
What Are the Effects of Moso Bamboo Expansion into Japanese Cedar on Arbuscular Mycorrhizal Fungi: Altering the Community Composition Rather than the Diversity
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site Description
2.2. Soil Sampling and the Determination of Chemical Characteristics
2.3. DNA Extraction, PCR Amplification and Molecular Identification of AMF
2.4. Statistical Analyses
3. Results
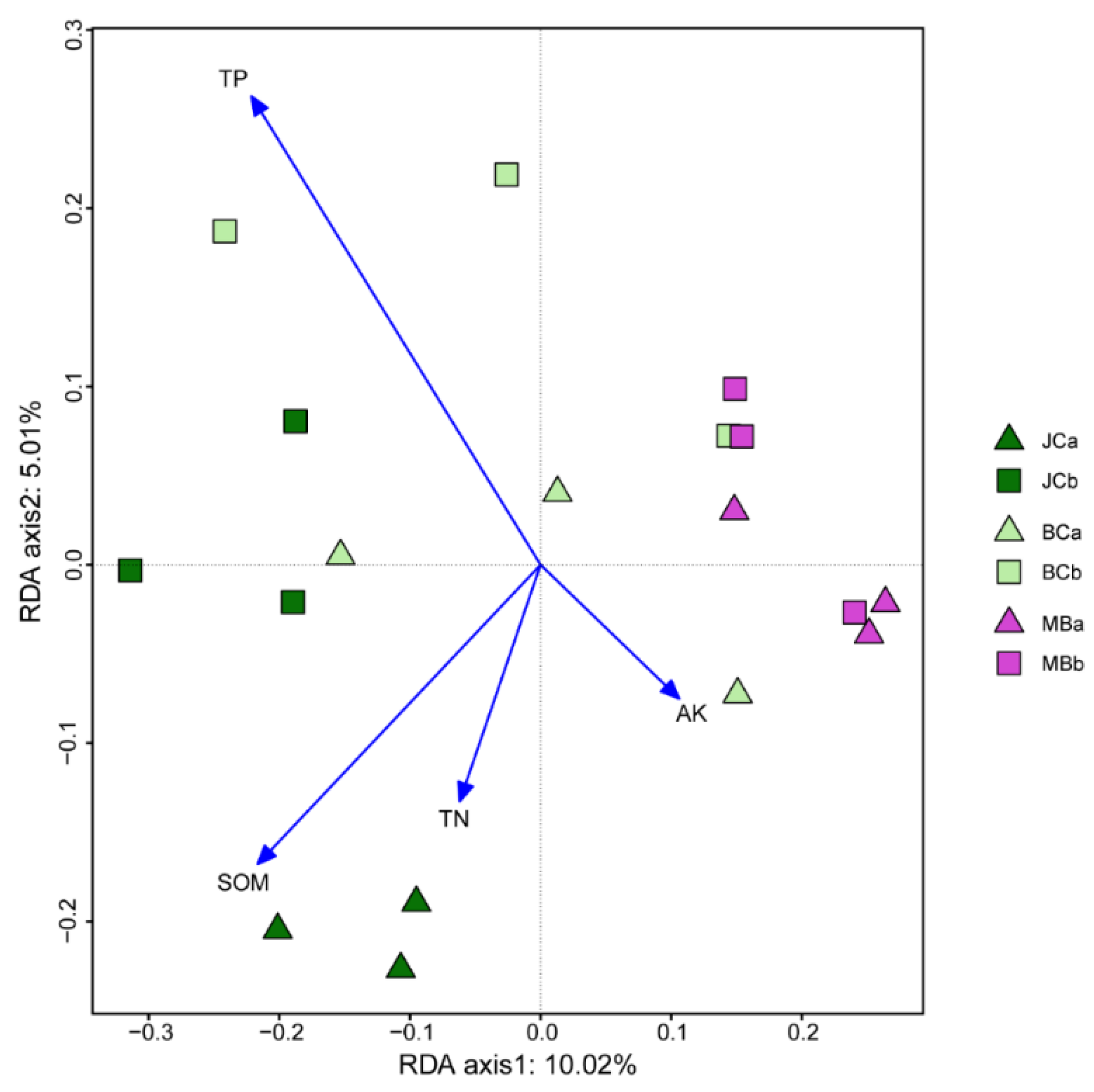
3.1. Chemical Properties of Soil
3.2. Overall Pyrosequencing Information
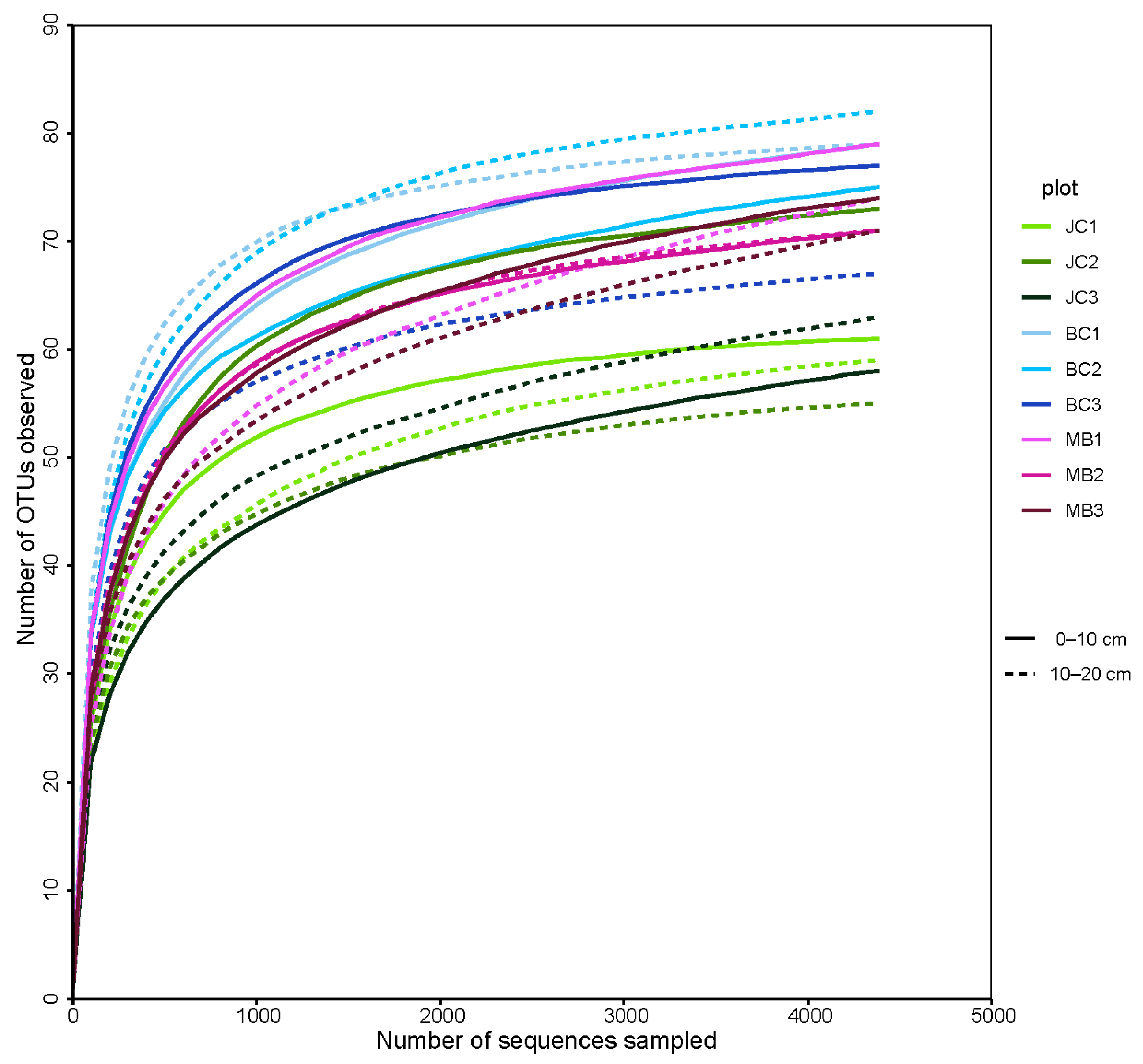
3.3. The Richness and α Diversity of AMF
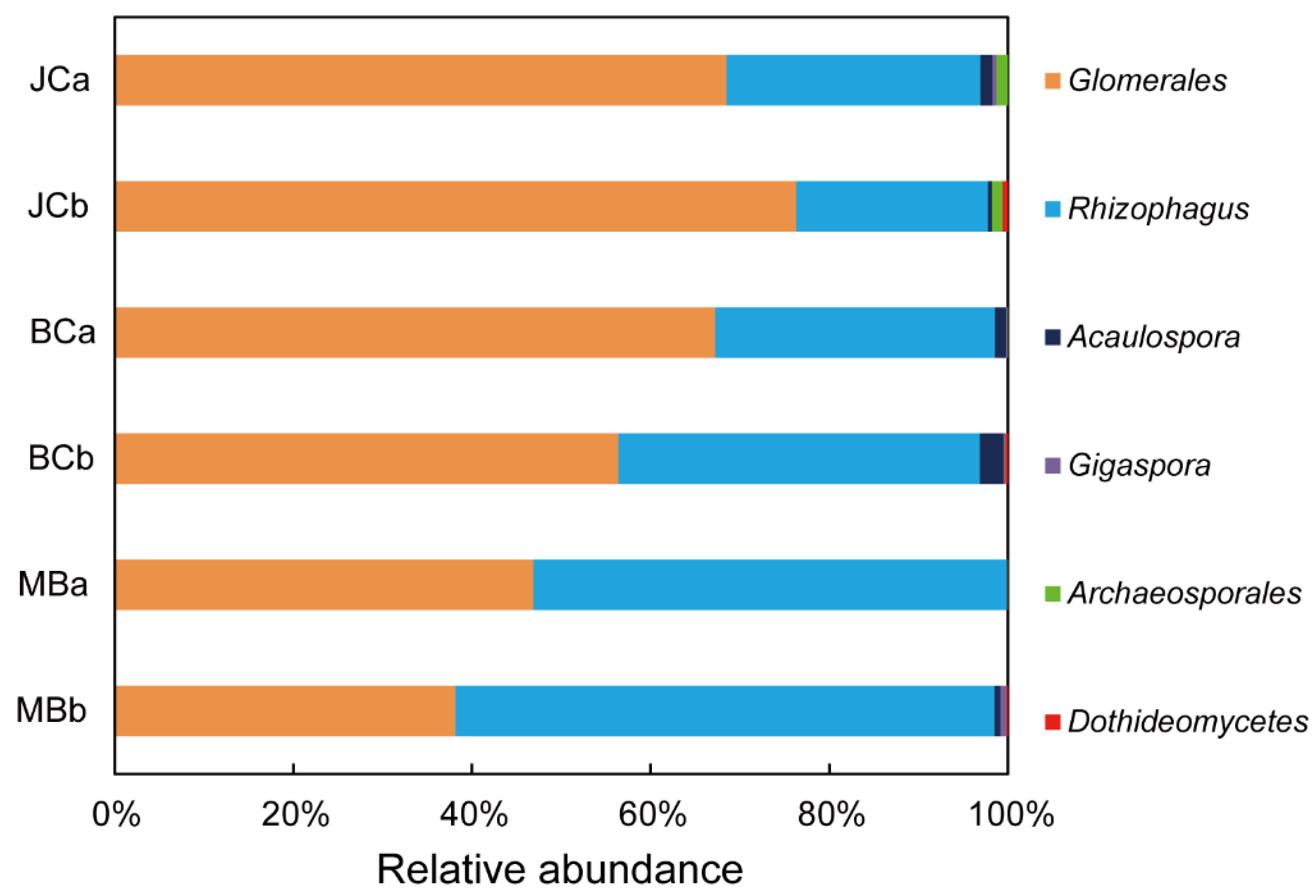
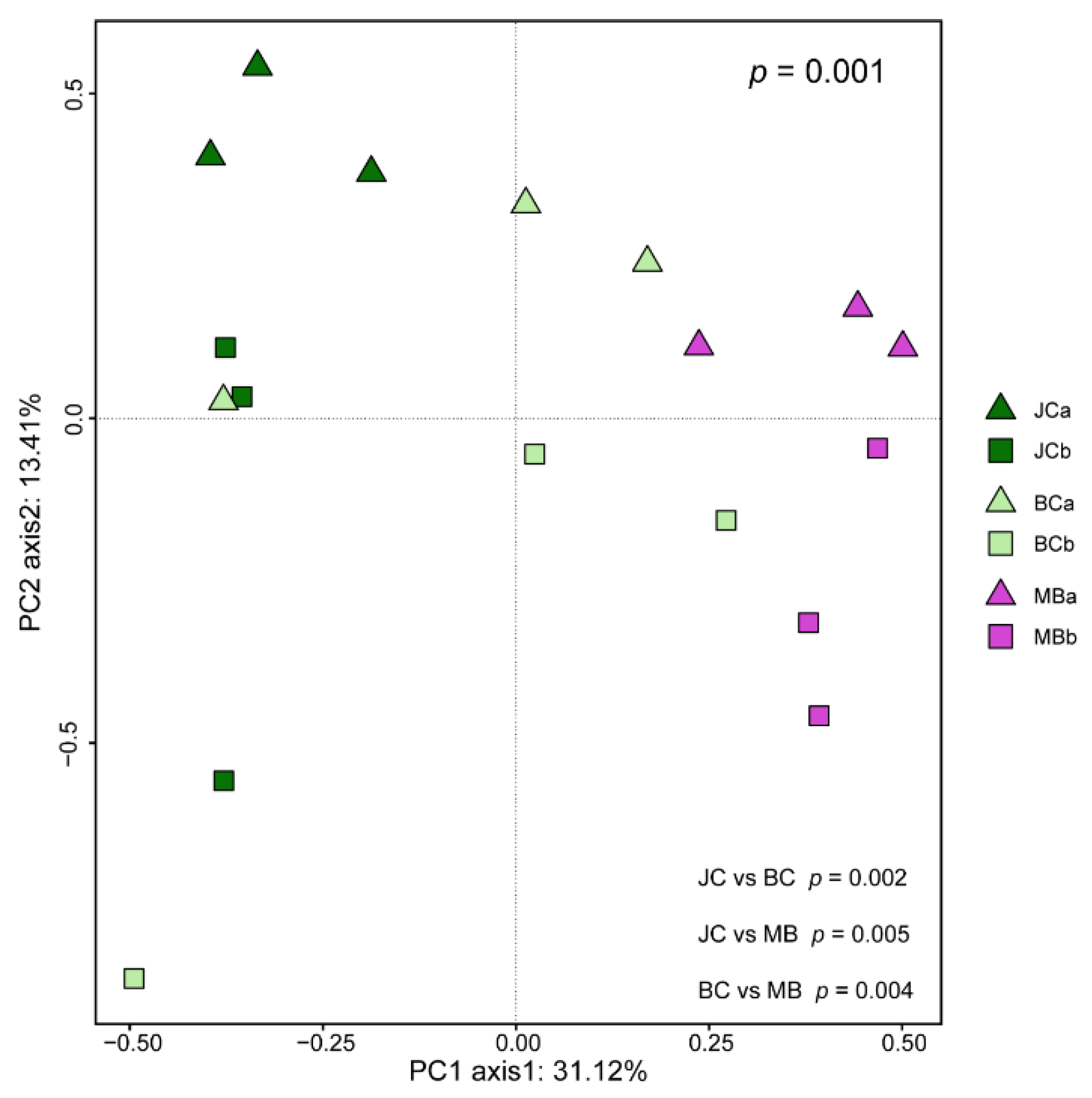
3.4. The Composition of AMF Community
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Heringer, G.; Faria, L.D.B.; Villa, P.M.; Araújo, A.U.; Botan, A.L.M.; Zenni, R.D. Urbanization affects the richness of invasive alien trees but has limited influence on species composition. Urban Ecosyst. 2021, 25, 753–763. [Google Scholar] [CrossRef]
- Powell, K.I.; Chase, J.M.; Knight, T.M. A synthesis of plant invasion effects on biodiversity across spatial scales. Am. J. Bot. 2011, 98, 539–548. [Google Scholar] [CrossRef] [Green Version]
- Song, Q.-N.; Ouyang, M.; Yang, Q.-P.; Lu, H.; Yang, G.-Y.; Chen, F.-S.; Shi, J.-M. Degradation of litter quality and decline of soil nitrogen mineralization after moso bamboo (Phyllostachys pubscens) expansion to neighboring broadleaved forest in subtropical China. Plant Soil 2016, 404, 113–124. [Google Scholar] [CrossRef]
- Xu, Q.-F.; Liang, C.-F.; Chen, J.-H.; Li, Y.-C.; Qin, H.; Fuhrmann, J.J. Rapid bamboo invasion (expansion) and its effects on biodiversity and soil processes+. Glob. Ecol. Conserv. 2020, 21, e00787. [Google Scholar] [CrossRef]
