
Vacuole Proteins with Optimized Microtubule Assembly Is Required for Fum1 Protein Localization and Fumonisin Biosynthesis in Mycotoxigenic Fungus Fusarium verticillioides

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Fum Proteins GFP and mCherry Tagging
2.3. Construction of FvVam7-mCherry and mCherry-FvRab7 Plasmids
2.4. Gene Deletion, FB1 and Split Luciferase Complementation Assay
2.5. Live Cell Imaging and Staining
3. Results
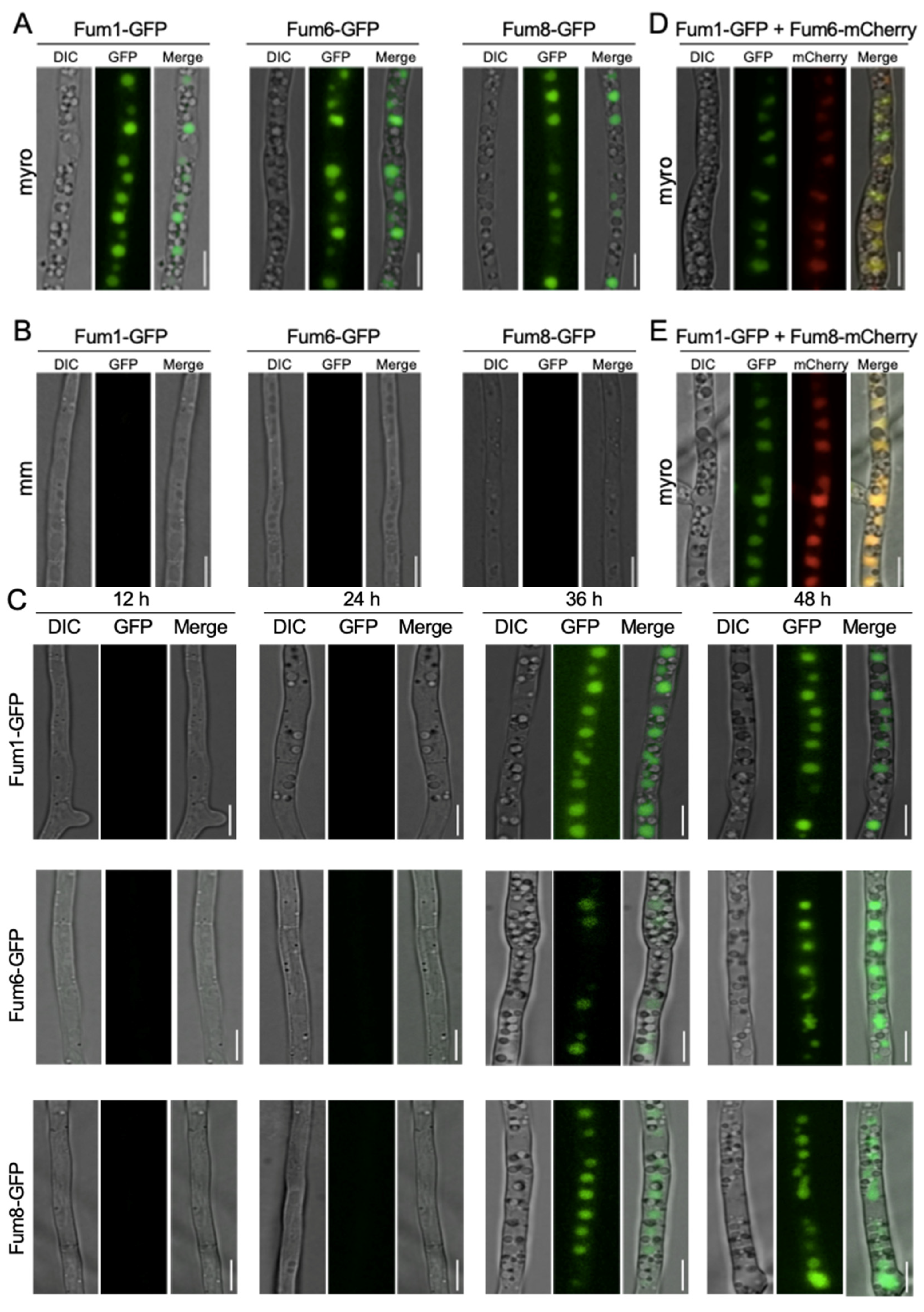
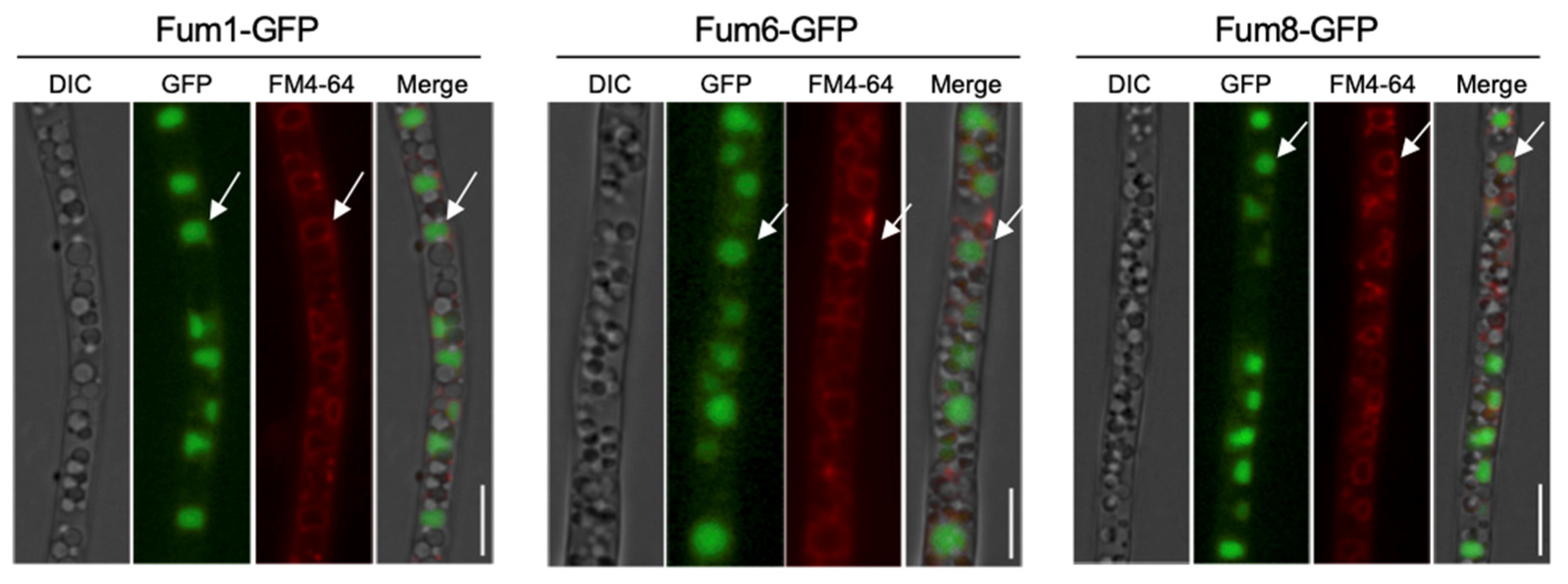
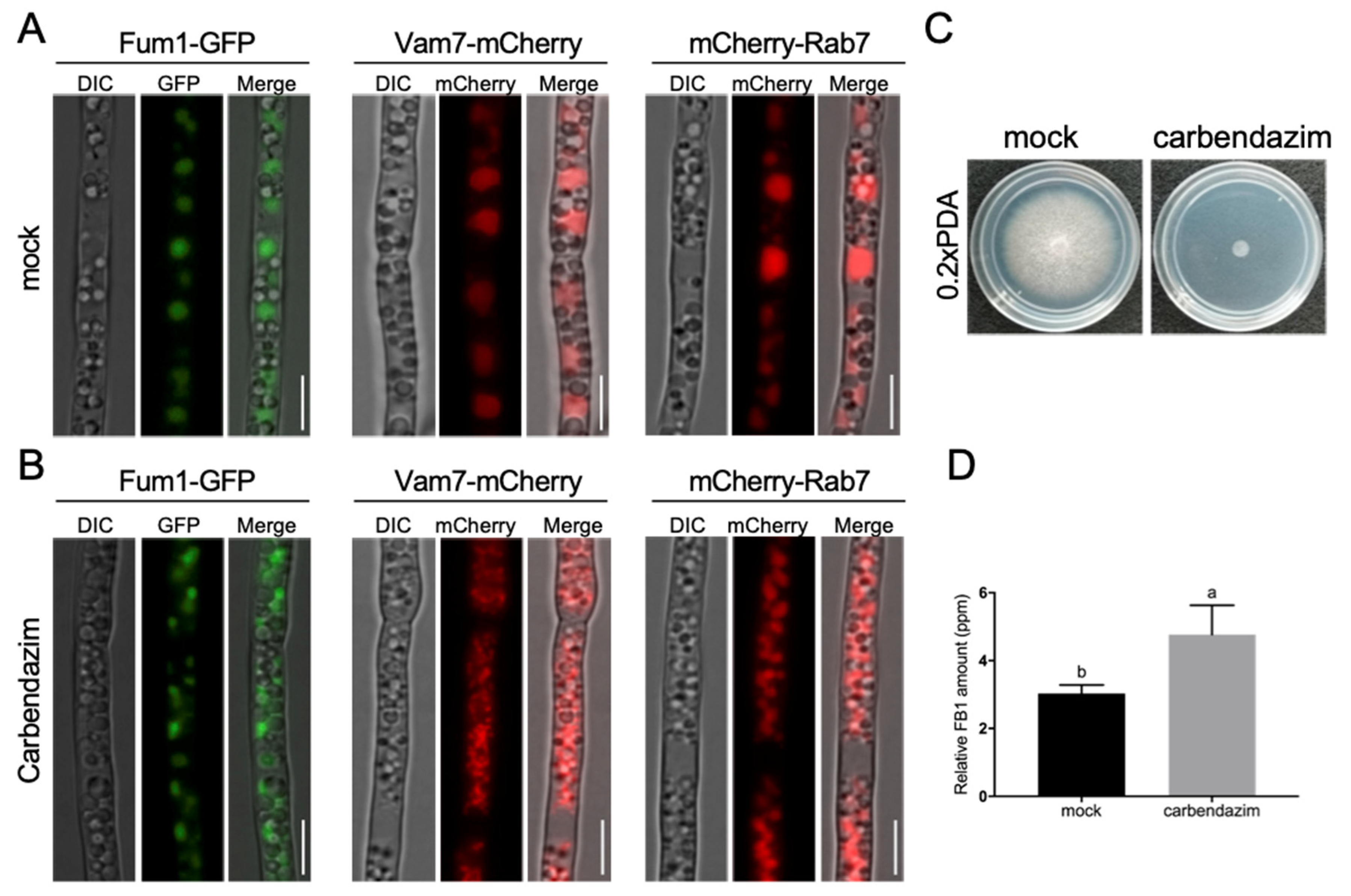
3.1. Subcellular Localization of Three FB1 Biosynthesis Enzymes
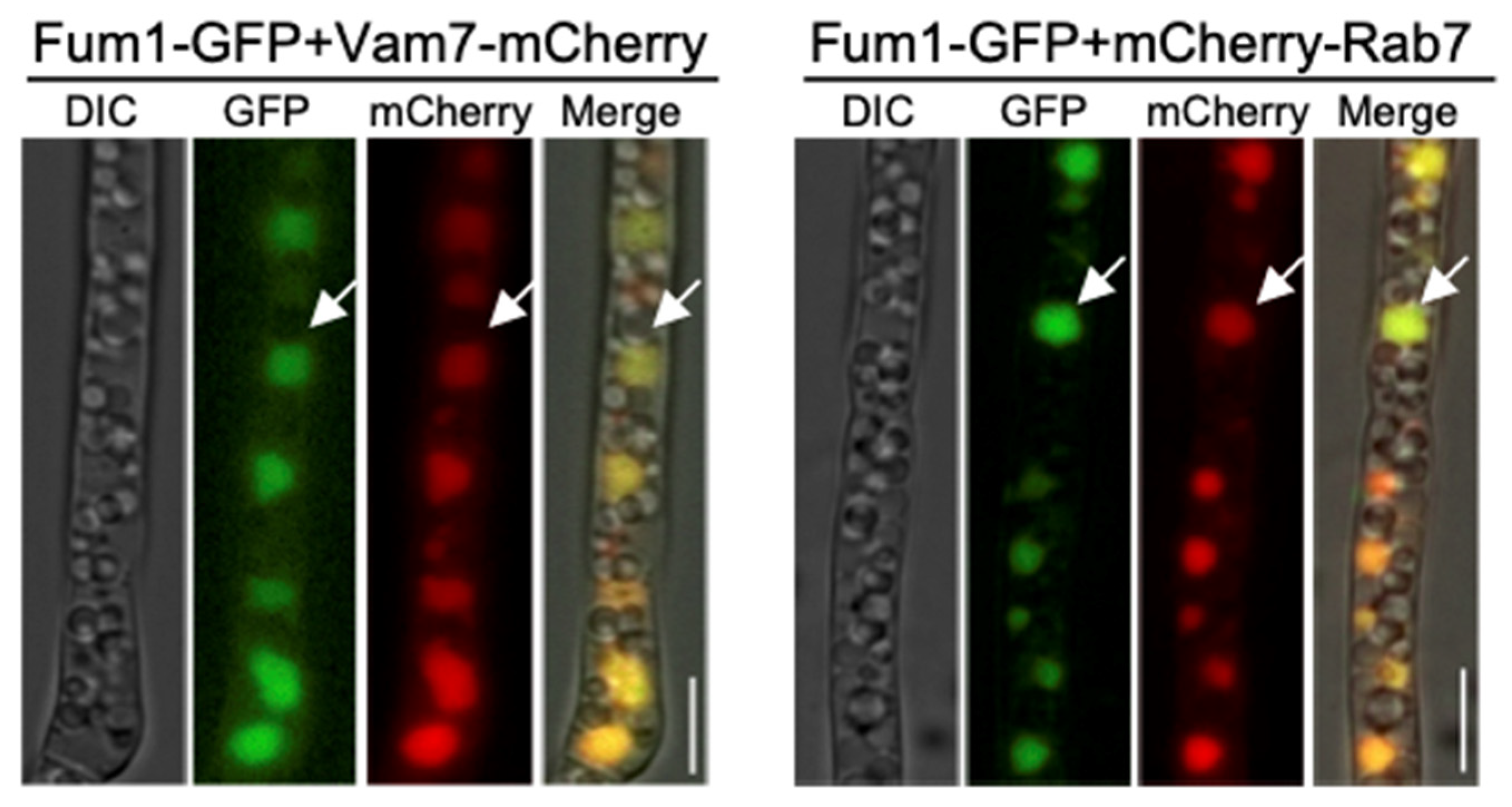
3.2. Co-Localization of Fum1, Fum6, and Fum8 in Vacuoles
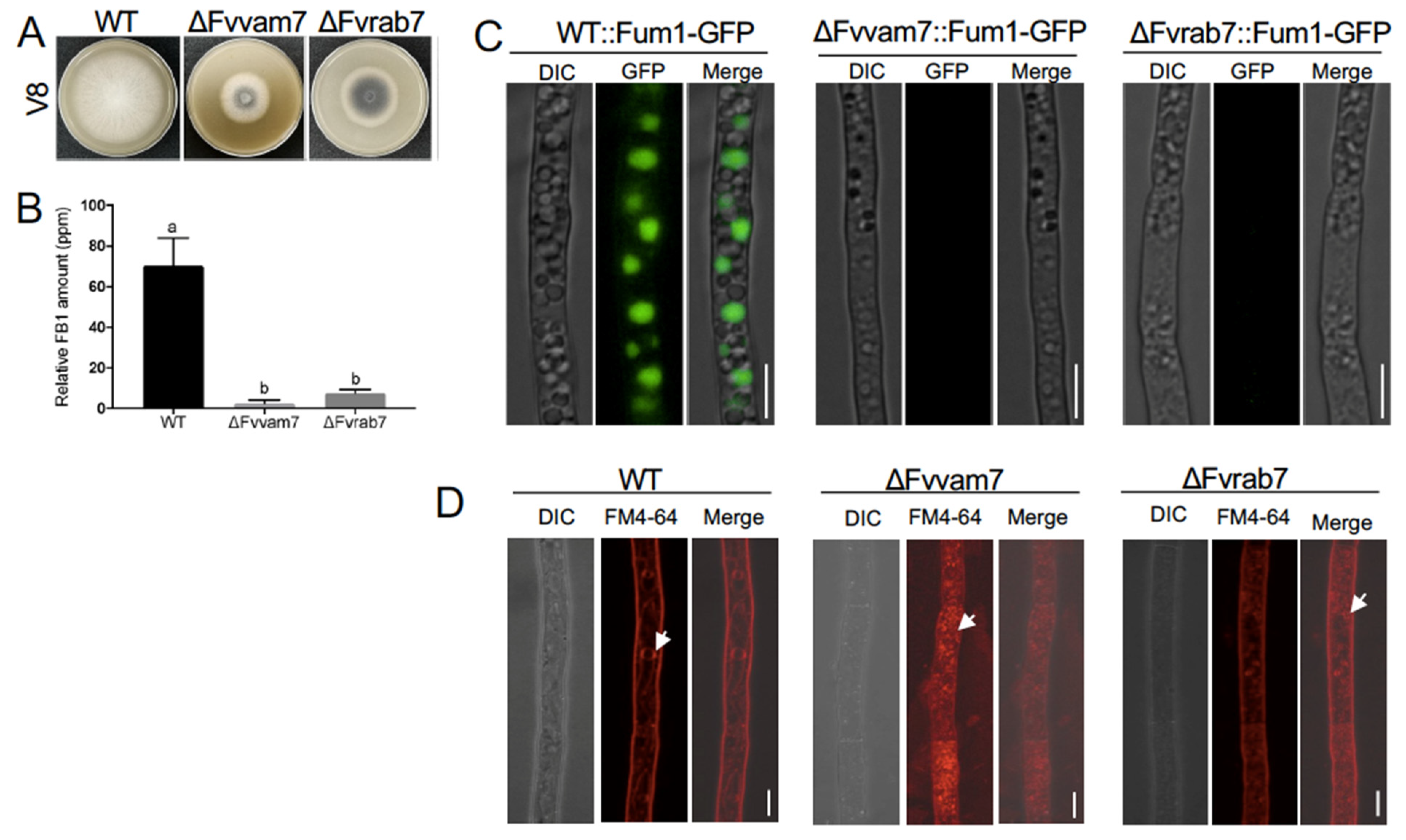
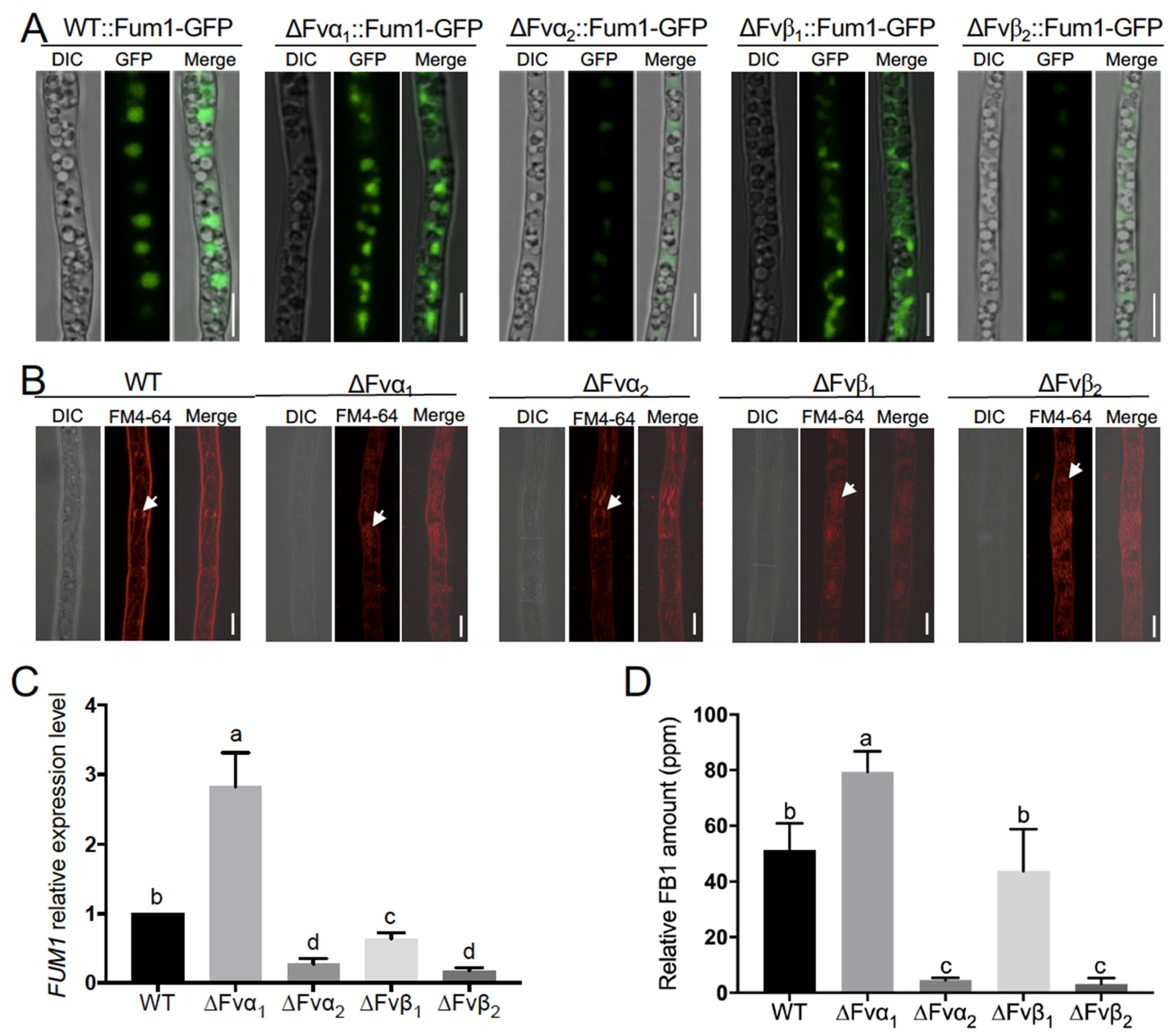
3.3. Two Vacuole Proteins Are Required for FB1 Biosynthesis
3.4. Proper Microtubule Assembly Regulates Vacuole Structure and FB1 Biosynthesis
3.5. Tubulin Fvα1 Is a Negative Regulator in FB1 Biosynthesis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal secondary metabolism—From biochemistry to genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Kistler, H.C.; Broz, K. Cellular compartmentalization of secondary metabolism. Front. Microbiol. 2015, 6, 68. [Google Scholar] [CrossRef] [Green Version]
- Evers, M.E.; Trip, H.; van den Berg, M.A.; Bovenberg, R.A.L.; Driessen, A.J.M. Compartmentalization and transport in beta-lactam antibiotics biosynthesis. In Molecular Biotechnology of Fungal Beta-Lactam Antibiotics and Related Peptide Synthetases; Springer: Berlin/Heidelberg, Germany, 2004; Volume 88, pp. 111–135. [Google Scholar]
