
Effect of Farnesol in Trichoderma Physiology and in Fungal–Plant Interaction
 , , , , , , and
, , , , , , and
Abstract
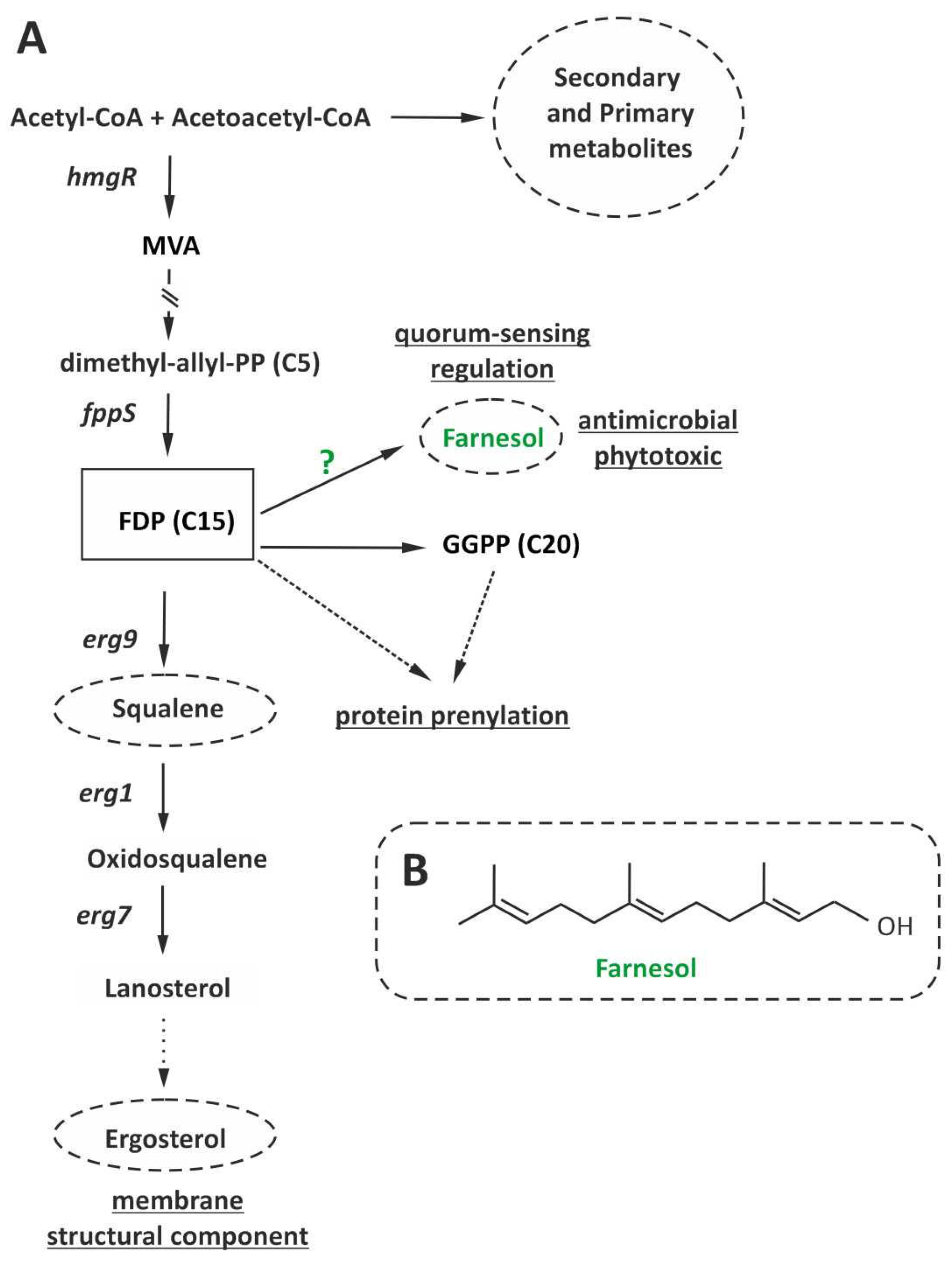
:1. Introduction
2. Materials and Methods
2.1. Strains Used and Culture Conditions
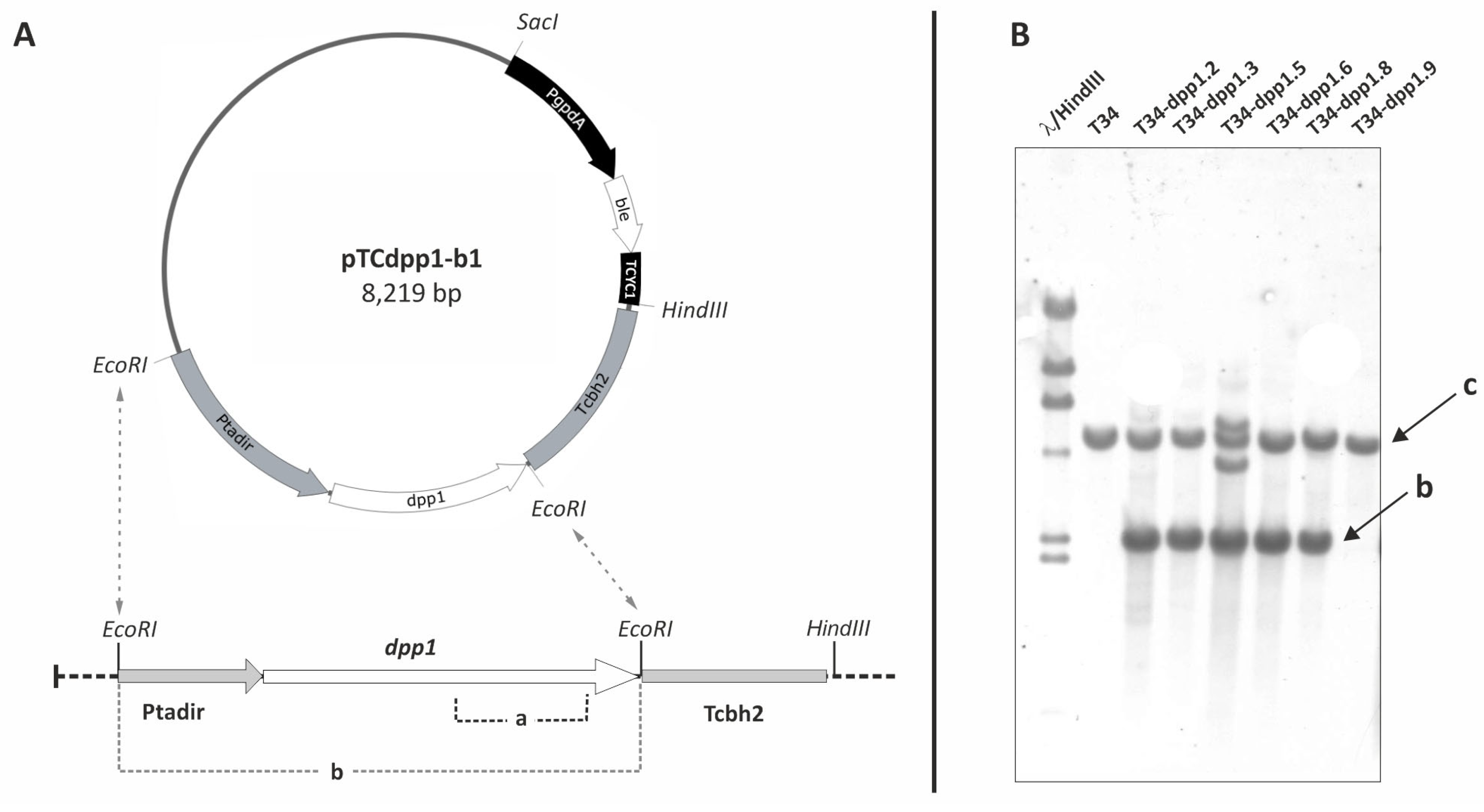
2.2. Overexpression of dpp1 Gene in T. harzianum CECT 2413 (T34)
2.3. Transformation of pTCdpp1-b1 Plasmid in T. harzianum T34
2.4. DNA Probes Labeling and Southern Hybridization
2.5. Real-Time qPCR (qPCR) Analysis
2.6. Metabolomic Characterization
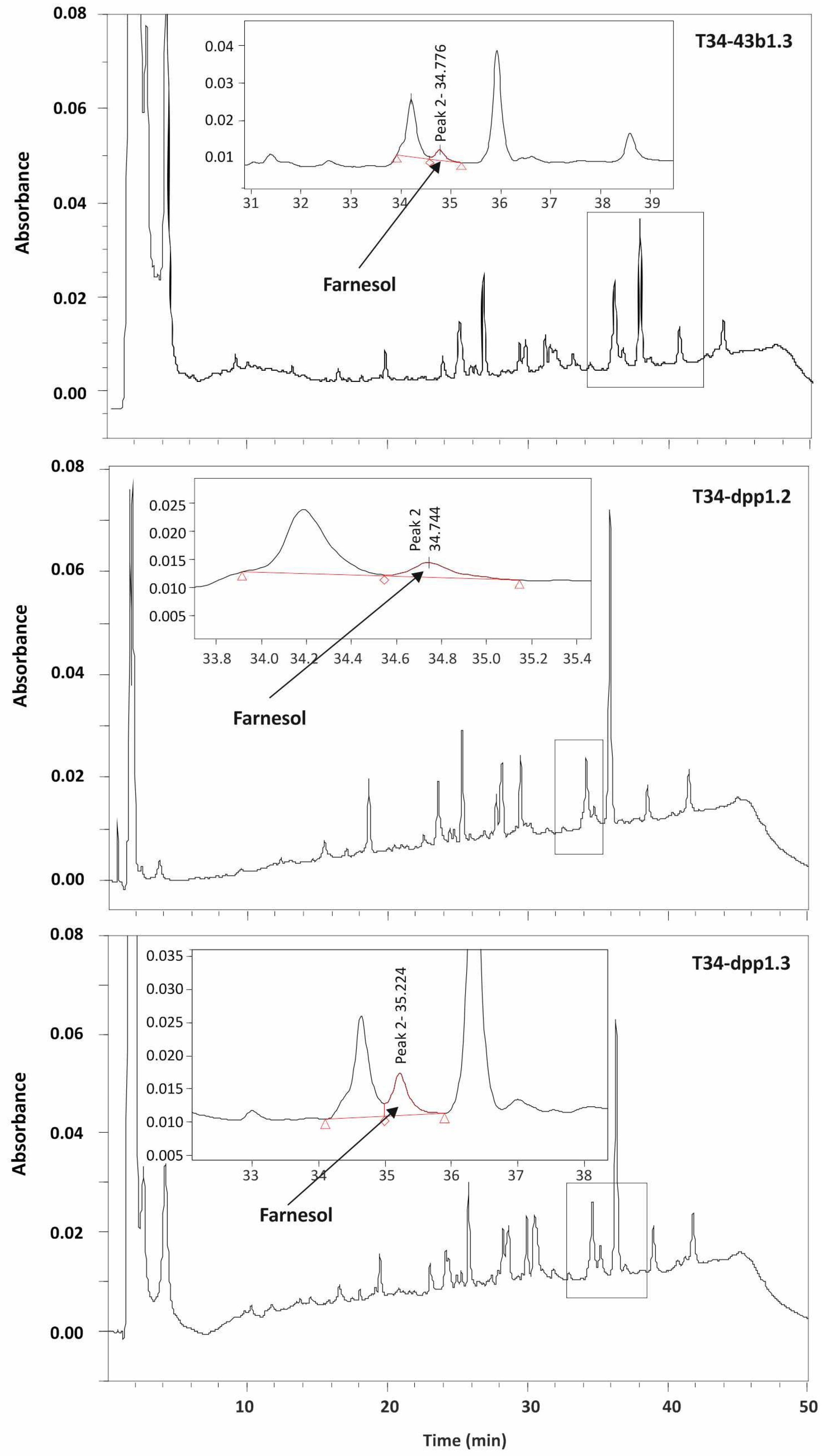
2.6.1. Quantification of Farnesol
2.6.2. Quantification of Ergosterol and Squalene
2.7. Antifungal Assays
2.7.1. Antifungal Assays on Tomato Leaves
2.7.2. Antifungal Assays on Membranes against Rhizoctonia solani
2.8. Tomato Plant Assays
2.8.1. In Vitro Analysis to Determine the Effect of Exogenous Farnesol on Tomato Growth and on Expression of Plant Defense- and Development-Related Genes
2.8.2. Inoculation of Tomato Seeds with Trichoderma Spores, Plant Growth and Infection with Botrytis cinerea B05.10
2.8.3. Tomato-Trichoderma Hydroponic Cultures for Analysis of the Effect of dpp1 Overexpression on the Ability to Colonize Tomato Roots
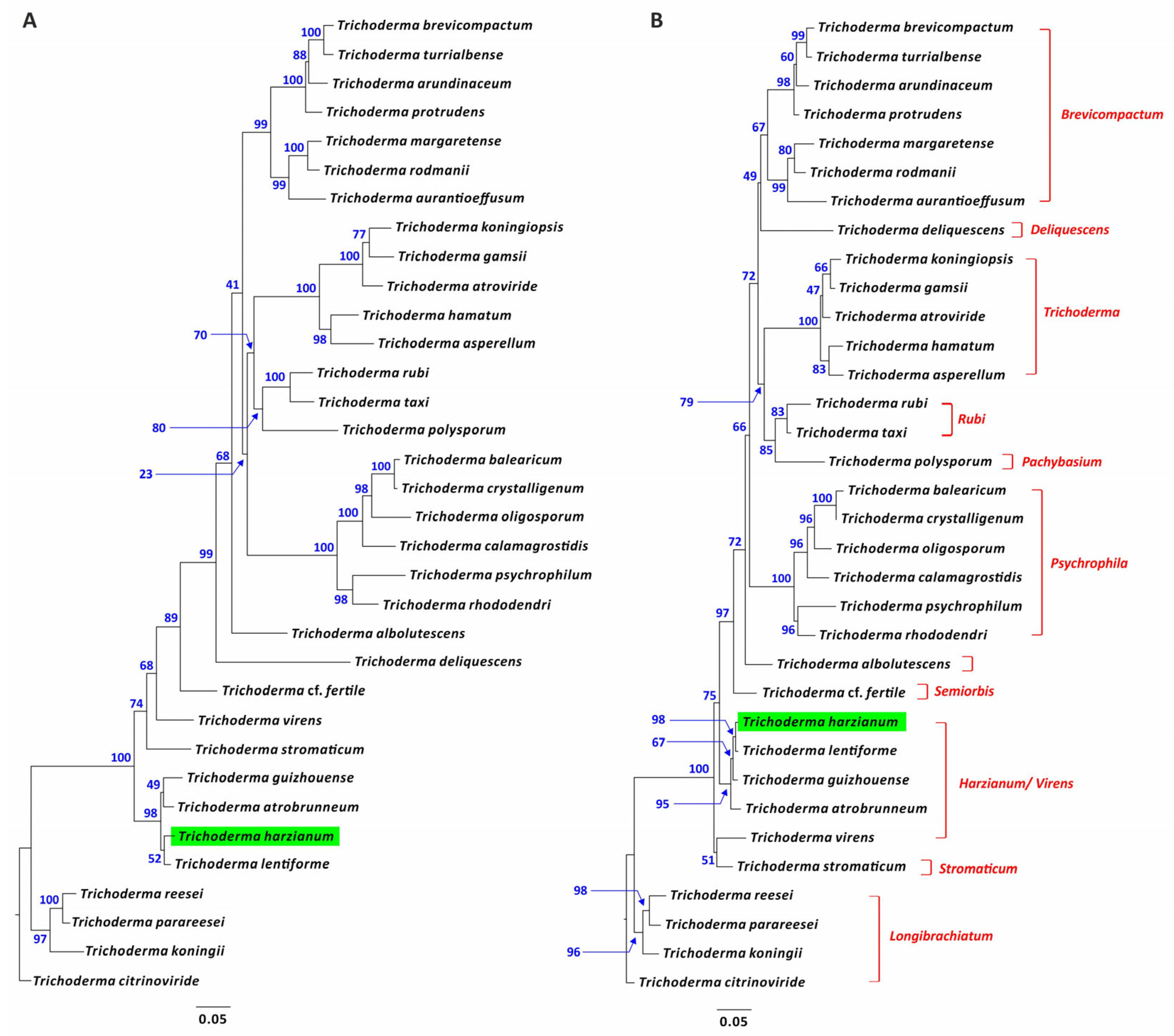
2.9. Phylogenetic Analysis
3. Results
3.1. Identification of T. harzianum Homologs of DPP1/LPP1
3.2. Effect of Overexpression of T. harzianum dpp1 on Fungal Growth
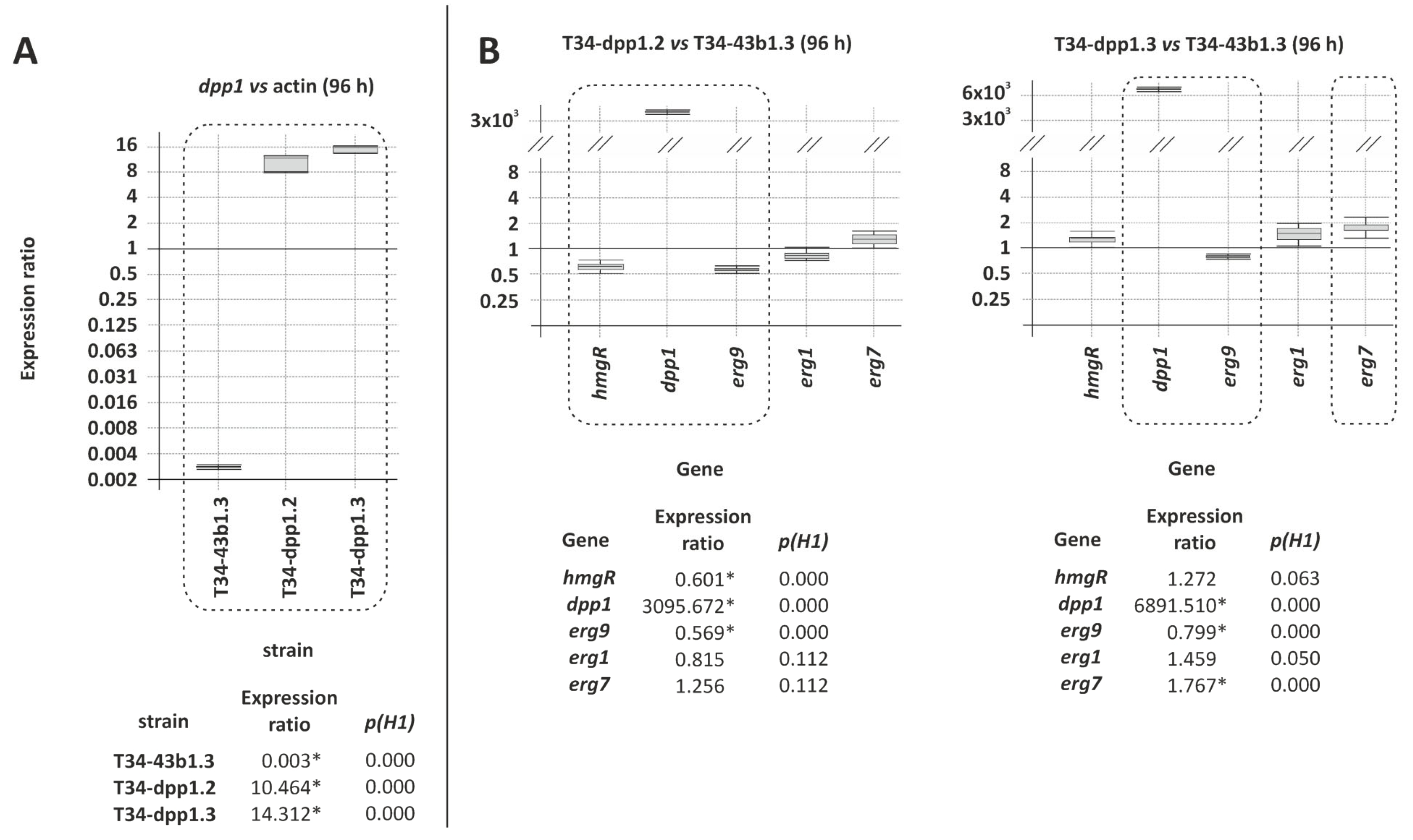
3.3. RT-qPCR Analysis of T. harzianum Terpene-Related Gene Expression
3.3.1. Expression of dpp1 in T34 Transformants with Plasmid pTCdpp1-b1
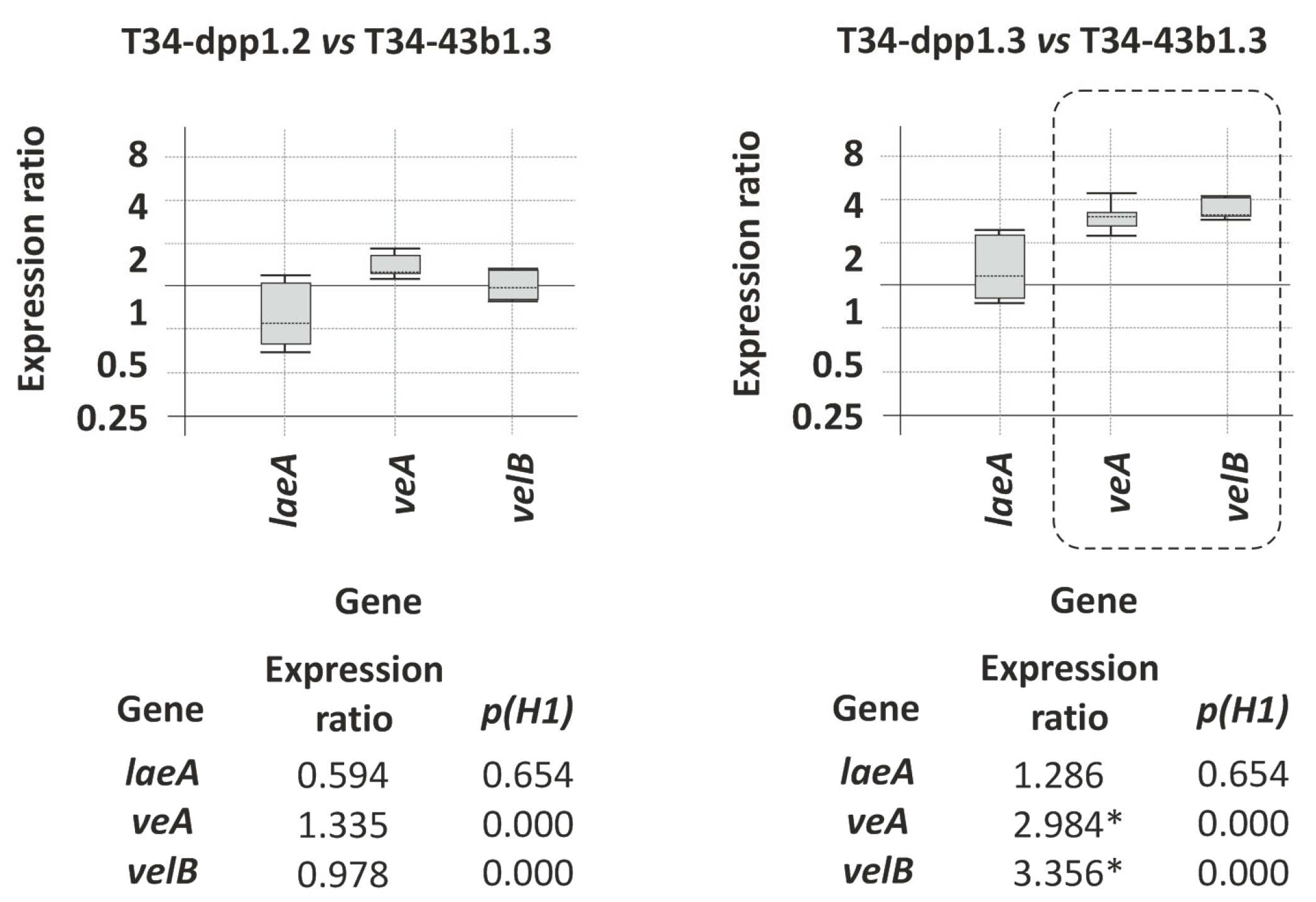
3.3.2. Effect of dpp1 Overexpression on the Transcription Level of Other Terpene Biosynthetic Genes