- Ouyang, M.; Yang, C.; Tian, D.; Pan, J.; Chen, G.; Su, H.; Yan, Z.; Ji, C.; Tang, Z.; Fang, J. A field-based estimation of moso bamboo forest biomass in China. For. Ecol. Manag. 2022, 505, 119885. [Google Scholar] [CrossRef]
- Lima, R.A.; Rother, D.C.; Muler, A.E.; Lepsch, I.F.; Rodrigues, R.R. Bamboo overabundance alters forest structure and dynamics in the Atlantic Forest hotspot. Biol. Conserv. 2012, 147, 32–39. [Google Scholar] [CrossRef]
- Yang, Q.P.; Guo, Y.R.; Lan, W.J.; Ni Song, Q.; Yang, G.Y. Addition Effects of Co-Expansion of Two Bamboos on Plant Diversity in Broad-Leaved Forests. Chin. J. Appl. Ecol. 2017, 28, 3155–3162. [Google Scholar]
- Ouyang, M.; Yang, Q.; Chen, X.; Yang, G.; Shi, J.; Fang, X. Effects of the Expansion of Phyllostachys edulis on Species Composition, Structure and Diversity of the Secondary Evergreen Broad-Leaved Forests. Biodivers. Sci. 2016, 24, 649–657. [Google Scholar] [CrossRef] [Green Version]
- Griscom, B.W.; Ashton, P.M.S. A self-perpetuating bamboo disturbance cycle in a neotropical forest. J. Trop. Ecol. 2006, 22, 587–597. [Google Scholar] [CrossRef]
- Yang, Q.; Wang, B.; Guo, Q.; Zhao, G.; Fang, K.; Liu, Y. Effects of Phyllostachys edulis expansion on carbon storage of evergreen broad-leaved forest in Dagangshan Mountain, Jiangxi. Acta Agric. Univ. Jiangxiensis 2011, 33, 529–536. [Google Scholar]
- Liu, X.; Fang, P.; Xiong, Y.; Peng, Q.; Yu, Z.; Luan, F.; Song, Q.; Fang, X.; Yang, Q.; Liu, J. Assessment of the influence of bamboo expansion on Si pools and fluxes in a disturbed subtropical evergreen broadleaved forest. Catena 2022, 213, 106136. [Google Scholar] [CrossRef]
