
Genotypic Analysis of the Population Structure in Malassezia globosa and Malassezia restricta

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolates
2.2. STR Design
2.3. DNA Extraction
2.4. PCR Amplification of STR Markers
2.5. Data Analysis
3. Results
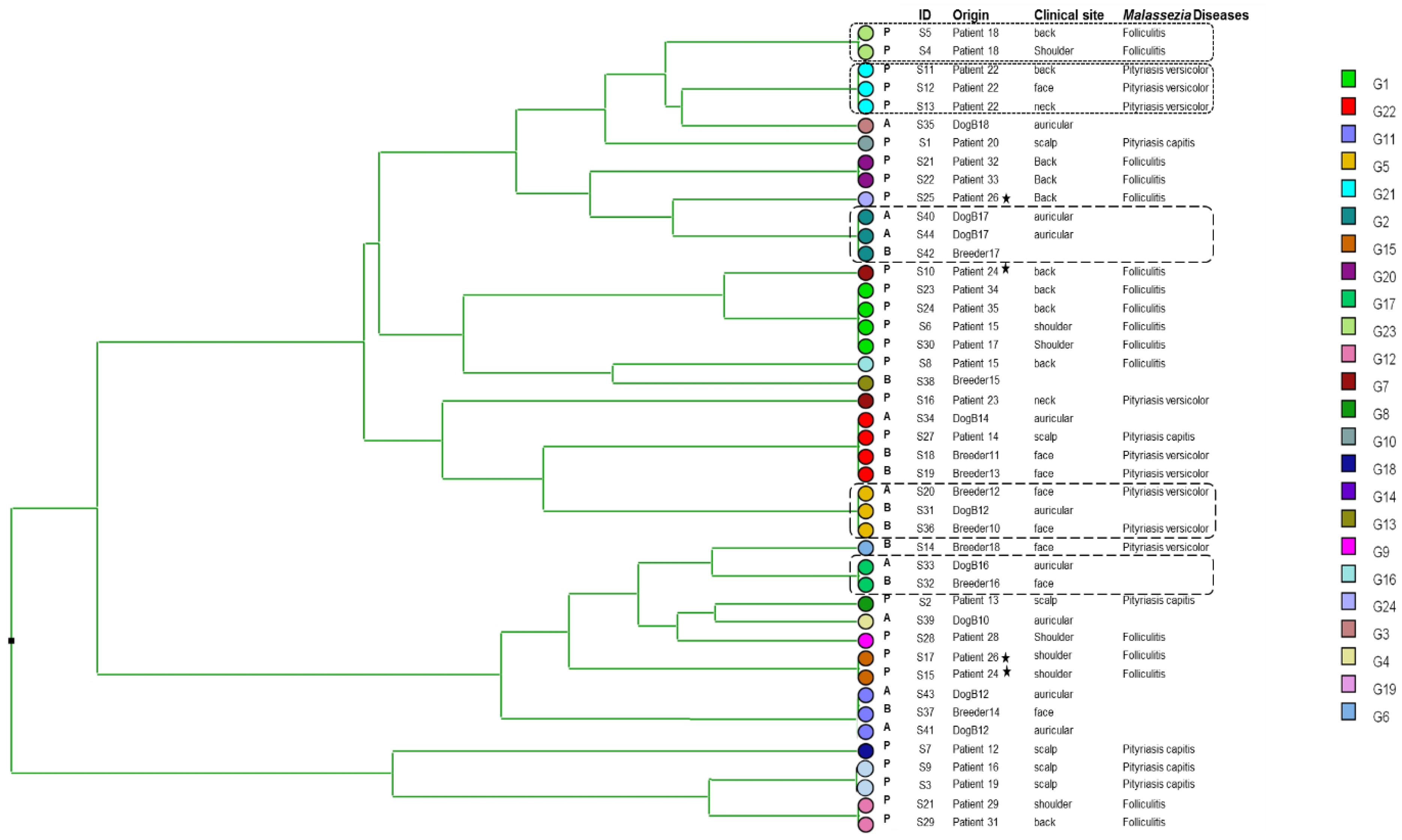
3.1. Genetic Diversity
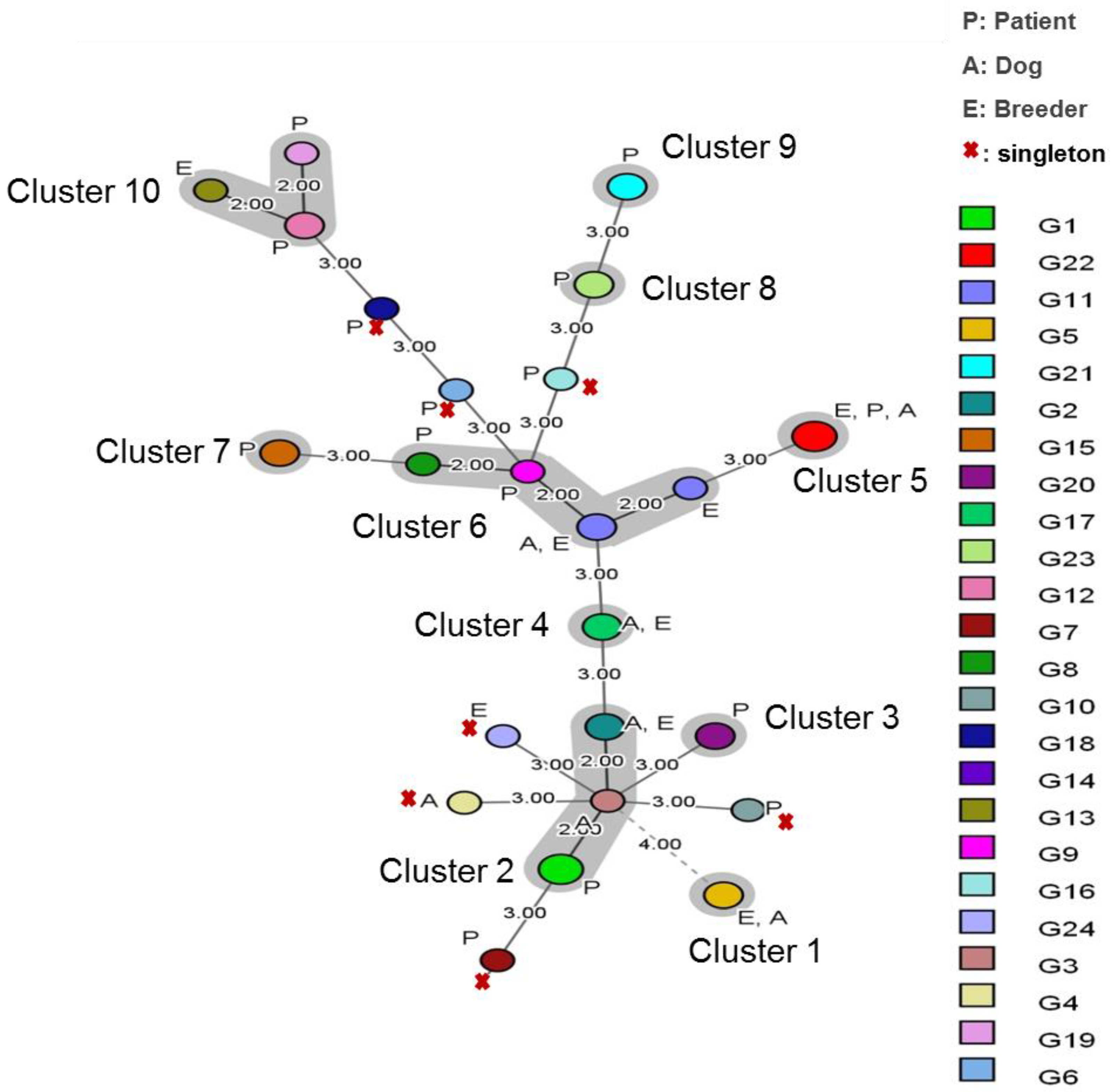
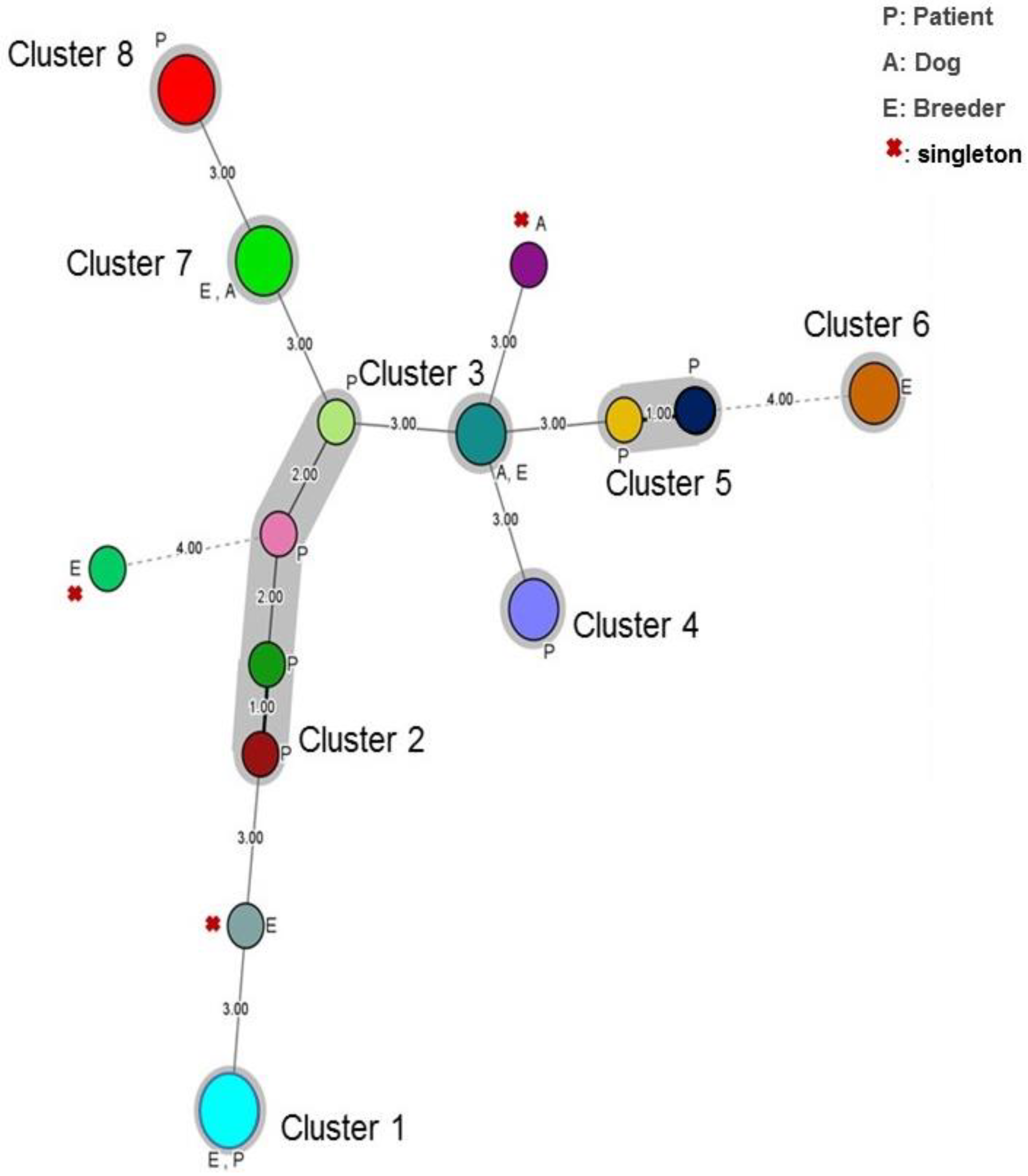
3.2. Cluster Analysis
3.3. Interpopulation Genetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sugita, T.; Nishikawa, A. Molecular and quantitative analyses of Malassezia microflora on the skin of atopic dermatitis patients and genotyping of M. globosa DNA. Nihon Ishinkin Gakkai Zasshi 2003, 44, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Sugita, T.; Takashima, M.; Kodama, M.; Tsuboi, R.; Nishikawa, A. Description of a new yeast species, Malassezia japonica, and its detection in patients with atopic dermatitis and healthy subjects. J. Clin. Microbiol. 2003, 41, 4695–4699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theelen, B.; Cafarchia, C.; Gaitanis, G.; Bassukas, I.D.; Boekhout, T.; Dawson, T.L. Corrigendum: Malassezia ecology, pathophysiology, and treatment. Med. Mycol. 2019, 57, e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prohic, A.; Jovovic Sadikovic, T.; Kuskunovic-Vlahovljak, S.; Baljic, R. Distribution of Malassezia Species in Patients with Different Dermatological Disorders and Healthy Individuals. Acta Dermatovenerol. Croat. 2016, 24, 274–281. [Google Scholar]
- Gupta, A.K.; Batra, R.; Bluhm, R.; Boekhout, T.; Dawson, T.L., Jr. Skin diseases associated with Malassezia species. J. Am. Acad. Dermatol. 2004, 51, 785–798. [Google Scholar] [CrossRef]
- Gupta, A.K.; Madzia, S.E.; Batra, R. Etiology and management of Seborrheic dermatitis. Dermatology 2004, 208, 89–93. [Google Scholar] [CrossRef]
- Gupta, A.K.; Bluhm, R. Seborrheic dermatitis. J. Eur. Acad. Dermatol. Venereol. 2004, 18, 13–26; quiz 19–20. [Google Scholar] [CrossRef]
