

Renewing Lost Genetic Variability with a Classical Yeast Genetics Approach
 , , , , and
, , , , and
Abstract
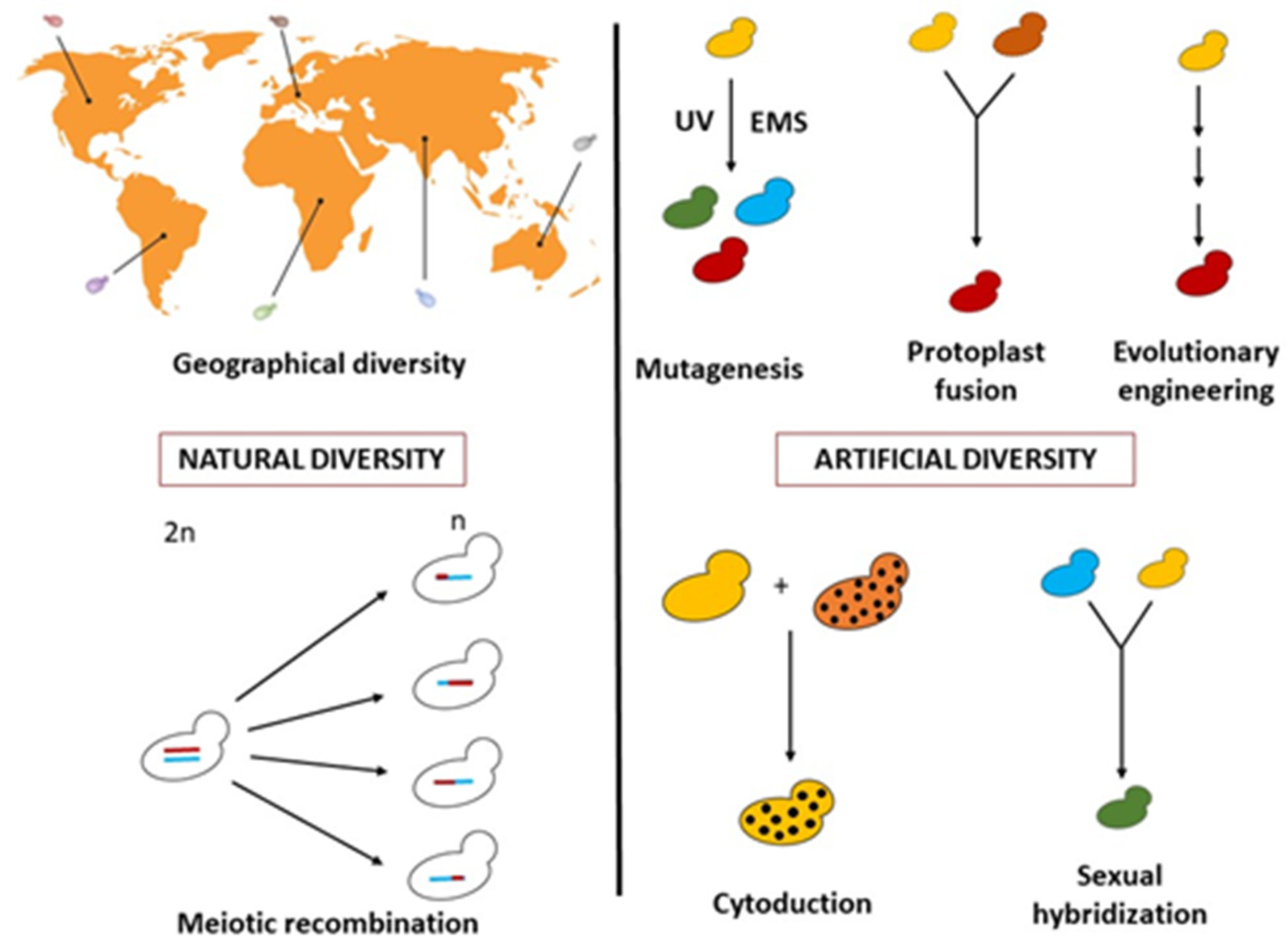
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Growth Conditions
2.2. Spore Production and Sporulation Efficiency
2.3. Screening of Temperature Tolerance of PS and Spores
2.4. Production of Mono-Spore Colonies
2.5. Phenotypic Variation in MSCs
2.6. Metabolomic Fingerprint at the End of Growth
2.7. Metabolomic Fingerprint under Stress
2.8. FTIR Data Analysis
3. Results and Discussion
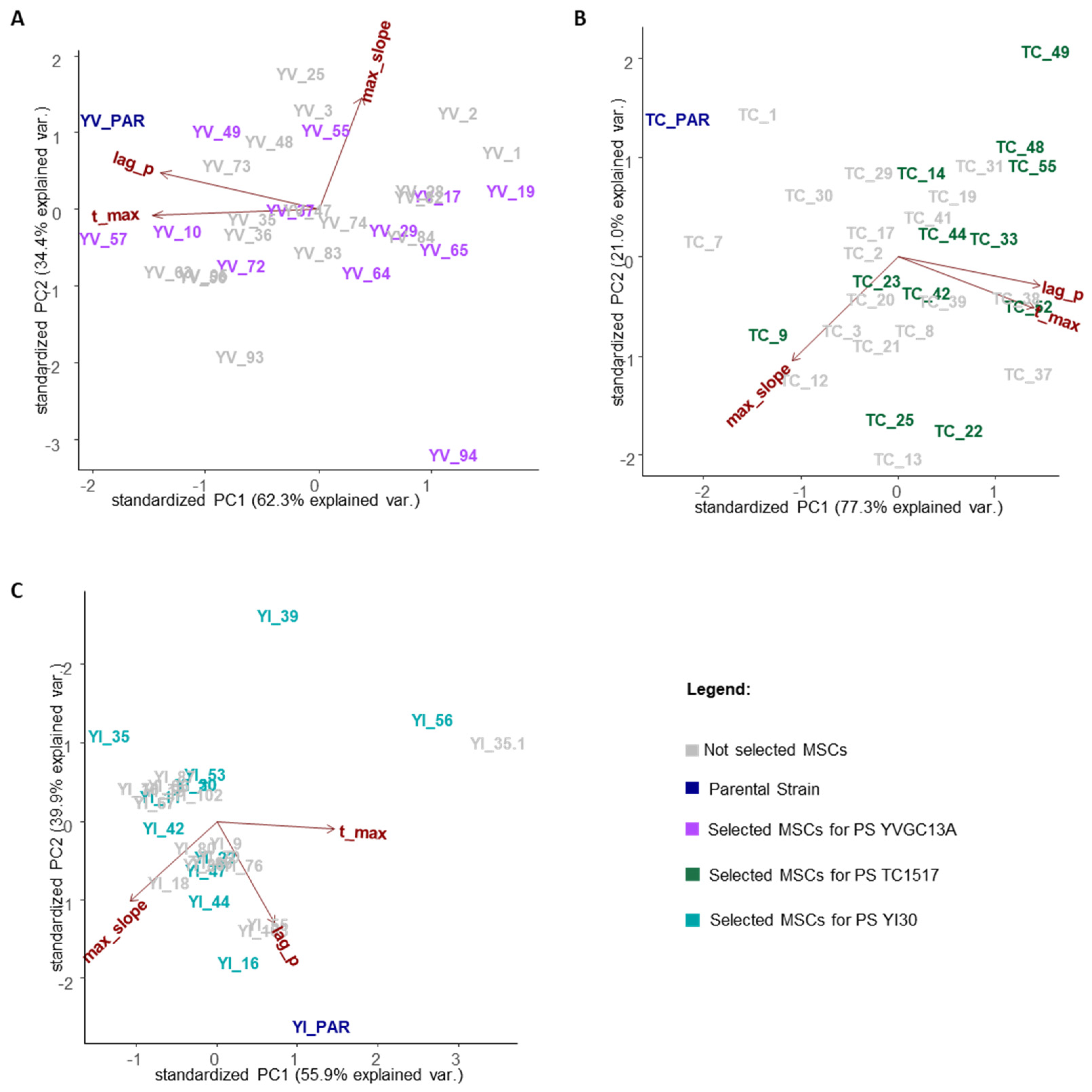
3.1. Efficiency of Sporulation (SE) and Development of an Easy and Effective Protocol for MSCs Production
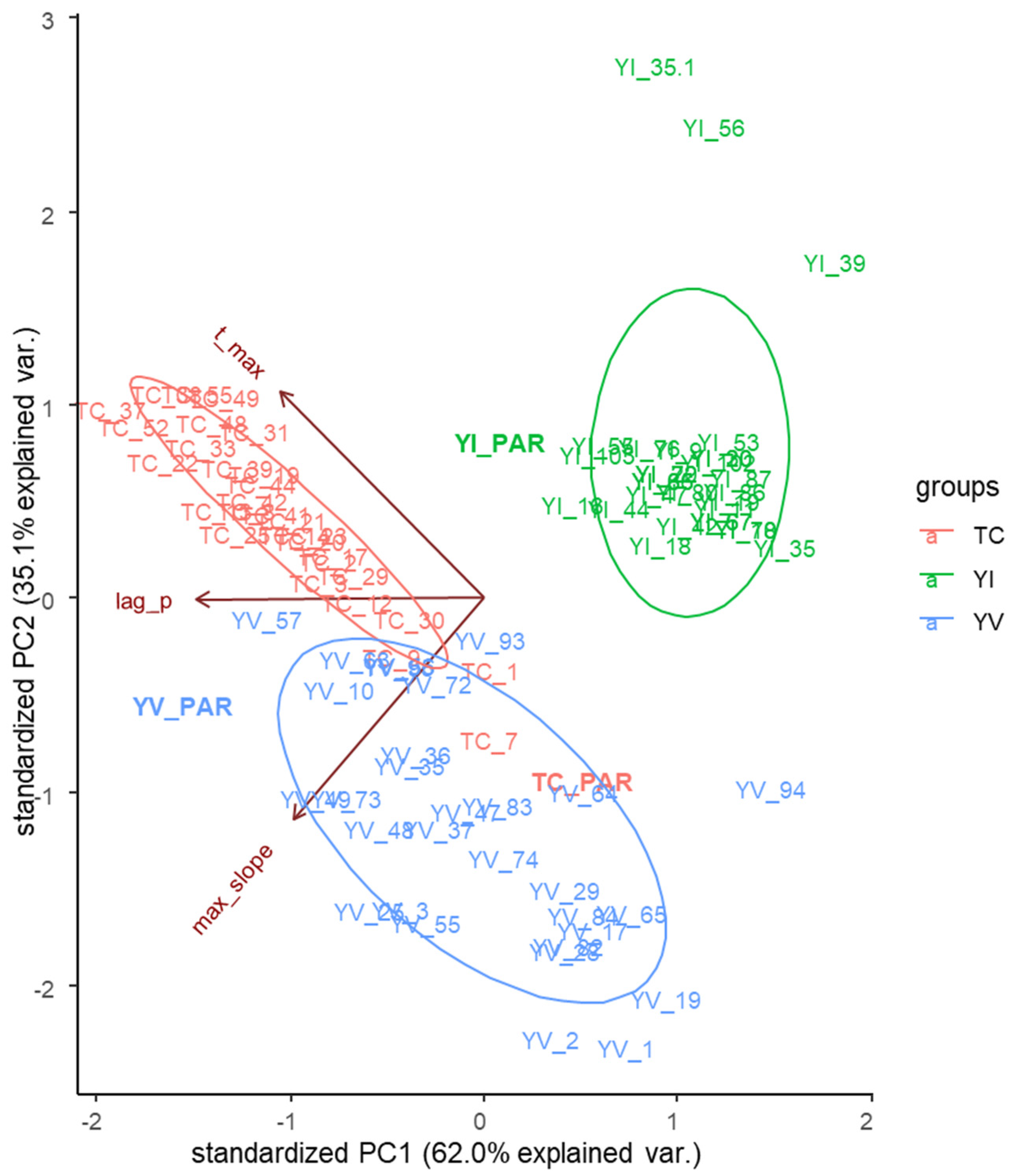
3.2. Growth of MSCs from Each Parental Strain in SD Broth
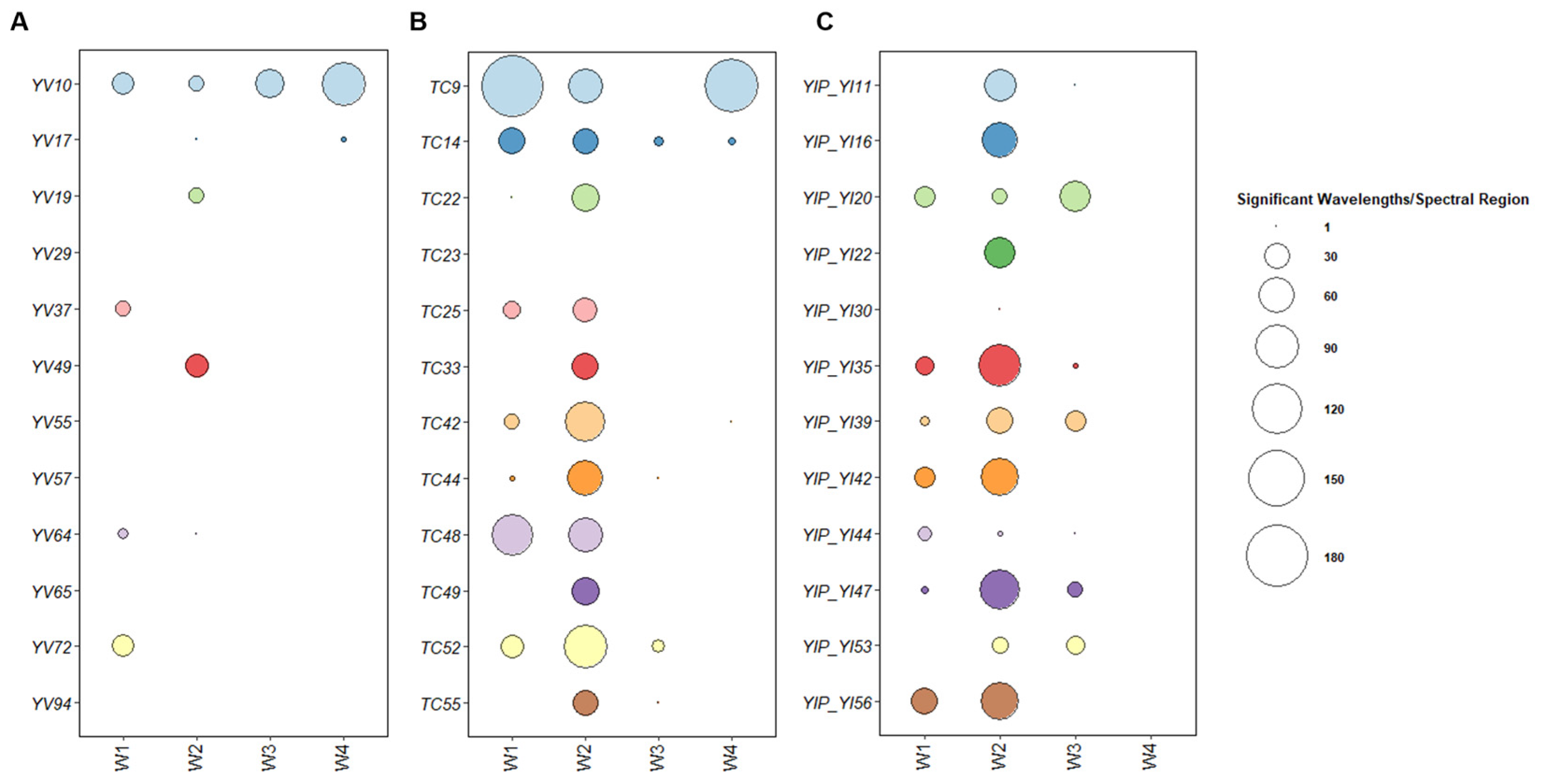
3.3. Metabolomic Fingerprinting of Selected MSCs
3.4. Growth and Metabolomic Phenotypes under Stressing Conditions
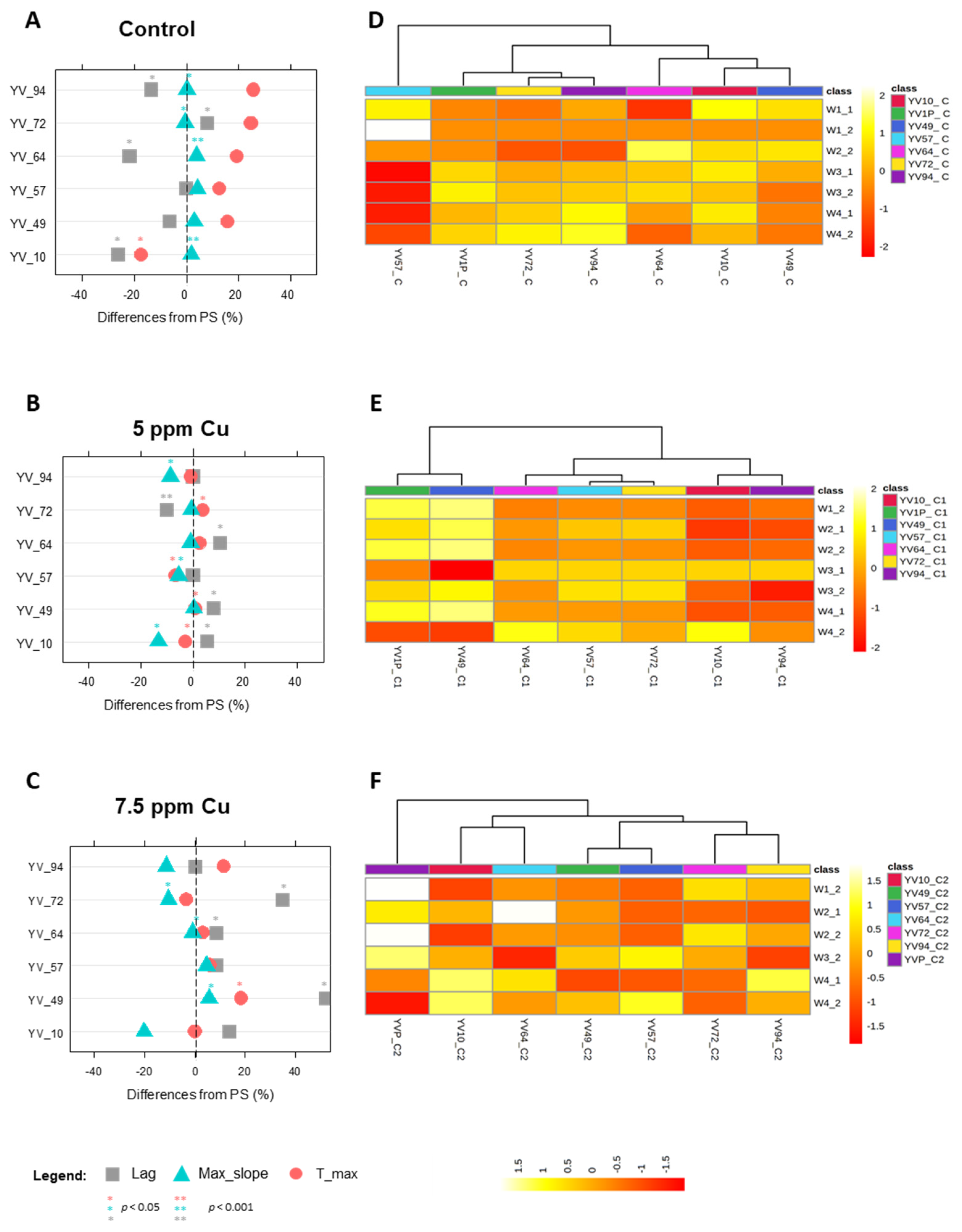
3.4.1. Phenotypes under Copper Sulphate Stress
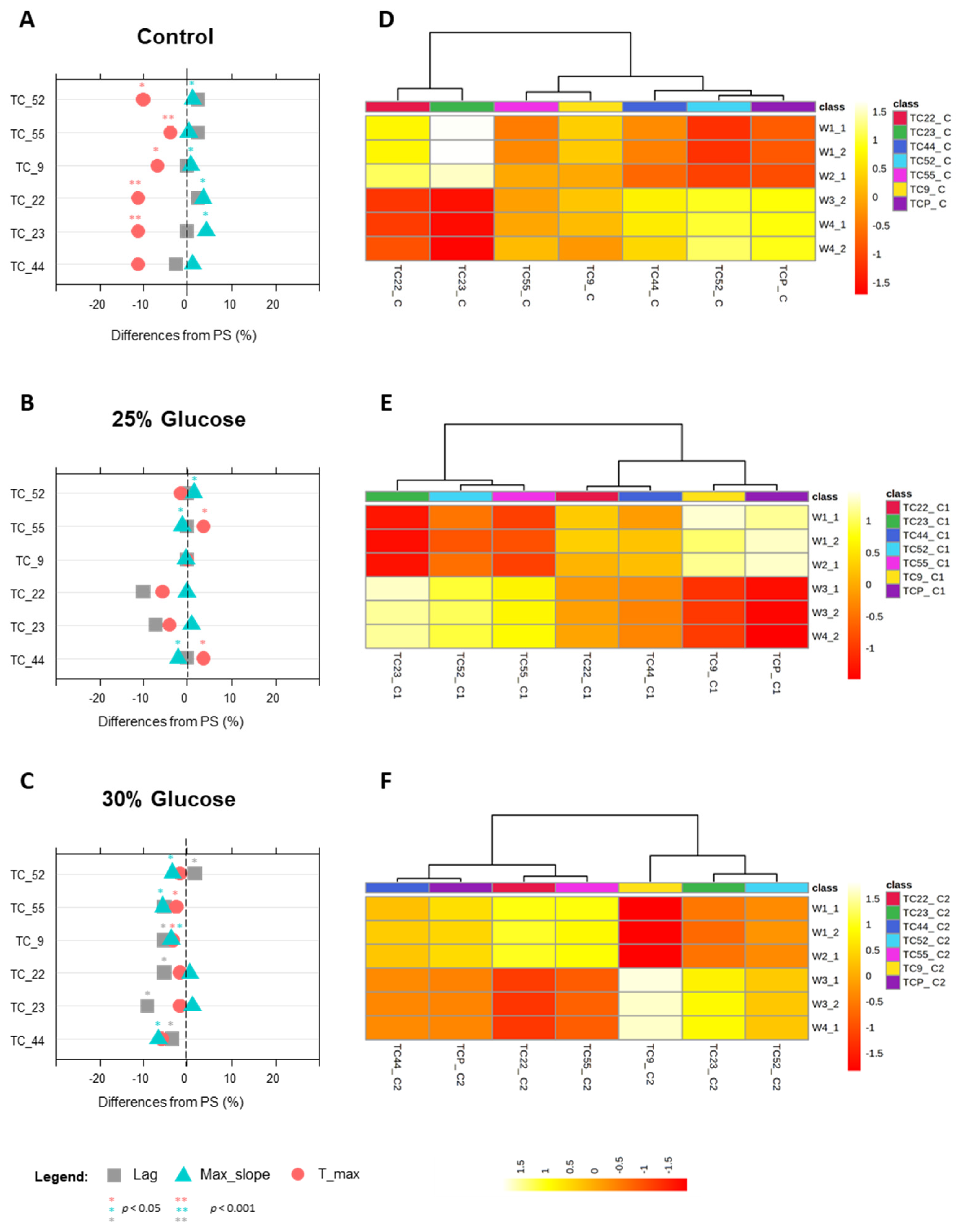
3.4.2. Phenotypes under Glucose Stress
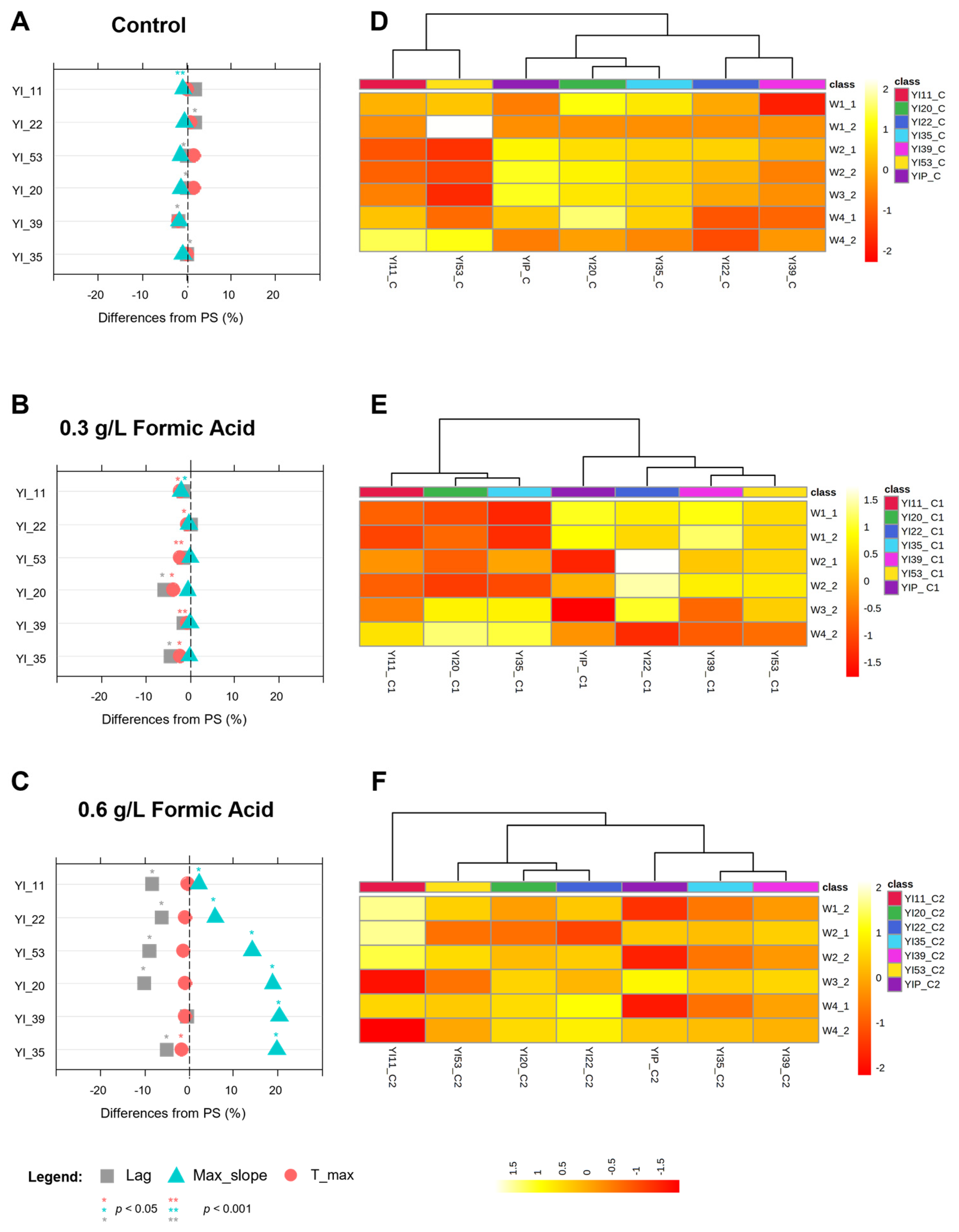
3.4.3. Phenotypes under Formic Acid Stress
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Attfield, P.V. Crucial Aspects of Metabolism and Cell Biology Relating to Industrial Production and Processing of Saccharomyces Biomass. Crit. Rev. Biotechnol. 2022, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Parapouli, M.; Vasileiadis, A.; Afendra, A.S.; Hatziloukas, E. Saccharomyces cerevisiae and its industrial applications. AIMS Microbiol. 2020, 6, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Steensels, J.; Gallone, B.; Voordeckers, K.; Verstrepen, K.J. Domestication of Industrial Microbes. Curr. Biol. 2019, 29, R381–R393. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.F.; Han, P.J.; Wang, Q.M.; Liu, W.Q.; Shi, J.Y.; Li, K.; Zhang, X.L.; Bai, F.Y. The Origin and Adaptive Evolution of Domesticated Populations of Yeast from Far East Asia. Nat. Commun. 2018, 9, 2690. [Google Scholar] [CrossRef] [Green Version]
