11] |
| D. wuzhishanensis | GZCC 22-0077T | ON527946 | ON527938 | ON533682 | [20] |
| D. xishuangbannaensis | KUMCC 17-0290T | MH260293 | MH275061 | MH412768 | [10] |
| D. yongxiuensis | JAUCC 4725T | OK562394 | OK562388 | OK562406 | [6] |
| D. yunjushanensis | JAUCC 4723T | OK562398 | OK562392 | OK562410 | [6] |
| D. yunnanensis | MFLUCC 20-0153T | MW081546 | MW081541 | MW084995 | [16] |
| Species | Conidiophores (μm) | Conida | Habitat | Host/Locality | References | |
|---|---|---|---|---|---|---|
| Size (µm) | Morphology | |||||
| Distoseptispora adscendens | 20–50 × 7–10 | 110–375 × 14–20 | Cylindrical to obclavate, rostrate, brown to dark brown, paler towards the apex, 16–62-distoseptate | Terrestrial | Rotten wood and dead branches of many woody plant species, China | [21,45] |
| D. amniculi | 90–180 × 3–4.5 | 85–167 × 9–11.8 | Obclavate, rostrate, olivaceous-brown, grayish-brown or mid-brown, sometimes with a secondary conidium, (7–)12–24-distoseptate | Freshwater | Unidentified submerged wood, Thailand | [11] |
| D. appendiculata | 62–86 × 4.5–5.5 | 67–89 × 10–16 | Obpyriform or obclavate, olivaceous or dark brown, with a gelatinous sheath around tip, 13–17-distoseptate | Freshwater | Unidentified submerged wood, Thailand | [22] |
| D. aqualignicola | 90–190(–240) × 5–8 | 41–94(–104) × 10.5–12.5 | Obclavate, rostrate, brown at the base, subhyaline to pale-brown at the apex, 4–8-euseptate | Freshwater | Unidentified submerged wood, China | [21] |
| D. aquamyces | (78–)91–198 × 4–7 | 30–95 × 7–12 | Obclavate to obpyriform, mostly rostrate, pale-brown to brown at the base, subhyaline to pale-brown at the apex, 4–10-euseptate, verrucose | Freshwater | Unidentified submerged wood, China | [21] |
| D. aquatica | 29–41 × 7–9 | 110–157 × 13.5–16.5 | Obclavate, dark brown with bluish-green to malachite green tinge, paler towards the apex, 15–28-distoseptate | Freshwater | Unidentified submerged wood, China | [1] |
| D. aquisubtropica | 16–83 × 5–11 | 43–278 × 11–19 | Obclavate or lanceolate, rostrate, pale brown or dark brown, olivaceous,16–31-distoseptate, verrucose | Freshwater | Unidentified submerged wood, China | [20] |
| D. atroviridis | 100–167 × 2.7–4 | 31–43 × 13–20 | Ellipsoidal to obovoid, dark green, subhyaline at the basal cell, 6-distoseptate | Freshwater | Unidentified submerged wood, China | [11] |
| D. bambusae | 40–96 × 4–5.5 | 45–74 × 5.5–9.5 | Obclavate, olivaceous or brown, 5–10-distoseptate | Terrestrial | Dead bamboo culms, China | [15] |
| D.bambusicola | 64–116 × 4–7 | 72–193 × 7.5–14.5 | Obclavate or lanceolate, rostrate, pale brown, up to 16-distoseptate | Freshwater | Unidentified submerged decaying bamboo culms, China | [18] |
| D. bangkokensis | 37–55 × 3–4 | 400–568 × 13–16 | Obclavate, rostrate, dark olivaceous to dark brown, conidia percurrent proliferation, which forms another conidium at the apex, multi-distoseptate | Freshwater | Unidentified submerged wood, Thailand | [17] |
| D. cangshanensis | 44–68 × 4–8 | 58–166(–287) × 10–14 | Obclavate or lanceolate, rostrate, olivaceous or brown, multi-distoseptate | Freshwater | Unidentified submerged wood, China | [9] |
