
Four New Wood-Inhabiting Fungal Species of Peniophoraceae (Russulales, Basidiomycota) from the Yunnan-Guizhou Plateau, China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphology
2.2. Molecular Phylogeny

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Specimen No. | GenBank Accession No. | References | Country | |
|---|---|---|---|---|---|
| ITS | nLSU | ||||
| Amylostereum areolatum | NH 8041 | AF506405 | AF506405 | [45] | Sweden |
| A. chailletii | NH 8031 | AF506406 | AF506406 | [45] | Sweden |
| A. laevigatum | NH 12863 | AF506407 | AF506407 | [45] | Sweden |
| Asterostroma bambusicola | He 4132 | KY263871 | this publication | Thailand | |
| A. cervicolor | He 2314 | KY263869 | this publication | China | |
| A. laxum | EL 33-99 | AF506410 | AF506410 | [45] | Sweden |
| A. vararioides | He 4140 | KY263870 | this publication | Thailand | |
| Auriscalpium vulgare | EL 33-95 | AF506375 | AF506375 | [45] | Sweden |
| Baltazaria galactina | CBS: 752.86 | MH873721 | [49] | France | |
| B. neogalactina | CBS: 755.86 | MH873724 | MH873724 | [49] | French |
| B. occidentalis | AFTOL-ID | DQ234539 | [50] | Canada | |
| B. podocarpi | Dai 9261 | KJ583221 | [51] | China | |
| Dentipratulum bialoviesense | GG 1645 | AF506389 | AF506389 | [45] | Sweden |
| Dichostereum durum | FG 1985 | AF506429 | AF506429 | [45] | Sweden |
| D. effuscatum | GG 930915 | AF506390 | AF506390 | [45] | Sweden |
| Gloeocystidiellum bisporum | KHL 11135 | AY048877 | AY048877 | [45] | Sweden |
| G. clavuligerum | FCUG 2159 | AF310088 | AF310088 | [52] | Spain |
| G. purpureum | Wu 9310-45 | AF441338 | AF441338 | [45] | China |
| Gloeocystidiopsis flammea | CBS: 324.66 | AF506437 | AF506437 | [45] | C. African Rep. |
| Gloeodontia columbiensis | NH 11118 | AF506444 | AF506444 | [45] | Spain |
| G. discolor | KHL 10099 | AF506445 | AF506445 | [45] | USA |
| G. eriobotryae | Dai 12080 | JQ349103 | [53] | China | |
| G. pyramidata | LR 15502 | AF506446 | AF506446 | [45] | Colombia |
| G. subasperispora | KHL 8695 | AF506404 | AF506404 | [45] | Norway |
| G. yunnanensis | SWFC 00010504 | MN908254 | [54] | China | |
| Gloeopeniophorella convolvens | KHL 10103 | AF506435 | AF506435 | [45] | USA |
| Gloiothele lactescens | EL 8-98 | AF506453 | AF506453 | [45] | Sweden |
| G. lamellosa | KHL 11031 | AF506454 | AF506454 | [45] | Venezuela |
| Heterobasidion annosum | 06129/6 | KJ583225 | [51] | Russia | |
| H. parviporum | 04121/3 | KJ583226 | [51] | Finland | |
| Lachnocladium schweinfurthianum | KM 49740 | MH260051 | [38] | Cameroon | |
| Lactarius leonis | SJ 91016 | AF506411 | AF506411 | [45] | Sweden |
| Lentinellus cochleatus | KGN 960928 | AF506417 | AF506417 | [45] | Sweden |
| L. ursinus | EL 73-97 | AF506419 | AF506419 | [45] | USA |
| L. vulpinus | KGN 980825 (GB) | AF347097 | AF347097 | [45] | Sweden |
| Megalocystidium luridum | KHL 8635 | AF506422 | AF506422 | [45] | Norway |
| Michenera artocreas | GHL-2016-Oct | MH204692 | [55] | USA | |
| M. incrustata | He 2630 | MH142907 | [55] | China | |
| Peniophora albobadia | CBS: 329.66 | MH858809 | MH858809 | [49] | France |
| P. bicornis | He 3609 | MK588763 | MK588763 | [39] | China |