- Gallone, B.; Steensels, J.; Prahl, T.; Soriaga, L.; Saels, V.; Herrera-Malaver, B.; Merlevede, A.; Roncoroni, M.; Voordeckers, K.; Miraglia, L.; et al. Domestication and Divergence of Saccharomyces cerevisiae Beer Yeasts. Cell 2016, 166, 1397–1410.e16. [Google Scholar] [CrossRef] [Green Version]
- Mozzachiodi, S.; Krogerus, K.; Gibson, B.; Nicolas, A.; Liti, G. Unlocking the Functional Potential of Polyploid Yeasts. Nat. Commun. 2022, 13, 1–13. [Google Scholar] [CrossRef]
- De Chiara, M.; Barré, B.P.; Persson, K.; Irizar, A.; Vischioni, C.; Khaiwal, S.; Stenberg, S.; Amadi, O.C.; Žun, G.; Doberšek, K.; et al. Domestication Reprogrammed the Budding Yeast Life Cycle. Nat. Ecol. Evol. 2022, 6, 448–460. [Google Scholar] [CrossRef]
- Gregory, T.R. Artificial Selection and Domestication: Modern Lessons from Darwin’s Enduring Analogy. Evol. Educ. Outreach 2008, 2, 5–27. [Google Scholar] [CrossRef] [Green Version]
- Zeller, U.; Göttert, T. The Relations between Evolution and Domestication Reconsidered-Implications for Systematics, Ecology, and Nature Conservation. Glob. Ecol. Conserv. 2019, 20, e00756. [Google Scholar] [CrossRef]
- Bigey, F.; Segond, D.; Friedrich, A.; Guezenec, S.; Bourgais, A.; Huyghe, L.; Agier, N.; Nidelet, T.; Sicard, D. Evidence for Two Main Domestication Trajectories in Saccharomyces cerevisiae Linked to Distinct Bread-Making Processes. Curr. Biol. 2021, 31, 722–732.e5. [Google Scholar] [CrossRef]
- Gorter De Vries, A.R.; Pronk, J.T.; Daran, J.M.G. Lager-Brewing Yeasts in the Era of Modern Genetics. FEMS Yeast Res. 2019, 19, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lengeler, K.B.; Stovicek, V.; Fennessy, R.T.; Katz, M.; Förster, J. Never Change a Brewing Yeast? Why Not, There Are Plenty to Choose from. Front. Genet. 2020, 11, 582789. [Google Scholar] [CrossRef] [PubMed]
- Favaro, L.; Jansen, T.; van Zyl, W.H. Exploring Industrial and Natural Saccharomyces cerevisiae Strains for the Bio-Based Economy from Biomass: The Case of Bioethanol. Crit. Rev. Biotechnol. 2019, 39, 800–816. [Google Scholar] [CrossRef] [PubMed]