3.4. Metabolomic Characterization of the T. harzianum dpp1-Overexpressed Transformants
3.4.1. Production of Farnesol by dpp1-Overexpressed Transformants
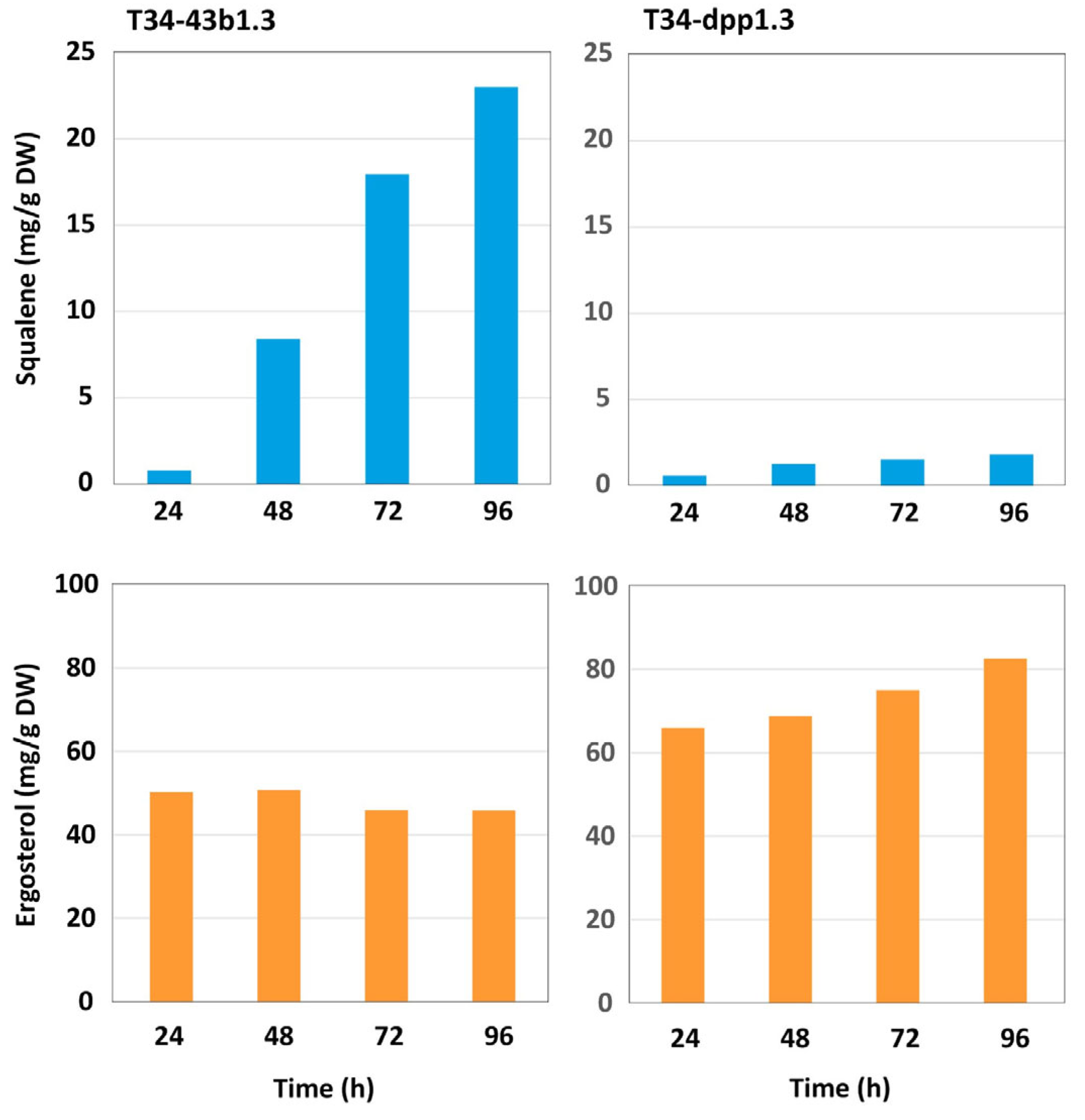
3.4.2. Production of Squalene and Ergosterol in dpp1-Overexpressing Transformants
3.5. Effect of External Addition of Farnesol on the Squalene-Ergosterol Levels in T34-43b1.3 versus T34-dpp1.3
3.6. Importance of Farnesol in the Regulation of Circadian Cycle-Related Genes in T. harzianum
3.7. Effect of the Increase in Farnesol Production on the Fungal Phenotype
3.8. Effect of dpp1 Overexpression on Trichoderma–Plant Interactions
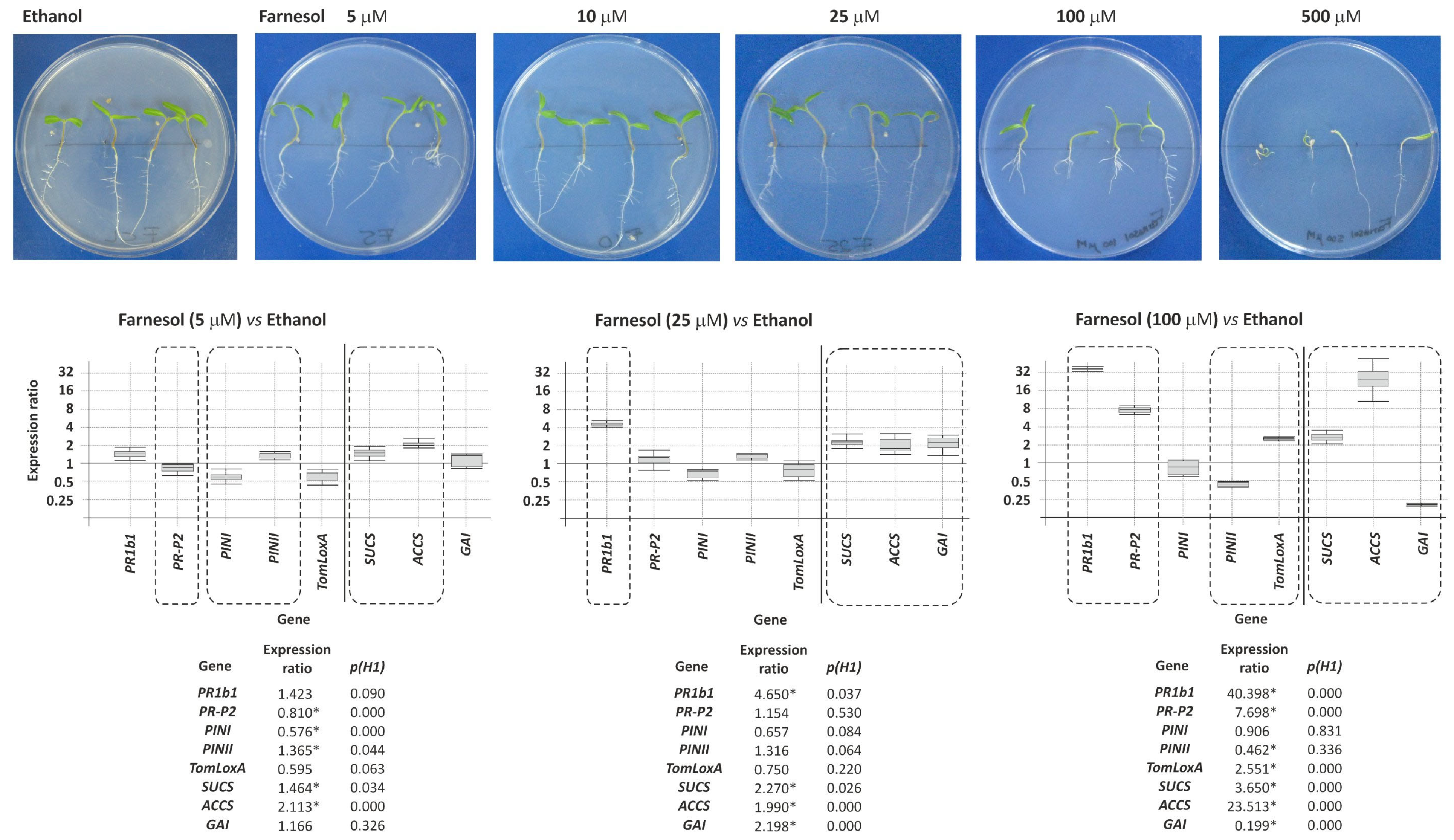
3.8.1. Effect of Exogenous Farnesol on Tomato Plants
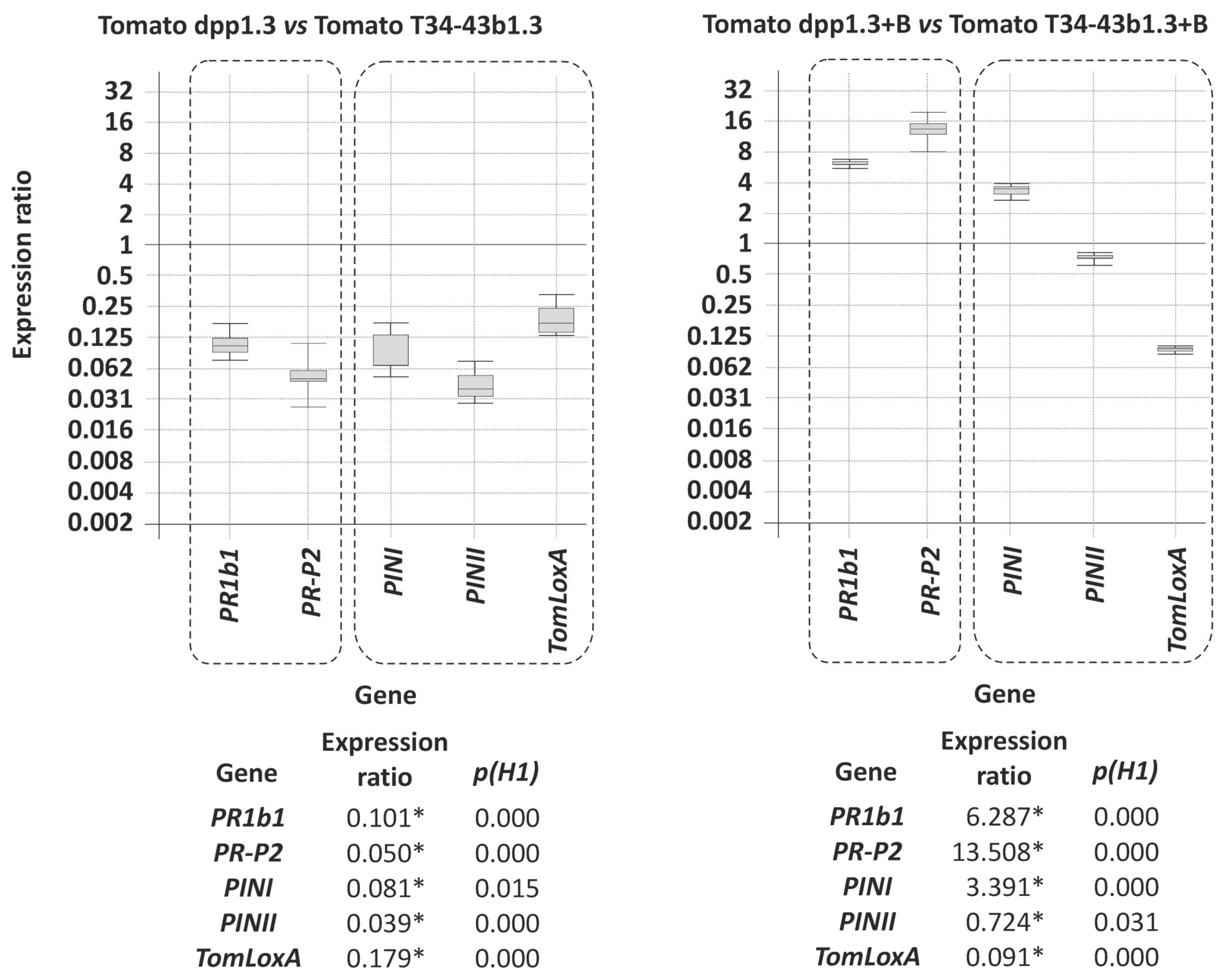
3.8.2. Effect of dpp1 Overexpression in T. harzianum on Plant Development
3.8.3. Effect of dpp1 Overexpression on Resistance of Tomato Plants to Botrytis cinerea
3.8.4. Effect of Increased Farnesol Production on Root Colonization
3.9. Effect of dpp1 Overexpression on the Trichoderma Antifungal Activity
3.9.1. Antifungal Activity against B. cinerea B05.10 on Tomato Leaves
3.9.2. Antifungal Assays on Membranes against R. solani
3.10. Phylogenetic Analysis of Trichoderma dpp1 Orthologs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Hermosa, H.; Cardoza, R.E.; Rubio, M.B.; Gutiérrez, S.; Monte, E. Secondary metabolism and antimicrobial metabolites of Trichoderma. In Biotechnology and Biology of Trichoderma; Gupka, V.K., Schmoll, M., Herrera-Estrella, A., Upadhyay, R.S., Druzhinina, I., Tuohy, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; Chapter 10; pp. 125–137. [Google Scholar]
- Sivasithamparam, K.; Ghisalberti, E.L. Secondary metabolism in Trichoderma and Gliocladium. In Trichoderma and Glicocladium; Harman, G.E., Hubicek, C.P., Eds.; Taylor & Francis: London, UK, 1998; Volume 1, pp. 139–191. [Google Scholar]