- Muller, W.H.; Bovenberg, R.A.L.; Groothuis, M.H.; Kattevilder, F.; Smaal, E.B.; Vandervoort, L.H.M.; Verkleij, A.J. Involvement of Microbodies in Penicillin Biosynthesis. Biochim. Biophys. Acta 1992, 1116, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Kurylowicz, W.; Kurzatkowski, W.; Kurzatkowski, J. Biosynthesis of Benzylpenicillin by Penicillium-Chrysogenum and its Golgi-Apparatus. Arch. Immunol. Ther. Exp. 1987, 35, 699–724. [Google Scholar]
- Lendenfeld, T.; Ghali, D.; Wolschek, M.; Kubicekpranz, E.M.; Kubicek, C.P. Subcellular Compartmentation of Penicillin Biosynthesis in Penicillium-Chrysogenum—The Amino-Acid Precursors Are Derived from the Vacuole. J. Biol. Chem. 1993, 268, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Chanda, A.; Roze, L.V.; Kang, S.; Artymovich, K.A.; Hicks, G.R.; Raikhel, N.V.; Calvo, A.M.; Linz, J.E. A key role for vesicles in fungal secondary metabolism. Proc. Natl. Acad. Sci. USA 2009, 106, 19533–19538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.-Y.; Linz, J.E. Functional expression and subcellular localization of the aflatoxin pathway enzyme Ver-1 fused to enhanced green fluorescent protein. Appl. Environ. Microbiol. 2008, 74, 6385–6396. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.-Y.; Linz, J.E. Functional expression and sub-cellular localization of the early aflatoxin pathway enzyme Nor-1 in Aspergillus parasiticus. Mycol. Res. 2009, 113, 591–601. [Google Scholar] [CrossRef] [Green Version]