- Legras, J.L.; Merdinoglu, D.; Cornuet, J.M.; Karst, F. Bread, Beer and Wine: Saccharomyces cerevisiae Diversity Reflects Human History. Mol. Ecol. 2007, 16, 2091–2102. [Google Scholar] [CrossRef] [PubMed]
- Gronchi, N.; Favaro, L.; Cagnin, L.; Brojanigo, S.; Pizzocchero, V.; Basaglia, M.; Casella, S. Novel Yeast Strains for the Efficient Saccharification and Fermentation of Starchy By-Products to Bioethanol. Energies 2019, 12, 714. [Google Scholar] [CrossRef] [Green Version]
- Saerens, S.M.G.; Duong, C.T.; Nevoigt, E. Genetic Improvement of Brewer’s Yeast: Current State, Perspectives and Limits. Appl. Microbiol. Biotechnol. 2010, 86, 1195–1212. [Google Scholar] [CrossRef]
- Stewart, G.G.; Hill, A.E.; Russell, I. 125th Anniversary Review: Developments in Brewing and Distilling Yeast Strains. J. Inst. Brew. 2013, 119, 202–220. [Google Scholar] [CrossRef]
- Steensels, J.; Snoek, T.; Meersman, E.; Nicolino, M.P.; Voordeckers, K.; Verstrepen, K.J. Improving Industrial Yeast Strains: Exploiting Natural and Artificial Diversity. FEMS Microbiol. Rev. 2014, 38, 947–995. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.M.; Liu, W.Q.; Liti, G.; Wang, S.A.; Bai, F.Y. Surprisingly Diverged Populations of Saccharomyces cerevisiae in Natural Environments Remote from Human Activity. Mol. Ecol. 2012, 21, 5404–5417. [Google Scholar] [CrossRef]
- Liti, G.; Carter, D.M.; Moses, A.M.; Warringer, J.; Parts, L.; James, S.A.; Davey, R.P.; Roberts, I.N.; Burt, A.; Koufopanou, V.; et al. Population Genomics of Domestic and Wild Yeasts. Nature 2009, 458, 337–341. [Google Scholar] [CrossRef] [Green Version]
- García-Ríos, E.; Guillamón, J.M. Genomic Adaptations of Saccharomyces Genus to Wine Niche. Microorganisms 2022, 10, 1811. [Google Scholar] [CrossRef] [PubMed]
