
Seven New Species of the Genus Geastrum (Geastrales, Geastraceae) in China

Abstract
:1. Introduction
2. Material and Methods
2.1. Morphological Study
2.2. DNA Extraction, Amplification and Sequencing
2.3. Phylogenetic Analyses
3. Results
3.1. Phylogeny
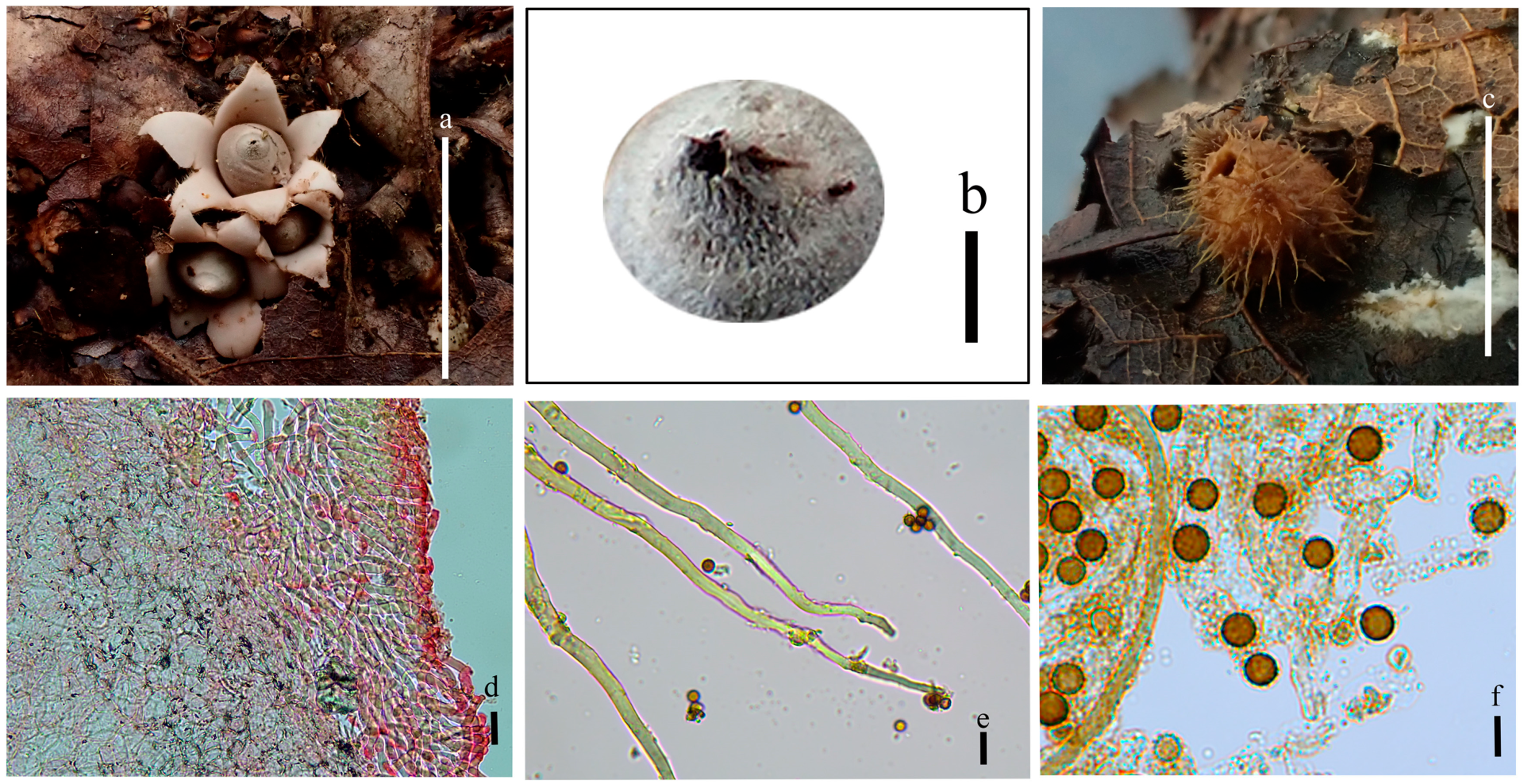
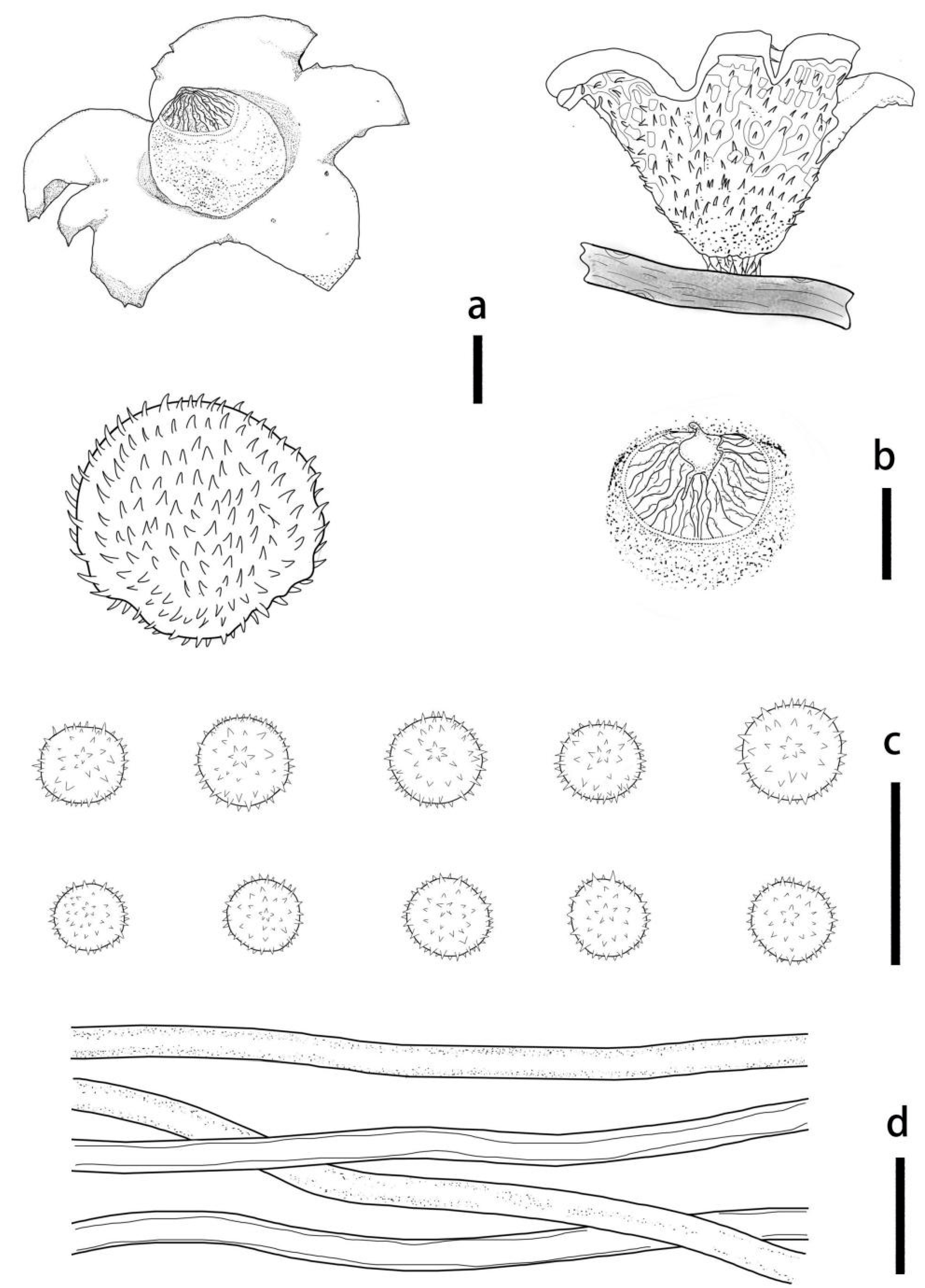
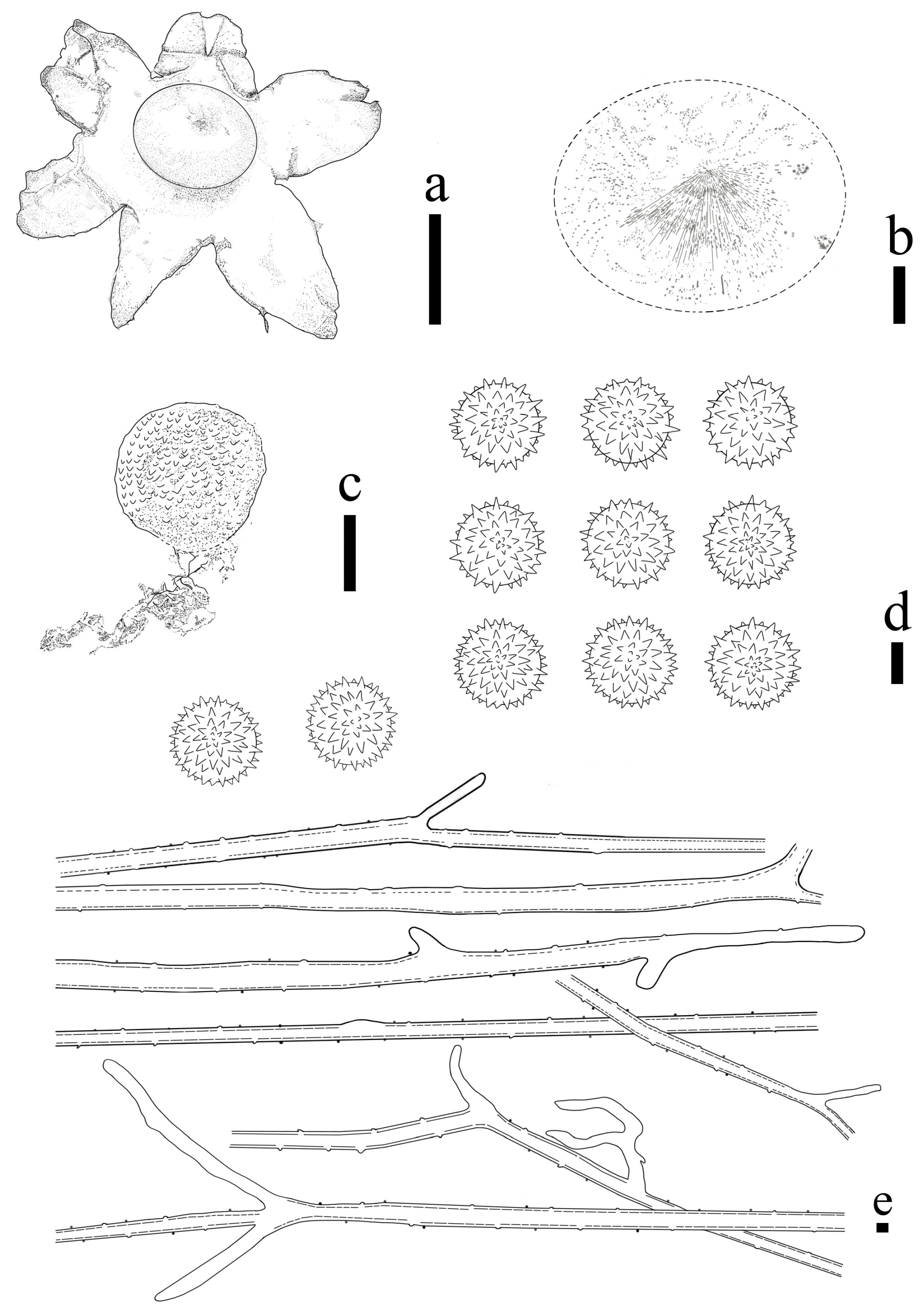
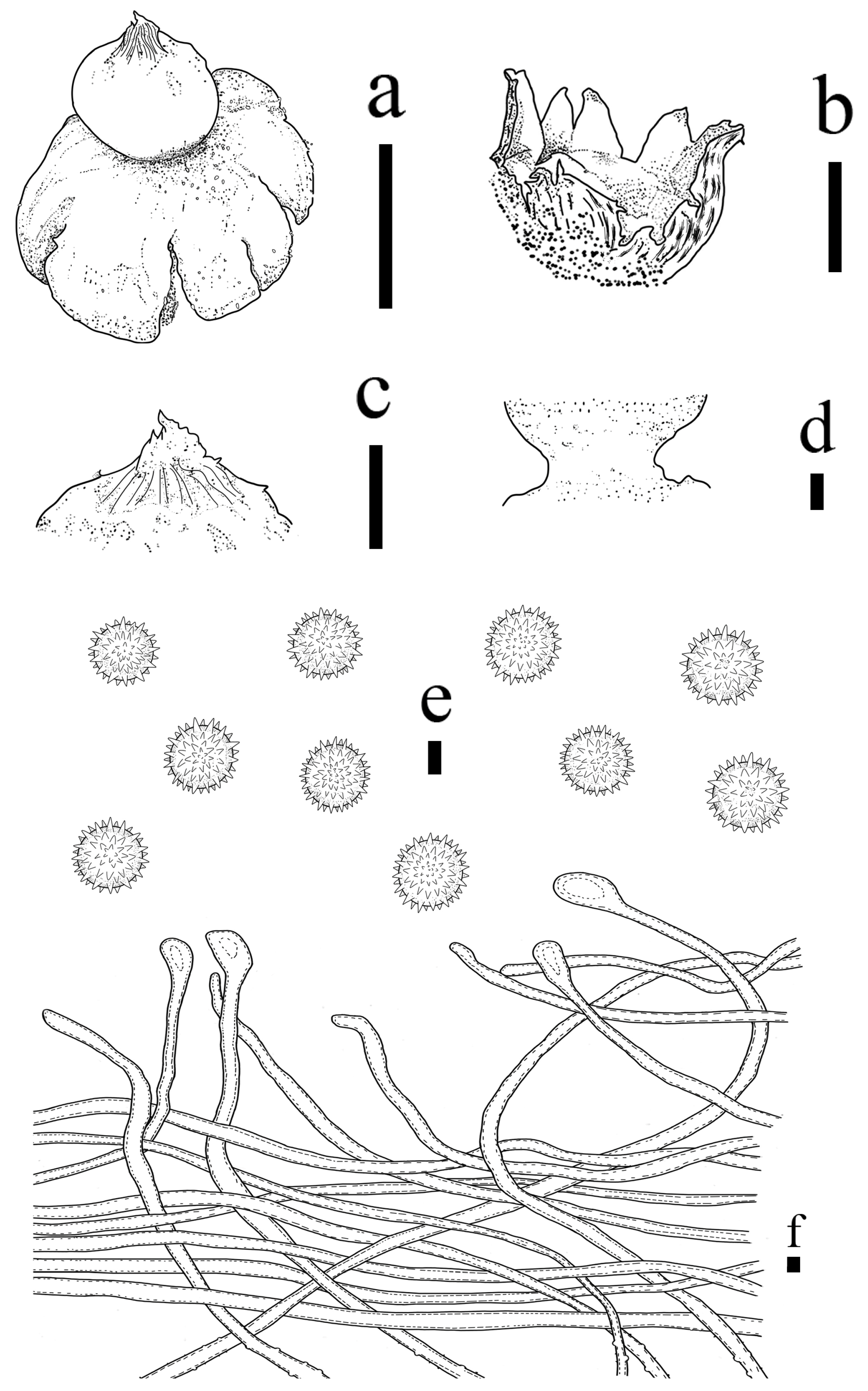
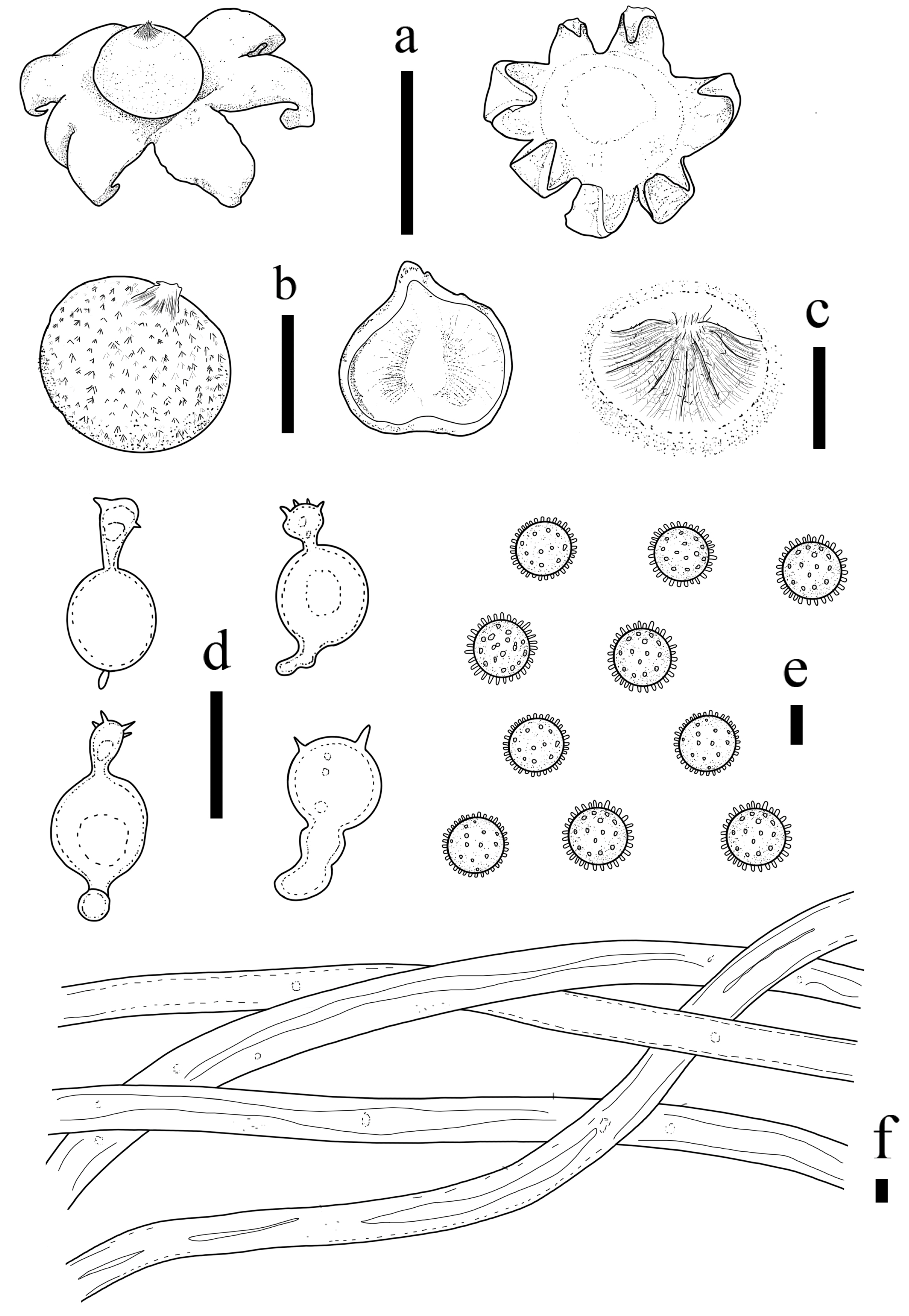
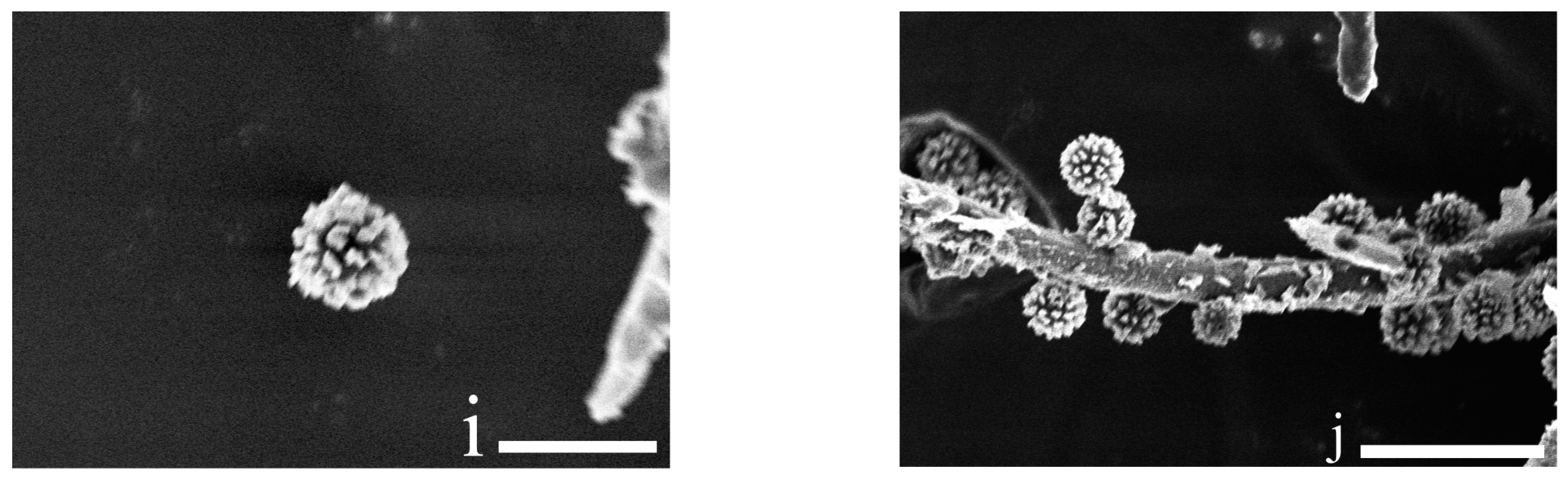
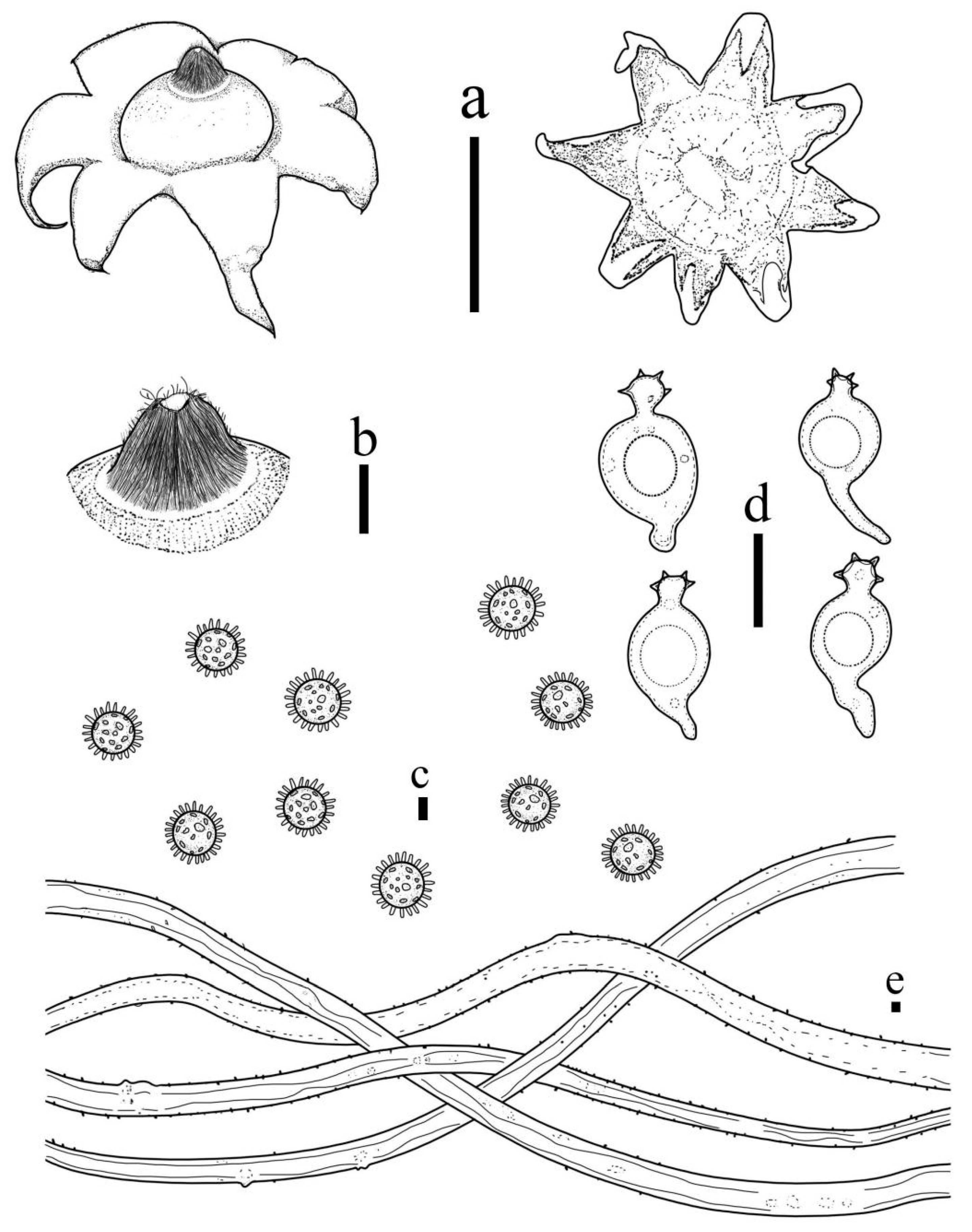
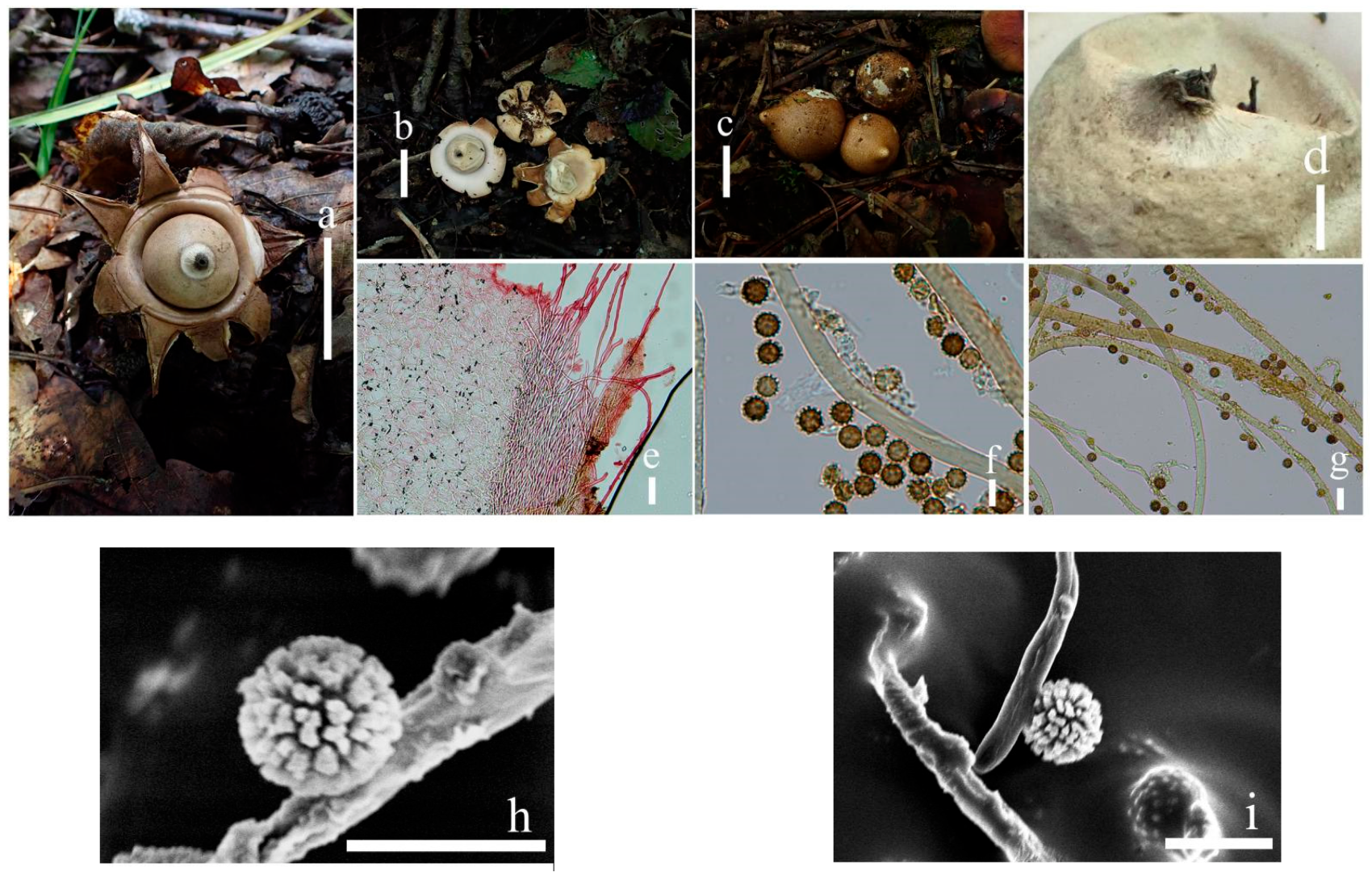
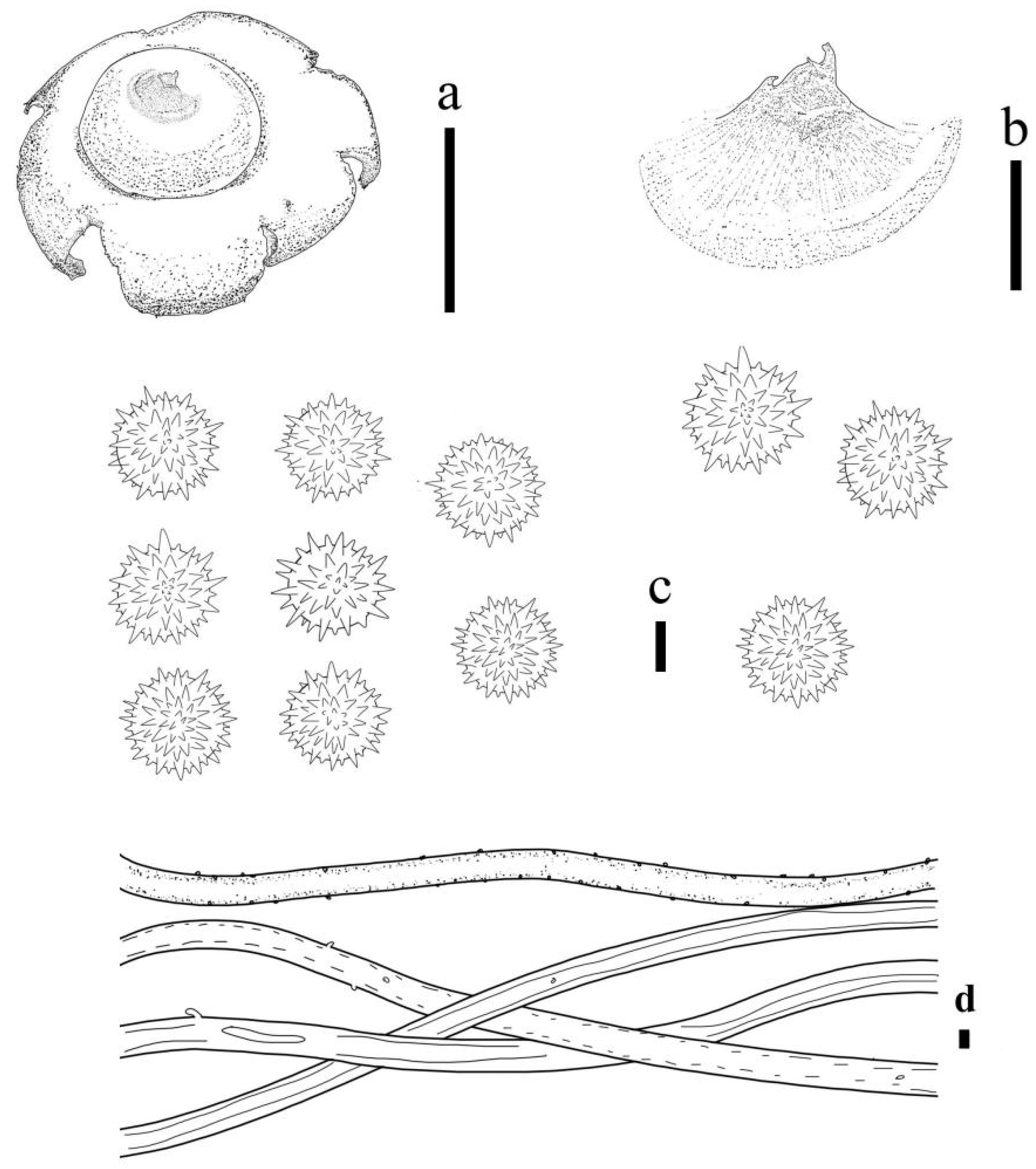
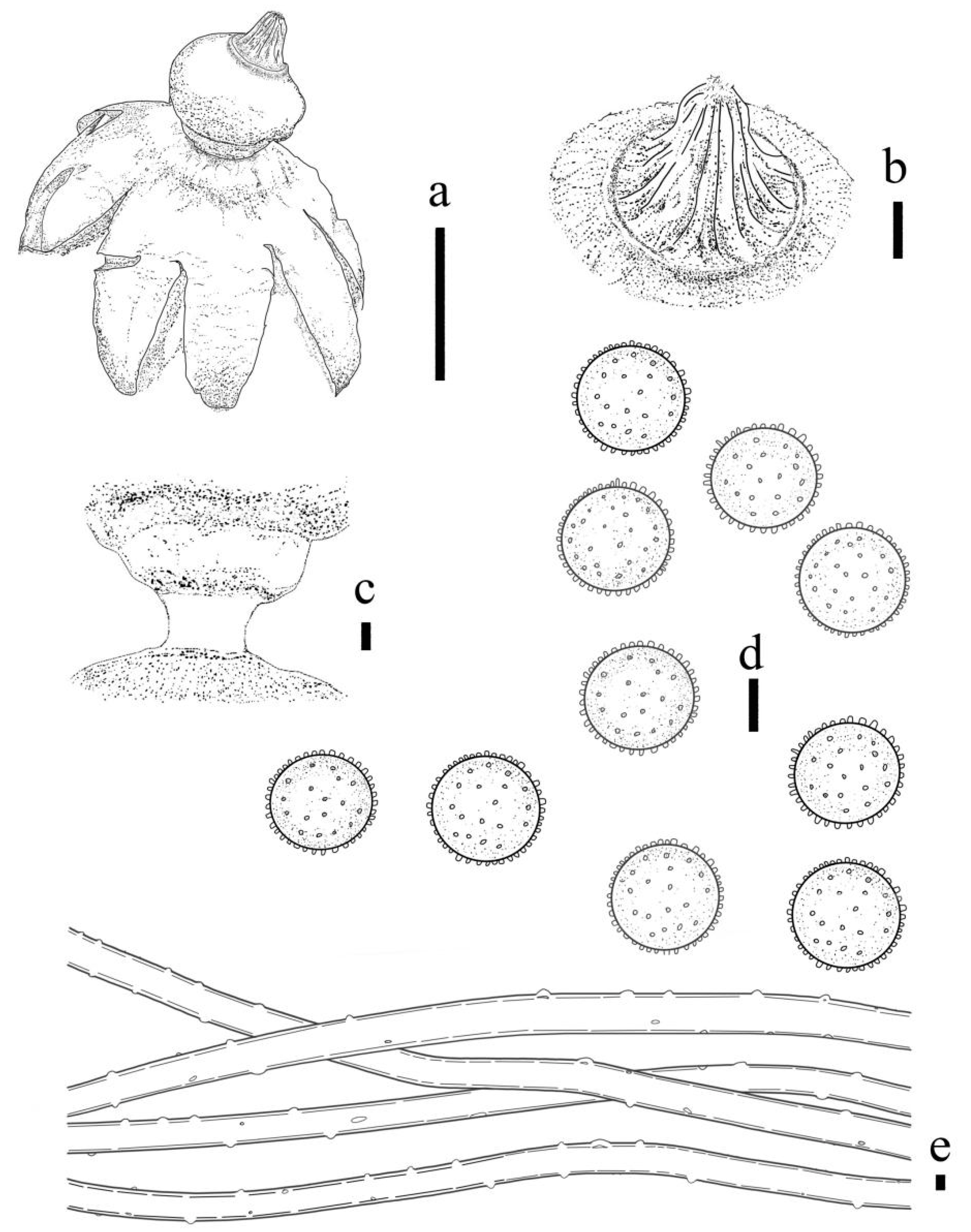
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ponce de León, P. A revision of the family Geastraceae. Fieldiana Bot. 1968, 31, 301–349. [Google Scholar]
- Ma, X.; Tang, S.; Cao, M. Magical Fungi-Geastrum. Life World 2010, 9, 2. [Google Scholar]
- Persoon, C.H. Neuer Versuch einer systematischen Einteilung der Schwämme. Neues Mag. Bot. 1794, 1, 63–128. [Google Scholar]