- Paulino, L.C.; Tseng, C.H.; Blaser, M.J. Analysis of Malassezia microbiota in healthy superficial human skin and in psoriatic lesions by multiplex real-time PCR. FEMS Yeast Res. 2008, 8, 460–471. [Google Scholar] [CrossRef] [Green Version]
- Yim, S.M.; Kim, J.Y.; Ko, J.H.; Lee, Y.W.; Choe, Y.B.; Ahn, K.J. Molecular analysis of Malassezia microflora on the skin of the patients with atopic dermatitis. Ann. Dermatol. 2010, 22, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Mirhendi, H.; Makimura, K.; Zomorodian, K.; Yamada, T.; Sugita, T.; Yamaguchi, H. A simple PCR-RFLP method for identification and differentiation of 11 Malassezia species. J. Microbiol. Methods 2005, 61, 281–284. [Google Scholar] [CrossRef]
- Gupta, A.K.; Boekhout, T.; Theelen, B.; Summerbell, R.; Batra, R. Identification and typing of Malassezia species by amplified fragment length polymorphism and sequence analyses of the internal transcribed spacer and large-subunit regions of ribosomal DNA. J. Clin. Microbiol. 2004, 42, 4253–4260. [Google Scholar] [CrossRef] [PubMed]
- Duarte, E.R.; Hamdan, J.S. RAPD differentiation of Malassezia spp. from cattle, dogs and humans. Mycoses 2010, 53, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Akaza, N.; Akamatsu, H.; Sasaki, Y.; Takeoka, S.; Kishi, M.; Mizutani, H.; Sano, A.; Hirokawa, K.; Nakata, S.; Matsunaga, K. Cutaneous Malassezia microbiota in atopic dermatitis patients differ by gender and body part. Dermatology 2010, 221, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Ilahi, A.; Hadrich, I.; Neji, S.; Trabelsi, H.; Makni, F.; Ayadi, A. Real-Time PCR Identification of Six Malassezia Species. Curr. Microbiol. 2017, 74, 671–677. [Google Scholar] [CrossRef]
- Vergnaud, G.; Denoeud, F. Minisatellites: Mutability and genome architecture. Genome Res. 2000, 10, 899–907. [Google Scholar] [CrossRef] [Green Version]
