
Fungistatic Activity Mediated by Volatile Organic Compounds Is Isolate-Dependent in Trichoderma sp. “atroviride B”
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Materials
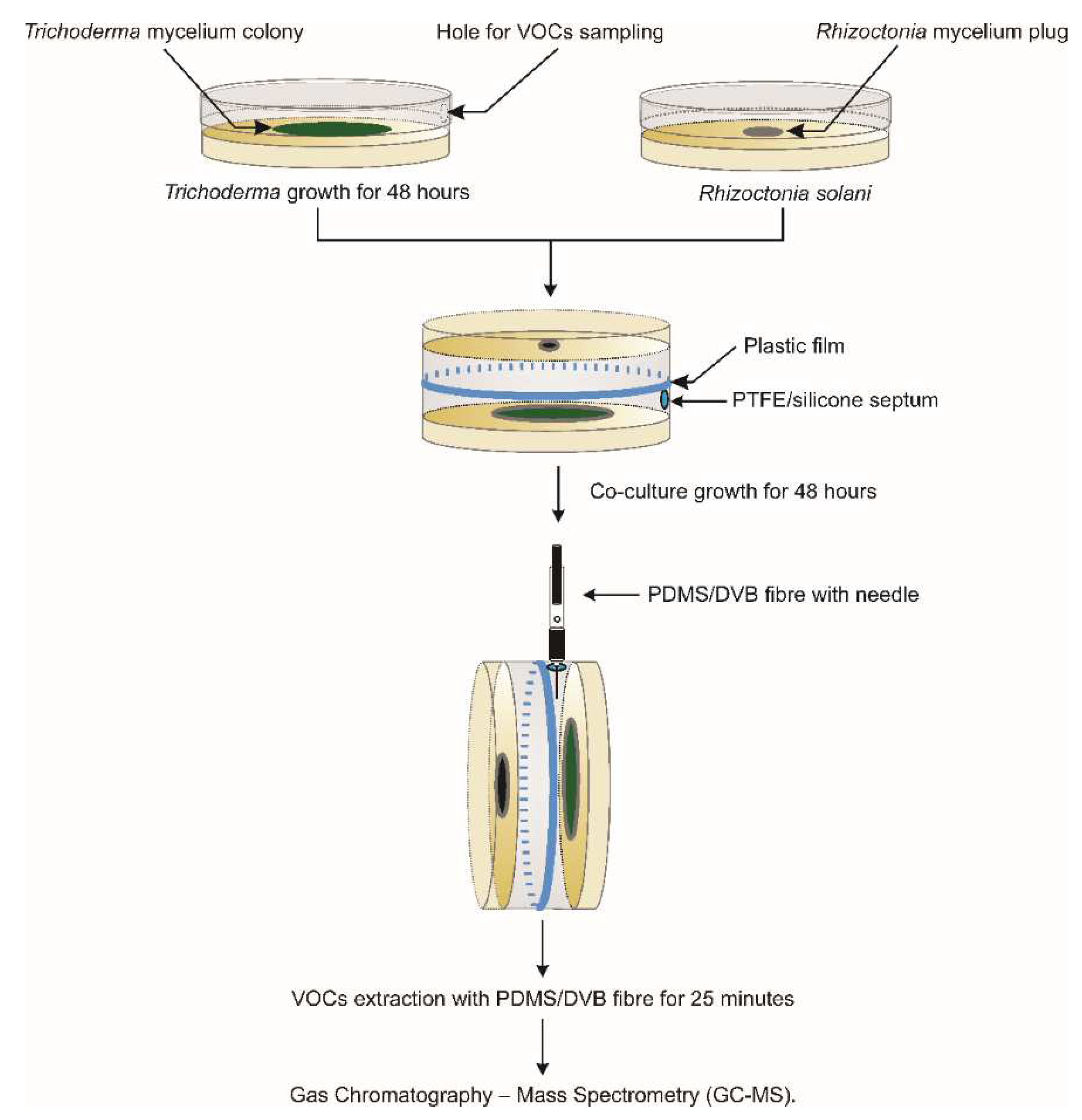
2.2. Inverted Plate Assays
2.3. Evaluation of Trichoderma Isolates
2.4. Analysis of VOCs
2.5. Bioactivity Assays of Pure Compounds
3. Results
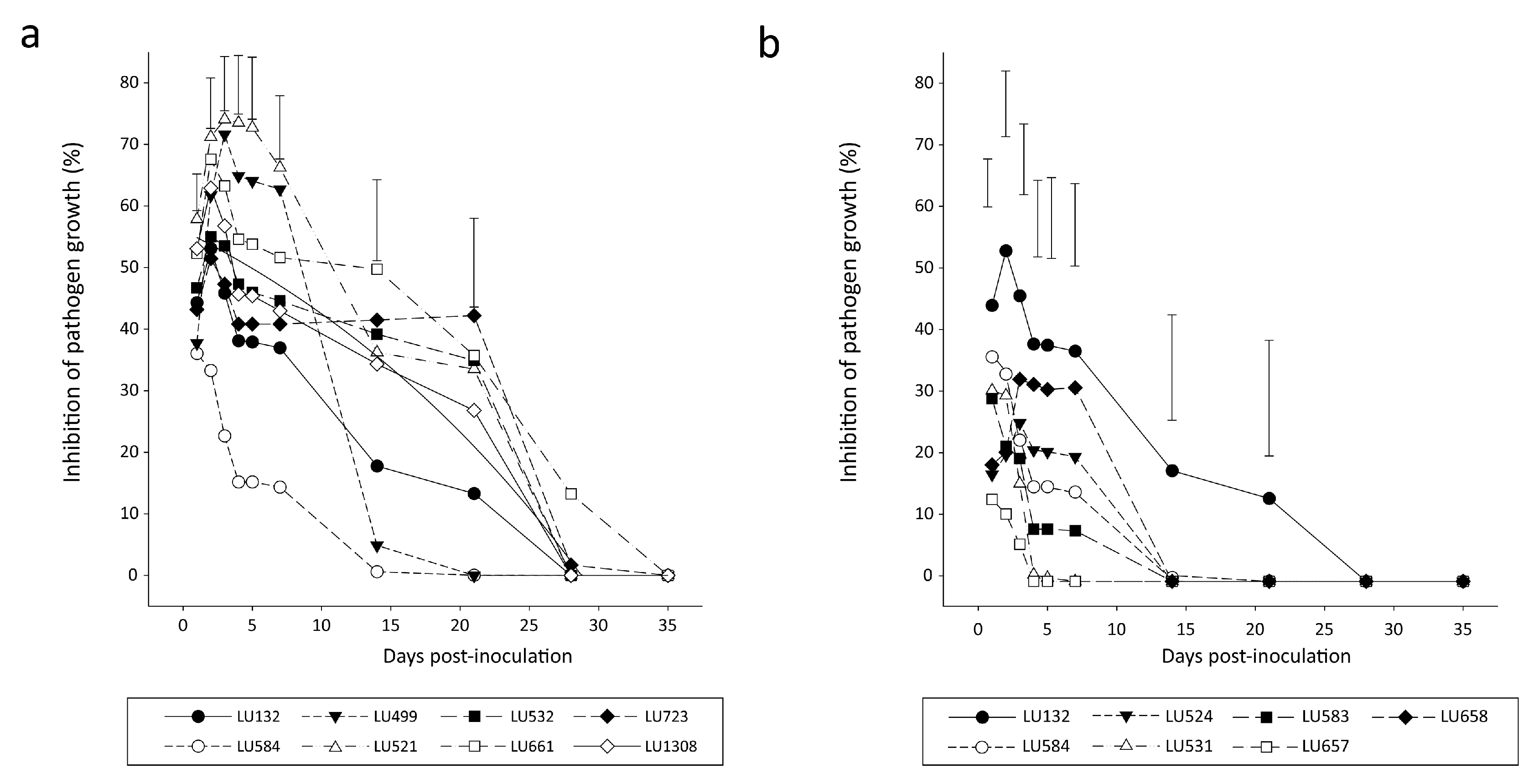
