
Beta Diversity of Arbuscular Mycorrhizal Communities Increases in Time after Crop Establishment of Peruvian Sacha Inchi (Plukenetia volubilis)
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
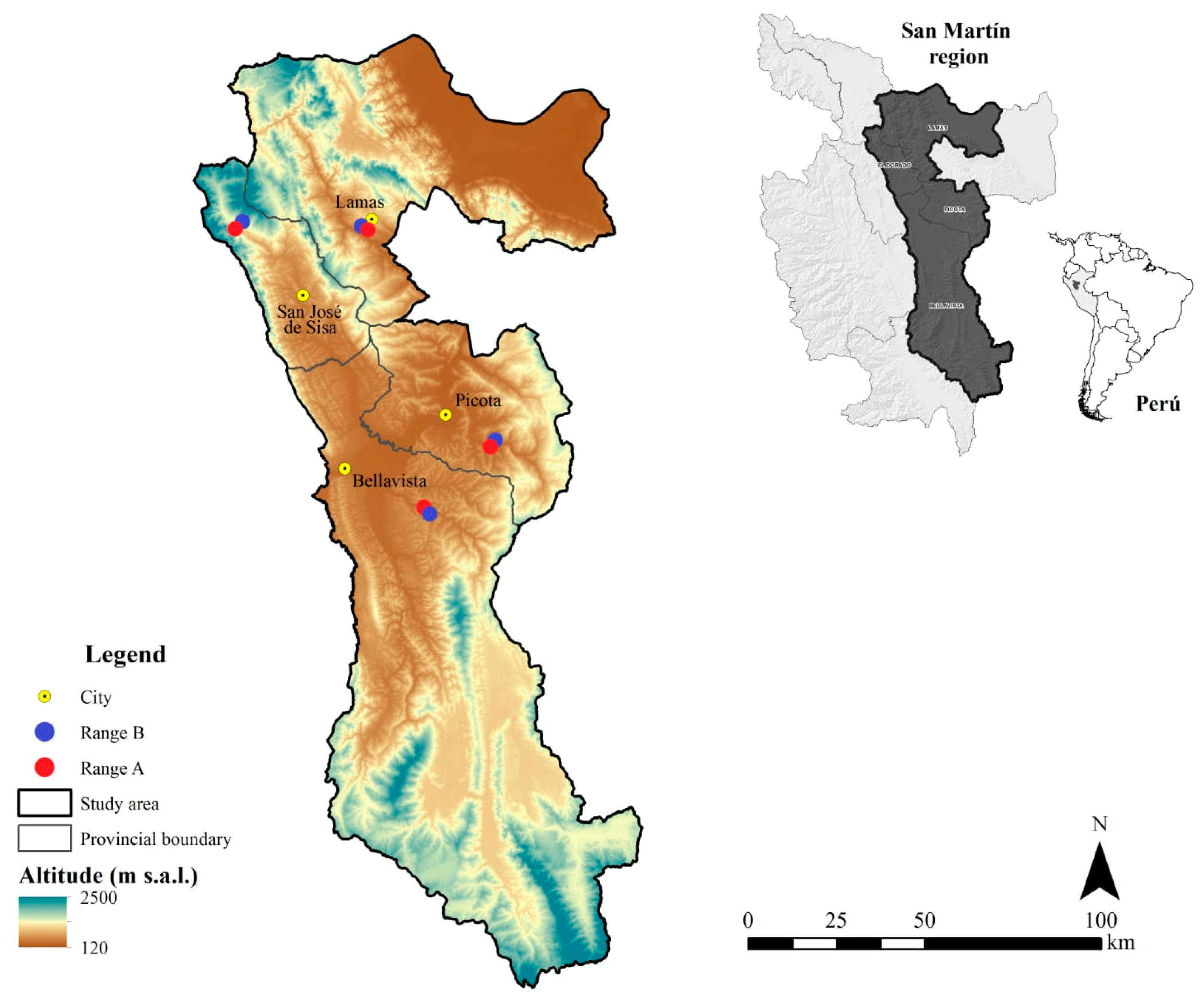