- Cardoza, R.E.; Vizcaíno, J.A.; Hermosa, M.R.; Sousa, S.; González, F.J.; Llobell, A.; Monte, E.; Gutiérrez, S. Cloning and characterization of the erg1 gene of Trichoderma harzianum: Effect of the erg1 silencing on ergosterol biosynthesis and resistance to terbinafine. Fungal Genet. Biol. 2006, 43, 164–178. [Google Scholar] [CrossRef]
- Cardoza, R.E.; Hermosa, M.R.; Vizcaíno, J.A.; González, F.; Llobell, A.; Monte, E.; Gutiérrez, S. Partial silencing of a hydroxy-methylglutaryl-CoA reductase-encoding gene in Trichoderma harzianum CECT 2413 results in a lower level of resistance to lovastatin and lower antifungal activity. Fungal Genet. Biol. 2007, 44, 269–283. [Google Scholar] [CrossRef]
- Cardoza, R.E.; Malmierca, M.G.; Gutiérrez, S. Overexpression of erg1 gene in Trichoderma harzianum CECT 2413: Effect on the induction of tomato defence-related genes. J. Appl. Microbiol. 2014, 117, 812–823. [Google Scholar] [CrossRef] [PubMed]
- Haney, S.L.; Wills, V.S.; Wiemer, D.F.; Holstein, S.A. Recent advances in the development of mammalian geranylgeranyl diphosphate synthase inhibitors. Molecules 2017, 22, 886. [Google Scholar] [CrossRef] [Green Version]
- Kruppa, M. Quorum sensing and Candida albicans. Mycoses 2008, 52, 1–10. [Google Scholar] [CrossRef]
- Nickerson, K.W.; Atkin, A.L.; Hornby, J.M. Quorum sensing in dimorphic fungi: Farnesol and beyond. Appl. Environ. Microbiol. 2006, 72, 3805–3813. [Google Scholar] [CrossRef] [Green Version]