3.1. Bioactivity of VOCs from Trichoderma sp. “atroviride B” against R. solani
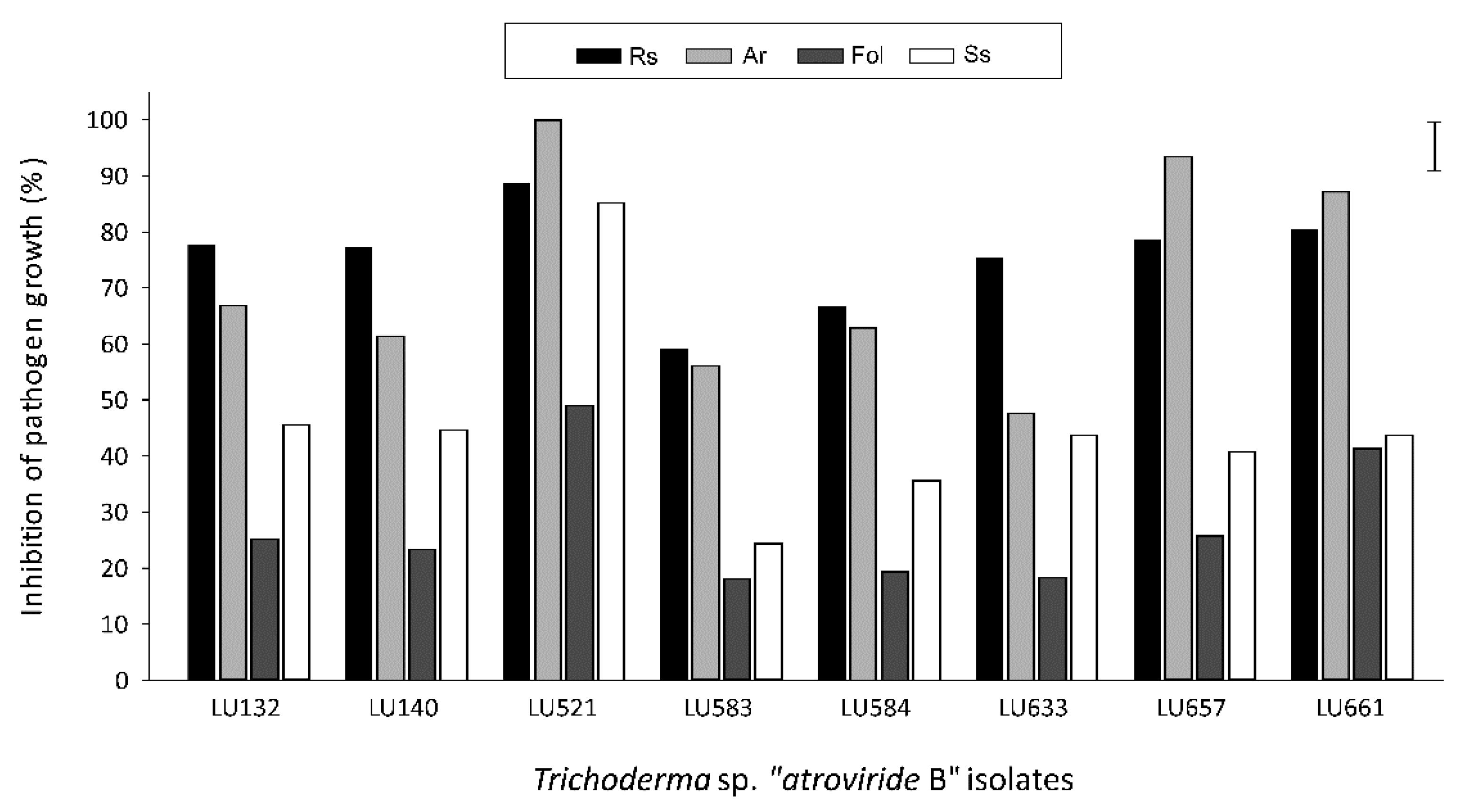
3.2. Bioactivity of VOCs from T. sp. “atroviride B” against other Pathogens
3.3. Analysis of VOCs from T. sp. “atroviride B”
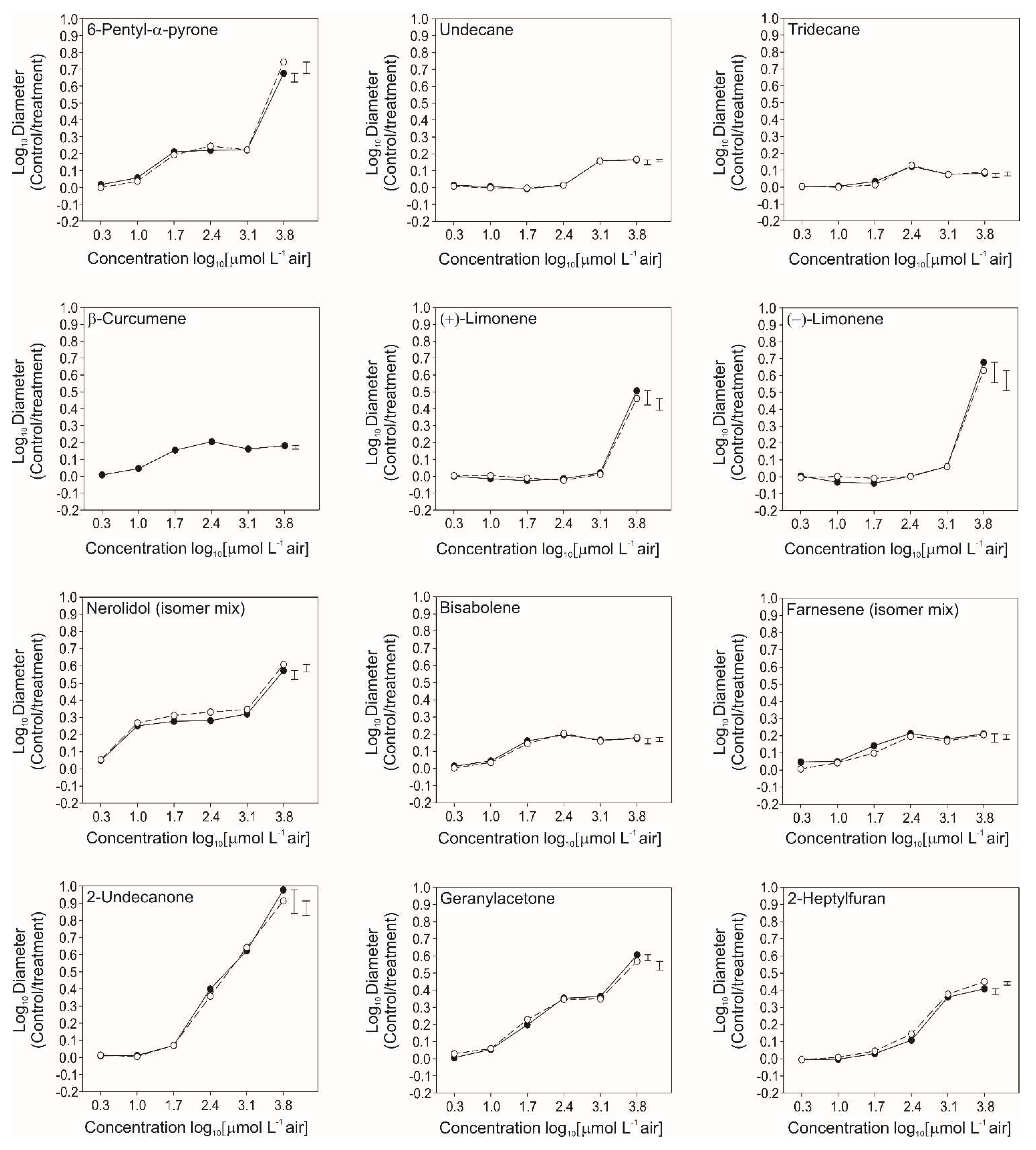
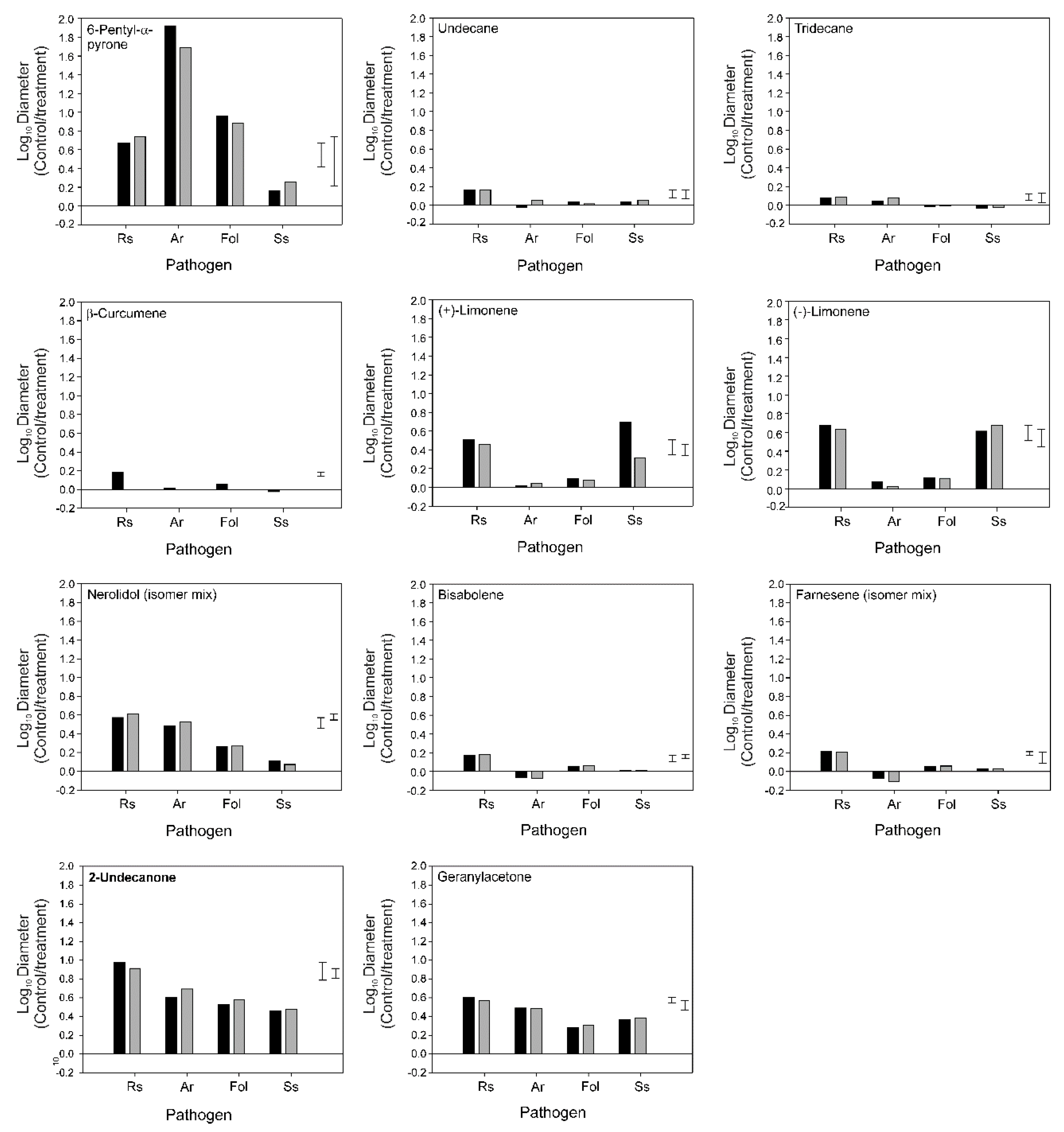
3.4. Bioactivity of Individual VOCs
4. Discussion
5. Conclusions
- Trichoderma sp. “atroviride B” isolates produce qualitatively and quantitatively different VOC blends.
- There is a direct correlation between the amount of VOCs emitted by T. sp. “atroviride B” and the fungistatic activity against R. solani.
- The blend of VOCs produced by Trichoderma has stronger fungistatic activity than the single VOCs tested.
- Plant pathogens respond species-specifically to single Trichoderma VOCs and the whole blend.
