
Genetic Variation of Puccinia triticina Populations in Iran from 2010 to 2017 as Revealed by SSR and ISSR Markers

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Puccinia triticina Isolates
2.2. DNA Extraction
2.3. ISSR Markers
2.4. SSR Markers
2.5. Data Analysis
3. Results
SSR and ISSR Polymorphisms
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roelfs, A.P.; Singh, R.P.; Saari, E.E. Rust Diseases of Wheat: Concepts and Methods of Disease Management; CIMMYT: Mexico City, Mexico, 1992; p. 81. [Google Scholar]
- Murray, T.; Parry, D.W.; Cattlin, N.D. Diseases of Small Grain Cereal Crops; Manson Publishing: London, UK, 1998; p. 142. [Google Scholar] [CrossRef]
- Bolton, M.D.; Kolmer, J.A.; Garvin, D.F. Wheat leaf rust caused by Puccinia triticina. Mol. Plant Pathol. 2008, 9, 563–575. [Google Scholar] [CrossRef]
- Kolmer, J.A. Tracking wheat rust on a continental scale. Curr. Opin. Plant Biol. 2005, 8, 441–449. [Google Scholar] [CrossRef]
- Kolmer, J.A.; Ordoñez, M.E.; Manisterski, J.; Anikster, Y. Genetic differentiation of Puccinia triticina populations in the middle east and genetic similarity with populations in Central Asia. Phytopathology 2011, 101, 870–877. [Google Scholar] [CrossRef] [Green Version]
- Kolmer, J. Leaf rust of wheat: Pathogen biology, variation and host resistance. Forests 2013, 4, 70–84. [Google Scholar] [CrossRef] [Green Version]
- Kolmer, J.A.; Ordoñez, M.E. Genetic differentiation of Puccinia triticina populations in Central Asia and the Caucasus. Phytopathology 2007, 97, 1141–1149. [Google Scholar] [CrossRef] [Green Version]
- Park, R.F.; Burdon, J.J.; Jahoor, A. Evidence for somatic hybridization in nature of Puccinia recondita f. sp. tritici. Mycol. Res. 1999, 103, 715–732. [Google Scholar] [CrossRef]
- Goyeau, H.; Park, R.; Schaeffer, B.; Lannou, C. Distribution of pathotypes with regard to host cultivars in french wheat leaf rust populations. Phytopathology 2006, 96, 264–273. [Google Scholar] [CrossRef] [Green Version]
- Nemati, Z.; Mostowfizadeh-Ghalamfarsa, R.; Dadkhodaie, A.; Mehrabi, R.; Steffenson, B.J. Virulence of leaf rust physiological races in Iran from 2010 to 2017. Plant Dis. 2020, 104, 363–372. [Google Scholar] [CrossRef]
- Kolmer, J.A.; Liu, J.Q.; Sies, M. Virulence and molecular polymorphism in Puccinia recondita f. sp. tritici in Canada. Phyto-Pathol. 1995, 85, 276–285. [Google Scholar] [CrossRef]
- Mantovani, P.; Maccaferri, M.; Tuberosa, R.; Kolmer, J. Virulence phenotypes and molecular genotypes in collections of Puccinia triticina from Italy. Plant Dis. 2010, 94, 420–424. [Google Scholar] [CrossRef]