- Lee, L.W.; Chiou, C.H.; Klomparens, K.L.; Cary, J.W.; Linz, J.E. Subcellular localization of aflatoxin biosynthetic enzymes Nor-1, Ver-1, and OmtA in time-dependent fractionated colonies of Aspergillus parasiticus. Arch. Microbiol. 2004, 181, 204–214. [Google Scholar] [CrossRef]
- Menke, J.; Weber, J.; Broz, K.; Kistler, H.C. Cellular development associated with induced mycotoxin synthesis in the filamentous fungus Fusarium graminearum. PLoS ONE 2013, 8, e63077. [Google Scholar] [CrossRef] [Green Version]
- Tang, G.; Chen, Y.; Xu, J.R.; Kistler, H.C.; Ma, Z. The fungal myosin I is essential for Fusarium toxisome formation. PLoS Pathog. 2018, 14, e1006827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rheeder, J.P.; Marasas, W.F.; Vismer, H.F. Production of fumonisin analogs by Fusarium species. Appl. Environ. Microbiol. 2002, 68, 2101–2105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelderblom, W.C.; Jaskiewicz, K.; Marasas, W.F.; Thiel, P.G.; Horak, R.M.; Vleggaar, R.; Kriek, N.P. Fumonisins--novel mycotoxins with cancer-promoting activity produced by Fusarium moniliforme. Appl. Environ. Microbiol. 1988, 54, 1806–1811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, P.E.; Desjardins, A.E.; Plattner, R.D. Fumonisins, mycotoxins produced by fusarium species: Biology, chemistry, and significance. Annu. Rev. Phytopathol. 1993, 31, 233–252. [Google Scholar] [CrossRef]