- Langford, M.L.; Hasim, S.; Nickerson, K.W.; Atkin, A.L. Activity and toxicity of farnesol towards Candida albicans are dependent on growth conditions. Antimicrob. Agents Chemother. 2010, 54, 940–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oshiro, J.; Han, G.-S.; Caramn, G.M. Diacylglycerol pyrophosphate phosphatase in Saccharomyces cerevisiae. Biochimica et Biophysica Acta. 2003, 1635, 1–9. [Google Scholar] [CrossRef]
- Toke, D.A.; Bennett, W.L.; Oshiro, J.; Wu, W.-I.; Voelker, D.R.; Carman, G.M. Isolation and characterization of the Saccharomyces cerevisiae LPP1 gene encoding a Mg2+-independent phosphatidate phosphatase. J. Biol. Chem. 1998, 273, 14331–14338. [Google Scholar] [CrossRef] [Green Version]
- Toke, D.A.; Bennett, W.L.; Sillon, D.A.; Wu, W.-I.; Chen, X.; Ostrander, D.B.; Oshiro, J.; Cremesti, A.; Voelker, D.R.; Fischl, A.S.; et al. Isolation and characterization of the Saccharomyces cerevisiae DPP1 gene encoding diacylglycerol pyrophosphate phosphatase. J. Biol. Chem. 1998, 273, 3278–3284. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, A.; Chen, X.; Rush, J.; Horazdowsky, B.; Waechter, C.J.; Carman, G.M.; Sternweis, P.C. The LPP1 and DPP1 gene products account for most of the isoprenoid phosphate phosphatase activities in Saccharomyces cerevisiae. J. Biol. Chem. 1999, 274, 14831–14837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindo, L.; McCormick, S.P.; Cardoza, R.E.; Kim, H.-S.; Brown, D.W.; Alexander, N.J.; Proctor, R.H.; Gutiérrez, S. Role of Trichoderma arundinaceum tri10 in regulation of terpene biosynthetic genes and in control of metabolic flux. Fungal Genet. Biol. 2019, 122, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Ryder, N.S.; Dupont, M.-C. Inhibition of squalene epoxidase by allylamine antimycotic compounds. A comparative study of the fungal and mammalian enzymes. Biochem. J. 1985, 230, 765–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornby, J.M.; Kebaara, B.W.; Nickerson, K.W. Farnesol biosynthesis in Candida albicans: Cellular response to sterol inhibition by zaragozic acid B. Antimicrob. Agents Chemother. 