- Liu, T.G.; Chen, W.Q. Race and virulence dynamics of Puccinia triticina in China during 2000–2006. Plant Dis. 2012, 96, 1601–1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolmer, J.A.; Kabdulova, M.G.; Mustafina, M.A.; Zhemchuzhina, N.S.; Dubovoy, V. Russian populations of Puccinia triticina in distant regions are not differentiated for virulence and molecular genotype. Plant Pathol. 2014, 64, 328–336. [Google Scholar] [CrossRef]
- Kolmer, J.A. Collections of Puccinia triticina in different provinces of China are highly related for virulence and molecular genotype. Phytopathology 2015, 105, 700–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gultyaeva, E.I.; Shaydayuk, E.L.; Kazartsev, I.A.; Kosman, E. Race characterization and molecular genotyping of Puccinia triticina populations from durum wheat in Russia. Plant Dis. 2021, 105, 1495–1504. [Google Scholar] [CrossRef]
- Khan, R.R.; Bariana, H.S.; Dholakia, B.B.; Naik, S.V.; Lagu, M.D.; Rathjen, A.J.; Bhavani, S.; Gupta, V.S. Molecular mapping of stem and leaf rust resistance in wheat. Theor. Appl. Genet. 2005, 111, 846–850. [Google Scholar] [CrossRef]
- Ordoñez, M.E.; Kolmer, J.A. Simple sequence repeat diversity of a world-wide collection of Puccinia triticina from durum wheat. Phytopathology 2007, 97, 574–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabo, L.J.; Kolmer, J.A. Development of simple sequence repeat markers for the plant pathogenic rust fungus Puccinia triticina. Mol. Ecol. Notes 2007, 7, 708–710. [Google Scholar] [CrossRef]
- Goyeau, H.; Berder, J.; Czerepak, C.; Gautier, A.; Lanen, C.; Lannou, C. Low diversity and fast evolution in the population of Puccinia triticina causing durum wheat leaf rust in France from 1999 to 2009, as revealed by an adapted differential set. Plant Pathol. 2012, 61, 761–772. [Google Scholar] [CrossRef]
- Dadrezaie, S.T.; Lababidi, S.; Nazari, K.; Goltapeh, E.M.; Afshari, F.; Alo, F.; Shams-Bakhsh, M.; Safaie, N. Molecular genetic diversity in Iranian populations of Puccinia triticina, the causal agent of wheat leaf rust. Am. J. Plant Sci. 2013, 04, 1375–1386. [Google Scholar] [CrossRef] [Green Version]
- Kosman, E. Measuring diversity: From individuals to populations. Eur. J. Plant Pathol. 2014, 138, 467–486. [Google Scholar] [CrossRef]
- Aoun, M.; Kolmer, J.A.; Breiland, M.; Richards, J.; Brueggeman, R.S.; Szabo, L.J.; Acevedo, M. Genotyping-by-sequencing for the study of genetic diversity in Puccinia triticina. Plant Dis. 2020, 104, 752–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Liu, T.; Liu, B.; Gao, L.; Chen, W. Population genetic structures of Puccinia triticina in five provinces of China. Eur. J. Plant Pathol. 2020, 156, 1135–1145. [Google Scholar] [CrossRef] [Green Version]