- Seike, T.; Sakata, N.; Matsuda, F.; Furusawa, C. Elevated Sporulation Efficiency in Fission Yeast Schizosaccharomyces japonicus Strains Isolated from Drosophila. J. Fungi 2021, 7, 350. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, R.K.; Romano, P.; Suzzi, G.; Polsinelli, M. Genome Renewal: A New Phenomenon Revealed from a Genetic Study of 43 Strains of Saccharomyces cerevisiae Derived from Natural Fermentation of Grape Musts. Yeast 1994, 10, 1543–1552. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Shi, G.; Chen, M.; Chen, G.; Wu, Q. Using Polygenic Risk Scores Related to Complex Traits to Predict Production Performance in Cross-Breeding of Yeast. J. Fungi 2022, 8, 914. [Google Scholar] [CrossRef] [PubMed]
- Codon, A.C.; Gasent-Ramirez, J.M.; Benitez, T. Factors Which Affect the Frequency of Sporulation and Tetrad Formation in Saccharomyces cerevisiae Baker’s Yeasts. Appl. Environ. Microbiol. 1995, 61, 630–638. [Google Scholar] [CrossRef] [Green Version]
- Favaro, L.; Basaglia, M.; Trento, A.; Van Rensburg, E.; García-Aparicio, M.; Van Zyl, W.H.; Casella, S. Exploring Grape Marc as Trove for New Thermotolerant and Inhibitor-Tolerant Saccharomyces cerevisiae Strains for Second-Generation Bioethanol Production. Biotechnol. Biofuels 2013, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Jansen, T.; Hoff, J.W.; Jolly, N.; van Zyl, W.H. Mating of Natural Saccharomyces cerevisiae Strains for Improved Glucose Fermentation and Lignocellulosic Inhibitor Tolerance. Folia Microbiol. 2018, 63, 155–168. [Google Scholar] [CrossRef]
- Morrison-Whittle, P.; Goddard, M.R. From Vineyard to Winery: A Source Map of Microbial Diversity Driving Wine Fermentation. Environ. Microbiol. 2018, 20, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Budroni, M.; Giordano, G.; Pinna, G.; Farris, G.A. A Genetic Study of Natural Flor Strains of Saccharomyces cerevisiae Isolated during Biological Ageing from Sardinian Wines. J. Appl. Microbiol. 2000, 89, 657–662. [Google Scholar] [CrossRef]
- Milani, E.A.; Gardner, R.C.; Silva, F.V.M. Thermal Resistance of Saccharomyces Yeast Ascospores in Beers. Int. J. Food Microbiol. 2015, 206, 75–80. [Google Scholar] [CrossRef]