2003, 47, 2366–2369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarathna, D.H.M.L.P.; Hornby, J.M.; Hoerrmann, N.; Parkhurst, A.M.; Duhamel, G.E.; Nickerson, K.W. Enhanced pathogenicity of Candida albicans pre-treated with subinhibitory concentrations of fluconazole in a mouse model of disseminated candidiasis. J. Antimicrob. Chemother. 2005, 56, 1156–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penttilä, M.; Nevalainen, H.; Rättö, M.; Salminen, E.; Knowles, J. A versatile transformation system for the cellulotytic filamentous fungus Trichoderma reesei. Gene 1987, 61, 155–164. [Google Scholar] [CrossRef]
- Cardoza, R.E.; McCormick, S.P.; Malmierca, M.G.; Olivera, E.R.; Alexander, N.J.; Monte, E.; Gutiérrez, S. Effect of trichothecene production of plant defense response and on fungal physiology: Overexpression of Trichoderma arundinaceum tri4 gene in T. harzianum. Appl. Environ. Microbiol. 2015, 81, 6355–6366. [Google Scholar] [CrossRef] [Green Version]
- Cardoza, R.E.; Vizcaíno, J.A.; Hermosa, M.R.; Monte, E.; Gutiérrez, S. A comparison of the phenotypic and genetic stability of recombinant Trichoderma spp. generated by protoplast- and Agrobacterium-mediated transformation. J. Microbiol. 2006, 44, 383–395. [Google Scholar]
- Lindo, L.; Cardoza, R.E.; Lorenzana, A.; Casquero, P.A.; Gutiérrez, S. Identification of plant genes putatively involved in the perception of fungal ergosterol-squalene. J. Integr. Plant Biol. 2020, 62, 927–947. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST©) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acid Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Tucci, M.; Ruocco, M.; De Masi, L.; De Palma, M.; Lorito, M. The beneficial effect of Trichoderma spp. on tomato is modulated by the plant genotype. Mol. Plant Pathol. 