- Ordoñez, M.E.; Kolmer, J.A. Differentiation of molecular genotypes and virulence phenotypes of Puccinia triticina from common wheat in North America. Phytopathology 2009, 99, 750–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosman, E.; Jokela, J. Dissimilarity of individual microsatellite profiles under different mutation models: Empirical approach. Ecol. Evol. 2019, 9, 4038–4054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosman, E.; Leonard, K.J. Similarity coefficients for molecular markers in studies of genetic relationships between individuals for haploid, diploid, and polyploid species. Mol. Ecol. 2005, 14, 415–424. [Google Scholar] [CrossRef]
- Niazmand, A.R.; Afshari, F.; Abbasi, M.; Rezaee, S. Study on pathotypes diversity and virulence factors of Puccinia triticina Eriksson, the causal agent of wheat brown rust in Iran. Iran. J. Plant Pathol. 2010, 46, 187–202. [Google Scholar]
- Nemati, Z.; Pourkhaloee, A.; Mostowfizadeh-Ghalamfarsa, R.; Khosh-Khui, M.; Jafari, M. A report of rust species on different hosts from Fars Province. In Proceedings of the 3rd Iranian Mycological Congress, Sanandaj, Iran, 26–28 August 2017; p. 81. [Google Scholar]
- Kolmer, J.A.; Long, D.L.; Hughes, M.E. Physiologic specialization of Puccinia triticina on wheat in the United States. Plant Dis. 2009, 93, 538–544. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.Q.; Kolmer, J.A. Molecular and virulence diversity and linkage disequilibria in asexual and sexual populations of the wheat leaf rust fungus, Puccinia recondita. Genome 1998, 41, 832–840. [Google Scholar] [CrossRef]
- Spackman, M.E.; Ogbonnaya, F.C.; Brown, J.S. Hypervariable RAPD, ISSR and SSR markers generate robust taxonomic groups among Puccinia striiformis formae speciales of importance to Australian agriculture. Australas. Plant Pathol. 2010, 39, 226–233. [Google Scholar] [CrossRef]
- Powell, W.; Morgante, M.; Andre, C.; Hanafey, M.; Vogel, J.; Tingey, S.; Rafalski, A. The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis. Mol. Breed. 1996, 2, 225–238. [Google Scholar] [CrossRef]