- Bahalul, M.; Kaneti, G.; Kashi, Y. Ether-Zymolyase Ascospore Isolation Procedure: An Efficient Protocol for Ascospores Isolation in Saccharomyces cerevisiae Yeast. Yeast 2010, 27, 999–1003. [Google Scholar] [CrossRef] [PubMed]
- Kamrad, S.; Rodríguez-López, M.; Cotobal, C.; Correia-Melo, C.; Ralser, M.; Bähler, J. Pyphe, a Python Toolbox for Assessing Microbial Growth and Cell Viability in High-Throughput Colony Screens. Elife 2020, 9, e55160. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical. R Found. Comput. 2022. Available online: https://www.R-project.org/ (accessed on 27 December 2022).
- Essendoubi, M.; Toubas, D.; Bouzaggou, M.; Pinon, J.M.; Manfait, M.; Sockalingum, G.D. Rapid Identification of Candida Species by FT-IR Microspectroscopy. Biochim. Biophys. Acta-Gen. Subj. 2005, 1724, 239–247. [Google Scholar] [CrossRef]
- Corte, L.; Roscini, L.; Pierantoni, D.C.; Pellegrino, R.M.; Emiliani, C.; Basaglia, M.; Favaro, L.; Casella, S.; Cardinali, G. Delta-Integration of Single Gene Shapes the Whole Metabolomic Short-Term Response to Ethanol of Recombinant Saccharomyces cerevisiae Strains. Metabolites 2020, 10, 140. [Google Scholar] [CrossRef] [Green Version]
- Pang, Z.; Chong, J.; Zhou, G.; De Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the Gap between Raw Spectra and Functional Insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Beuchat, L.R. Combined Effect of Growth Medium, Age of Cells and Phase of Sporulation on Heat Resistance and Recovery of Hansenula anomala. Mycopathologia 1984, 87, 129–134. [Google Scholar] [CrossRef]
- McClary, D.O.; Nulty, W.L.; Miller, G.R. Effect of Potassium Versus Sodium in the Sporulation of Saccharomyces. J. Bacteriol. 1959, 78, 362–368. [Google Scholar] [CrossRef] [Green Version]
- Neiman, A.M. Ascospore Formation in the Yeast Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2005, 69, 565–584. [Google Scholar] [CrossRef] [Green Version]