- Luan, J.; Li, S.; Dong, W.; Liu, Y.; Wang, Y.; Liu, S. Litter decomposition affected by bamboo expansion is modulated by litter-mixing and microbial composition. Funct. Ecol. 2021, 35, 2562–2574. [Google Scholar] [CrossRef]
- Wu, C.; Mo, Q.; Wang, H.; Zhang, Z.; Huang, G.; Ye, Q.; Zou, Q.; Kong, F.; Liu, Y.; Wang, G.G. Moso bamboo (Phyllostachys edulis (Carriere) J. Houzeau) invasion affects soil phosphorus dynamics in adjacent coniferous forests in subtropical China. Ann. For. Sci. 2018, 75, 24. [Google Scholar] [CrossRef] [Green Version]
- Hooper, D.U.; Bignell, D.; Brown, V.; Brussaard, L.; Dangerfield, J.; Wall, D.; Wardle, D.; Coleman, D.; Giller, K.; Lavelle, P.; et al. Interactions between Aboveground and Belowground Biodiversity in Terrestrial Ecosystems: Patterns, Mechanisms, and Feedbacks. Bioscience 2000, 50, 1049–1061. [Google Scholar] [CrossRef]
- Liu, W.; Liao, L.; Liu, Y.; Wang, Q.; Murray, P.J.; Jiang, X.; Zou, G.; Cai, J.; Zhao, X. Effects of Phyllostachys pubescens Expansion on Underground Soil Fauna Community and Soil Food Web in a Cryptomeria japonica Plantation, Lushan Mountain, Subtropical China. J. Soils Sediments 2021, 21, 2212–2227. [Google Scholar] [CrossRef]
- Liu, X.; Siemann, E.; Cui, C.; Liu, Y.; Guo, X.; Zhang, L. Moso bamboo (Phyllostachys edulis) invasion effects on litter, soil and microbial PLFA characteristics depend on sites and invaded forests. Plant Soil 2019, 438, 85–99. [Google Scholar] [CrossRef]
- Li, S.; Xie, D.; Ge, X.; Dong, W.; Luan, J. Altered diversity and functioning of soil and root-associated microbiomes by an invasive native plant. Plant Soil 2022, 473, 235–249. [Google Scholar] [CrossRef]