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gal-Hemed, I.; Atanasova, L.; Komon-Zelazowska, M.; Druzhinina, I.S.; Viterbo, A.; Yarden, O. Marine isolates of Trichoderma spp. as potential halotolerant agents of biological control for arid-zone agriculture. Appl. Environ. Microbiol. 2011, 77, 5100–5109. [Google Scholar] [CrossRef] [PubMed]
- Błaszczyk, L.; Strakowska, J.; Chełkowski, J.; Gąbka-Buszek, A.; Kaczmarek, J. Trichoderma species occurring on wood with decay symptoms in mountain forests in Central Europe: Genetic and enzymatic characterization. J. Appl. Genet. 2016, 57, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Montoya-Gonzalez, A.H.; Quijano-Vicente, G.; Morales-Maza, A.; Ortiz-Uribe, N.; Hernandez-Martinez, R. Isolation of Trichoderma spp. from Desert Soil, Biocontrol Potential Evaluation and Liquid Culture Production of Conidia Using Agricultural Fertilizers. J. Fertil. Pestic. 2016, 7, 1. [Google Scholar] [CrossRef]
- Kim, K.; Heo, Y.M.; Jang, S.; Lee, H.; Kwon, S.-L.; Park, M.S.; Lim, Y.W.; Kim, J.-J. Diversity of Trichoderma spp. in Marine Environments and Their Biological Potential for Sustainable Industrial Applications. Sustainability 2020, 12, 4327. [Google Scholar] [CrossRef]
- Borut, S.Y.; Johnson, T., Jr. Some biological observations on fungi in estuarine sediments. Mycologia 1962, 54, 181–193. [Google Scholar] [CrossRef]
- Mendoza-Mendoza, A.; Zaid, R.; Lawry, R.; Hermosa, R.; Monte, E.; Horwitz, B.A.; Mukherjee, P.K. Molecular dialogues between Trichoderma and roots: Role of the fungal secretome. Fungal. Biol. Rev. 2018, 32, 62–85. [Google Scholar] [CrossRef]
- Braithwaite, M.; Clouston, A.; Minchin, R.; Yardley, J.; Nieto-Jacobo, M.F.; Mendoza-Mendoza, A.; Steyaert, J.; Hill, R.; Marshall, J.; Stewart, A. The density-dependent effect of initial nematode population levels on the efficacy of Trichoderma as a bio-nematicide against Meloidogyne hapla on tomato. Australas. Plant Path. 2016, 45, 473–479. [Google Scholar] [CrossRef]
- Su, D.; Ding, L.; He, S. Marine-derived Trichoderma species as a promising source of bioactive secondary metabolites. Mini-Rev. Med. Chem. 2018, 18, 1702–1713. [Google Scholar] [CrossRef]
- Poveda, J.; Hermosa, R.; Monte, E.; Nicolás, C. Trichoderma harzianum favours the access of arbuscular mycorrhizal fungi to non-host Brassicaceae roots and increases plant productivity. Sci. Rep. 2019, 9, 11650. [Google Scholar] [CrossRef]
- Lace, B.; Genre, A.; Woo, S.; Faccio, A.; Lorito, M.; Bonfante, P. Gate crashing arbuscular mycorrhizas: In vivo imaging shows the extensive colonization of both symbionts by Trichoderma atroviride. Environ. Microbiol. Rep. 2015, 7, 64–77. [Google Scholar] [CrossRef]
- Druzhinina, I.S.; Seidl-Seiboth, V.; Herrera-Estrella, A.; Horwitz, B.A.; Kenerley, C.M.; Monte, E.; Mukherjee, P.K.; Zeilinger, S.; Grigoriev, I.V.; Kubicek, C.P. Trichoderma: The genomics of opportunistic success. Nat. Rev. Microbiol. 2011, 9, 749–759. [Google Scholar] [CrossRef]
- Geraldine, A.M.; Lopes, F.A.C.; Carvalho, D.D.C.; Barbosa, E.T.; Rodrigues, A.R.; Brandão, R.S.; Ulhoa, C.J.; Lobo Junior, M. Cell wall-degrading enzymes and parasitism of sclerotia are key factors on field biocontrol of white mold by Trichoderma spp. Biol. Control 2013, 67, 308–316. [Google Scholar] [CrossRef]
- Nieto-Jacobo, M.F.; Steyaert, J.M.; Salazar-Badillo, F.B.; Nguyen, D.V.; Rostás, M.; Braithwaite, M.; De Souza, J.T.; Jimenez-Bremont, J.F.; Ohkura, M.; Stewart, A.; et al. Environmental Growth Conditions of Trichoderma spp. Affects Indole Acetic Acid Derivatives, Volatile Organic Compounds, and Plant Growth Promotion. Front. Plant Sci. 2017, 8, 102. [Google Scholar] [CrossRef]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “Secrets” of a Multitalented Biocontrol Agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Cai, F.; Druzhinina, I.S. In honor of John Bissett: Authoritative guidelines on molecular identification of Trichoderma. Fungal Divers. 2021, 107, 1–69. [Google Scholar] [CrossRef]
- Braithwaite, M.; Johnston, P.R.; Ball, S.L.; Nourozi, F.; Hay, A.J.; Shoukouhi, P.; Chomic, A.; Lange, C.; Ohkura, M.; Nieto-Jacobo, M.F.; et al. Trichoderma down under: Species diversity and occurrence of Trichoderma in New Zealand. Australas. Plant Path. 2017, 46, 11–30. [Google Scholar] [CrossRef]
- Mendoza-Mendoza, A.; Nogueira-López, G.; Arizmendi, F.; Cripps-Guazzone, N.; Nieto-Jacobo, M.; Lawry, R.; Kandula, D.; Salazar-Badillo, F.; Salas-Muñoz, S.; Mauricio-Castillo, J. Mechanisms of growth promotion by members of the rhizosphere fungal genus Trichoderma. Adv. PGPR Res. 2017, 1, 1–15. [Google Scholar]
- Daryaei, A.; Jones, E.E.; Ghazalibiglar, H.; Glare, T.R.; Falloon, R.E. Culturing conditions affect biological control activity of Trichoderma atroviride against Rhizoctonia solani in ryegrass. J. Appl. Microbiol. 2016, 121, 461–472. [Google Scholar] [CrossRef]