| P. bicornis | He 4767 | MK588764 | MK588764 | [39] | China |
| P. borbonica | He 4597 | MK588766 | MK588766 | [39] | China |
| P. borbonica | He 4606 | MK588765 | MK588765 | [39] | China |
| P. cinerea | B 1020 | MN475151 | MN475151 | [39] | USA |
| P. crassitunicata | CBS: 663.91 | MH862292 | MH862292 | [49] | France |
| P. duplex | B 1022 | MN475153 | MN475153 | [39] | USA |
| P. erikssonii | CBS: 287.58 | MH857788 | MH857788 | [39] | France |
| P.erikssonii | Cui 11871 | MK588771 | MK588811 | [39] | China |
| P. exima | B 1011 | MN475155 | MN475155 | [39] | USA |
| P. exima | T-523 | MK588772 | MK588772 | [39] | USA |
| P. fasticata | CBS: 942.96 | MH862624 | MH862624 | [39] | Ethiopia |
| P. fissilis | CBS: 681.91 | MH862298 | MH862298 | [39] | France |
| P. fissilis | CBS: 684.91 | MH862299 | MH862299 | [39] | Netherlands |
| P. gabonensis | CBS: 673.91 | MH862293 | MH862293 | [39] | Gabon |
| P. gilbertsonii | CBS: 357.95 | MH862528 | MH862528 | [39] | USA |
| P. gilbertsonii | CBS: 360.95 | MH862530 | MH862530 | [39] | USA |
| P. halimi | CBS: 862.84 | MH861843 | MH861843 | [39] | France |
| P. incarnata | NH 10271 | AF506425 | AF506425 | [45] | Denmark |
| P. incarnata | CBS: 430.72 | MH860518 | MH872230 | [39] | Netherlands |
| P. junipericola | He 2462 | MK588773 | MK588773 | this publication | China |
| P. laeta | CBS: 256.56 | MH857617 | MH857617 | [39] | France |
| P. laeta | CBS: 255.56 | MH857616 | MH857616 | [39] | France |
| P. laxitexta | LGMF 1159 | JX559580 | [39] | Brazil | |
| P. laxitexta | BAFC 3309 | FJ882040 | [39] | Argentina | |
| P. laxitexta | BAFC: 4687 | MN518328 | [39] | Argentina | |
| P. lilacea | CBS: 337.66 | MH858813 | MH858813 | [39] | Armenia |
| P. lycii | CBS: 264.56 | MH857624 | MH857624 | [39] | France |
| P. lycii | CBS: 261.56 | MH857621 | MH857621 | [39] | France |
| P. malaiensis | He 4870 | MK588775 | MK588775 | [39] | China |
| P. manshurica | He 2956 | MK588776 | MK588776 | [39] | China |
| P. manshurica | He 3729 | MK588777 | MK588777 | [39] | China |
| P. meridionalis | CBS: 289.58 | MH857789 | MH857789 | [49] | France |
| P. molesta | CBS: 678.91 | MH862296 | MH862296 | [39] | Cote d’Ivoire |
| P. molesta | CBS: 676.91 | MH862294 | MH862294 | [39] | Gabon |
| P. molesta | CBS: 677.91 | MH862295 | MH862295 | [39] | Gabon |
| P. monticola | CBS: 649.91 | MH862289 | MH862289 | [39] | France |
| P. nuda | AFTOL-ID 660 | DQ411533 | [39] | USA | |
| P. nuda | LZ15-07 | MT859929 | MT859929 | this publication | China |
| P. ovalispora | CBS: 653.91 | MH873971 | [39] | Netherlands | |
| P. ovalispora | CBS: 653.91 | MH862290 | MH862290 | [39] | Netherlands |
| P. parvocystidiata | CBS: 716.91 | MH862305 | MH862305 | [39] | France |
| P. parvocystidiata | CBS: 717.91 | MH862306 | MH862306 | [39] | France |
| P. piceae | B 1010 | MN475158 | MN475158 | this publication | USA |
| P. pilatiana | CBS: 269.56 | MH857627 | MH857627 | [39] | France |
| P. pilatiana | CBS: 265.56 | MH857625 | MH857625 | [39] | France |
| P. pilatiana | CBS: 266.56 | MH857626 | MH857626 | [39] | France |
| P. pini | CBS: 273.56 | MH857631 | MH857631 | [39] | France |