- De Toni, G.B. Revisio monographica generis Geasteris Mich.e tribu Gasteromycetum. Rev. Mycol. 1887, 9, 61–77. [Google Scholar]
- Hollós, L. Magyarorszag Gasteromycetai; FranklinTársulat: Budapest, Hungary, 1903. [Google Scholar]
- Staněk, V.J. Geastraceae. In Flora ČSR B1: Gasteromycetes; Pilát, A., Ed.; Českolovenská Akademie Věd: Prague, Czech Republic, 1958; pp. 392–526, 777–795. [Google Scholar]
- Sunhede, S. Geastraceae (Basidiomycotina). Morphology, ecology and systematics with special emphasis on the North European species. Synop. Fungorum 1989, 1, 1–534. [Google Scholar]
- Dissing, H.; Lange, M. Gasteromycetes of Congo. Bull. Jard. Bot. État Brux. 1962, 32, 325–416. [Google Scholar] [CrossRef]
- Dörfelt, H. Die Erdsterne: Geastraceae und Astraeaceae; Ziemsen: Wittenberg Lutherstadt, Germany, 1985. [Google Scholar]
- Lloyd, C.G. The Geastrae. Bull. Lloyd Libr. Bot. 1902, 2, 1–43. [Google Scholar]
- Zamora, J.C.; Calonge, F.D.; Martín, M.P. Integrative taxonomy reveals an unexpected diversity in Geastrum section Geastrum (Geastrales, Basidiomycota). Persoonia 2015, 34, 130–165. [Google Scholar] [CrossRef]
- Jeppson, M.; Nilsson, R.H.; Larsson, E. European Earthstars in Geastraceae (Geastrales, Phallomycetidae) a Systematic Approach Using Morphology and Molecular Sequence Data. Syst. Biodivers. 2013, 11, 437–465. Available online: https://www.researchgate.net/publication/263190804 (accessed on 6 November 2022). [CrossRef]
- Jeppson, M. Jordstjärnor. Mykol. Publ. 2013, 6, 1–228. [Google Scholar]
- Zamora, J.C.; Calonge, F.D.; Martín, M.P. New sources of taxonomic information for earthstars (Geastrum, Geastraceae, Basidiomycota): Phenoloxidases and rhizomorph crystals. Phytotaxa 2013, 132, 1–20. [Google Scholar] [CrossRef]
- Zamora, J.C.; Kuhar, F.; Castiglia, V.; Papinutti, L. On Geastrum argentinum, a forgotten species. Mycoscience 2014, 55, 177–182. [Google Scholar] [CrossRef]
- Zamora, J.C. Proposal to conserve the name Geastrum (Geastrales, Basidiomycota) with a conserved type. Taxon 2014, 63, 679–680. [Google Scholar] [CrossRef]