- Li, Y.; Li, Y.; Chang, S.X.; Xu, Q.; Guo, Z.; Gao, Q.; Qin, Z.; Yang, Y.; Chen, J.; Liang, X. Bamboo invasion of broadleaf forests altered soil fungal community closely linked to changes in soil organic C chemical composition and mineral N production. Plant Soil 2017, 418, 507–521. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: New York, NY, USA, 2008. [Google Scholar]
- Bowles, T.M.; Jackson, L.E.; Cavagnaro, T.R. Mycorrhizal fungi enhance plant nutrient acquisition and modulate nitrogen loss with variable water regimes. Glob. Chang. Biol. 2018, 24, e171–e182. [Google Scholar] [CrossRef]
- Hazard, C.; Johnson, D. Does Genotypic and Species Diversity of Mycorrhizal Plants and Fungi Affect Ecosystem Function? New Phytol. 2018, 220, 1122–1128. [Google Scholar] [CrossRef] [Green Version]
- Konvalinková, T.; Püschel, D.; Řezáčová, V.; Gryndlerová, H.; Jansa, J. Carbon Flow from Plant to Arbuscular Mycorrhizal Fungi Is Reduced under Phosphorus Fertilization. Plant Soil 2017, 419, 319–333. [Google Scholar] [CrossRef]
- El-Sawah, A.M.; Abdel-Fattah, G.G.; Holford, P.; Korany, S.M.; Alsherif, E.A.; AbdElgawad, H.; Ulhassan, Z.; Jośko, I.; Ali, B.; Sheteiwy, M.S. Funneliformis constrictum modulates polyamine metabolism to enhance tolerance of Zea mays L. to salinity. Microbiol. Res. 2023, 266, 127254. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; El-Sawah, A.M.; Korany, S.M.; Alsherif, E.A.; Mowafy, A.M.; Chen, J.; Jośko, I.; Selim, S.; AbdElgawad, H. Arbuscular Mycorrhizal Fungus “Rhizophagus irregularis” impacts on physiological and biochemical responses of ryegrass and chickpea plants under beryllium stress. Environ. Pollut. 2022, 315, 120356. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; Ali, D.F.I.; Xiong, Y.