- Schouls, L.M.; van der Heide, H.G.; Vauterin, L.; Vauterin, P.; Mooi, F.R. Multiple-locus variable-number tandem repeat analysis of Dutch Bordetella pertussis strains reveals rapid genetic changes with clonal expansion during the late 1990s. J. Bacteriol. 2004, 186, 5496–5505. [Google Scholar] [CrossRef] [Green Version]
- Top, J.; Schouls, L.M.; Bonten, M.J.; Willems, R.J. Multiple-locus variable-number tandem repeat analysis, a novel typing scheme to study the genetic relatedness and epidemiology of Enterococcus faecium isolates. J. Clin. Microbiol. 2004, 42, 4503–4511. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.W.; Watanabe, H.; Phung, D.C.; Tung, S.K.; Lee, Y.S.; Terajima, J.; Liang, S.Y.; Chiou, C.S. Multilocus variable-number tandem repeat analysis for molecular typing and phylogenetic analysis of Shigella flexneri. BMC Microbiol. 2009, 9, 278. [Google Scholar] [CrossRef] [Green Version]
- Thierry, S.; Wang, D.; Arne, P.; Deville, M.; De Bruin, B.; Nieguitsila, A.; Pourcel, C.; Laroucau, K.; Chermette, R.; Huang, W.; et al. Multiple-locus variable-number tandem repeat analysis for molecular typing of Aspergillus fumigatus. BMC Microbiol. 2010, 10, 315. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.Y.; Hadj-Henni, L.; Thierry, S.; Arne, P.; Chermette, R.; Botterel, F.; Hadrich, I.; Makni, F.; Ayadi, A.; Ranque, S.; et al. Simple and highly discriminatory VNTR-based multiplex PCR for tracing sources of Aspergillus flavus isolates. PLoS ONE 2012, 7, e44204. [Google Scholar] [CrossRef]
- Hadrich, I.; Neji, S.; Drira, I.; Trabelsi, H.; Mahfoud, N.; Ranque, S.; Makni, F.; Ayadi, A. Microsatellite typing of Aspergillus flavus in patients with various clinical presentations of aspergillosis. Med. Mycol. 2013, 51, 586–591. [Google Scholar] [CrossRef] [PubMed]
- Drira, I.; Hadrich, I.; Neji, S.; Mahfouth, N.; Trabelsi, H.; Sellami, H.; Makni, F.; Ayadi, A. Highly discriminatory variable-number tandem-repeat markers for genotyping of Trichophyton interdigitale strains. J. Clin. Microbiol. 2014, 52, 3290–3296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neji, S.; Hadrich, I.; Ilahi, A.; Trabelsi, H.; Chelly, H.; Mahfoudh, N.; Cheikhrouhou, F.; Sellami, H.; Makni, F.; Ayadi, A. Molecular Genotyping of Candida parapsilosis Species Complex. Mycopathologia 2018, 183, 765–775. [Google Scholar] [CrossRef]