- de Souza, S.G.H.; Carpentieri-Pípolo, V.; Ruas, C.D.F.; Carvalho, V.D.P.; Ruas, P.M.; Gerage, A.C. Comparative analysis of genetic diversity among the maize inbred lines (Zea mays L.) obtained by RAPD and SSR markers. Braz. Arch. Biol. Technol. 2008, 51, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Vieira, E.S.N.; Pinho, V.D.R.V.; Carvalho, M.G.G.; Esselink, D.G.; Vosman, B. Development of microsatellite markers for identifying Brazilian Coffea arabica varieties. Genet. Mol. Biol. 2010, 33, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Arbaoui, M.; Kraic, J.; Huszar, J. Genetic variation of Sclerotinia sclerotiorum isolates from different conditions. J. Agric. Sci. 2008, 54, 36–39. [Google Scholar]
- Sharma, M.; Gupta, S.K.; Sharma, T.R. Characterization of variability in Rhizoctonia solani by using morphological and molecular markers. J. Phytopathol. 2005, 153, 449–456. [Google Scholar] [CrossRef]
- Xue, S.; Xuecheng, Z.; Yunxiang, M.; Zhenghong, S.; Song, Q. Identification of phase and sex-related ISSR markers of red alga Gracilaria lemaneiformis. J. Ocean Univ. China 2006, 5, 82–84. [Google Scholar] [CrossRef]
- Vasseur, V.; Rey, P.; Bellanger, E.; Brygoo, Y.; Tirilly, Y. Molecular characterization of Pythium group F isolates by ribosomal-and intermicrosatellite-DNA regions analysis. Eur. J. Plant Pathol. 2005, 112, 301–310. [Google Scholar] [CrossRef]
- Geleta, M.; Bryngelsson, T. Inter simple sequence repeat (ISSR) based analysis of genetic diversity of Lobelia rhynchopetalum (Campanulaceae). Hereditas 2009, 146, 122–130. [Google Scholar] [CrossRef]
- Gupta, M.; Chyi, Y.-S.; Romero-Severson, J.; Owen, J.L. Amplification of DNA markers from evolutionarily diverse genomes using single primers of simple-sequence repeats. Theor. Appl. Genet. 1994, 89, 998–1006. [Google Scholar] [CrossRef]
- Dinolfo, M.I.; Stenglein, S.A.; Moreno, M.V.; Nicholson, P.; Jennings, P.; Salerno, G.L. ISSR markers detect high genetic varia-tion among Fusarium poae isolates from Argentina and England. Eur. J. Plant Pathol. 2010, 127, 483–491. [Google Scholar] [CrossRef]
- Bassam, B.J.; Gresshoff, P.M. Silver staining of DNA in polyacrylamide gels. Nat. Protoc. 2007, 2, 2649–2654. [Google Scholar] [CrossRef]
- Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res. 1967, 27, 209–220. [Google Scholar] [PubMed]
- Liu, K.; Muse, S.V. PowerMarker: Integrated analysis environment for genetic marker data. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multi locus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.O.D.; Smouse, P.E. Genalex 6: Genetic analysis in excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using multilocus genotype data: Linked loci and correlated allele frequencies. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Kolmer, J.A.; Hanzalova, A.; Goyeau, H.; Bayles, R.; Morgounov, A. Genetic differentiation of the wheat leaf rust fungus Puccinia triticina in Europe. Plant Pathol. 2012, 62, 21–31. [Google Scholar] [CrossRef]