-C.; Brestic, M.; Skalicky, M.; Hamoud, Y.A.; Ulhassan, Z.; Shaghaleh, H.; AbdElgawad, H.; Farooq, M.; et al. Physiological and biochemical responses of soybean plants inoculated with Arbuscular mycorrhizal fungi and Bradyrhizobium under drought stress. BMC Plant Biol. 2021, 21, 195. [Google Scholar] [CrossRef]
- Tian, B.; Pei, Y.; Huang, W.; Ding, J.; Siemann, E. Increasing flavonoid concentrations in root exudates enhance associations between arbuscular mycorrhizal fungi and an invasive plant. ISME J. 2021, 15, 1919–1930. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Niu, L.; Wu, Q.; Chen, J.; Li, Y.; Liang, C.; Xu, Q.; Fuhrmann, J.J.; Shen, Y. Bamboo forest expansion increases soil organic carbon through its effect on soil arbuscular mycorrhizal fungal community and abundance. Plant Soil 2017, 420, 407–421. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Zobel, M. How mycorrhizal associations drive plant population and community biology. Science 2020, 367, eaba1223. [Google Scholar] [CrossRef] [PubMed]
- Zou, G.; Liu, Y.; Kong, F.; Liao, L.; Deng, G.; Jiang, X.; Cai, J.; Liu, W. Depression of the soil arbuscular mycorrhizal fungal community by the canopy gaps in a Japanese cedar (Cryptomeria japonica) plantation on Lushan Mountain, subtropical China. PeerJ 2021, 9, e10905. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.E.; Groten, K. The Costs and Benefits of Plant-Arbuscular Mycorrhizal Fungal Interactions. Annu. Rev. Plant Biol. 2022, 73, 649–672. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, L. Scientific Survey and Study of Biodiversity on the Lushan Nature Reserve in Jiangxi Province; Science Press: Beijing, China, 2010. [Google Scholar]