- Zamora, J.C.; de Diego Calonge, F.; Hosaka, K.; Martín, M.P. Systematics of the genus Geastrum (Fungi: Basidiomycota) revisited. Taxon 2014, 63, 477–497. [Google Scholar] [CrossRef]
- Assis, N.M.; Freitas-Neto, J.F.; Sousa, J.O.; Fr, B.; Baseia, L. Geastrum hyalinum (Basidiomycota, Geastraceae), a new species from Brazilian Southern Amazon. Stud. Fungi 2019, 4, 83–89. [Google Scholar] [CrossRef]
- Finy, P.; Papp, V.; Knapp, D.G.; Bóka, K.; Kovács, G.M.; Dima, B. Geastrum dolomiticum, a new earthstar species from Central Europe. Plant Syst. Evol. 2021, 307, 43. [Google Scholar] [CrossRef]
- Teng, S.C. Fungi of China; Science Press: Beijing, China, 1963; pp. 678–682. [Google Scholar]
- Tai, F.L. Sylloge Fungorum Sinicorum; Science Press; Academia Sinica: Beijing, China, 1979; pp. 471–473. [Google Scholar]
- Liu, B. The Gasteromycetes of China. Beihefte Nova Hedwigia 1984, 76, 1–235. [Google Scholar]
- Yuan, M.S.; Sun, P.Q. ; Sichuan Mushroom; Sichuan Science and Technology Press: Sichuan, China, 1995; pp. 625–627. [Google Scholar]