| D. caricis | 35–90 × 6–7 | (55–)65–85(–100) × 15–16(–17) | Obclavate, brown, septa with central pore, basal cell pale brown, 5–10-distoseptate | Terrestrial | Leaves of Carex sp., Thailand | [26] |
| D. chinensis | 16.5–44 × 5.5–9 | 81–283 × 10–19 | Obclavate or lanceolate, rostrate, olivaceous to dark brown, up to 40-distoseptate | Freshwater | Unidentified submerged wood, China | [12] |
| D. clematidis | 22–40 × 4–10 | 120–210 × 12–20 | Oblong, obclavate, cylindrical or rostrate, brown with green tinge, lighter towards the apex, 28–35-distoseptate | Terrestrial | Dried stem of Clematis sikkimensis, Thailand | [41] |
| D. crassispora | 14–27 × 6–10 | 95–197(–214) × 13–24 | Obclavate, rostrate, brown with a green tinge, 15–36(–41)-distoseptate | Freshwater | Unidentified submerged wood, China | [21] |
| D. curvularia | 11–28 × 5–9 | (60–)100–200(–314) × 12–19 | Obclavate, rostrate, brown with a green tinge, (9–)16–48(–59)-distoseptate | Freshwater | Unidentified submerged wood, China | [21] |
| D. cylindricospora | 105–157 × 6.5–8.5 | 136.5–278 × 8.5–11 | Cylindrical to elongated, greenish-brown to dark brown, hyaline at the apex, 20–65-distoseptate | Freshwater | Unidentified submerged wood, China | [19] |
| D. dehongensis | 45–80 × 4–5 | 17–30 × 7.5–10 | Obpyriform to obclavate, broad cylindrical or irregular, olivaceous, 3–5-distoseptate | Freshwater | Unidentified submerged wood, China | [3] |
| D. effusa | 72–171 × 5–6.5 | 35.5–113 × 7–12.5 | Obclavate, rostrate, olivaceous-brown to dark brown, sometimes slightly paler at the apex, 4–9-distoseptate | Freshwater | Unidentified submerged wood, China | [11] |
| D. euseptata | 19–28 × 4–5 | 37–54 × 8–9 | Obpyriform to obclavate, olivaceous, becoming paler towards the apex, 4–7-euseptate | Freshwater | Unidentified submerged wood, China | [16] |
| D. fasciculata | 12–16 × 5–6 | 46–200 × 10–16.5 | Subcylindrical to obclavate, olivaceous when young, dark brown when mature, 10–40-distoseptate | Freshwater | Unidentified submerged wood, Thailand | [7] |
| D. fluminicola | 21–33 × 5.5–6.5 | 125–250 × 13–15 | Oblong, obclavate, cylindrical or rostrate, brown with green tinge, paler towards the apex, 17–34-distoseptate | Freshwater | Unidentified submerged wood, China | [1] |
| D. fusiformis | 40–110 × 3.5–5.8 | 35–52 × 13.5–22 | Ellipsoidal to fusiform, dark olivaceous-brown to dark brown, pale brown at both ends, 6–8-distoseptate | Freshwater | Unidentified submerged wood, China | [11] |
| D. guizhouensis | 21–50 × 4–9 | 90–273 × 15–21 | Obclavate, brown to dark brown, usually paler towards apex, 10–38-distoseptate | Terrestrial | Unidentified decaying wood, China | [12] |
| D. guttulata | 55–90(–145) × 3.5–5.5 | 75–130(–165) × 7–11 | Obclavate or lanceolate, rostrate, mid- to dark brown or olivaceous, 11–14(–20)-euseptate | Freshwater | Unidentified submerged wood, Thailand | [2] |