- Yang, F.Y.; Li, G.Z.; Zhang, D.E.; Christie, P.; Li, X.L.; Gai, J.P. Geographical and plant genotype effects on the formation of arbuscular mycorrhiza in Avena sativa and Avena nuda at different soil depths. Biol. Fertil. Soils 2010, 46, 435–443. [Google Scholar] [CrossRef] [Green Version]
- Lu, R. Methods of Soil and Agro-Chemical Analysis; China Agriculture Science and Technique Press: Beijing, China, 2000. [Google Scholar]
- Lee, J.; Lee, S.; Young, J.P.W. Improved PCR primers for the detection and identification of arbuscular mycorrhizal fungi. FEMS Microbiol. Ecol. 2008, 65, 339–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, L.; Lalonde, M.; Bruns, T.D. Specific amplification of 18S fungal ribosomal genes from vesicular-arbuscular endomycorrhizal fungi colonizing roots. Appl. Environ. Microbiol. 1992, 58, 291–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helgason, T.; Daniell, T.J.; Husband, R.; Fitter, A.H.; Young, J.P.W. Ploughing up the Wood-Wide Web? Nature 1998, 394, 431. [Google Scholar] [CrossRef] [PubMed]
- Xiang, D.; Verbruggen, E.; Hu, Y.; Veresoglou, S.D.; Rillig, M.C.; Zhou, W.; Xu, T.; Li, H.; Hao, Z.; Chen, Y.; et al. Land use influences arbuscular mycorrhizal fungal communities in the farming–pastoral ecotone of northern China. New Phytol. 2014, 204, 968–978. [Google Scholar] [CrossRef] [PubMed]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, J.; Zou, Y.; Zhang, J.; Peres-Neto, P.R. Generalizing hierarchical and variation partitioning in multiple regression and canonical analyses using the rdacca.hp R package. Methods Ecol. Evol. 2022, 13, 782–788. [Google Scholar] [CrossRef]
- R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2019.