- Proctor, R.H.; Desjardins, A.E.; Plattner, R.D.; Hohn, T.M. A polyketide synthase gene required for biosynthesis of fumonisin mycotoxins in Gibberella fujikuroi mating population A. Fungal Genet. Biol. 1999, 27, 100–112. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.A.; Proctor, R.H.; Plattner, R.D. Characterization of four clustered and coregulated genes associated with fumonisin biosynthesis in Fusarium verticillioides. Fungal Genet. Biol. 2001, 34, 155–165. [Google Scholar] [CrossRef]
- Woloshuk, C.P.; Shim, W.B. Aflatoxins, fumonisins, and trichothecenes: A convergence of knowledge. FEMS Microbiol. Rev. 2013, 37, 94–109. [Google Scholar] [CrossRef] [Green Version]
- Blacutt, A.A.; Gold, S.E.; Voss, K.A.; Gao, M.; Glenn, A.E. Fusarium verticillioides: Advancements in Understanding the Toxicity, Virulence, and Niche Adaptations of a Model Mycotoxigenic Pathogen of Maize. Phytopathology 2018, 108, 312–326. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Huang, J.; Zhang, H.; Shim, W.B. A Rab GTPase protein FvSec4 is necessary for fumonisin B1 biosynthesis and virulence in Fusarium verticillioides. Curr. Genet. 2020, 66, 205–216. [Google Scholar] [CrossRef]
- Yan, H.; Shim, W.B. Characterization of non-canonical G beta-like protein FvGbb2 and its relationship with heterotrimeric G proteins in Fusarium verticillioides. Environ. Microbiol. 2020, 22, 615–628. [Google Scholar] [CrossRef]