- Rousset, F. genepop’007: A complete re-implementation of the genepop software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Hunter, P.R.; Gaston, M.A. Numerical index of the discriminatory ability of typing systems: An application of Simpson’s index of diversity. J. Clin. Microbiol. 1988, 26, 2465–2466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goudet, J. FSTAT Version 1.2: Un programme informatique pour calculer la statistique F. J. Hérédité 1995, 86, 485–486. [Google Scholar]
- Polzin, T.; Daneschmand, S.V. On Steiner trees and minimum spanning trees in hypergraphs. Oper. Res. Lett. 2003, 31, 12–20. [Google Scholar] [CrossRef]
- Dolatabadi, S.; de Hoog, G.S.; Meis, J.F.; Walther, G. Species boundaries and nomenclature of Rhizopus arrhizus (syn. R. oryzae). Mycoses 2014, 57 (Suppl. 3), 108–127. [Google Scholar] [CrossRef]
- Nyilasi, I.; Papp, T.; Csernetics, A.; Krizsan, K.; Nagy, E.; Vagvolgyi, C. High-affinity iron permease (FTR1) gene sequence-based molecular identification of clinically important Zygomycetes. Clin. Microbiol. Infect. 2008, 14, 393–397. [Google Scholar] [CrossRef] [Green Version]
- Puig, L.; Bragulat, M.R.; Castella, G.; Cabanes, F.J. Phenotypic and genetic diversity of Malassezia furfur from domestic and zoo animals. Med. Mycol. 2018, 56, 941–949. [Google Scholar] [CrossRef]
- Theelen, B.; Cafarchia, C.; Gaitanis, G.; Bassukas, I.D.; Boekhout, T.; Dawson, T.L., Jr. Malassezia ecology, pathophysiology, and treatment. Med. Mycol. 2018, 56, S10–S25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czyzewska, U.; Bartoszewicz, M.; Siemieniuk, M.; Tylicki, A. Genetic relationships and population structure of Malassezia pachydermatis strains isolated from dogs with otitis externa and healthy dogs. Mycologia 2018, 110, 666–676. [Google Scholar] [CrossRef]