- Petersen, J.G.L.; Olson, L.W.; Zickler, D. Synchronous Sporulation of Saccharomyces cerevisiae. Carlsb. Res. Commun. 1978, 43, 241–253. [Google Scholar] [CrossRef] [Green Version]
- Tremaine, J.H.; Miller, J.J. Effect of Yeast Extract, Peptone, and Certain Nitrogen Compounds on Sporulation of Saccharomyces cerevisiae. Mycopathol. Mycol. Appl. 1956, 7, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Dawes, I.W.; Hardie, I.D. Selective Killing of Vegetative Cells in Sporulated Yeast Cultures by Exposure to Diethyl Ether. MGG Mol. Gen. Genet. 1974, 131, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Bilinski, C.; Russell, I.; Stewart, G.G. Analysis of Sporulation in Brewer’s Yeast: Induction of Tetrad Formation. J. Inst. Brew. 1986, 92, 594–598. [Google Scholar] [CrossRef]
- Rachon, G.; Raleigh, C.P.; Pawlowsky, K. Heat Resistance of Yeast Ascospores and Their Utilisation for the Validation of Pasteurisation Processes for Beers. J. Inst. Brew. 2021, 127, 149–159. [Google Scholar] [CrossRef]
- Camarasa, C.; Sanchez, I.; Brial, P.; Bigey, F.; Dequin, S. Phenotypic Landscape of Saccharomyces cerevisiae during Wine Fermentation: Evidence for Origin-Dependent Metabolic Traits. PLoS ONE 2011, 6, e25147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitanovic, E. Use of cu fungicides in vineyards and olive groves. In Fungicides for Plant and Animal Diseases; Dhanasekaran, D., Thajuddin, N., Panneerselvam, A., Eds.; IntechOpen: Rijeka, Croatia, 2012; p. Ch. 14. [Google Scholar]
- Xie, D.; Sun, Y.; Lei, Y. Effect of Glucose Levels on Carbon Flow Rate, Antioxidant Status, and Enzyme Activity of Yeast during Fermentation. J. Sci. Food Agric. 2022, 102, 5333–5347. [Google Scholar] [CrossRef]
- Cagnin, L.; Gronchi, N.; Basaglia, M.; Favaro, L.; Casella, S. Selection of Superior Yeast Strains for the Fermentation of Lignocellulosic Steam-Exploded Residues. Front. Microbiol. 2021, 12, 1–12. [Google Scholar] [CrossRef]
- Parrou, J.L.; Teste, M.A.; François, J. Effects of Various Types of Stress on the Metabolism of Reserve Carbohydrates in Saccharomyces cerevisiae: Genetic Evidence for a Stress-Induced Recycling of Glycogen and Trehalose. Microbiology 1997, 143, 1891–1900. [Google Scholar] [CrossRef] [Green Version]