| D. hydei | 87–145 × 3–7 | 32–58 × 10–15 | Obpyriform to fusiform, olivaceous to brown, with a hyaline, globose, gelatinous sheath around tip, 7–9-distoseptate | Terrestrial | Decaying bamboo culms, Thailand | [5] |
| D. lancangjiangensis | 144–204 × 5–6 | 64–84 × 9–10 | Narrowly obclavate or obspathulate, brown to dark brown, becoming paler or hyaline towards apex, 3–10-euseptate | Freshwater | Unidentified submerged wood, China | [17] |
| D. leonensis | 110–130 × 6–8 | 50–75 × 15–18 | Obclavate, fusiform or ellipsoidal, rostrate, medium brown, becoming pale brown towards the apex, 8–10-distoseptate | Terrestrial | Dead culms of grasses or dead branches, China | [21,45] |
| D. lignicola | 84–124 × 4–5 | 60–108 × 7–9 | Obclavate, solitary or catenate, brown, 5–9-euseptate | Freshwater | Unidentified submerged wood, Thailand | [22] |
| D. longispora | 17–37 × 6–10 | 189–297 × 16–23 | Obclavate, elongated, brown to yellowish-brown, 31–56-distoseptate | Freshwater | Unidentified submerged wood, China | [14] |
| D. martinii | 50–110 × 3.5–4.5 | 15–20 × 11–16 | Transversal ellipsoid, oblate or subglobose, muriform, pale brown to brown | Terrestrial | Unidentified dead branches, China | [8] |
| D. meilingensis | 69–192 × 4–7 | 32–64.5 × (7–)9–12.5 | Obclavate, mostly bright brown when mature, 5–7-distoseptate | Freshwater | Decaying bamboo culms, China | [6] |
| D. mengsongensis | 17–54 × 4.5–7 | 86–200 × 6–13 | Obclavate, brown to dark brown, sometimes with percurrent proliferation and forming another conidium from the conidial apex, 15–31-distoseptate | Terrestrial | Unidentified dead branches, China | This study |
| D. multiseptata | 23–65 × 4.5–8.5 | 95–290 × 11–20 | Obclavate, rostrate, dark olivaceous-green, multi-distoseptate | Freshwater | Unidentified submerged wood, Thailand | [42] |
| D. nabanheensis | 29–42 × 8–10 | 102–214.5 × (7–)11–14.5 | Obclavate, brown to dark brown, 18–31-distoseptate | Terrestrial | Unidentified dead branches, China | This study |
| D. neorostrata | 93–117 × 5.5–6.5 | 109–147 × 13–15 | Obclavate, rostrate, dark olivaceous to mid- or dark brown, pale brown towards apex, multi-distoseptate | Freshwater | Unidentified submerged wood, Thailand | [22] |
| D. nonrostrata | 105–160 × 4.5–7 | 22–51 × 8–14 | Oblong, obclavate or narrowly obpyriform, mostly non-rostrate, rarely rostrate, pale-olivaceous or pale-brown, 4–10-distoseptate | Freshwater | Unidentified submerged wood, China | [21] |
| D. obclavata | 117.5–162.5 × 5–7 | 46–66 × 9–11 | Obclavate, olivaceous to pale or dark brown, 9-11-distoseptate | Freshwater | Unidentified submerged wood, Thailand | [22] |
| D. obpyriformis | 97–119 × 5–7 | 53–71 × 12–16 | Obpyriform, olivaceous to pale or dark brown, 9–11-distoseptate | Freshwater | Unidentified submerged wood, China | [9] |
| D. pachyconidia | 11–27 × 4–9 | 42–136 × 14–22 | Obclavate, lanceolate, rostrate or not, pale-brown with a green tinge, 8–21-distoseptate | Freshwater | Unidentified submerged wood, China | [21] |
| D. palmarum | 90–165 × 4–7 | 35–180 × 7–11 | Oblong, obclavate, cylindrical or rostrate, greenish-black to brown, paler towards the apex, 7–27-distoseptate | Terrestrial | Rachis of Cocos nucifera, Thailand | [3] |