- Cafarchia, C.; Gasser, R.B.; Latrofa, M.S.; Parisi, A.; Campbell, B.E.; Otranto, D. Genetic variants of Malassezia pachydermatis from canine skin: Body distribution and phospholipase activity. FEMS Yeast Res. 2008, 8, 451–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celis, A.M.; Vos, A.M.; Triana, S.; Medina, C.A.; Escobar, N.; Restrepo, S.; Wosten, H.A.; de Cock, H. Highly efficient transformation system for Malassezia furfur and Malassezia pachydermatis using Agrobacterium tumefaciens-mediated transformation. J. Microbiol. Methods 2017, 134, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hiruma, M.; Cho, O.; Hiruma, M.; Kurakado, S.; Sugita, T.; Ikeda, S. Genotype analyses of human commensal scalp fungi, Malassezia globosa, and Malassezia restricta on the scalps of patients with dandruff and healthy subjects. Mycopathologia 2014, 177, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.L.; Cafarchia, C.; Otranto, D.; Ferreira, R.R.; Bianchi, S.P.; Latrofa, M.S.; Parisi, A.; Ferreiro, L. Genetic variability and phospholipase production of Malassezia pachydermatis isolated from dogs with diverse grades of skin lesions. Med. Mycol. 2010, 48, 889–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilahi, A.; Hadrich, I.; Goudjil, S.; Kongolo, G.; Chazal, C.; Leke, A.; Ayadi, A.; Chouaki, T.; Ranque, S. Molecular epidemiology of a Malassezia pachydermatis neonatal unit outbreak. Med. Mycol. 2018, 56, 69–77. [Google Scholar] [CrossRef]
- Cafarchia, C.; Latrofa, M.S.; Figueredo, L.A.; da Silva Machado, M.L.; Ferreiro, L.; Guillot, J.; Boekhout, T.; Otranto, D. Physiological and molecular characterization of atypical lipid-dependent Malassezia yeasts from a dog with skin lesions: Adaptation to a new host? Med. Mycol. 2011, 49, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Cafarchia, C.; Gasser, R.B.; Figueredo, L.A.; Latrofa, M.S.; Otranto, D. Advances in the identification of Malassezia. Mol. Cell. Probes 2011, 25, 1–7. [Google Scholar] [CrossRef]