- Christensen, S.; Borrego, E.; Shim, W.B.; Isakeit, T.; Kolomiets, M. Quantification of fungal colonization, sporogenesis, and production of mycotoxins using kernel bioassays. J. Vis. Exp 2012, 3727. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.K.; Cho, E.J.; Jo, S.; Sung, B.R.; Lee, S.; Yun, S.H. A split luciferase complementation assay for studying in vivo protein-protein interactions in filamentous ascomycetes. Curr. Genet. 2012, 58, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Alexander, N.J.; Proctor, R.H.; McCormick, S.P. Genes, gene clusters, and biosynthesis of trichothecenes and fumonisins in Fusarium. Toxin Rev. 2009, 28, 198–215. [Google Scholar] [CrossRef]
- Zhang, H.; Yan, H.; Shim, W.B. Fusarium verticillioides SNARE protein FvSyn1 harbours two key functional motifs that play selective roles in fungal development and virulence. Microbiology 2019, 165, 1075–1085. [Google Scholar] [CrossRef]
- Fischer-Parton, S.; Parton, R.M.; Hickey, P.C.; Dijksterhuis, J.; Atkinson, H.A.; Read, N.D. Confocal microscopy of FM4-64 as a tool for analysing endocytosis and vesicle trafficking in living fungal hyphae. J. Microsc. 2000, 198, 246–259. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, B.; Liu, L.; Chen, H.; Zhang, H.; Zheng, X.; Zhang, Z. FgMon1, a guanine nucleotide exchange factor of FgRab7, is important for vacuole fusion, autophagy and plant infection in Fusarium graminearum. Sci. Rep. 2015, 5, 18101. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Li, B.; Fang, Q.; Li, Y.; Zheng, X.; Zhang, Z. SNARE protein FgVam7 controls growth, asexual and sexual development, and plant infection in Fusarium graminearum. Mol. Plant Pathol. 2016, 17, 108–119. [Google Scholar] [CrossRef]