| P. pini | CBS: 270.56 | MH857628 | MH857628 | [39] | France |
| P. pithya | CBS: 275.56 | MH857633 | MH857633 | [49] | France |
| P. polygonia | He 3668 | MH669233 | [56] | China | |
| P. polygonia | CBS: 404.50 | MH856684 | MH856684 | [39] | France |
| P. proxima | CBS: 406.50 | MH856686 | MH856686 | [39] | France |
| P. proxima | CBS: 405.50 | MH856685 | MH856685 | [39] | France |
| P. pseudonuda | FCUG 2384 | GU322866 | this publication | Sweden | |
| P. pseudonuda | FCUG 2390 | GU322865 | this publication | Sweden | |
| P. pseudopini | B 1024 | MN475163 | MN475163 | this publication | USA |
| P. pseudoversicolor | CBS: 125881 | MH864303 | MH864303 | [39] | France |
| P. quercina | CBS: 407.50 | MH856687 | MH868204 | [39] | France |
| P. quercina | CBS: 408.50 | MH856688 | MH856688 | [39] | France |
| P. quercina | CBS: 409.50 | MH856689 | MH856689 | [39] | France |
| P. reidii | CBS: 397.83 | MH861616 | MH861616 | [39] | France |
| P. rosealba | CLZhao 3513 | ON786559 | OP380690 | present study | China |
| P. rosealba | CLZhao 9401 * | ON786560 | present study | China | |
| P. rufa | B 1014 | MN475165 | MN475165 | this publication | USA |
| P. rufa | CBS: 351.59 | MH857891 | MH869432 | [39] | Canada |
| P. rufomarginata | CBS: 281.56 | MH857639 | MH857639 | [39] | France |
| P. rufomarginata | CBS: 282.56 | MH857640 | MH857640 | [39] | France |
| P. septentrionalis | CBS: 294.58 | MH857791 | MH857791 | [39] | Canada |
| P. simulans | CBS: 875.84 | MH861850 | MH861850 | [39] | France |
| P. simulans | CBS: 874.84 | MH861849 | MH861849 | [39] | France |
| P. subsalmonea | CBS: 697.91 | MH862303 | MH862303 | [39] | Netherlands |
| P. subsalmonea | CBS: 696.91 | MH862302 | MH862302 | [39] | Netherlands |
| P. taiwanensis | Wu 9209-14 | MK588794 | MK588794 | [39] | China |
| P. tamaricicola | CBS: 438.62 | MH858203 | MH858203 | [39] | Morocco |
| P. tamaricicola | CBS: 439.62 | MH858204 | MH858204 | [39] | Morocco |
| P. tamaricicola | CBS: 441.62 | MH858205 | MH858205 | [39] | Morocco |
| P. versicolor | CBS: 358.61 | MH858082 | MH858082 | [39] | Morocco |
| P. violaceolivida | CBS: 348.52 | MH857077 | MH857077 | [39] | France |
| P. yunnanensis | CLZhao 3978 | OP380617 | OP380689 | present study | China |
| P. yunnanensis | CLZhao 7347 * | OP380616 | present study | China | |
| P. yunnanensis | CLZhao 8135 | OP380615 | present study | China | |
| Russula violacea | SJ 93009 | AF506465 | AF506465 | [45] | Sweden |
| Scytinostroma portentosum | EL 11-99 | AF506470 | AF506470 | [45] | Sweden |
| Sistotrema brinkmannii | NH 11412 | AF506473 | AF506473 | [45] | Turkey |
| S. coronilla | NH 7598 | AF506475 | AF506475 | [45] | Canada |
| Stereum hirsutum | NH 7960 | AF506479 | AF506479 | [45] | Romania |
| Vararia abortiphysa | CBS: 632.81 | MH861387 | MH861387 | [49] | Gabon |
| V. ambigua | CBS: 634.81 | MH861388 | MH873137 | [49] | France |
| V. amphithallica | CBS: 687.81 | MH861431 | MH861431 | [49] | France |
| V. aurantiaca | CBS: 642.81 | MH861394 | MH861394 | [49] | Gabon |
| V. aurantiaca | CBS: 641.81 | MH861393 | MH861393 | [49] | France |
| V. breviphysa | CBS: 644.81 | MH861396 | MH861396 | [49] | Gabon |