- Soares, R.C.; Zani, M.B.; Arruda, A.C.; Arruda, L.H.; Paulino, L.C. Malassezia intra-specific diversity and potentially new species in the skin microbiota from Brazilian healthy subjects and seborrheic dermatitis patients. PLoS ONE 2015, 10, e0117921. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Marker Name | Primer Sequences (5′ to 3′) | DNA Motifs (bp) | Number of Alleles | Fragment Size (bp) | Simpson Diversity Index D | Marker Location |
|---|---|---|---|---|---|---|---|
| M. globosa | STR-MG1 | F: FAM-CACGCTAAACTGCTGGATGA | 2 | 6 | 242-256-270-286-296-302 | 0.829 | Ch9 |
| R: CGAATATGAACCCCATGGAC | |||||||
| STR-MG2 | F: HEX-CGGTAGTACGATTGGCCCTA | 2 | 5 | 198-228-232-272-282 | 0.741 | Ch3 | |
| R: CCACATCATACGAGCCACAC | |||||||
| STR-MG3 | F: TET-GGATGAAGAGGCTCGCTATG | 3 | 5 | 168-177-189-213-234 | 0.722 | Ch3 | |
| R: ACAGGCGTAGAAGCCAAAGA | |||||||
| STR-MG4 | F: GGCGGTGGTAAGCACTCTGG | 66 | 6 | 250-300-350-400-450-500 | 0.807 | Ch1 | |
| R: ATGGGTGGTCACCACATGG | |||||||
| STR-MG5 | F: AGGGGCTTCACGGTCATGTG | 86 | 4 | 300-400-500-700 | 0.649 | Ch3 | |
| R: CATTGGAGCCTGGTGTGACG | |||||||
| STR-MG6 | F: AACCAACGACGTCCAGAAAC | 176 | 3 | 300-450-700 | 0.581 | Ch5 | |
| R: GTTAAGCTCGCTTGCTCGAAT | |||||||
| M. restricta | STR-MR1 | F: FAM-TAGCACTATCCCAACGTACC | 2 | 5 | 242-250-254-258-286 | 0.774 | Ch7 |
| R: GTTCCGTGTGTCATGATTGC | |||||||
| STR-MR2 | F: HEX-CGCAGAAAACCTAGAGACG | 3 | 5 | 258-264-288-297-336 | 0.818 | Ch3 | |
| R: CTCGTGCGTGTGAGTATTGG | |||||||
| STR-MR3 | F: TET-GCCTTGGATGCACTGGTATT | 3 | 4 | 278-314-338-341 | 0.642 | Ch2 | |
| R: CACGGCGTCAGGAACAAGAG | |||||||
| STR-MR4 | F: ACGGGTTCGAACGGTGAG | 66 | 5 | 250-350-450-500-600 | 0.775 | Ch4 | |
| R: ATGATTGCTTGCGTTGACTG | |||||||
| STR-MR5 | F: TATGGGTGCTGCCAGAGTCG | 162 | 3 | 300-600-750 | 0.549 | Ch4 | |
| R: GTCGAAGGAGATTCACGGCG | |||||||
| STR-MR6 | F: CCCACCACCAACTAACAACA | 228 | 3 | 250-500-750 | 0.521 | Ch5 | |