- McLean, K.L.; Swaminathan, J.; Frampton, C.M.; Hunt, J.S.; Ridgway, H.J.; Stewart, A. Effect of formulation on the rhizosphere competence and biocontrol ability of Trichoderma atroviride C52. Plant Pathol. 2005, 54, 212–218. [Google Scholar] [CrossRef]
- Card, S.D.; Walter, M.; Jaspers, M.V.; Sztejnberg, A.; Stewart, A. Targeted selection of antagonistic microorganisms for control of Botrytis cinerea of strawberry in New Zealand. Australas. Plant Path. 2009, 38, 183–192. [Google Scholar] [CrossRef]
- Maag, D.; Kandula, D.R.W.; Müller, C.; Mendoza-Mendoza, A.; Wratten, S.D.; Stewart, A.; Rostás, M. Trichoderma atroviride LU132 promotes plant growth but not induced systemic resistance to Plutella xylostella in oilseed rape. BioControl 2014, 59, 241–252. [Google Scholar] [CrossRef]
- Yardley, J. Understanding Protoplast Technology as a Tool to Enhance Biocontrol. Ph.D. Thesis, Lincoln University, Lincoln, New Zealand, 2018. [Google Scholar]
- Martínez-Medina, A.; Van Wees, S.C.M.; Pieterse, C.M.J. Airborne signals from Trichoderma fungi stimulate iron uptake responses in roots resulting in priming of jasmonic acid-dependent defences in shoots of Arabidopsis thaliana and Solanum lycopersicum. Plant Cell Environ. 2017, 40, 2691–2705. [Google Scholar] [CrossRef] [PubMed]
- Audrain, B.; Létoffé, S.; Ghigo, J.-M. Airborne Bacterial Interactions: Functions Out of Thin Air? Front. Microbiol. 2015, 6, 1476. [Google Scholar] [CrossRef] [PubMed]
- Garbeva, P.; Weisskopf, L. Airborne medicine: Bacterial volatiles and their influence on plant health. New Phytol. 2020, 226, 32–43. [Google Scholar] [CrossRef]
- Schulz-Bohm, K.; Martín-Sánchez, L.; Garbeva, P. Microbial Volatiles: Small Molecules with an Important Role in Intra- and Inter-Kingdom Interactions. Front. Microbiol. 2017, 8, 2484. [Google Scholar] [CrossRef]
- Cruz-Magalhães, V.; Nieto-Jacobo, M.F.; van Zijll de Jong, E.; Rostás, M.; Padilla-Arizmendi, F.; Kandula, D.; Kandula, J.; Hampton, J.; Herrera-Estrella, A.; Steyaert, J.M.; et al. The NADPH Oxidases Nox1 and Nox2 Differentially Regulate Volatile Organic Compounds, Fungistatic Activity, Plant Growth Promotion and Nutrient Assimilation in Trichoderma atroviride. Front. Microbiol. 2019, 9, 3271. [Google Scholar] [CrossRef]
- Lee, S.; Yap, M.; Behringer, G.; Hung, R.; Bennett, J.W. Volatile organic compounds emitted by Trichoderma species mediate plant growth. Fungal Biol. Biotechnol. 2016, 3, 7. [Google Scholar] [CrossRef]
- Elsherbiny, E.A.; Amin, B.H.; Aleem, B.; Kingsley, K.L.; Bennett, J.W. Trichoderma Volatile Organic Compounds as a Biofumigation Tool against Late Blight Pathogen Phytophthora infestans in Postharvest Potato Tubers. J. Agric. Food Chem. 2020, 68, 8163–8171. [Google Scholar] [CrossRef]
- Sridharan, A.; Thankappan, S.; Karthikeyan, G.; Uthandi, S. Comprehensive profiling of the VOCs of Trichoderma longibrachiatum EF5 while interacting with Sclerotium rolfsii and Macrophomina phaseolina. Microbiol. Res. 2020, 236, 126436. [Google Scholar]
- Jalali, F.; Zafari, D.; Salari, H. Volatile organic compounds of some Trichoderma spp. increase growth and induce salt tolerance in Arabidopsis thaliana. Fungal Ecol. 2017, 29, 67–75. [Google Scholar] [CrossRef]
- Kottb, M.; Gigolashvili, T.; Großkinsky, D.K.; Piechulla, B. Trichoderma volatiles effecting Arabidopsis: From inhibition to protection against phytopathogenic fungi. Front. Microbiol. 2015, 6, 995. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Ghirardo, A.; Weber, B.; Schnitzler, J.P.; Benz, J.P.; Rosenkranz, M. Trichoderma Species Differ in Their Volatile Profiles and in Antagonism Toward Ectomycorrhiza Laccaria bicolor. Front. Microbiol. 2019, 10, 891. [Google Scholar] [CrossRef] [Green Version]
- Kandula, D.R.; Jones, E.E.; Stewart, A.; McLean, K.L.; Hampton, J.G. Trichoderma species for biocontrol of soil-borne plant pathogens of pasture species. Biocontrol. Sci. Techn. 2015, 25, 1052–1069. [Google Scholar] [CrossRef]
- Umar, A.; Kandula, D.R.; Hampton, J.G.; Rolston, M.P.; Chng, S.F. Potential biological control of take-all disease in perennial ryegrass. N. Z. Plant Prot. 2019, 72, 213–220. [Google Scholar] [CrossRef]
- Baskarathevan, J. Botryosphaeriaceous Infection in New Zealand Vineyards: Identification, Population Structure and Genetic Diversity. Ph.D. Thesis, Lincoln University, Lincoln, New Zealand, 2011. [Google Scholar]
- Cruz-Magalhães, V.; Nieto-Jacobo, M.F.; Rostás, M.; Echaide-Aquino, J.F.; Esquivel-Naranjo, E.U.; Stewart, A.; Loguercio, L.L.; Mendoza-Mendoza, A. Histidine kinase two-component response regulators Ssk1, Skn7 and Rim15 differentially control growth, developmental and volatile organic compounds emissions as stress responses in Trichoderma atroviride. Curr. Res. Microb. Sci. 2022, 3, 100139. [Google Scholar] [CrossRef]