- Oda, Y.; Hirata, A.; Sano, T.; Fujita, T.; Hiwatashi, Y.; Sato, Y.; Kadota, A.; Hasebe, M.; Hasezawa, S. Microtubules regulate dynamic organization of vacuoles in Physcomitrella patens. Plant Cell Physiol. 2009, 50, 855–868. [Google Scholar] [CrossRef] [Green Version]
- Ligrone, R.; Duckett, J.G. The leafy stems of Sphagnum (Bryophyta) contain highly differentiated polarized cells with axial arrays of endoplasmic microtubules. New Phytol. 1998, 140, 567–579. [Google Scholar] [CrossRef]
- Pressel, S.; Ligrone, R.; Duckett, J.G. Cellular differentiation in moss protonemata: A morphological and experimental study. Ann. Bot. 2008, 102, 227–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vedrenne, C.; Hauri, H.P. Morphogenesis of the endoplasmic reticulum: Beyond active membrane expansion. Traffic 2006, 7, 639–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ligrone, R.; Duckett, J.G. Polarity and endoplasmic microtubules in food-conducting cells of mosses: An experimental study. New Phytol. 1996, 134, 503–516. [Google Scholar] [CrossRef]
- Janevska, S.; Ferling, I.; Jojic, K.; Rautschek, J.; Hoefgen, S.; Proctor, R.H.; Hillmann, F.; Valiante, V. Self-Protection against the Sphingolipid Biosynthesis Inhibitor Fumonisin B1 Is Conferred by a FUM Cluster-Encoded Ceramide Synthase. mBio 2020, 11, e00455-20. [Google Scholar] [CrossRef]