- Gultyaeva, E.I.; Aristova, M.K.; Shaidayuk, E.L.; Mironenko, N.V.; Kazartsev, I.A.; Akhmetova, A.; Kosman, E. Genetic differentiation of Puccinia triticina Erikss in Russia. Russ. J. Genet. 2017, 53, 998–1005. [Google Scholar] [CrossRef]
- Kolmer, J.A.; Mirza, J.I.; Imtiaz, M.; Shah, S.J.A. Genetic differentiation of the wheat leaf rust fungus Puccinia triticina in Pakistan and genetic relationship to other worldwide populations. Phytopathology 2017, 107, 786–790. [Google Scholar] [CrossRef] [Green Version]
- Czajowski, G.; Kosman, E.; Słowacki, P.; Park, R.F.; Czembor, P. Pathogenic and genetic diversity of Puccinia triticina from triticale in Poland between 2012 and 2015. Plant Pathol. 2021, 70, 2148–2164. [Google Scholar] [CrossRef]
- Czajowski, G.; Kosman, E.; Slowacki, P.; Park, R.F.; Czembor, P. Assessing SSR markers for utility and informativeness in genetics studies of brown rust fungi on wheat, triticale and rye. Plant Pathol. 2021, 70, 1110–1122. [Google Scholar] [CrossRef]
- Nemati, Z.; Mostowfizadeh-Ghalamfarsa, R.; Dadkhodaie, A.; Mehrabi, R.; Steffenson, B.J. Host range of various leaf rust populations in Iran. Iran. J. Plant Pathol. 2019, 54, 305–316. [Google Scholar]
- Kolmer, J.A.; Herman, A.; Ordoñez, M.E.; German, S.; Morgounov, A.; Pretorius, Z.; Visser, B.; Anikster, Y.; Acevedo, M. Endemic and panglobal genetic groups, and divergence of host-associated forms in worldwide collections of the wheat leaf rust fungus Puccinia triticina as determined by genotyping by sequencing. Heredity 2020, 124, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Quinones, M.A.; Larter, E.N.; Samborski, D.J. The inheritance of resistance to Puccinia recondita in hexaploid triticale. Can. J. Genet. Cytol. 1972, 14, 495–505. [Google Scholar] [CrossRef]
- Nazareno, E.; Feng, l.; Madeleine, S.; Park, R.F.; Kianian, S.F.; Figueroa, M. Puccinia coronata f.sp. avenae: A threat to global oat production. Mol. Plant Pathol. 2017, 19, 1047–1060. [Google Scholar] [CrossRef] [Green Version]
- Mehnaz, M.; Dracatos, P.M.; Park, R.F.; Singh, D. Mining middle eastern and central Asian barley germplasm to understand diversity for resistance to Puccinia hordei, causal agent of leaf rust. Agronomy 2021, 11, 2146. [Google Scholar] [CrossRef]
- Wahl, I.; Anikster, Y.; Manisterski, J.; Segal, A. Evolution at the center of origin. In The Cereal Rusts; Bushnell, I.W.R., Roelfs, A.P., Eds.; Academic Press: New York, NY, USA, 1984; pp. 39–77. [Google Scholar] [CrossRef]