- Hohmann, P.; Jones, E.E.; Hill, R.A.; Stewart, A. Understanding Trichoderma in the root system of Pinus radiata: Associations between rhizosphere colonisation and growth promotion for commercially grown seedlings. Fungal Biol. 2011, 115, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Cripps-Guazzone, N.; Jones, E.E.; Condron, L.M.; McLean, K.L.; Stewart, A.; Ridgway, H.J. Rhizosphere and endophytic colonisation of ryegrass and sweet corn roots by the isolate Trichoderma atroviride LU132 at different soil pHs. N. Z. Plant Prot. 2016, 69, 78–85. [Google Scholar] [CrossRef]
- Massawe, V.C.; Hanif, A.; Farzand, A.; Mburu, D.K.; Ochola, S.O.; Wu, L.; Tahir, H.A.S.; Gu, Q.; Wu, H.; Gao, X. Volatile Compounds of Endophytic Bacillus spp. have Biocontrol Activity Against Sclerotinia sclerotiorum. Phytopathology 2018, 108, 1373–1385. [Google Scholar] [CrossRef]
- Wang, C.; Pi, L.; Jiang, S.; Yang, M.; Shu, C.; Zhou, E. ROS and trehalose regulate sclerotial development in Rhizoctonia solani AG-1 IA. Fungal Biol. 2018, 122, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Inamdar, A.A.; Morath, S.; Bennett, J.W. Fungal Volatile Organic Compounds: More Than Just a Funky Smell? Annu. Rev. Microbiol. 2020, 74, 101–116. [Google Scholar] [CrossRef]
- Speckbacher, V.; Ruzsanyi, V.; Wigger, M.; Zeilinger, S. The Trichoderma atroviride Strains P1 and IMI 206040 Differ in Their Light-Response and VOC Production. Molecules 2020, 25, 208. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.W.; Inamdar, A.A. Are Some Fungal Volatile Organic Compounds (VOCs) Mycotoxins? Toxins 2015, 7, 3785–3804. [Google Scholar] [CrossRef] [Green Version]
- Atriztán-Hernández, K.; Moreno-Pedraza, A.; Winkler, R.; Markow, T.; Herrera-Estrella, A. Trichoderma atroviride from Predator to Prey: Role of the Mitogen-Activated Protein Kinase Tmk3 in Fungal Chemical Defense against Fungivory by Drosophila melanogaster Larvae. Appl. Environ. Microbiol. 2019, 85, e01825-18. [Google Scholar] [CrossRef]
- González-Pérez, E.; Ortega-Amaro, M.A.; Salazar-Badillo, F.B.; Bautista, E.; Douterlungne, D.; Jiménez-Bremont, J.F. The Arabidopsis-Trichoderma interaction reveals that the fungal growth medium is an important factor in plant growth induction. Sci. Rep. 2018, 8, 16427. [Google Scholar] [CrossRef]
- Li, N.; Alfiky, A.; Wang, W.; Islam, M.; Nourollahi, K.; Liu, X.; Kang, S. Volatile Compound-Mediated Recognition and Inhibition Between Trichoderma Biocontrol Agents and Fusarium oxysporum. Front. Microbiol. 2018, 9, 2614. [Google Scholar] [CrossRef]
- Cordovez, V.; Mommer, L.; Moisan, K.; Lucas-Barbosa, D.; Pierik, R.; Mumm, R.; Carrion, V.J.; Raaijmakers, J.M. Plant Phenotypic and Transcriptional Changes Induced by Volatiles from the Fungal Root Pathogen Rhizoctonia solani. Front. Plant Sci. 2017, 8, 1262. [Google Scholar] [CrossRef] [PubMed]
- Moisan, K.; Raaijmakers, J.M.; Dicke, M.; Lucas-Barbosa, D.; Cordovez, V. Volatiles from soil-borne fungi affect directional growth of roots. Plant Cell Environ. 2021, 44, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Lazazzara, V.; Vicelli, B.; Bueschl, C.; Parich, A.; Pertot, I.; Schuhmacher, R.; Perazzolli, M. Trichoderma spp. volatile organic compounds protect grapevine plants by activating defense-related processes against downy mildew. Physiol. Plant. 2021, 172, 1950–1965. [Google Scholar] [CrossRef]
- Zhang, F.; Yang, X.; Ran, W.; Shen, Q. Fusarium oxysporum induces the production of proteins and volatile organic compounds by Trichoderma harzianum T-E5. FEMS Microbiol. Lett. 2014, 359, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Inayati, A.; Sulistyowati, L.; Aini, L.Q.; Yusnawan, E. Antifungal activity of volatile organic compounds from Trichoderma virens. AIP Conf. Proc. 2019, 2120, 080012. [Google Scholar] [CrossRef]
- Moreno-Ruiz, D.; Fuchs, A.; Missbach, K.; Schuhmacher, R.; Zeilinger, S. Influence of Different Light Regimes on the Mycoparasitic Activity and 6-Pentyl-α-pyrone Biosynthesis in Two Strains of Trichoderma atroviride. Pathogens 2020, 9, 860. [Google Scholar] [CrossRef] [PubMed]
- Siddiquee, S. Chapter 11—Recent Advancements on the Role and Analysis of Volatile Compounds (VOCs) from Trichoderma. In Biotechnology and Biology of Trichoderma; Gupta, V.K., Schmoll, M., Herrera-Estrella, A., Upadhyay, R.S., Druzhinina, I., Tuohy, M.G., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 139–175. [Google Scholar]
- Hong, J.-H.; Lee, J.; Min, M.; Ryu, S.-M.; Lee, D.; Kim, G.-H.; Kim, J.-J. 6-Pentyl-α-pyrone as an anti-sapstain compound produced by Trichoderma gamsii KUC1747 inhibits the germination of ophiostomatoid fungi. Holzforschung 2014, 68, 769–774. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Replicates | Average Inhibition of Pathogen Growth (%) b | Occurrence of Pathogen Sclerotia (%) | |
|---|---|---|---|---|
| Patented strains | ||||
| LU132 | 40 | 18.30 | 13.5 | ***c |