| V. calami | CBS: 646.81 | MH861398 | MH861398 | [49] | France |
| V. calami | CBS: 648.81 | MH861399 | MH861399 | [49] | France |
| V. callichroa | CBS: 744.91 | MH874000 | MH874000 | [49] | France |
| V. cinnamomea | CBS: 642.84 | MH873488 | MH873488 | [49] | Madagascar |
| V. cinnamomea | CBS: 641.84 | MH861794 | MH861794 | [49] | Madagascar |
| V. cremea | CBS: 651.81 | MH873147 | MH873147 | [49] | France |
| V. daweishanensis | CLZhao 17911 | OP380613 | OP615103 | present study | China |
| V. daweishanensis | CLZhao 17936 * | OP380614 | OP380688 | present study | China |
| V. dussii | CBS: 655.81 | MH861405 | MH861405 | [49] | France |
| V. dussii | CBS: 652.81 | MH873148 | MH873148 | [49] | France |
| V. ellipsospora | HHB-19503 | MW740328 | MW740328 | this publication | New Zealand |
| V. fragilis | CLZhao 2628 | OP380611 | present study | China | |
| V. fragilis | CLZhao 16475 * | OP380612 | OP380687 | present study | China |
| V. fusispora | PDD: 119539 | OL709443 | OL709443 | this publication | New Zealand |
| V. gallica | CBS: 234.91 | MH862250 | [49] | Canada | |
| V. gallica | CBS: 656.81 | MH861406 | MH873152 | [49] | France |
| V. gillesii | CBS: 660.81 | MH873153 | MH873153 | [49] | Cote d’Ivoire |
| V. gomezii | CBS: 661.81 | MH873154 | MH873154 | [49] | French |
| V. gracilispora | CBS: 664.81 | MH861412 | MH861412 | [49] | Gabon |
| V. gracilispora | CBS: 663.81 | MH861411 | [49] | Gabon | |
| V. insolita | CBS: 668.81 | MH861413 | MH861413 | [49] | France |
| V. intricata | CBS: 673.81 | MH861418 | MH861418 | [49] | France |
| V. investiens | FP-151122 | MH971976 | MH971977 | [56] | USA |
| V. malaysiana | CBS: 644.84 | MH873490 | MH873490 | [49] | Singapore |
| V. minispora | CBS: 682.81 | MH861426 | MH861426 | [49] | France |
| V. ochroleuca | CBS: 465.61 | MH858109 | MH858109 | [49] | France |
| V. ochroleuca | JS 24400 | AF506485 | AF506485 | [45] | Norway |
| V. parmastoi | CBS: 879.84 | MH861852 | MH861852 | [49] | Uzbekistan |
| V. perplexa | CBS: 695.81 | MH861438 | MH861438 | [49] | France |
| V. pectinata | CBS: 685.81 | MH861429 | [49] | Cote d’Ivoire | |
| V. pirispora | CBS: 720.86 | MH862016 | MH862016 | [49] | France |
| V. rhombospora | CBS: 743.81 | MH861470 | MH861470 | [49] | France |
| V. rosulenta | CBS: 743.86 | MH862028 | [49] | France | |
| V. rugosispora | CBS: 697.81 | MH861440 | MH861440 | [49] | Gabon |
| V. sigmatospora | CBS: 748.91 | MH874001 | MH874001 | [49] | Netherlands |
| V. sphaericospora | CBS: 700.81 | MH873185 | MH873185 | [49] | Gabon |
| V. sphaericospora | CBS: 703.81 | MH861446 | MH861446 | [49] | Gabon |
| V. trinidadensis | CBS: 651.84 | MH861803 | MH861803 | [49] | Madagascar |
| V. trinidadensis | CBS: 650.84 | MH873495 | MH873495 | [49] | Madagascar |
| V. tropica | CBS: 704.81 | MH861447 | MH873189 | [49] | France |
| V. vassilievae | UC2022892 | KP814203 | KP814203 | this publication | USA |
| V. verrucosa | CBS 706.81 | MH861449 | MH861449 | [49] | France |
| Vesiculomyces citrinus | EL 53-97 | AF506486 | AF506486 | [45] | Sweden |
3. Results
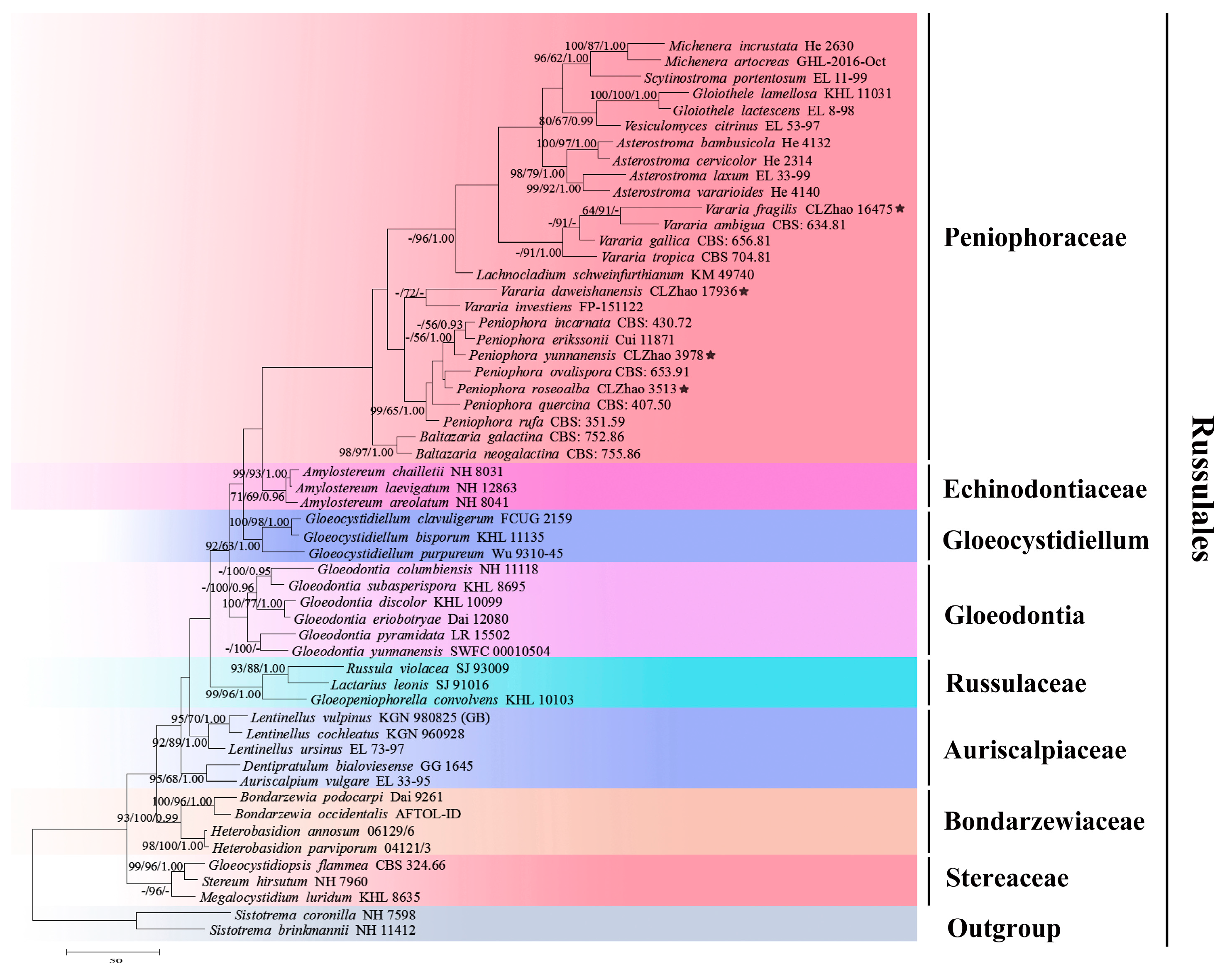
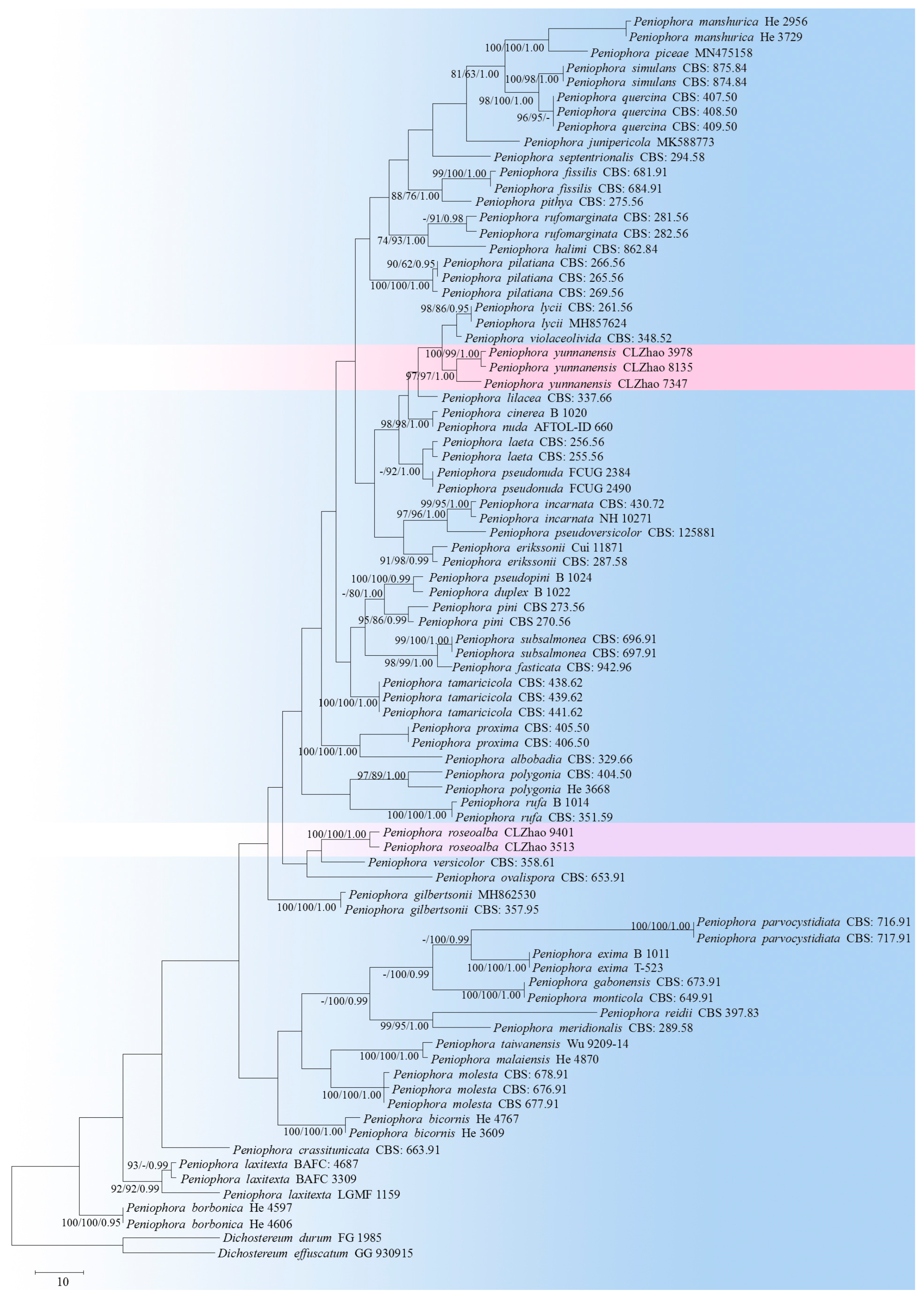
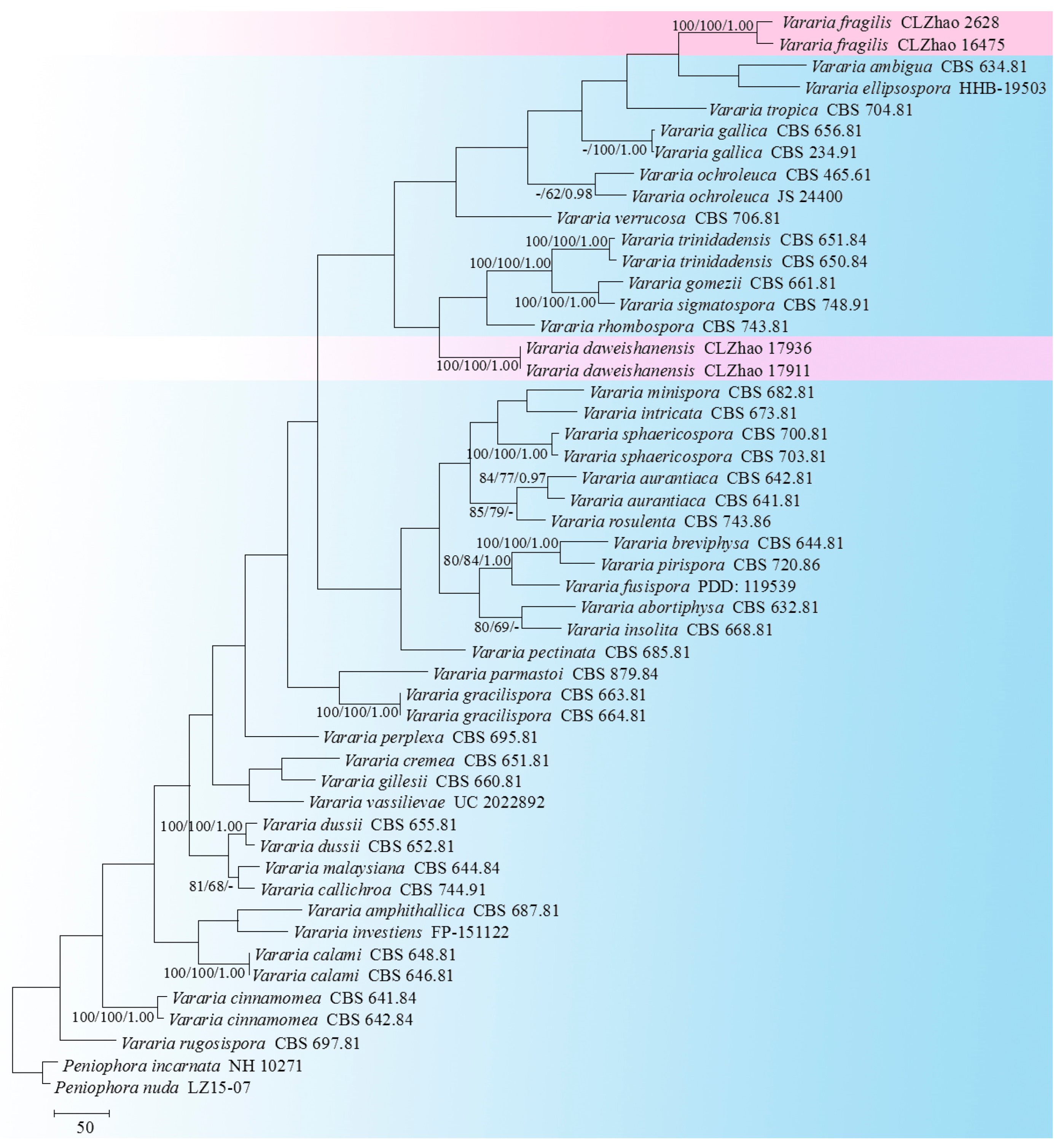