2011, 12, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, B.B.; Gruissem, W.; Jones, R.L. Biochemistry and Molecular Biology of Plants; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Bradford, M.M. Rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Ghimire, G.P.; Lee, H.C.; Sohng, J.K. Improved squalene production via modulation of the methylerythritol 4-phosphate pathway and heterologous expression of genes from Streptomyces peucetius ATCC 27952 in Escherichia coli. Appl. Environ. Microbiol. 2009, 75, 7291–7293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malmierca, M.G.; Cardoza, R.E.; Alexander, N.J.; McCormick, S.P.; Hermosa, R.; Monte, E.; Gutiérrez, S. Involvement of Trichoderma trichothecenes in the biocontrol activity and induction of plant defense-related genes. Appl. Environ. Microbiol. 2012, 78, 4856–4868. [Google Scholar] [CrossRef] [Green Version]
- Malmierca, M.G.; Barua, J.; McCormick, S.P.; Izquierdo-Bueno, I.; Cardoza, R.E.; Alexander, N.J.; Hermosa, R.; Collado, I.G.; Monte, E.; Gutiérrez, S. Novel aspinolide production by Trichoderma arundinaceum with a potential role in Botrytis cinerea antagonistic activity and plant defence priming. Environ. Microbiol. 2015, 17, 1103–1118. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Bnyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2014, 32, 268–274. [Google Scholar] [CrossRef]
- Gutiérrez, S.; McCormick, S.P.; Cardoza, R.E.; Kim, H.-S.; Lindo-Yugueros, L.; Vaughan, M.M.; Carro-Huerga, G.; Busman, M.; Sáenz de Miera, L.E.; Jaklitsch, W.M.; et al. Distribution, function, and evolution of a gene essential for trichothecene toxin biosynthesis in Trichoderma. Front. Microbiol. 2021, 12, 791641. [Google Scholar] [CrossRef]