| LU140 | 40 | 14.40 | 36.8 | *** |
| LU584 | 40 | 5.75 | 25.0 | *** |
| LU633 | 40 | 14.43 | 18.9 | *** |
| Other isolates | ||||
| LU661 | 5 | 35.74 | 60.0 | |
| LU521 | 5 | 33.61 | 0.0 | *** |
| LU723 | 5 | 30.44 | 50.0 | |
| LU532 | 5 | 28.44 | 20.0 | *** |
| LU1308 | 5 | 25.91 | 60.0 | |
| LU141 | 5 | 24.46 | 0.0 | *** |
| LU579 | 5 | 21.51 | 33.3 | * |
| LU666 | 5 | 20.25 | 25.0 | ** |
| LU131 | 5 | 19.81 | 20.0 | *** |
| LU1345 | 5 | 19.10 | 50.0 | |
| LU1357 | 5 | 18.82 | 20.0 | *** |
| LU499 | 5 | 18.51 | 40.0 | * |
| LU522 | 5 | 18.28 | 40.0 | * |
| LU665 | 5 | 17.51 | 0.0 | *** |
| LU572 | 5 | 17.23 | 20.0 | *** |
| LU659 | 5 | 17.13 | 60.0 | |
| LU510 | 5 | 17.11 | 0.0 | *** |
| LU298 | 5 | 16.74 | 33.3 | * |
| LU1341 | 5 | 15.78 | 25.0 | ** |
| LU1330 | 5 | 15.74 | 40.0 | * |
| LU562 | 5 | 15.00 | 0.0 | *** |
| LU497 | 5 | 14.40 | 20.0 | *** |
| LU739 | 5 | 13.70 | 75.0 | |
| LU660 | 5 | 13.32 | 25.0 | ** |
| LU634 | 5 | 12.29 | 20.0 | *** |
| LU668 | 5 | 11.89 | 40.0 | * |
| LU575 | 5 | 11.02 | 50.0 | |
| LU590 | 5 | 10.91 | 0.0 | *** |
| LU577 | 5 | 10.23 | 40.0 | * |
| LU992 | 5 | 9.97 | 20.0 | *** |
| LU568 | 5 | 9.41 | 0.0 | *** |
| LU573 | 5 | 8.48 | 60.0 | |
| LU658 | 5 | 8.24 | 0.0 | *** |
| LU136 | 5 | 8.21 | 20.0 | *** |
| LU563 | 5 | 7.99 | 50.0 | |
| LU987 | 5 | 7.28 | 25.0 | ** |
| LU656 | 5 | 7.24 | 20.0 | *** |
| LU574 | 5 | 6.90 | 20.0 | *** |
| LU725 | 5 | 6.66 | 80.0 | |
| LU578 | 5 | 6.45 | 40.0 | * |
| LU300 | 5 | 6.34 | 20.0 | *** |
| LU587 | 5 | 6.21 | 20.0 | *** |
| LU147 | 5 | 6.02 | 20.0 | *** |
| LU588 | 5 | 5.98 | 40.0 | * |
| LU591 | 5 | 5.93 | 60.0 | |
| LU524 | 5 | 5.78 | 40.0 | * |
| LU582 | 5 | 5.49 | 40.0 | * |
| LU580 | 5 | 5.34 | 40.0 | * |
| LU741 | 5 | 5.15 | 20.0 | *** |
| LU581 | 5 | 5.03 | 20.0 | *** |
| LU589 | 5 | 5.03 | 40.0 | * |
| LU586 | 5 | 5.01 | 25.0 | ** |
| LU583 | 5 | 3.34 | 40.0 | * |
| LU531 | 5 | 1.85 | 20.0 | *** |
| LU657 | 5 | 0.69 | 40.0 | * |
| R. solani alone c | 40 | N/A | 92.5 | |
| LSD (5%) | 5 v. 5 | 8.41 | ||
| 5 v. 40 | 6.31 | |||
| 40 v. 40 | 2.97 | |||
| Treatment | Replicates | Trichoderma Growth (mm, Diameter) at 2 Days | Log10 Days for Trichoderma Colonization (Back Transformed) | Log10 Days for Trichoderma Sporulation (Back Transformed) |
|---|---|---|---|---|
| Patented strains | ||||
| LU132 | 40 | (74.0) a | (0.30) [2.0] | 0.64 [4.4] |
| LU140 | 40 | (74.0) | (0.30) [2.0] | 0.63 [4.3] |
| LU584 | 40 | (74.0) | (0.30) [2.0] | 0.62 [4.2] |
| LU633 | 40 | (74.0) | (0.30) [2.0] | 0.60 [4.0] |
| Top performers | ||||
| LU499 | 5 | (74.0) | (0.30) [2.0] | 1.36 [22.9] |
| LU521 | 5 | (74.0) | (0.30) [2.0] | 0.64 [4.4] |
| LU532 | 5 | 40.2 | 0.67 [4.6] | 0.64 [4.4] |
| LU661 | 5 | (74.0) | (0.30) [2.0] | 0.75 [5.6] |
| LU723 | 5 | 41.2 | 0.58 [3.8] | NS b |
| LU1308 | 5 | (74.0) | (0.30) [2.0] | 0.66 [4.6] |
| Poor performers | ||||
| LU524 | 5 | (74.0) | (0.30) [2.0] | 0.48 [3.0] |
| LU531 | 5 | 18.0 | 1.10 [12.7] | 0.66 [4.6] |
| LU583 | 5 | (74.0) | (0.30) [2.0] | 0.60 [4.0] |
| LU657 | 5 | 15.4 | 0.82 [6.6] | 0.97 [9.3] |
| LU658 | 5 | (74.0) | (0.30) [2.0] | 0.53 [3.4] |
| LSD (5%) | 5 v. 5 | 5.1 | 0.16 | 0.05 |
| 5 v. 40 | 3.8 | 0.12 | 0.04 | |
| 40 v. 40 | 1.8 | 0.06 | 0.02 | |
| LSEffect (5%) | 5 v. 5 | 3.6 | 0.11 | |
| 5 v. 40 | 2.7 | 0.08 | ||
| 40 v. 40 | 1.3 | 0.04 |
| VOC No | Putative Name | Synonym (or Nearest Match for Unknown) | Similarity (%) | Retention Index |
|---|---|---|---|---|
| 1 | 2-Heptanone | 91 | 900 | |
| 2 | 2-Heptanol | 95 | 904 | |
| 3 | Pentyl acetate | Acetic acid, pentyl ester | 97 | 917 |
| 4 | 3-Octanone | 96 | 989 | |
| 5 | (±)-Limonene a | 4-Isopropenyl-1-methylcyclohexene; | 91 | 1032 |
| 6 | Unknown | (4-Hydroxyphenyl)acetonitrile | 83 | 1093 |
| 7 | 2-Nonanone | 96 | 1094 | |
| 8 | Undecane a | 92 | 1099 | |
| 9 | Unknown | 1118 | ||
| 10 | Unknown | 3,7-Dimethyl-6-octenal | 81 | 1156 |
| 11 | 1-Butyl-4-methoxybenzene or | 4-Butylphenyl methyl ether | 91 | 1249 |
| 12 | (S)-(-)-Citronellic acid | Methyl 3,7-dimethyl-6-octenoate | 92 | 1262 |
| 13 | Unknown | 3-Dodecen-1-ol, acetate, (E)- | 83 | 1285 |
| 14 | 2-Undecanone a | 96 | 1295 | |
| 15 | Tridecane a | 95 | 1298 | |
| 16 | Unknown | 1311 | ||
| 17 | Unknown | 1399 | ||
| 18 | α-Bergamotene | 2,6-Dimethyl-6-(4-methyl-3-penten-1-yl)bicyclo [3.1.1]hept-2-ene | 94 | 1444 |
| 19 | Geranylacetone a | 6,10-Dimethyl-5,9-undecadien-2-one | 95 | 1457 |
| 20 | cis-β-Farnesene a | (6Z)-7,11-Dimethyl-3-methylene-1,6,10-dodecatriene | 91 | 1460 |
| 21 | 6-Pentyl-α-pyrone a | 6-Pentyl-2H-pyran-2-one | 92 | 1476 |
| 22 | (-)-Germacrene D | 8-Isopropyl-1-methyl-5-methylene-1,6-cyclodecadiene | 96 | 1496 |
| 23 | Unknown | cis-γ-Bisabolene or β-Curcumene | 90 | 1502 |
| 24 | Unknown | 1512 | ||
| 25 | β-Bisabolene a | (4S)-1-Methyl-4-(6-methyl-1,5-heptadien-2-yl)cyclohexene | 93 | 1517 |