- de Brito Francisco, R.; Martinoia, E. The Vacuolar Transportome of Plant Specialized Metabolites. Plant Cell Physiol. 2018, 59, 1326–1336. [Google Scholar] [CrossRef] [Green Version]
- Shim, W.B.; Woloshuk, C.P. Nitrogen repression of fumonisin B1 biosynthesis in Gibberella fujikuroi. FEMS Microbiol. Lett. 1999, 177, 109–116. [Google Scholar] [CrossRef]
- Abenza, J.F.; Galindo, A.; Pinar, M.; Pantazopoulou, A.; de los Rios, V.; Penalva, M.A. Endosomal maturation by Rab conversion in Aspergillus nidulans is coupled to dynein-mediated basipetal movement. Mol. Biol. Cell 2012, 23, 1889–1901. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Zheng, W.; Wu, C.; Yang, J.; Xi, Y.; Xie, Q.; Zhao, X.; Deng, X.; Lu, G.; Li, G.; et al. Rab GTPases are essential for membrane trafficking-dependent growth and pathogenicity in Fusarium graminearum. Environ. Microbiol. 2015, 17, 4580–4599. [Google Scholar] [CrossRef]
- Keller, A.; Peltzer, J.; Carpentier, G.; Horvath, I.; Olah, J.; Duchesnay, A.; Orosz, F.; Ovadi, J. Interactions of enolase isoforms with tubulin and microtubules during myogenesis. Biochim. Biophys. Acta 2007, 1770, 919–926. [Google Scholar] [CrossRef]
- Cassimeris, L.; Silva, V.C.; Miller, E.; Ton, Q.; Molnar, C.; Fong, J. Fueled by microtubules: Does tubulin dimer/polymer partitioning regulate intracellular metabolism? Cytoskeleton 2012, 69, 133–143. [Google Scholar] [CrossRef]