- Adamo, G.M.; Brocca, S.; Passolunghi, S.; Salvato, B.; Lotti, M. Laboratory Evolution of Copper Tolerant Yeast Strains. Microb. Cell Fact. 2012, 11, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Grosfeld, E.V.; Bidiuk, V.A.; Mitkevich, O.V.; Ghazy, E.S.M.O.; Kushnirov, V.V.; Alexandrov, A.I. A Systematic Survey of Characteristic Features of Yeast Cell Death Triggered by External Factors. J. Fungi 2021, 7, 886. [Google Scholar] [CrossRef]
- Favaro, L.; Corte, L.; Roscini, L.; Cagnin, L.; Tiecco, M.; Colabella, C.; Berti, A.; Basaglia, M.; Cardinali, G.; Casella, S. A Novel FTIR-Based Approach to Evaluate the Interactions between Lignocellulosic Inhibitory Compounds and Their Effect on Yeast Metabolism. RSC Adv. 2016, 6, 47981–47989. [Google Scholar] [CrossRef]
- Corte, L.; Tiecco, M.; Roscini, L.; De Vincenzi, S.; Colabella, C.; Germani, R.; Tascini, C.; Cardinali, G. FTIR Metabolomic Fingerprint Reveals Different Modes of Action Exerted by Structural Variants of N-Alkyltropinium Bromide Surfactants on Escherichia coli and Listeria innocua Cells. PLoS ONE 2015, 10, e0115275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corte, L.; Roscini, L.; Zadra, C.; Antonielli, L.; Tancini, B.; Magini, A.; Emiliani, C.; Cardinali, G. Effect of pH on Potassium Metabisulphite Biocidic Activity against Yeast and Human Cell Cultures. Food Chem. 2012, 134, 1327–1336. [Google Scholar] [CrossRef] [PubMed]
- Benbadis, L.; Cot, M.; Rigoulet, M.; Francois, J. Isolation of Two Cell Populations from Yeast during High-Level Alcoholic Fermentation That Resemble Quiescent and Nonquiescent Cells from the Stationary Phase on Glucose. FEMS Yeast Res. 2009, 9, 1172–1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holman, H.Y.N.; Wozei, E.; Lin, Z.; Comolli, L.R.; Ball, D.A.; Borglin, S.; Fields, M.W.; Hazen, T.C.; Downing, K.H. Real-Time Molecular Monitoring of Chemical Environment in Obligate Anaerobes during Oxygen Adaptive Response. Proc. Natl. Acad. Sci. USA 2009, 106, 12599–12604. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Xie, C.Y.; Yang, B.X.; Gou, M.; Xia, Z.Y.; Sun, Z.Y.; Tang, Y.Q. The Response Mechanisms of Industrial Saccharomyces cerevisiae to Acetic Acid and Formic Acid during Mixed Glucose and Xylose Fermentation. Process Biochem. 2020, 91, 319–329. [Google Scholar] [CrossRef]
- Du, C.; Li, Y.; Xiang, R.; Yuan, W. Formate Dehydrogenase Improves the Resistance to Formic Acid and Acetic Acid Simultaneously in Saccharomyces cerevisiae. Int. J. Mol. Sci. 2022, 23, 3406. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Background | Geographical Location | Genotype | Reference | Stressing Agent | Low Stress | High Stress |
|---|---|---|---|---|---|---|---|
| TC1517 | Grape marcs | Italy | 2n, homotallic | [26] | Glucose (g L−1) | 250 | 300 |
| YI30 | Industrial distillery | South Africa | 2n, homotallic | [27] | Formic acid (g L−1) | 0.3 | 0.6 |
| YVGC13A | Vineyard, isolated from vine bark | Canada | 2n, homotallic | University of Perugia | Copper sulfate (Cu-ppm) | 5 | 7.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gupte, A.P.; Pierantoni, D.C.; Conti, A.; Donati, L.; Basaglia, M.; Casella, S.; Favaro, L.; Corte, L.; Cardinali, G. Renewing Lost Genetic Variability with a Classical Yeast Genetics Approach. J. Fungi 2023, 9, 264. https://doi.org/10.3390/jof9020264
Gupte AP, Pierantoni DC, Conti A, Donati L, Basaglia M, Casella S, Favaro L, Corte L, Cardinali G. Renewing Lost Genetic Variability with a Classical Yeast Genetics Approach. Journal of Fungi. 2023; 9(2):264. https://doi.org/10.3390/jof9020264
Chicago/Turabian StyleGupte, Ameya Pankaj, Debora Casagrande Pierantoni, Angela Conti, Leonardo Donati, Marina Basaglia, Sergio Casella, Lorenzo Favaro, Laura Corte, and Gianluigi Cardinali. 2023. "Renewing Lost Genetic Variability with a Classical Yeast Genetics Approach" Journal of Fungi 9, no. 2: 264. https://doi.org/10.3390/jof9020264