- Mao, X.L. Macrofungi of China; Henan Technology Press: Henan, China, 2000; pp. 244–245. [Google Scholar]
- Li, Y.; Tolgor, B. Mushroom in Changbai Mountain; Science Press: Beijing, China, 2003; pp. 296–297. [Google Scholar]
- Zhou, T.X.; Chen, Y.H.; Zhao, L.Z.; Fu, H.; Yang, B. Flora Fungorum Sinicorum. Vol. 36. Geastraceae, Nidulariaceae; Science Press: Beijing, China, 2007; pp. 1–167. [Google Scholar]
- Li, Y.; Li, T.H.; Yang, Z.L.; Bau, T.; Dai, Y.C. Atlas of Chinese Macrofungal Resources; Central China Farmer’s Publishing House: Zhengzhou, China, 2015; pp. 1–1351. [Google Scholar]
- Han, B.X.; Bau, T. New records of Geastrum from China. Mycosystema 2016, 35, 1416–1424. [Google Scholar] [CrossRef]
- Zhou, H.; Li, J.; Hou, C. Two new species of the genus Geastrum from the Yanshan Montains in China. Mycosystema 2022, 41, 1–16. [Google Scholar] [CrossRef]
- Cai, Q.; Cui, Y.Y.; Yang, Z.L. Lethal Amanita species in China. Mycologia 2016, 108, 993–1009. [Google Scholar] [CrossRef]
- Cui, Y.Y.; Cai, Q.; Tang, L.; Liu, J.W.; Yang, Z.L. The family Amanitaceae: Molecular phylogeny, higher-rank taxonomy and the species in China. Fungal Divers. 2018, 91, 5–230. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Methuen: London, UK, 1978; p. 252. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and directsequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes–application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Geml, J.; Davis, D.D.; Geiser, D.M. Systematics of the genus Sphaerobolus based on molecular and morphological data, with the description of Sphaerobolus ingoldii sp. nov. Mycologia 2005, 97, 680–694. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Molec. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. raxmlGUI 2.0: A graphical interface and toolkit for phylogenetic analyses using RAxML. Methods Ecol. Evol. 2020, 12, 373–377. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree v1.4.3. 2016. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 27 December 2022).