| D. phangngaensis | 18–30(–40) × 4.3–6.5 | 165–350 × 14–19 | Obclavate, rostrate, dark olivaceous to mid- or dark brown, multi-distoseptate | Freshwater | Unidentified submerged wood, Thailand | [2] |
| D. rayongensis | 75–125 × 3.5–5.5 | (36–)60–106(–120) × 9–14.5 | Obclavate or obspathulate, rostrate, pale brown or pale olivaceous, becoming paler or hyaline towards the apex, sometimes with percurrent proliferation and forming another conidium from the conidial apex, mostly 9–13-euseptate, rarely 14–15-septate | Freshwater | Unidentified submerged wood, Thailand | [28] |
| D. rostrata | 82–126 × 5–7 | 115–155 × 9–11 | Obclavate or lanceolate, rostrate, olivaceous to pale brown, (15–)18–23-distoseptate | Freshwater | Unidentified submerged wood, China | [9] |
| D. saprophytica | 50–140 × 3.2–4.2 | 14.5–30 × 4.5–7.5 | Subcylindrical to obclavate, olivaceous to brown, solitary or occasionally catenate, 2–6-distoseptate | Freshwater | Unidentified submerged wood, Thailand | [7] |
| D. septata | 23–86 × 3–7 | 22–179 × 10–16 | Obclavate, rostrate, pale brown or dark brown, olivaceous-green, usually paler towards apex, verrucose, up to 25-distoseptate | Freshwater | Unidentified submerged wood, China | [20] |
| D. sinensis | 23.5–56.5 × 3.5–7 | 40–107(–137) × 10–12 | Obclavate, brown to dark brown, apical cell paler, 10–25-distoseptate | Terrestrial | Unidentified dead branches, China | This study |
| D. songkhlaensis | 70–90 × 4–5.5 | 44–125 × 9–14.5 | Obclavate, olivaceous to brown, 9–16-distoseptate | Freshwater | Unidentified submerged wood, Thailand | [7] |
| D. suoluoensis | 80–250 × 4.5–5.8 | (65–)80–125(–145) × 8–13 | Narrowly obclavate or obspathulate, yellowish-brown or dark olivaceous, becoming paler or hyaline towards the apex, verrucose, sometimes with percurrent proliferation, which forms another conidium from the conidial apex, 8–10-euseptate | Freshwater | Unidentified submerged wood, China | [2] |
| D. tectonae | Up to 40 × 4–6 | (90–)130–140(–170) × (11–)13–14(–16) | Cylindric-obclavate, dark reddish-brown, slightly paler towards the apex, verruculose, 20–28-distoseptate | Terrestrial | Dead twig of Tectona grandis, Thailand | [42] |
| D. tectonigena | Up to 110 × 5–11 | (83–)148–225(–360) × (10–)11–12(–13) | Cylindric-obclavate, dark reddish-brown and slightly paler towards the apex, verruculose, 20–46-distoseptate | Terrestrial | Dead twig of Tectona grandis, Thailand | [42] |
| D. thailandica | 15–26 × 3–6 | 130–230 × 13.5–17 | Oblong, obclavate, cylindrical or rostrate, reddish-brown to brown, pale brown towards the apex, 35–52-distoseptate | Terrestrial | Dead leaves of Pandanus sp., Thailand | [10] |
| D. thysanolaenae | 30–80 × 3.5–5.5 | 21.5–80 × 6.5–12.8 | Narrow and elongated obclavate, light to dark brown, paler at the apex, 8–14-distoseptate | Terrestrial | Dead culms of Thysanolaena maxima, China | [13] |
| D. tropica | 60–151 × 3.5–7 | 39–75 × 7.5–10.5 | Obclavate, rostrate, olivaceous-brown or dark brown, verrucose, 5–7 distoseptate | Terrestrial | Unidentified dead wood, China | [20] |