3.1. Molecular Phylogeny
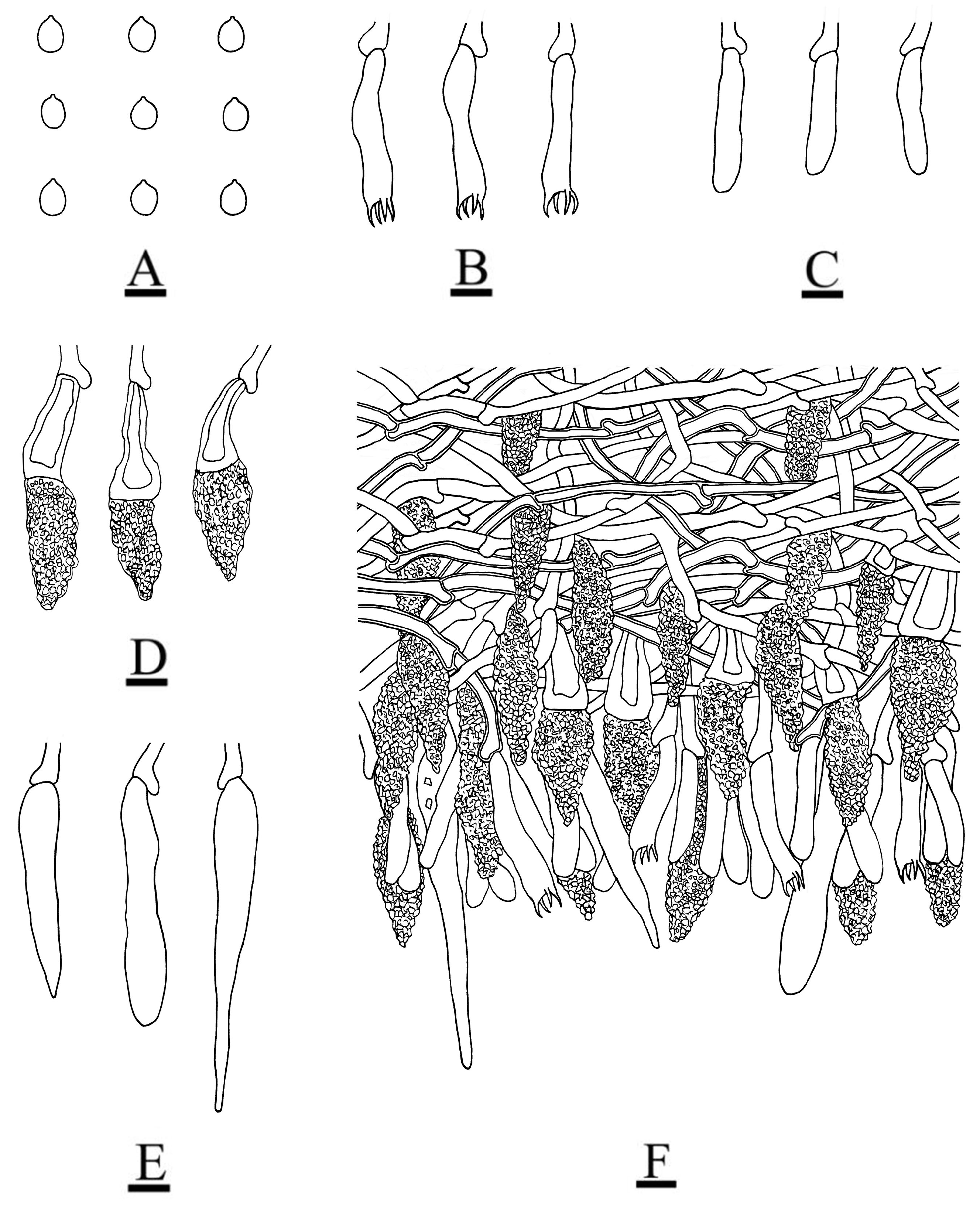
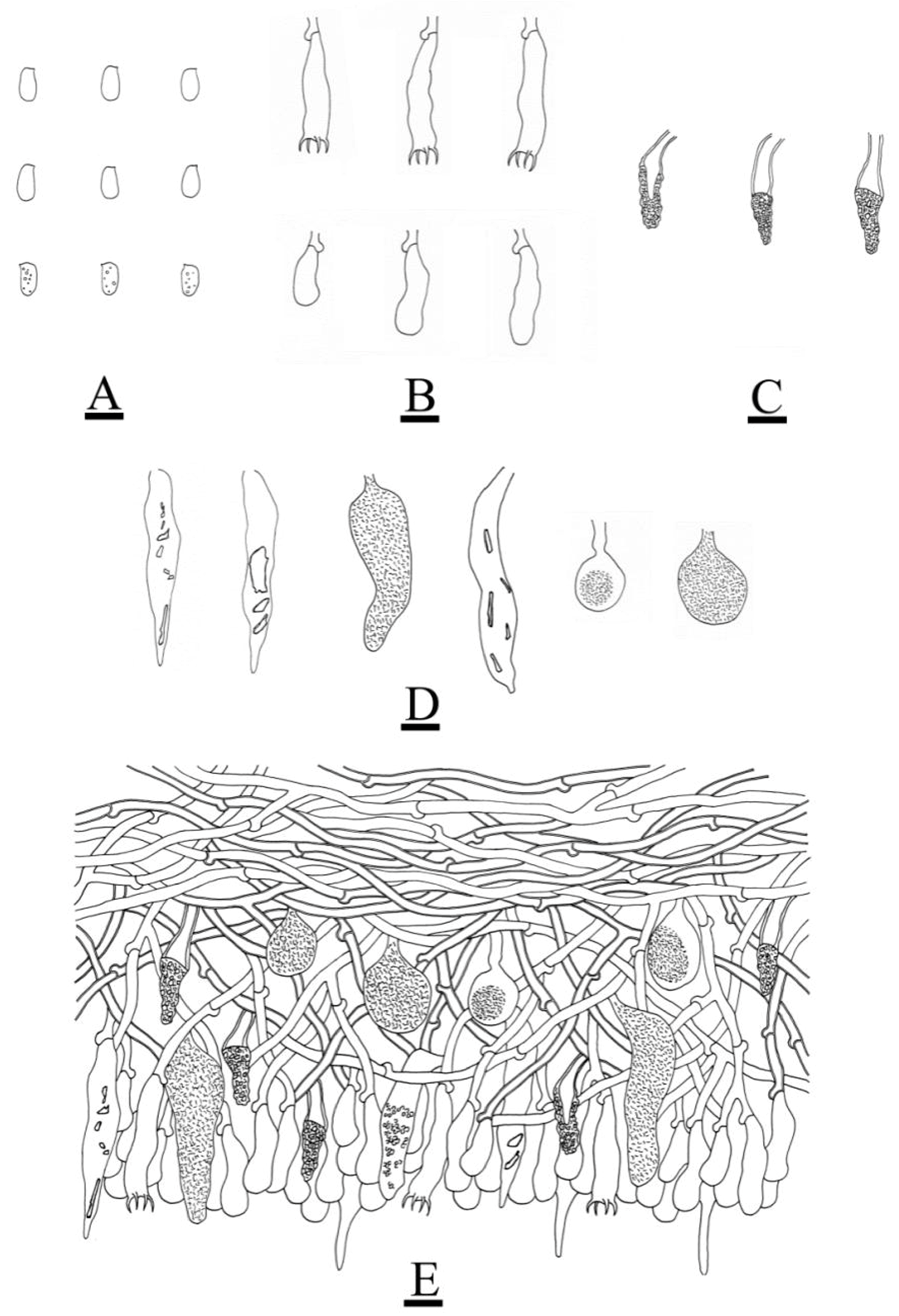
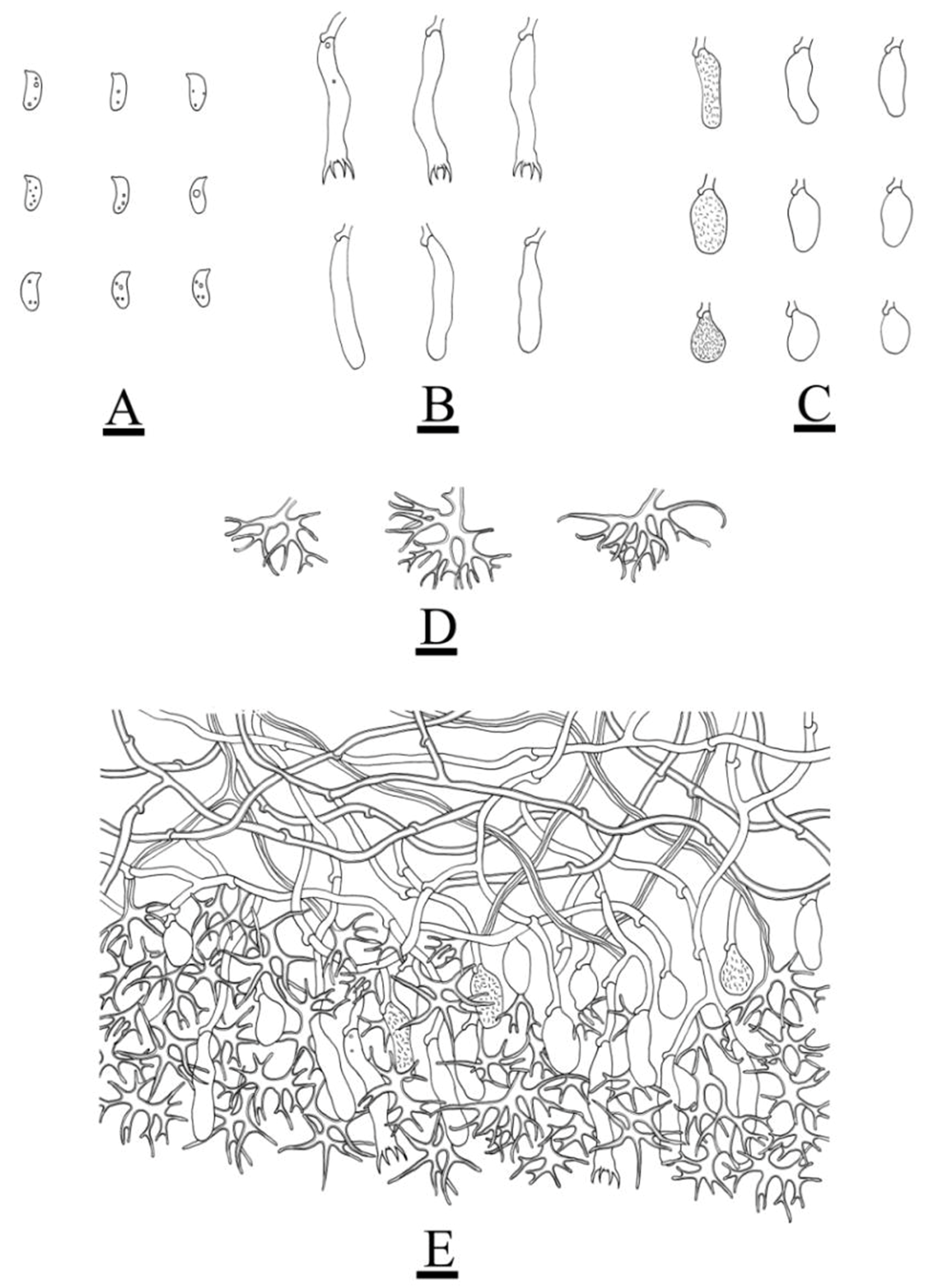
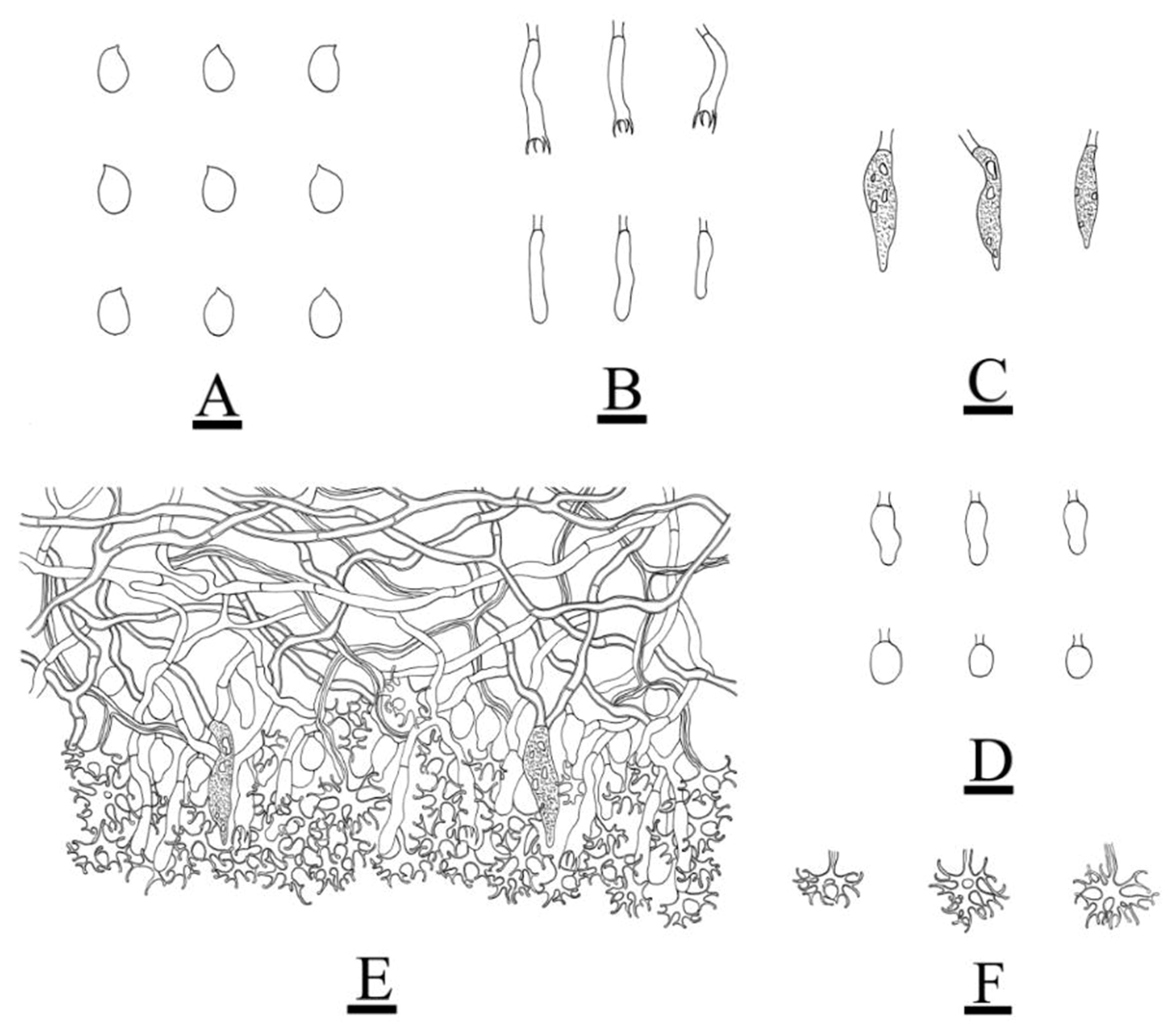
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tedersoo, L.; Bahram, M.; Põlme, S.; Koljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008; p. 771. [Google Scholar]
- Cooke, M.C. On Peniophora. Grevillea 1879, 8, 17–21. [Google Scholar]
- Burt, E.A. The Thelephoraceae of North America. XIV. Peniophora. Ann. Mo. Bot. Gard. 1925, 12, 213–357. [Google Scholar] [CrossRef]
- Hennings, P. Fungi Africae orientalis III. Bot. Jahrbücher Für Syst. Pflanzengesch. Pflanzengeogr. 1904, 34, 39–57. [Google Scholar]
- Hjortstam, K.; Ryvarden, L. Some new and noteworthy Basidiomycetes (Aphyllophorales) from Nepal. Mycotaxon 1984, 20, 133–151. [Google Scholar]
- Hjortstam, K.; Roberts, P.J.; Spooner, B.M. Corticioid fungi from Brunei Darussalam. Kew Bull. Addit. Ser. 1998, 53, 805–827. [Google Scholar] [CrossRef]
- Bourdot, H. Corticiés nouveaux de la flore mycologique de France III. Rev. Sci. Bourbon. Cent. Fr. 1910, 23, 3–15. [Google Scholar]
- Boidin, J. Hétérobasidiomycètes saprophytes et Homobasidiomycètes résupinés. II. Catalogue raisonné des espèces pyrénéennes de la région de Luchon (Haute-Garonnes). Bull. Société D’histoire Nat. Toulouse 1957, 92, 277–292. [Google Scholar]
- Wu, S.H. Three new species of corticioid fungi from Taiwan. Bot. Stud. 2007, 48, 325–330. [Google Scholar] [CrossRef]
- Ranojevic, N. Zweiter beitrag zur pilzflora serbiens. Ann. Mycol. 1910, 8, 347–402. [Google Scholar]
- Dhingra, G.S. Peniophora hallenbergii sp. nov. from India. Mycotaxon 2013, 126, 235–237. [Google Scholar] [CrossRef]
- Boidin, J.; Lanquetin, P.; Gilles, G. Les Peniophoraceae de la zone intertropicale (Basidiomycetes, Aphyllophorales). Bull. Société Mycol. Fr. 1991, 107, 91–156. [Google Scholar]
- Parmasto, E.; Parmasto, I. Variation in basidiospores in the Hymenomycetes and its significance to their taxonomy. Bibl. Mycol. 1987, 115, 1–168. [Google Scholar]
- Wu, S.H. A study of Peniophora species with simple-septate hyphae occurring in Taiwan. Mycotaxon 2003, 85, 187–199. [Google Scholar]
- Popoff, O.F.; Wright, J.E. Two new corticioid fungi (Aphyllophorales) from NE Argentina and Paraguay. Mycotaxon 1994, 51, 317–324. [Google Scholar]
- Gorjón, S.P.; Jesus, M.A. Some new species and new records of corticioid fungi (Basidiomycota) from the Brazilian Amazon. Phytotaxa 2012, 67, 38–54. [Google Scholar] [CrossRef] [Green Version]
- Jackson, H.S.; Dearden, E.R. Studies of Canadian Thelephoraceae. III. Some new species from British Columbia. Can. J. Res. 1949, 27, 147–156. [Google Scholar] [CrossRef]
- Bernicchia, A.; Gorjón, S.P. Fungi Europaei 12: Corticiaceae s.l.; Edizioni Candusso: Alassio, Italy, 2010. [Google Scholar]
- Boidin, J.; Lanquetin, P. Vararia subgenus Vararia (Basidiomycetes, Lachnocladiaceae): Étude spèciale des espèces d’Afrique intertropicale. Bull. Soc. Mycol. 1975, 91, 457–513. [Google Scholar]
- Boidin, J.; Lanquetin, P.; Gilles, G. Application du concept biologique del’espèce aux Basidiomycètes. Le genre Vararia section Vararia au Gabon. Cryptogam. Mycol. 1980, 1, 265–384. [Google Scholar]