- Wiese, M.V. Compendium of Wheat Diseases, 2nd ed.; APS Press, The American Phytopathological Society: St. Paul, MN, USA, 1987; p. 41. [Google Scholar]
- Neu, C.; Keller, B.; Feuillet, C. Cytological and molecular analysis of the Hordeum vulgare-Puccinia triticina nonhost interaction. Mol. Plant-Microbe Interact. 2003, 16, 626–633. [Google Scholar] [CrossRef] [Green Version]
- Niks, R.E.; Marcel, T.C. Nonhost and basal resistance: How to explain specificity? New Phytol. 2009, 182, 817–828. [Google Scholar] [CrossRef]
- Bettgenhaeuser, J.; Gilbert, B.; Ayliffe, M.; Moscou, M.J. Nonhost resistance to rust pathogens–a continuation of continua. Front Plant Sci. 2014, 5, 664. [Google Scholar] [CrossRef] [Green Version]
- Porras-Hurtado, L.; Ruiz, Y.; Santos, C.; Phillips, C.; Carracedo, Á.; Lareu, M.V. An overview of structure: Applications, parameter settings, and supporting software. Front. Genet. 2013, 4, 98. [Google Scholar] [CrossRef] [Green Version]
- D’Oliveira, B.D.; Samborski, D.J. Aecial stage of Puccinia recondita on Ranunculaceae and Boraginaceae in Portugal. In Proceedings of the First European Brown Rust Conference; Macer, R.C., Wolfe, M.S., Eds.; Plant Breeding Institute: Cambridge, UK, 1966; pp. 133–150. [Google Scholar]
- Pakravan, M.; Alipanah, H.; Soleimani, N. A revision of the genus Thalictrum L. in Iran. Iran. J. Bot. 2014, 20, 170–178. [Google Scholar]


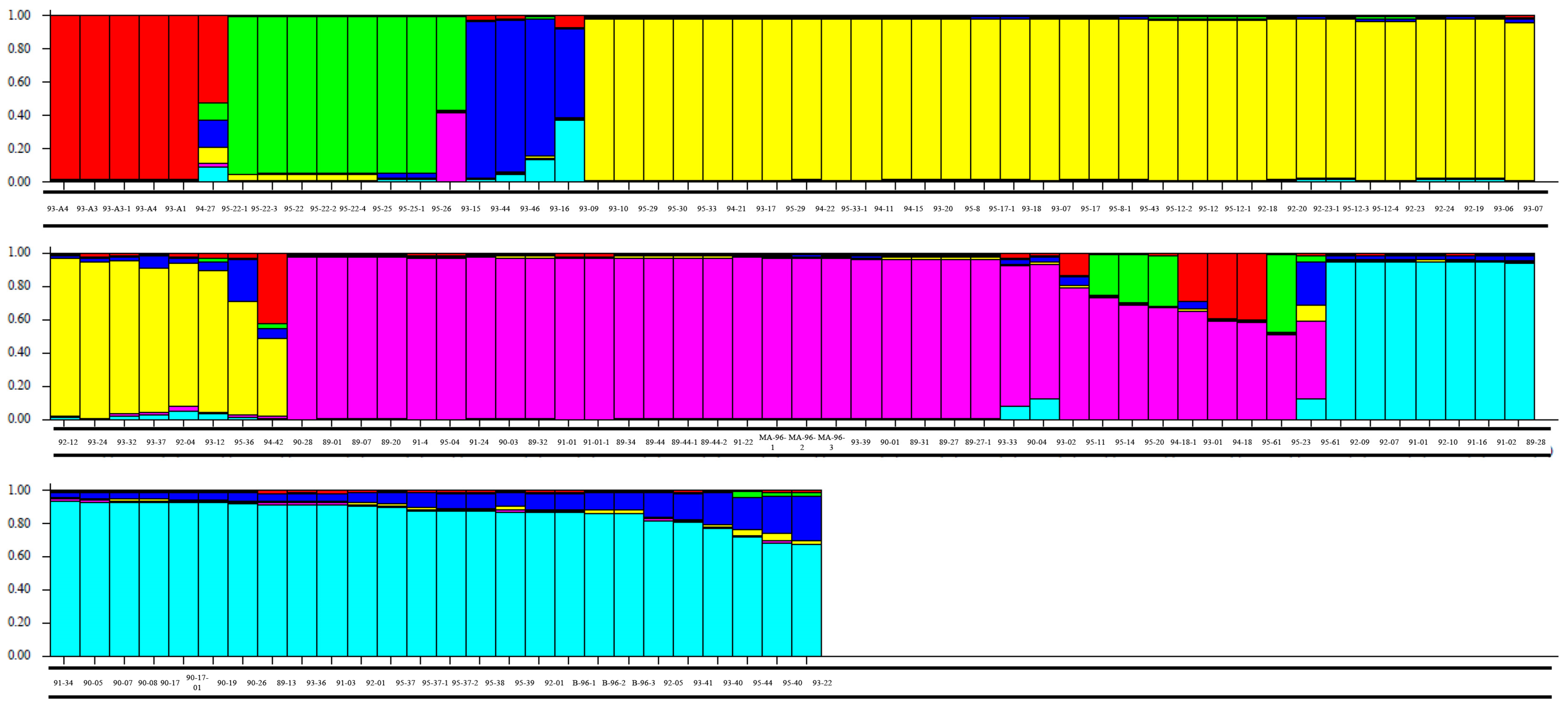{kind=link}
| Primer | Primer Sequence | Reference |
|---|---|---|
| P1 * | 5′-(GACA) 4AT-3′ | [36] |
| P2 * | 5′-(GACC) 4TT-3′ | [36] |
| P5 * | 5′-(ACTG) 3ACG-3′ | [37] |
| P7 | 5′-(TGTC) 5-3′ | [38] |
| P8 | 5′-(CAC) 5-3′ | [39] |
| P9 | 5′-(CCA) 5-3′ | [39] |
| P10 * | 5′-(GACC) 4-3′ | [39] |
| P17 * | 5′-(GGAGA) 3-3′ | [40] |
| M1 * | 5′-(ACTG) 4-3′ | [41] |
| M2 * | 5′-(GACAC) 3-3′ | [41] |
| M6 | 5′-(GCC) 3-3′ | [42] |
| M7 * | 5′-(GAG) 5-3′ | [42] |
| Groups of Isolates | SSR Markers | ISSR Markers |
|---|---|---|
| Wild Barley 2017 | 76.47% | 2.38% |
| Durum Wheat 2016 | 58.82% | 34.92% |
| Barley 2016 | 70.59% | 44.44% |
| Oat 2016 | 58.82% | 0.00% |
| Triticale 2016 | 58.82% | 0.00% |
| Bread Wheat 2016 | 88.24% | 74.60% |
| Bread Wheat 2015 | 76.47% | 50.79% |
| Bread Wheat 2014 | 94.12% | 75.40% |
| Bread Wheat 2013 | 82.35% | 65.08% |
| Bread Wheat 2012 | 76.47% | 65.87% |
| Bread Wheat 2011 | 82.35% | 69.84% |
| Bread Wheat 2010 | 76.47% | 51.59% |
| Mean | 75.00% | 44.58% |
| SE | 3.33% | 8.39% |
| Locus Number | Repeat Size | Missing Data | Min | Max | Number of Homozygotes | Number of Heterozygotes | Proportion of Homozygotes | N. Alleles |
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 0 | 128 | 130 | 0 | 126 | 0 | 2 |