| 26 | β-Sesquiphellandrene | (6S)-3-Methylene-6-[(2S)-6-methyl-5-hepten-2-yl]cyclohexene | 91 | 1534 |
| 27 | Unknown | (E)-6-Pent-1-enylpyran-2-one | 85 | 1540 |
| 28 | trans-Nerolidol a | (6E)-3,7,11-Trimethyl-1,6,10-dodecatrien-3-ol | 96 | 1570 |
| 29 | Methyl trans,cis-farnesate | 6,10-Dodecadienoic acid, 3,7,11-trimethyl-, methyl ester, (E)-(S)- | 92 | 1720 |
| 30 | Unknown | 1995 | ||
| 31 | Verticiol | [1R-(1R*,3E,7E,11R*,12R*)]-4,8,12,15,15-Pentamethyl- bicyclo[9.3.1]pentadeca-3,7-dien-12-ol | 90 | ND b |
| 32 | Unknown | ND b |
| Treatment | Inhibition of Pathogen Growth (%) a | Total Number of VOCs | Total Quantity of VOCs (Peak Area × 107) |
|---|---|---|---|
| LU521 | 78.2 | 28 | 7.1 |
| LU661 | 67.7 | 28 | 8.8 |
| LU132 | 65.8 | 22 | 5.3 |
| LU140 | 65.3 | 22 | 5.3 |
| LU633 | 62.5 | 23 | 5.6 |
| LU657 | 59.9 | 24 | 4.1 |
| LU584 | 53.7 | 23 | 3.7 |
| LU583 | 46.1 | 19 | 3.3 |
| LSD (5%) | 6.7 | 3 | 1.2 |
| VOC No. a | Relative Peak Area (1 × 105) b | |||||||
|---|---|---|---|---|---|---|---|---|
| LU132 | LU140 | LU521 | LU583 | LU584 | LU633 | LU657 | LU661 | |
| 1 | 59.7 ± 10.7 | 60.5 ± 11.7 | 41 ± 5.9 | 32.9 ± 6.7 | 27.9 ± 4.8 | 98.2 ± 22.7 | 36.3 ± 5.8 | 183.0 ± 39.3 |
| 2 | 1.9 ± 1.2 | 2.6 ± 1.2 | ND c | ND | ND | 6.3 ± 1.9 | 1.2 ± 0.7 | 10.9 ± 3.2 |
| 3 | 6.6 ± 2.3 | 4.8 ± 1.5 | ND | 0.2 ± 0.2 | 0.3 ± 0.3 | 0.4 ± 0.4 | 0.5 ± 0.3 | 0.7 ± 0.3 |
| 4 | 3.5 ± 1.9 | 2.5 ± 1.0 | 10 ± 0.5 | 4.0 ± 2.4 | 2.4 ± 0.4 | 6.6 ± 2.1 | 2.9 ± 1.4 | 5.7 ± 1.7 |
| 5 | 2.6 ± 0.2 | 2.8 ± 0.4 | 4 ± 0.3 | 1.2 ± 0.4 | 1.9 ± 0.2 | 2.1 ± 0.2 | ND | 4.0 ± 0.4 |
| 6 | 3.5 ± 1.0 | 3.2 ± 0.8 | 0.7 ± 0.1 | 1.4 ± 0.3 | 1.9 ± 0.6 | 4.2 ± 1.4 | 3.4 ± 0.8 | 7.4 ± 1.7 |
| 7 | 3.6 ± 0.8 | 3.4 ± 0.8 | 0.5 ± 0.1 | 1.8 ± 0.5 | 1.6 ± 0.4 | 4.8 ± 1.2 | 2.2 ± 0.3 | 9.9 ± 2.2 |
| 8 | 1.2 ± 0.5 | 1.2 ± 0.4 | 1.5 ± 0.4 | 0.8 ± 0.4 | 1.0 ± 0.3 | 1.1 ± 0.4 | 1.0 ± 0.4 | 1.8 ± 0.7 |
| 9 | 28.5 ± 4.6 | 22.4 ± 4.8 | 111.1 ± 18.2 | 5.4 ± 1.1 | 12.7 ± 2.9 | 17.8 ± 4.4 | 30.8 ± 5.8 | 96.5 ± 16.4 |
| 10 | ND | ND | ND | ND | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.4 ± 0.2 | 0.4 ± 0.2 |
| 11 | 0.1 ± 0.1 | ND | 0.3 ± 0.09 | 0.1 ± 0.1 | 0.3 ± 0.1 | ND | 0.1 ± 0.1 | 0.5 ± 0.1 |
| 12 | ND | ND | 1.1 ± 0.7 | ND | ND | ND | ND | ND |
| 13 | ND | ND | ND | ND | ND | ND | ND | 0.4 ± 0.2 |
| 14 | ND | ND | 6.4 ± 0.5 | 0.02 ± 0.02 | 0.02 ± 0.02 | 1.4 ± 0.3 | 0.2 ± 0.2 | 2.8 ± 0.4 |
| 15 | 3.8 ± 1.3 | 4.0 ± 1.2 | 3.8 ± 1.7 | 2.2 ± 1.0 | 2.1 ± 0.8 | 2.7 ± 0.7 | 2.5 ± 0.6 | 4.8 ± 1.7 |
| 16 | ND | ND | 1.3 ± 0.1 | ND | ND | ND | ND | 1.2 ± 0.3 |
| 17 | ND | 0.1 ± 0.1 | 0.12 ± 0.1 | ND | ND | ND | 0.1 ± 0.1 | 0.2 ± 0.1 |
| 18 | 6.9 ± 1.3 | 6.5 ± 0.9 | 34.0 ± 9.9 | 1.2 ± 0.2 | 1.5 ± 0.3 | 3.0 ± 0.4 | 1.2 ± 0.4 | 24.4 ± 6.6 |
| 19 | 0.4 ± 0.1 | 0.4 ± 0.1 | 1.3 ± 0.1 | ND | 0.2 ± 0.1 | 0.5 ± 0.1 | 1.5 ± 0.2 | 1.9 ± 0.4 |
| 20 | 1.2 ± 0.3 | 1.2 ± 0.3 | 3.0 ± 0.3 | ND | 0.4 ± 0.3 | 0.6 ± 0.3 | ND | 3.6 ± 1.1 |
| 21 | 390.2 ± 59.2 | 394.3 ± 49.1 | 456.2 ± 65.5 | 273.5 ± 42.4 | 300.9 ± 51.1 | 398.0 ± 58.7 | 308.0 ± 36.6 | 489.6 ± 68.7 |
| 22 | 2.2 ± 0.6 | 2.1 ± 0.5 | 2.2 ± 0.6 | 1.3 ± 0.4 | 1.8 ± 0.5 | 1.1 ± 0.7 | 2.8 ± 1.0 | 3.7 ± 0.6 |
| 23 | 2.5 ± 0.5 | 3.1 ± 0.2 | 8.6 ± 0.8 | 2.0 ± 1.0 | 1.8 ± 0.6 | 3.6 ± 0.8 | 3.4 ± 0.9 | 4.2 ± 0.8 |
| 24 | 1.5 ± 0.3 | 1.6 ± 0.2 | 1.8 ± 0.3 | 0.6 ± 0.3 | 0.9 ± 0.3 | 1.1 ± 0.2 | 1.4 ± 0.2 | 2.8 ± 0.6 |
| 25 | 0.8 ± 0.1 | 0.8 ± 0.1 | 2.8 ± 0.3 | 0.3 ± 0.2 | 0.3 ± 0.2 | 0.4 ± 0.2 | 0.3 ± 0.2 | 1.4 ± 0.3 |
| 26 | ND | ND | 0.4 ± 0.1 | ND | ND | ND | 0.8 ± 0.1 | ND |
| 27 | 6.8 ± 0.8 | 7.1 ± 0.6 | 7.5 ± 0.6 | 4.2 ± 0.5 | 5.1 ± 0.9 | 7.5 ± 0.8 | 6.4 ± 0.3 | 10.6 ± 0.7 |
| 28 | 0.7 ± 0.1 | 0.7 ± 0.1 | 2.1 ± 0.1 | 0.2 ± 0.1 | 0.6 ± 0.2 | 0.5 ± 0.1 | 0.9 ± 0.2 | 1.9 ± 0.2 |
| 29 | ND | ND | 0.7 ± 0.0 | ND | ND | ND | ND | ND |
| 30 | ND | ND | 1.1 ± 0.2 | ND | ND | ND | ND | 0.4 ± 0.1 |
| 31 | 0.8 ± 0.3 | 0.7 ± 0.1 | 13.0 ± 2.7 | ND | 0.5 ± 0.2 | 0.8 ± 0.3 | 1.2 ± 0.1 | 4.6 ± 1.1 |
| 32 | ND | ND | 0.3 ± 0.1 | ND | ND | ND | ND | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Zijll de Jong, E.; Kandula, J.; Rostás, M.; Kandula, D.; Hampton, J.; Mendoza-Mendoza, A. Fungistatic Activity Mediated by Volatile Organic Compounds Is Isolate-Dependent in Trichoderma sp. “atroviride B”. J. Fungi 2023, 9, 238. https://doi.org/10.3390/jof9020238
van Zijll de Jong E, Kandula J, Rostás M, Kandula D, Hampton J, Mendoza-Mendoza A. Fungistatic Activity Mediated by Volatile Organic Compounds Is Isolate-Dependent in Trichoderma sp. “atroviride B”. Journal of Fungi. 2023; 9(2):238. https://doi.org/10.3390/jof9020238
Chicago/Turabian Stylevan Zijll de Jong, Eline, Janaki Kandula, Michael Rostás, Diwakar Kandula, John Hampton, and Artemio Mendoza-Mendoza. 2023. "Fungistatic Activity Mediated by Volatile Organic Compounds Is Isolate-Dependent in Trichoderma sp. “atroviride B”" Journal of Fungi 9, no. 2: 238. https://doi.org/10.3390/jof9020238