- Zhao, Y.; Liang, C.; Shao, S.; Chen, J.; Qin, H.; Xu, Q. Linkages of litter and soil C:N:P stoichiometry with soil microbial resource limitation and community structure in a subtropical broadleaf forest invaded by Moso bamboo. Plant Soil 2021, 465, 473–490. [Google Scholar] [CrossRef]
- Wang, H.-C.; Tian, G.; Chiu, C.-Y. Invasion of moso bamboo into a Japanese cedar plantation affects the chemical composition and humification of soil organic matter. Sci. Rep. 2016, 6, 32211. [Google Scholar] [CrossRef] [Green Version]
- Chang, E.-H.; Chiu, C.-Y. Changes in soil microbial community structure and activity in a cedar plantation invaded by moso bamboo. Appl. Soil Ecol. 2015, 91, 1–7. [Google Scholar] [CrossRef]
- Nakane, K. Soil carbon cycling in a Japanese cedar (Cryptomeria japonica) plantation. For. Ecol. Manag. 1995, 72, 185–197. [Google Scholar] [CrossRef]
- Ouyang, M.; Tian, D.; Pan, J.; Chen, G.; Su, H.; Yan, Z.; Yang, Q.; Ji, C.; Tang, Z.; Fang, J. Moso bamboo (Phyllostachys edulis) invasion increases forest soil pH in subtropical China. Catena 2022, 215, 106339. [Google Scholar] [CrossRef]
- Zarei, M.; Hempel, S.; Wubet, T.; Schäfer, T.; Savaghebi, G.; Jouzani, G.S.; Nekouei, M.K.; Buscot, F. Molecular diversity of arbuscular mycorrhizal fungi in relation to soil chemical properties and heavy metal contamination. Environ. Pollut. 2010, 158, 2757–2765. [Google Scholar] [CrossRef] [PubMed]
- Inderjit; van der Putten, W. H. Impacts of Soil Microbial Communities on Exotic Plant Invasions. Trends Ecol. Evol. 2010, 25, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liang, X.; Tang, C.; Li, Y.; Chen, Z.; Chang, S.X.; Guo, Z.; Shen, Y.; Xu, Q. Moso bamboo invasion into broadleaf forests is associated with greater abundance and activity of soil autotrophic bacteria. Plant Soil 2018, 428, 163–177. [Google Scholar] [CrossRef]
- Lin, Y.-T.; Tang, S.-L.; Pai, C.-W.; Whitman, W.B.; Coleman, D.; Chiu, C.-Y. Changes in the Soil Bacterial Communities in a Cedar Plantation Invaded by Moso Bamboo. Microb. Ecol. 2014, 67, 421–429. [Google Scholar] [CrossRef]
- Fukushima, K.; Usui, N.; Ogawa, R.; Tokuchi, N. Impacts of moso bamboo (Phyllostachys pubescens) invasion on dry matter and carbon and nitrogen stocks in a broad-leaved secondary forest located in Kyoto, western Japan. Plant Species Biol. 2015, 30, 81–95. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Yang, Q.-P.; Song, Q.-N.; Yu, D.-K.; Yang, G.-Y.; Qi, H.-Y.; Shi, J.-M. Strategy of fine root expansion of Phyllostachys pubescens population into evergreen broad-leaved forest. Chin. J. Plant Ecol. 2013, 37, 230–238. [Google Scholar] [CrossRef]
- Leinster, T.; Cobbold, C.A. Measuring diversity: The importance of species similarity. Ecology 2012, 93, 477–489. [Google Scholar] [CrossRef] [Green Version]
- Wei, Q.; Yin, R.; Huang, J.; Vogler, A.P.; Li, Y.; Miao, X.; Kardol, P. The diversity of soil mesofauna declines after bamboo invasion in subtropical China. Sci. Total Environ. 2021, 789, 147982. [Google Scholar] [CrossRef]
- Balvanera, P.; Pfisterer, A.B.; Buchmann, N.; He, J.S.; Nakashizuka, T.; Raffaelli, D.; Schmid, B. Quantifying the Evidence for Biodiversity Effects on Ecosystem Functioning and Services. Ecol. Lett. 2006, 9, 1146–1156. [Google Scholar] [CrossRef] [Green Version]
- Croll, D.; Wille, L.; Gamper, H.A.; Mathimaran, N.; Lammers, P.J.; Corradi, N.; Sanders, I.R. Genetic diversity and host plant preferences revealed by simple sequence repeat and mitochondrial markers in a population of the arbuscular mycorrhizal fungus Glomus intraradices. New Phytol. 2008, 178, 672–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horn, S.; Hempel, S.; Verbruggen, E.; Rillig, M.C.; Caruso, T. Linking the Community Structure of Arbuscular Mycorrhizal Fungi and Plants: A Story of Interdependence? ISME J. 2017, 11, 1400–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wang, G.G.; Zhang, B.; Yuan, Z.; Fu, Z.; Yuan, Y.; Zhu, L.; Ma, S.; Zhang, J. Arbuscular Mycorrhizal Fungi Associated with Tree Species in a Planted Forest of Eastern China. Forests 2019, 10, 424. [Google Scholar] [CrossRef] [Green Version]