| R: AAACACGGACCACACAACAC |
| Pop E | Pop A | |
|---|---|---|
| Pop P | 0.018 | 0.045 |
| Pop E | 0.057 |
| Cluster (%) | I | II | III | IV | V | VI | VII | VIII | IX | X | Singleton | p |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Patient | - | 15.38 | 7.69 | - | 3.84 | 3/26 | 7.69 | 7.69 | 11.53 | 7.69 | 26.92 | >0.05 |
| Breeder | 11.11 | 11.11 | - | - | 11.11 | 33.33 | - | - | - | 11.11 | 11.11 | 0.16 |
| Dog | 22.22 | 33.33 | - | 11.11 | 11.11 | 11.11 | - | - | - | - | 11.11 | 0.16 |
| Global Site effect | >0.05 | |||||||||||
| Malassezia folliculitis | 25 | 12.5 | - | - | 6.25 | 12.5 | 12.5 | - | 12.5 | 18.75 | - | |
| Pityriasis versicolor | - | - | - | 22.22 | 22.22 | - | - | 33.33 | 11.11 | 11.11 | - | |
| Pityriasis capitis | - | - | - | 20 | 20 | - | - | - | - | 60 | 0.01 | |
| Healthy human | 25 | - | - | 25 | - | 25 | - | - | - | - | 25 | - |
| Global Year effect | <0.05 | |||||||||||
| 2016 | 13.63 | 18.18 | - | 9.09 | 18.88 | 13.63 | - | - | - | 9.09 | 18.88 | - |
| 2017 | - | - | - | - | - | 40 | - | 40 | - | - | 20 | - |
| 2018 | - | 23.52 | 11.67 | - | - | 5.88 | 11.67 | - | 17.64 | 5.88 | 23.52 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hadrich, I.; Khemakhem, N.; Ilahi, A.; Trabelsi, H.; Sellami, H.; Makni, F.; Neji, S.; Ayadi, A. Genotypic Analysis of the Population Structure in Malassezia globosa and Malassezia restricta. J. Fungi 2023, 9, 263. https://doi.org/10.3390/jof9020263
Hadrich I, Khemakhem N, Ilahi A, Trabelsi H, Sellami H, Makni F, Neji S, Ayadi A. Genotypic Analysis of the Population Structure in Malassezia globosa and Malassezia restricta. Journal of Fungi. 2023; 9(2):263. https://doi.org/10.3390/jof9020263
Chicago/Turabian StyleHadrich, Ines, Nahed Khemakhem, Amin Ilahi, Houaida Trabelsi, Hayet Sellami, Fattouma Makni, Sourour Neji, and Ali Ayadi. 2023. "Genotypic Analysis of the Population Structure in Malassezia globosa and Malassezia restricta" Journal of Fungi 9, no. 2: 263. https://doi.org/10.3390/jof9020263