| D. verrucosa | 92–250 × 4.7–6.3 | 41–63 × 8.8–12.6 | Obclavate, rostrate, olivaceous-brown, becoming paler at the apex, verrucose, 6–8-euseptate | Freshwater | Unidentified submerged wood, China | [11] |
| D. wuzhishanensis | 16–56 × 5–7 | 76–143 × 11–17 | Obclavate, rostrate, pale brown or dark brown, olivaceous-green and yellow, usually paler towards apex, verrucose, up to 22-distoseptate | Freshwater | Unidentified submerged wood, China | [20] |
| D. xishuangbannaensis | 12–17 × 2–5 | 160–305 × 8–15 | Cylindrical-obclavate, green brown to brown, up to 40-distoseptate | Terrestrial | Decaying leaves of Pandanus utilis, China | [10] |
| D. yongxiuensis | 112–253 × 4–9 | 46–74(–86) × 10–13(–16) | Obclavate or obspathulate, olivaceous to yellowish-brown or brown, becoming paler or hyaline towards the apex, 6–9-euseptate | Freshwater | Decaying bamboo culms, China | [6] |
| D. yunjushanensis | 100–175 × 5.5–10 | 39–67.5(–77) × (7–)9.5–13.5(–16.5) | Obpyriform or obclavate, olivaceous when young, dark brown when mature, sometimes with the percurrent proliferation which forms another conidium from the conidial apex, 7–13-distoseptate | Freshwater | Decaying bamboo culms, China | [6] |
| D. yunnanensis | 131–175 × 6–7 | 58–108 × 8–10 | Obclavate, rostrate, mid-olivaceous to brown, becoming paler towards the apex, 6–10-euseptate | Freshwater | Unidentified submerged wood, China | [16] |
| Species | Asci (μm) | Ascospores | Habitat | Host/Locality | References | ||
|---|---|---|---|---|---|---|---|
| Size (μm) | Septation | Characteristics | |||||
| Distoseptispora hyalina | 145–190 × 8–11 | (19.5–)23–26(–28.5) × 4.5–7 | 0–3 | Fusiform, hyaline, guttulate, and with a mucilaginous sheath | Freshwater | Unidentified submerged wood, Thailand | [11] |
| D. licualae | 120–210 × 8–15 | 20–30 × 5–10 | 0 | Inequilateral to fusiform, hyaline, guttulate, and with a thick mucilaginous sheath | Terrestrial | Dead leaves of Licuala glabra, Thailand | [43] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Hu, Y.; Luo, X.; Xu, Z.; Castañeda-Ruíz, R.F.; Xia, J.; Zhang, X.; Zhang, L.; Cui, R.; Ma, J. Morphological and Phylogenetic Analyses Reveal Three New Species of Distoseptispora (Distoseptisporaceae, Distoseptisporales) from Yunnan, China. J. Fungi 2023, 9, 470. https://doi.org/10.3390/jof9040470
Liu J, Hu Y, Luo X, Xu Z, Castañeda-Ruíz RF, Xia J, Zhang X, Zhang L, Cui R, Ma J. Morphological and Phylogenetic Analyses Reveal Three New Species of Distoseptispora (Distoseptisporaceae, Distoseptisporales) from Yunnan, China. Journal of Fungi. 2023; 9(4):470. https://doi.org/10.3390/jof9040470
Chicago/Turabian StyleLiu, Jingwen, Yafen Hu, Xingxing Luo, Zhaohuan Xu, Rafael F. Castañeda-Ruíz, Jiwen Xia, Xiuguo Zhang, Lianhu Zhang, Ruqiang Cui, and Jian Ma. 2023. "Morphological and Phylogenetic Analyses Reveal Three New Species of Distoseptispora (Distoseptisporaceae, Distoseptisporales) from Yunnan, China" Journal of Fungi 9, no. 4: 470. https://doi.org/10.3390/jof9040470