- Cunningham, G.H. Thelephoraceae of New Zealand. Part IV. The genui Vararia. Trans. Roy. Soc. NZ. 1955, 82, 973–985. [Google Scholar]
- Gilbertson, R.L. Some species of Vararia from temperate North America. Pap. Mich. Acad. Sci. 1965, 50, 161–184. [Google Scholar]
- Boidin, J. Basidiomycètes Lachnocladiaceae résupinés de la Republique Centrafricaine. Cah. Maboké 1967, 5, 23–35. [Google Scholar]
- Boidin, J.; Lanquetin, P. Compléments au genre Vararia P. Karst. (Basidiomycètes). Pers. -Mol. Phylogeny Evol. Fungi 1984, 12, 243–262. [Google Scholar]
- Pouzar, Z. Taxonomic studies in resupinate fungi I. Česká Mykol 1982, 36, 141–145. [Google Scholar]
- Boidin, J.; Gilles, G. Contribution à la connaissance du genre Vararia (Basidiomycotina). Bull. Soc. Mycol. 1999, 115, 115–139. [Google Scholar]
- Duhem, B.; Buyck, B. On two new tropical Vararia (Russulales, Basidiomycota) with extremely small, racemose dichohyphidia. Cryptogam. Mycol. 2012, 33, 427–437. [Google Scholar] [CrossRef]
- Sanyal, S.K.; Dhingra, G.S.; Singh, A.P. Vararia longicystidiata sp. nov.(Agaricomycetes) from India. Mycotaxon 2012, 120, 357–360. [Google Scholar] [CrossRef]
- Nakasone, K.K. Taxonomic studies in Chrysoderma, Corneromyces, Dendrophysellum, Hyphoradulum, and Mycobonia. Mycotaxon 2015, 130, 369–397. [Google Scholar] [CrossRef]
- Liu, S.L.; He, S.H. The genus Vararia (Russulales, Basidiomycota) in China. Two new species and two new Chinese records. Nord. J. Bot. 2016, 34, 553–558. [Google Scholar] [CrossRef]
- Dai, Y.C. A revised checklist of corticioid and hydnoid fungi in China for 2010. Mycoscience 2011, 52, 69–79. [Google Scholar] [CrossRef]
- Dai, Y.C.; Yang, Z.L.; Cui, B.K.; Wu, G.; Yuan, H.S.; Zhou, L.W.; He, S.H.; Ge, Z.W.; Wu, F.; Wei, Y.L.; et al. Diversity and systematics of the important macrofungi in Chinese forests. Mycosystema 2021, 40, 770–805. [Google Scholar] [CrossRef]
- Liu, S.L. Taxonomy and Phylogeny of Vararia and Related Genera in China. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2019. [Google Scholar]
- Larsson, K.H.; Larsson, E.; Kõljalg, U. High phylogenetic diversity among corticioid homobasidiomycetes. Mycol. Res. 2004, 108, 983–1002. [Google Scholar] [CrossRef] [PubMed]
- Binder, M.; Hibbett, D.S.; Larsson, K.H.; Larsson, E.; Langer, E.; Langer, G. The phylogenetic distribution of resupinate forms across the major clades of mushroom-forming fungi (Homobasidiomycetes). Syst. Biodivers. 2005, 3, 113–157. [Google Scholar] [CrossRef]
- Leal-Dutra, C.A.; Neves, M.A.; Griffith, G.W.; Reck, M.A.; Clasen, L.A.; Dentinger, B.T.M. Reclassification of Parapterulicium corner (Pterulaceae, Agaricales), contributions to Lachnocladiaceae and Peniophoraceae (Russulales) and introduction of Baltazaria gen. nov. MycoKeys 2018, 37, 39–56. [Google Scholar] [CrossRef] [PubMed]
- Luo, K.Y.; Chen, Z.Y.; Zhao, C.L. Phylogenetic and taxonomic analyses of three new wood-inhabiting fungi of Xylodon (Basidiomycota) in a forest ecological system. J. Fungi 2022, 8, 405. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.H. Farvekort. In The Danish Mycological Society’s Colour-Chart; Foreningen til Svampekundskabens Fremme: Greve, Germany, 1996; pp. 1–6. [Google Scholar]