- Hemmes, D.E.; Desjardin, D.E. Earthstars (Geastrum, Myriostoma) of the Hawaiian Islands Including Two New Species, Geastrum litchiforme and Geastrum reticulatum. Pac. Sci. 2011, 65, 477–496. [Google Scholar] [CrossRef]
- Accioly, T.; Sousa, J.O.; Moreau, P.A.; Lecuru, C.; Silva, B.D.; Roy, M.; Monique, G.; Iuri, G.B.; Martin, M.P. Hidden fungal diversity from the Neotropics: Geastrum hirsutum, G. schweinitzii (Basidiomycota, Geastrales) and their allies. PLoS ONE 2019, 14, e0211388. [Google Scholar] [CrossRef]
- Sousa, J.O.; Baracho, G.S.; Baseia, I.G. Geastrum laevisporum: A new earthstar fungus with uncommon smooth spores. Mycosphere 2015, 6, 501–507. [Google Scholar] [CrossRef]
- Kotlaba, F.; Zehnálek, P. Sixteen gasteromycetes collected in Cuba 50 years ago. Czech Mycol. 2018, 70, 185–209. [Google Scholar] [CrossRef]
- Johansson, K.A. Blomjordstjärna (Geastrum floriforme) funnen i Västergötland. Sven. Mykol. 2008, 29, 74–79. [Google Scholar]
- Kasuya, T.; Hosaka, K.; Uno, K.; Makoto, K. Phylogenetic placement of Geastrum melanocephalum and polyphyly of Geastrum triplex. Mycoscience 2012, 53, 411–426. [Google Scholar] [CrossRef]
- Dai, Y.C.; Yang, Z.L.; Cui, B.K.; Wu, G.; Yuan, H.S.; Zhou, L.W.; He, S.H.; Ge, Z.W.; Wu, F.; Wei, Y.L.; et al. Diversity and systematics of the important macrofungi in Chinese forests. Mycosystema 2021, 40, 770–805. [Google Scholar] [CrossRef]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Designation | Dosage (μL) |
|---|---|
| DNA template | 4.0 |
| Forward primer | 1.0 |
| Reverse primer | 1.0 |
| 2× M5 HiPer plusTaqHiFi PCR mix (with green dye) | 12.5 |
| Nuclease-free ddH2O | 11.5 |
| Species | Geographic | Voucher | GenBank Accessions No. | |
|---|---|---|---|---|
| Origin | Number | ITS | nrLSU | |
| G. albonigrum | Panama | MA-Fungi 36140-2 | KF988349 | KF988468 |
| G. argentinum | Argentina | MA-Fungi 82605 | KF988353 | KF988473 |
| G. argentinum | Argentina | LPS 48446 | KF988352 | KF988472 |
| G. cf. arenarium | Spain | Herb. Zamora 76 | KF988338 | KF988470 |
| G. cf. arenarium | Spain | MA-Fungi 68191 | KF988350 | KF988469 |
| G. austrominimum | Australia | MEL 2292062 | KP687491 | KP687452 |
| G. austrominimum | Australia | MEL:2276089 | KP687490 | KP687451 |
| G. beijingense | China | BJTC 248 | MZ508872 | - |
| G. beijingense | China | BJTC 073 | MZ508873 | - |
| G. benitoi. | Spain | MA:Fungi 87324 | KP687494 | KP687455 |
| G. berkeleyi | Spain | MA-Fungi 74668 | KF988354 | KF988474 |
| G. berkeleyi | Slovakia | MJ8673/MJ867 | KC581985 | KC581985 |
| G. brunneocapillatum | Brazil | UFRN:Fungos:2286 | MH634996 | MH635029 |
| G. cf. calceum | Brazil | UFRN-Fungos 723 | KF988340 | KF988477 |
| G. campestre | USA | MICH 28566 | KF988358 | KF988480 |
| G. coronatum | Hungary | PRM:842868(holo) | KP687495 | KP687456 |
| G. coronatum | Spain | Zamora 181 | KP687496 | KP687457 |
| G. courtecuissei. | France | LIP:FH 2004090503 | MH635003 | MH635033 |
| G. corollinum | Spain | MA-Fungi 5746 | KF988359 | KF988481 |
| G. corollinum | Sweden | Herb. Sunhede 7744 | KF988360 | KF988482 |
| G. dolomiticum | Hungary: Fejér | FP20150908(holotype) | MT569463 | MT569455 |
| G. dolomiticum | Hungary: Veszprém | FP20151015 | MT569464 | MT569456 |
| G. elegans | Spain | Herb. Zamora 189 | KF988366 | KF988488 |
| G. elegans | Sweden | UPS F-560810 | KF988367 | KF988489 |
| G. fimbriatum | Spain | Herb. Zamora 234 | KF988369 | KF988491 |
| G. fimbriatum | Sweden | Herb. Sunhede 7739 | KF988370 | KF988492 |
| G. flexuosum | Sweden | UPS F-119844 | KF988371 | KF988493 |
| G. floriforme | Spain | Herb.Zamora 453 | KF988373 | KF988495 |
| G. floriforme | Spain | MA-Fungi 69173 | KF988372 | KF988494 |
| G. fornicatum | Spain | Herb.Zamora 255 | KF988374 | KF988496 |
| G. fornicatum | Spain | MA-Fungi 30749 | KF988375 | KF988497 |
| G. fuscogleba | USA | NY Trappe 9500 | KF988377 | KF988499 |
| G. fuscogleba | USA | NY Trappe 1071 | KF988376 | KF988498 |
| G. glaucescens | Argentina | MA-Fungi 83762 | KF988378 | KF988500 |
| G. glaucescens | Argentina | MA-Fungi 83763 | KF988379 | KF988501 |
| G. gorgonicum | Cape Verde | MA-Fungi 92118 | MN754045 | MN754083 |
| G. gorgonicum | Cape Verde | MA-Fungi 92116 | MN754046 | MN754084 |
| G. hansagiense | Hungary | BP110893 | MN582739 | MN582739 |
| G. hansagiense | Hungary | GBL1 | MN582753 | - |
| G. hariotii | Agentina | MA-Fungi 83765 | KF988381 | KF988504 |
| G. hariotii | Dominican | MA-Fungi 80070 | - | KF988503 |
| G. aff. hariotii | Brazil | MA-Fungi 78296 | KF988382 | KF988505 |
| G. hieronymi | Argentina | MA-Fungi 83767 | KF988344 | KF988509 |
| G. hieronymi | Agentina | MA-Fungi 83766 | KF988384 | KF988508 |
| G. hungaricum | Czech | Sunhede 5993 | KP687500 | KP687461 |
| G. hungaricum | Spain | Zamora 611 | KP687501 | KP687462 |
| G. ishikawae | Brazil | UFRN Fungos 2785 | - | NG.060682 |
| G. javanicum | Brazil | UFRN-Fungos 1215 | KJ127031 | - |
| G. kotlabae | Spain | MA-Fungi 39563 | KF988385 | KF988510 |
| G. kotlabae | Spain | Herb.Zamora 440 | KF988386 | KF988511 |
| G. kuharii | Argentina | MA:Fungi:86913 | KP687502 | KP687463 |
| G. aff. lageniforme | Argentina | MA-Fungi 83768 | KF988389 | KF988516 |
| G. aff. lageniforme | Niger | COFC Hama 327 | KF988390 | KF988517 |
| G. aff. lageniforme | Argentina | MA-Fungi 83770 | KF988391 | KF988518 |
| G. lageniforme | Spain | Herb. Zamora 316 | KF988339 | KF988514 |
| G. lageniforme | Spain | Herb. Zamora 207 | KF988388 | KF988513 |
| G. laneum | China | HMJAU65711 | OP964640 | OP964638 |
| G. laneum | China | HMJAU65704 (type) | OP964641 | OP964639 |
| G. laneum | China | HMJAU65705 | OP964641 | - |
| G. litchi | China | HMJAU65716 (type) | OQ360756 | OP964619 |