- Brakhage, A.A. Regulation of fungal secondary metabolism. Nat. Rev. Microbiol. 2013, 11, 21–32. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Steindorff, A.S.; Chenthamara, K.; Manganiello, G.; Henrissat, B.; Zhang, J.; Cai, F.; Kopchinskiy, A.G.; Kubicek, E.M.; Kuo, A.; et al. Evolution and comparative genomics of the most common Trichoderma species. BMC Genomics. 2019, 20, 485. [Google Scholar] [CrossRef] [Green Version]
- Paradise, E.M.; Kirby, J.; Chan, R.; Keasling, J.D. Redirection of flux through the FPP branch-point in Saccharomyces cerevisiae by down-regulating squalene synthase. Biotechnol. Bioeng. 2008, 100, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Szkopinska, A.; Grabinska, K.; Delourme, D.; Karst, F.; Rytka, J.; Palamarczyk, G. Polyprenol formation in the yeast Saccharomyces cerevisiae: Effect of farnesyl diphosphate synthase overexpression. J. Lipid. Res. 1997, 38, 962–968. [Google Scholar] [CrossRef] [PubMed]
- Song, L. Detection of farnesyl diphosphate accumulation in yeast ERG9 mutants. Anal. Biochem. 2003, 317, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Enjalbert, B.; Whiteway, M. Release from quorum-sensing molecules triggers hyphal formation during Candida albicans resumption of growth. Eukaryot. Cell. 2005, 4, 1203–1210. [Google Scholar] [CrossRef] [Green Version]
- Mayo-Prieto, S.; Izquierdo, H.; González, O.; Rodríguez-González, A.; Lorenzana, A.; Gutiérrez, S.; Casquero, P.A. Effect of farnesol, a compound produced by Trichoderma when growing on bean (Phaseolus vulgaris L.). Planta Med. 2016, 81, S1–S381. [Google Scholar] [CrossRef]