- Qu, M.H.; Wang, D.Q.; Zhao, C.L. A phylogenetic and taxonomic study on Xylodon (Hymenochaetales): Focusing on three new Xylodon species from southern China. J. Fungi 2022, 8, 35. [Google Scholar] [CrossRef]
- Dai, Y.C. Polypore diversity in China with an annotated checklist of Chinese polypores. Mycoscience 2012, 53, 49–80. [Google Scholar] [CrossRef]
- Zhao, C.L.; Wu, Z.Q. Ceriporiopsis kunmingensis sp. nov. (Polyporales, Basidiomycota) evidenced by morphological characters and phylogenetic analysis. Mycol. Prog. 2017, 16, 93–100. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Larsson, E.; Larsson, K.H. Phylogenetic relationships of russuloid basidiomycetes with emphasis on aphyllophoralean taxa. Mycologia 2003, 95, 1037–1065. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP*: Phylogenetic analysis using Parsimony (*and Other Methods); Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Felsenstein, J. Confidence intervals on phylogenetics: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES Science Gateway: Enabling high-impact science for phylogenetics researchers with limited resources. Assoc. Comput. Mach. 2012, 39, 1–8. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Matheny, P.B.; Wang, Z.; Binder, M.; Curtis, J.M.; Lim, Y.W.; Nilsson, R.H.; Hughes, K.W.; Hofstetter, V.; Ammirati, J.F.; Schoch, C.L.; et al. Contributions of rpb2 and tef1 to the phylogeny of mushrooms and allies (Basidiomycota, Fungi). Mol. Phylogenet. Evol. 2007, 43, 430–451. [Google Scholar] [CrossRef]
- Chen, J.J.; Shen, L.L. Amylosporus succulentus sp. nov. (Russulales, Basidiomycota) evidenced by morphological characters and phylogenetic analysis. Cryptogam. Mycol. 2014, 35, 271–282. [Google Scholar] [CrossRef]
- Larsson, E.; Hallenberg, N. Species delimitation in the Gloeocystidiellum porosum-clavuligerum complex inferred from compatibility studies and nuclear rDNA sequence data. Mycologia 2001, 93, 907–914. [Google Scholar] [CrossRef]
- Zhou, L.W.; Dai, Y.C. Taxonomy and phylogeny of wood-inhabiting hydnoid species in Russulales: Two new genera, three new species and two new combinations. Mycologia 2013, 105, 636–649. [Google Scholar] [CrossRef]
- Chen, L.; Shi, Z.J.; Wu, C.H.; Zhao, C.L. Gloeodontia yunnanensis sp. nov. (Russulales, Basidiomycota) from China, evidenced by morphological characters and phylogenetic analyses. Phytotaxa 2020, 43, 111–118. [Google Scholar] [CrossRef]
- Liu, S.L.; Nakasone, K.K.; He, S.H. Michenera incrustata sp. nov. (Peniophoraceae, Russulales) from southern China. Nova Hedwig. 2019, 108, 197–206. [Google Scholar] [CrossRef]
- Liu, S.L.; He, S.H. Taxonomy and phylogeny of Dichostereum (Russulales), with descriptions of three new species from southern China. MycoKeys 2018, 40, 111–126. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saccardo, P.A.; Sydow, P. Supplementum universale, Pars V. Sylloge Fungorum 1902, 16, 1–1291. [Google Scholar]
- Andreasen, M.; Hallenberg, N. A taxonomic survey of the Peniophoraceae. Synop Fungorum 2009, 26, 56–119. [Google Scholar]