| G. litchi | China | HMJAU65717 | - | OP964620 |
| G. marginatum | Spain | ERRO 2012112609 | KP687504 | KP687465 |
| G. marginatum | Czech | PRM:842884 (holo) | KP687507 | KP687468 |
| G. melanocephalum | Spain | Herb. Zamora 34 | KF988395 | KF988522 |
| G. melanocephalum | Sweden | Herb. Sunhede 7737 | KF988396 | KF988523 |
| G. melanorhynchum | China | HMJAU65765 | OP964616 | - |
| G. melanorhynchum | China | HMJAU65764 (type) | OP964617 | OP964614 |
| G. melanorhynchum | China | HMJAU65768 | OP964618 | OP964615 |
| G. meridionale | Spain | Herb. Zamora 252 (holo) | KF988412 | KF988540 |
| G. microphole | China | HMJAU65720 (type) | OP964636 | OP964643 |
| G. microphole | China | HMJAU65721 | OP964637 | OP964644 |
| G. mirabile | Japan | TNS:KH-JPN10-714 | JN845109 | JN845227 |
| G. minutisporum | Argentina | CORD15 | KM260665 | - |
| G. minutisporum | Argentina | CORD14 | KM260664 | - |
| G. minimum | Sweden | MA-Fungi 86669 | KF988405 | KF988533 |
| G. mongolicum | China | HMJAU65762 | OP964647 | OP964645 |
| G. mongolicum | China | HMJAU65763 (type) | OP964648 | OP964646 |
| G. morganii | Canada | Herb. Lebeuf HRL0177 (holo) | KF988406 | KF988534 |
| G. neoamericanum | Brazil | UFRN:Fungos:2302 (holo) | MH635001 | MH635040 |
| G. neoamericanum | French | LIP:JLC12030103 | MH635014 | MH635038 |
| G. ovalisporum | Bolivia | MA-Fungi 47184 | KF988411 | KF988539 |
| G. ovalisporum | Argentina | MA:Fungi 86670 | - | KP687476 |
| G. oxysepalum | China | HMJAU65730 | OP964629 | - |
| G. oxysepalum | China | HMJAU65735 | OP964630 | - |
| G. oxysepalum | China | HMJAU65734 | OP964631 | OP964621 |
| G. oxysepalum | China | HMJAU65727 (type) | OP964632 | OP964622 |
| G. oxysepalum | China | HMJAU65728 | OP964633 | OP964623 |
| G. parvistriatum | Spain | JCZ 272 | JN943162 | JN939572 |
| G. parvisporum | Argentina | BAFC:51926 | MG196037 | MG196035 |
| G. parvisporum | Argentina | MA-Fungi 83793 | KF988461 | KF988596 |
| G. pectinatum | Spain, Lugo | MA:Fungi:28156 | KP687516 | KP687478 |
| G. pleosporum | Cameroon | MA-Fungi 56971 | KF988416 | KF988544 |
| G. pouzarii | Czechoslovakia | MA-Fungi 2944 | KF988417 | KF988545 |
| G. pouzarii | Czechoslovakia | Herb. Sunhede 7494 | KF988418 | KF988546 |
| G. pseudostriatum | Sweden | MJ050919O | KC581990 | - |
| G. pseudostriatum | Sweden | MJ8240 | KC581991 | - |
| G. pseudostriatum | Sweden | MJ7573O1 | KC581992 | KC581992 |
| G. pseudostriatum | Sweden | BP 22110 | NR132884 | |
| G. pseudolimbatum | Spain | Herb. Zamora 231 | KF988419 | KF988547 |
| G. pseudolimbatum | Sweden | UPS F-560804 | KF988420 | KF988548 |
| G. pseudosaccatum | China | HMJAU65778 | OP964624 | - |
| G. pseudosaccatum | China | HMJAU65781 | OP964625 | OP964635 |
| G. pseudosaccatum | China | HMJAU65772 | OP964626 | - |
| G. pseudosaccatum | China | HMJAU65774 | OP964627 | - |
| G. pseudosaccatum | China | HMJAU65769 (type) | OP964628 | OP964634 |
| G. quadrifidum | Spain | Zamora 300 | KP687524 | KP687486 |
| G. quadrifidum | Sweden | MA86671 | KF988422 | KF988550 |
| G. rubropusillum | Brazil | UFRN:Fungos:2308 | MH634994 | MH635027 |
| G. rubellum | France | LIP:CL/MART 8067B | MH635009 | - |
| G. rubellum | Brazil | UFRN:Fungos:2844 | MH634999 | MH635031 |
| G. rufescens | Spain | Herb. Zamora 253 | KF988424 | KF988552 |
| G. rufescens | Spain | Herb. Zamora 274 | KF988425 | KF988553 |
| G. rusticum | Brazil | UFRN Fungos 1217 | - | NG060634 |
| G. saccatum | Japan | UPS F-530056 | KF988428 | KF988558 |
| G. schweinitzii | Argentina | MA-Fungi 83779 | KF988437 | KF988567 |
| G. schmidelii | Sweden | UPSF-560805 | KF988435 | KF988565 |
| G. schmidelii | China | HMAS 84118 | MZ50883 | MZ509381 |
| G. setiferum | Baseia | MA-Fungi 83781 | - | KF988571 |
| G. setiferum | Argentina | MA-Fungi 83782 | - | KF988572 |
| G. smardae | Spain | Herb. Zamora 527 | KF988441 | KF988574 |
| G. smardae | Canada | Herb. Lebeuf HRL 0160 | KF988440 | KF988573 |
| G. striatum | Sweden | MA-Fungi 86672 | KF988443 | KF988577 |
| G. spc070607 | China | HMJAU65787 | - | OP967186 |
| G. spc70808 | China | HMJAU65788 | - | OP967185 |
| G. spc70906 | China | HMJAU65790 | OP972576 | - |
| G. sp22DQ21 | China | HMJAU65789 | OP972575 | OP967188 |
| G. spQWQ8767 | China | HMJAU65786 | - | OP967183 |
| G. spQWQ155634 | China | HMJAU65785 | - | OP967187 |
| G. tenuipes | Australia | CANB:775658 | KP687527 | KP687489 |
| G. tenuipes | Australia | CANB:738350 | KP687526 | KP687488 |
| G. thanatophilum | USA | MICH 72012 | KF988364 | KF988486 |
| G. triplex | Madagascar | UPS F-014630-213863 | KF988444 | KF988578 |
| G. triplex | Argentina | MA-Fungi 83784 | KF988445 | KF988579 |
| G. velutinum | China | BJTC 598 | MZ508877 | - |
| G. violaceum | Agentina | MA-Fungi 82487 | KF988451 | KF988586 |
| G. violaceum | Agentina | BAFC 51671 | KF988450 | KF988585 |
| G. yanshanense | China | BJTC 381 | MZ508878 | MZ509383 |
| G. yanshanense | China | BJTC 057 | MZ508879 | MZ509384 |
| Sphaerobolus iowensis | USA | SS11 | AY487958 | AY439014 |
| S. iowensis | Indiana | SS1 | AY487950 | - |
| Schenella pityophila | Spain | Herb. Zamora 530 | KF988346 | KF988464 |
| S. pityophila | Spain | Herb. Zamora 531 | KF988347 | KF988465 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Bau, T. Seven New Species of the Genus Geastrum (Geastrales, Geastraceae) in China. J. Fungi 2023, 9, 251. https://doi.org/10.3390/jof9020251
Wang X, Bau T. Seven New Species of the Genus Geastrum (Geastrales, Geastraceae) in China. Journal of Fungi. 2023; 9(2):251. https://doi.org/10.3390/jof9020251
Chicago/Turabian StyleWang, Xin, and Tolgor Bau. 2023. "Seven New Species of the Genus Geastrum (Geastrales, Geastraceae) in China" Journal of Fungi 9, no. 2: 251. https://doi.org/10.3390/jof9020251