| 2 | 3 | 0 | 360 | 366 | 3 | 123 | 0.024 | 3 |
| 3 | 2 | 0 | 302 | 304 | 51 | 75 | 0.405 | 2 |
| 4 | 3 | 0 | 297 | 303 | 0 | 126 | 0 | 3 |
| 5 | 4 | 0 | 319 | 327 | 35 | 91 | 0.278 | 4 |
| 6 | 5 | 0 | 307 | 317 | 33 | 93 | 0.262 | 5 |
| 7 | 3 | 0 | 393 | 402 | 0 | 126 | 0 | 3 |
| 8 | 2 | 0 | 348 | 380 | 11 | 115 | 0.087 | 2 |
| 9 | 3 | 0 | 242 | 252 | 8 | 118 | 0.063 | 3 |
| 10 | 2 | 0 | 470 | 476 | 118 | 8 | 0.937 | 2 |
| 11 | 2 | 0 | 384 | 388 | 47 | 79 | 0.373 | 2 |
| 12 | 4 | 0 | 265 | 272 | 0 | 126 | 0 | 4 |
| 13 | 3 | 0 | 242 | 250 | 0 | 126 | 0 | 3 |
| 14 | 2 | 0 | 227 | 232 | 1 | 125 | 0.008 | 3 |
| 15 | 3 | 0 | 213 | 217 | 15 | 111 | 0.119 | 3 |
| 16 | 5 | 0 | 214 | 224 | 11 | 115 | 0.087 | 5 |
| 17 | 4 | 0 | 211 | 220 | 0 | 126 | 0 | 4 |
| 18 | 4 | 0 | 345 | 400 | 0 | 126 | 0 | 4 |
| 19 | 4 | 0 | 340 | 347 | 22 | 104 | 0.175 | 3 |
| 20 | 3 | 0 | 335 | 337 | 0 | 126 | 0 | 3 |
| Matrix Correlation | IAM SSR | SMMc SSR | SMMv SSR | DICE ISSR | JACCARD ISSR | MIS MATCH ISSR | DICE Phenotype | JACCARD Phenotype | MIS MATCH Phenotype |
|---|---|---|---|---|---|---|---|---|---|
| IAM SSR | 1 | ||||||||
| SMMc SSR | 0.9187 | 1 | |||||||
| SMMv SSR | 0.92277 | 0.99452 | 1 | ||||||
| DICE ISSR | 0.3328 | 0.49538 | 0.47167 | 1 | |||||
| JACCARD ISSR | 0.34361 | 0.49409 | 0.47025 | 0.99814 | 1 | ||||
| MIS MATCH ISSR | 0.28984 | 0.44524 | 0.43044 | 0.95371 | 0.95843 | 1 | |||
| DICE Phenotype | 0.10095 | 0.26786 | 0.28764 | 0.28271 | 0.28888 | 0.21861 | 1 | ||
| JACCARD Phenotype | 0.10095 | 0.26786 | 0.28764 | 0.28271 | 0.28888 | 0.21861 | 1 | 1 | |
| MIS MATCH Phenotype | 0.05111 | 0.18975 | 0.18438 | 0.19746 | 0.20227 | 0.14982 | 0.91504 | 0.91504 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nemati, Z.; Dadkhodaie, A.; Mostowfizadeh-Ghalamfarsa, R.; Mehrabi, R.; Cacciola, S.O. Genetic Variation of Puccinia triticina Populations in Iran from 2010 to 2017 as Revealed by SSR and ISSR Markers. J. Fungi 2023, 9, 388. https://doi.org/10.3390/jof9030388
Nemati Z, Dadkhodaie A, Mostowfizadeh-Ghalamfarsa R, Mehrabi R, Cacciola SO. Genetic Variation of Puccinia triticina Populations in Iran from 2010 to 2017 as Revealed by SSR and ISSR Markers. Journal of Fungi. 2023; 9(3):388. https://doi.org/10.3390/jof9030388
Chicago/Turabian StyleNemati, Zahra, Ali Dadkhodaie, Reza Mostowfizadeh-Ghalamfarsa, Rahim Mehrabi, and Santa Olga Cacciola. 2023. "Genetic Variation of Puccinia triticina Populations in Iran from 2010 to 2017 as Revealed by SSR and ISSR Markers" Journal of Fungi 9, no. 3: 388. https://doi.org/10.3390/jof9030388