- Boidin, J.; Lanquetin, P. Les genres Dichostereum et Vararia en Guadeloupe (Basidiomycetes, Lachnocladiaceae). Mycotaxon 1977, 6, 277–336. [Google Scholar]
- Boidin, J.; Gilles, G.; Lanquetin, P. Basidiomycètes Aphyllophorales de l’Île de la Réunion. IX—Les genres Dichostereum Pilat et Vararia Karsten. Bull. Société Mycol. Fr. 1987, 103, 119–135. [Google Scholar]
- Welden, A.L. West Indian species of Vararia with notes on extralimital species. Mycologia 1965, 57, 502–520. [Google Scholar] [CrossRef]
- Donk, M.A. Notes on resupinate Hymenomycetes IV. Fungus 1957, 27, 1–29. [Google Scholar]
- Karasiński, D. Polish resupinate Russulales: The genus Vararia. Acta Mycol. 2010, 45, 45–56. [Google Scholar] [CrossRef]
- Salcedo, I.; Sarrionandia, E.; Olariaga, I. Contribution to the knowledge of the Aphyllophorales (Basidiomycota) of the Basque Country (Spain). V. Nova Hedwig. 2006, 82, 81–90. [Google Scholar] [CrossRef]
- Núñez, M.; Ryvarden, L. East Asian polypores 2. Synop. Fungorum 2001, 14, 165–522. [Google Scholar]
- Dai, Y.C.; Cui, B.K.; Si, J.; He, S.H.; Hyde, K.D.; Yuan, H.S.; Liu, X.Y.; Zhou, L.W. Dynamics of the worldwide number of fungi with emphasis on fungal diversity in China. Mycol. Prog. 2015, 14, 62. [Google Scholar] [CrossRef]
- Dai, Y.C. Two new polypores from tropical China, and renaming two species in Polyporus and Phellinus. Mycoscience 2012, 53, 40–44. [Google Scholar] [CrossRef]
- Ryvarden, L.; Melo, I. Poroid fungi of Europe. Syn. Fung. 2014, 31, 1–455. [Google Scholar]
- Lin, S.H.; Chen, Z.C. The Corticiaceae and the resupinate Hydnaceae of Taiwan. Taiwania 1990, 35, 69–111. [Google Scholar]
- Wu, S.H. A study of Peniophora species in Taiwan with clamped hyphae. Bot. Bull. Acad. Sin. 2002, 43, 241–250. [Google Scholar]
- Dai, Y.C.; Liu, H.G.; Wu, F.; Cui, B.K.; Si, J.; He, S.H.; Yuan, Y.; Zhou, M.; Zhao, Q.; Liu, S.H.; et al. Resources and Diversity of Wood Decay Fungi in Yunnan, 1st ed.; Science Press: Beijing, China, 2022; pp. 454–455. [Google Scholar]
- Wu, F.; Zhou, L.W.; Vlasák, J.; Dai, Y.C. Global diversity and systematics of Hymenochaetaceae with poroid hymenophore. Fungal Divers. 2022, 113, 1–192. [Google Scholar] [CrossRef]
- Wu, F.; Man, X.W.; Tohtirjap, A.; Dai, Y.C. A comparison of polypore funga and species composition in forest ecosystems of China, North America, and Europe. For. Ecosyst. 2022, 9, 100051. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, L.; Zhang, X.; Deng, Y.; Zhao, C. Four New Wood-Inhabiting Fungal Species of Peniophoraceae (Russulales, Basidiomycota) from the Yunnan-Guizhou Plateau, China. J. Fungi 2022, 8, 1227. https://doi.org/10.3390/jof8111227
Zou L, Zhang X, Deng Y, Zhao C. Four New Wood-Inhabiting Fungal Species of Peniophoraceae (Russulales, Basidiomycota) from the Yunnan-Guizhou Plateau, China. Journal of Fungi. 2022; 8(11):1227. https://doi.org/10.3390/jof8111227
Chicago/Turabian StyleZou, Lei, Xiaolu Zhang, Yinglian Deng, and Changlin Zhao. 2022. "Four New Wood-Inhabiting Fungal Species of Peniophoraceae (Russulales, Basidiomycota) from the Yunnan-Guizhou Plateau, China" Journal of Fungi 8, no. 11: 1227. https://doi.org/10.3390/jof8111227