- Durrieu, C.; Bernier-Valentin, F.; Rousset, B. Microtubules bind glyceraldehyde 3-phosphate dehydrogenase and modulate its enzyme activity and quaternary structure. Arch. Biochem. Biophys. 1987, 252, 32–40. [Google Scholar] [CrossRef] [PubMed]
- An, S.; Deng, Y.; Tomsho, J.W.; Kyoung, M.; Benkovic, S.J. Microtubule-assisted mechanism for functional metabolic macromolecular complex formation. Proc. Natl. Acad. Sci. USA 2010, 107, 12872–12876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Duan, Y.; Zhang, J.; Lu, F.; Zhu, Y.; Shim, W.B.; Zhou, M. Microtubule-assisted mechanism for toxisome assembly in Fusarium graminearum. Mol. Plant Pathol. 2021, 22, 163–174. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, H.; Zhou, Z.; Zhang, H.; Shim, W.B. Vacuole Proteins with Optimized Microtubule Assembly Is Required for Fum1 Protein Localization and Fumonisin Biosynthesis in Mycotoxigenic Fungus Fusarium verticillioides. J. Fungi 2023, 9, 268. https://doi.org/10.3390/jof9020268
Yan H, Zhou Z, Zhang H, Shim WB. Vacuole Proteins with Optimized Microtubule Assembly Is Required for Fum1 Protein Localization and Fumonisin Biosynthesis in Mycotoxigenic Fungus Fusarium verticillioides. Journal of Fungi. 2023; 9(2):268. https://doi.org/10.3390/jof9020268
Chicago/Turabian StyleYan, Huijuan, Zehua Zhou, Huan Zhang, and Won Bo Shim. 2023. "Vacuole Proteins with Optimized Microtubule Assembly Is Required for Fum1 Protein Localization and Fumonisin Biosynthesis in Mycotoxigenic Fungus Fusarium verticillioides" Journal of Fungi 9, no. 2: 268. https://doi.org/10.3390/jof9020268