- Ma, M.; Ongena, M.; Wang, Q.; Guan, D.; Cao, F.; Jiang, X.; Li, J. Chronic fertilization of 37 years alters the phylogenetic structure of soil arbuscular mycorrhizal fungi in Chinese Mollisols. AMB Express 2018, 8, 57. [Google Scholar] [CrossRef] [Green Version]
- Biermann, B.; Linderman, R.G. Use of Vesicular-Arbuscular Mycor-Rhizal Roots, Intraradical Vesicles and Extraradical Vesicles Asinoculum. New Phytol. 1983, 95, 97–105. [Google Scholar] [CrossRef]
- Öpik, M.; Metsis, M.; Daniell, T.J.; Zobel, M.; Moora, M. Large-scale parallel 454 sequencing reveals host ecological group specificity of arbuscular mycorrhizal fungi in a boreonemoral forest. New Phytol. 2009, 184, 424–437. [Google Scholar] [CrossRef] [PubMed]
- Zou, N.; Shi, W.; Hou, L.; Kronzucker, H.J.; Huang, L.; Gu, H.; Yang, Q.; Deng, G.; Yang, G. Superior growth, N uptake and NH4+ tolerance in the giant bamboo Phyllostachys edulis over the broad-leaved tree Castanopsis fargesii at elevated NH4+ may underlie community succession and favor the expansion of bamboo. Tree Physiol. 2020, 40, 1606–1622. [Google Scholar] [CrossRef]
- Akama, A.; Mizoguchi, T.; Nagakura, J. Relationship between Nitrogen Utilization Characteristics and Suitable Growth Area for Pinus densiflora Sieb. Et Zucc. A Consideration on Nutritional Physiology of Forest Trees. Bull. For. For. Prod. Res. Inst. 2020, 19, 221–244. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stands | Soil Depth (cm) | SOM (g·kg−1) | TN (g·kg−1) | TP (mg·kg−1) | AK (mg·kg−1) | pH Value |
|---|---|---|---|---|---|---|
| JC | 0~10 | 98.06 ± 13.50 a | 3.32 ± 0.47 a | 184.66 ± 19.29 c | 71.65 ± 13.39 ab | 4.66 ± 0.07 a |
| 10~20 | 78.68 ± 20.94 a | 2.54 ± 0.59 a | 298.10 ± 35.00 b | 48.05 ± 12.22 b | 4.73 ± 0.14 a | |
| BC | 0~10 | 78.37 ± 16.70 a | 3.24 ± 0.63 a | 258.92 ± 23.52 bc | 84.41 ± 8.58 ab | 4.70 ± 0.05 a |
| 10~20 | 54.73 ± 11.63 a | 2.32 ± 0.44 a | 399.92 ± 30.42 a | 42.95 ± 3.36 b | 4.76 ± 0.03 a | |
| MB | 0~10 | 80.19 ± 4.23 a | 3.47 ± 0.15 a | 194.47 ± 20.29 c | 116.93 ± 16.58 a | 4.78 ± 0.08 a |
| 10~20 | 54.10 ± 0.06 a | 2.15 ± 0.06 a | 233.13 ± 13.84 bc | 49.64 ± 11.53 b | 4.90 ± 0.07 a |
| Stands | Soil Depth (cm) | Richness | Shannon-Wiener | Simpson | Evenness | ||||
|---|---|---|---|---|---|---|---|---|---|
| JC | 0~10 | 63.00 ± 4.58 bc | B | 2.84 ± 0.14 b | B | 0.891 ± 0.023 a | B | 0.687 ± 0.023 b | B |
| 10~20 | 59.00 ± 2.31 c | 2.80 ± 0.07 b | 0.894 ± 0.006 a | 0.686 ± 0.013 b | |||||
| BC | 0~10 | 77.00 ± 1.15 a | A | 3.40 ± 0.02 a | A | 0.943 ± 0.006 a | A | 0.782 ± 0.004 a | A |
| 10~20 | 76.00 ± 4.58 a | 3.40 ± 0.14 a | 0.940 ± 0.011 a | 0.784 ± 0.027 a | |||||
| MB | 0~10 | 74.33 ± 2.60 a | A | 3.13 ± 0.14 ab | B | 0.910 ± 0.018 a | B | 0.725 ± 0.047 ab | B |
| 10~20 | 71.67 ± 0.67 ab | 2.95 ± 0.13 b | 0.891 ± 0.022 a | 0.691 ± 0.031 b |
| Model | Variable | p Value | Relative Contribution (%) | |
|---|---|---|---|---|
| p value | 0.001 | SOM | 0.019 | 44.43 |
| TN | 0.011 | 18.33 | ||
| Adj R2 | 0.192 | TP | 0.016 | 37.24 |
| AK | 0.795 | 0.1 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, G.; Wu, B.; Chen, B.; Yang, Y.; Feng, Y.; Huang, J.; Liu, Y.; Murray, P.J.; Liu, W. What Are the Effects of Moso Bamboo Expansion into Japanese Cedar on Arbuscular Mycorrhizal Fungi: Altering the Community Composition Rather than the Diversity. J. Fungi 2023, 9, 273. https://doi.org/10.3390/jof9020273
Zou G, Wu B, Chen B, Yang Y, Feng Y, Huang J, Liu Y, Murray PJ, Liu W. What Are the Effects of Moso Bamboo Expansion into Japanese Cedar on Arbuscular Mycorrhizal Fungi: Altering the Community Composition Rather than the Diversity. Journal of Fungi. 2023; 9(2):273. https://doi.org/10.3390/jof9020273
Chicago/Turabian StyleZou, Guiwu, Binsheng Wu, Baodong Chen, Yaying Yang, Yan Feng, Jiahui Huang, Yuanqiu Liu, Philip J. Murray, and Wei Liu. 2023. "What Are the Effects of Moso Bamboo Expansion into Japanese Cedar on Arbuscular Mycorrhizal Fungi: Altering the Community Composition Rather than the Diversity" Journal of Fungi 9, no. 2: 273. https://doi.org/10.3390/jof9020273