- Malmierca, M.G.; Cardoza, R.E.; Alexander, N.J.; McCormick, S.P.; Collado, I.G.; Hermosa, R.; Monte, E.; Gutiérrez, S. Relevance of trichothecenes in fungal physiology: Disruption of tri5 in Trichoderma arundinaceum. Fungal Genet. Biol. 2013, 53, 22–33. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Concentration (μg/mL) | Specific Concentration (μg/mg protein) | % Variation |
|---|---|---|---|
| T34 | 0.0444 | 0.0037 | - |
| T34-dpp1.2 | 0.0811 | 0.0061 | +64.86% |
| T34-dpp1.3 | 0.4983 | 0.0358 | +867.57% |
| Sample | Concentration (μg/mL) | Specific Concentration (μg/mg Dry Weight) | % Variation |
|---|---|---|---|
| T34 | 0.6102 | 0.0122 | - |
| T34-dpp1.2 | 0.8230 | 0.0165 | +35.24% |
| T34-dpp1.3 | 1.9278 | 0.0386 | +216.40% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardoza, R.E.; McCormick, S.P.; Lindo, L.; Mayo-Prieto, S.; González-Cazón, D.; Martínez-Reyes, N.; Carro-Huerga, G.; Rodríguez-González, Á.; Proctor, R.H.; Casquero, P.A.; et al. Effect of Farnesol in Trichoderma Physiology and in Fungal–Plant Interaction. J. Fungi 2022, 8, 1266. https://doi.org/10.3390/jof8121266
Cardoza RE, McCormick SP, Lindo L, Mayo-Prieto S, González-Cazón D, Martínez-Reyes N, Carro-Huerga G, Rodríguez-González Á, Proctor RH, Casquero PA, et al. Effect of Farnesol in Trichoderma Physiology and in Fungal–Plant Interaction. Journal of Fungi. 2022; 8(12):1266. https://doi.org/10.3390/jof8121266
Chicago/Turabian StyleCardoza, Rosa E., Susan P. McCormick, Laura Lindo, Sara Mayo-Prieto, David González-Cazón, Natalia Martínez-Reyes, Guzmán Carro-Huerga, Álvaro Rodríguez-González, Robert H. Proctor, Pedro A. Casquero, and et al. 2022. "Effect of Farnesol in Trichoderma Physiology and in Fungal–Plant Interaction" Journal of Fungi 8, no. 12: 1266. https://doi.org/10.3390/jof8121266