2.1. Study Place
2.2. Sampling Design
2.3. Soil Analysis
2.4. Root Molecular Analysis
2.5. Statistical Analyses
3. Results
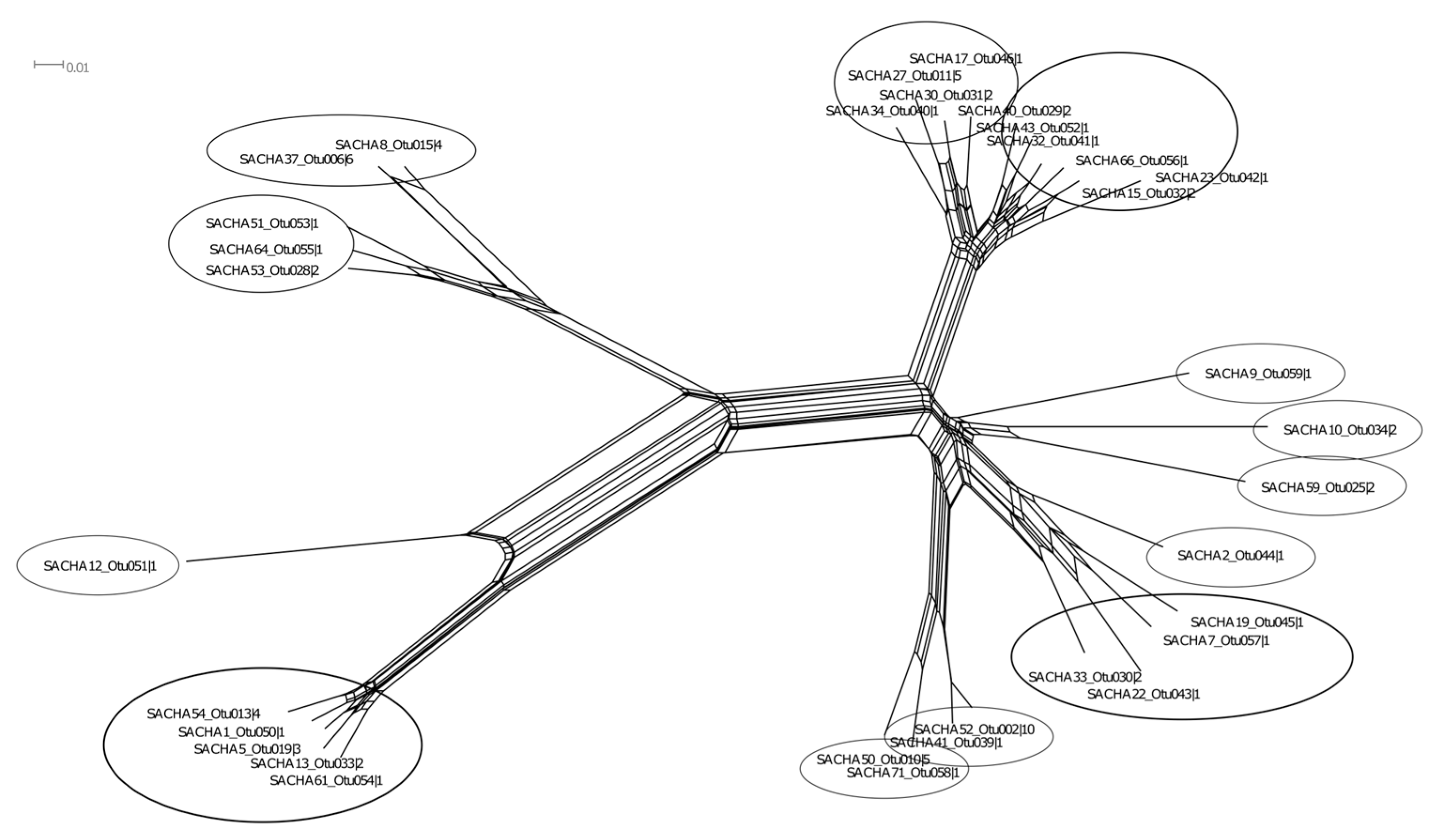
3.1. Characterization of AM Fungal Community
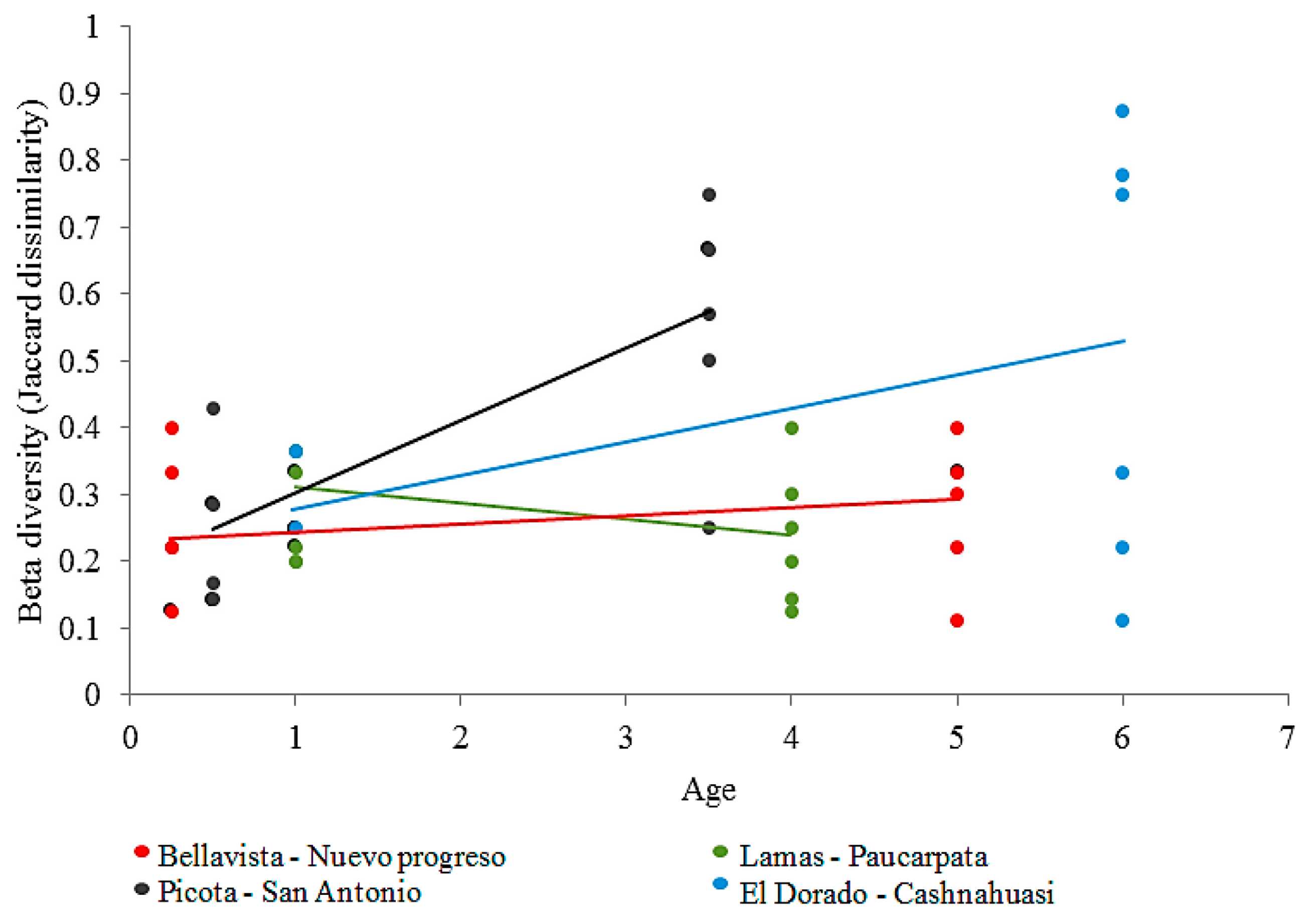
3.2. Effect of Age on Diversity of AM Fungal Communities
3.3. Drivers of Community Assembly (Database)
3.4. Drivers of Community Assembly (Peak Profile)
4. Discussion
4.1. Variability of Beta Diversity and Composition of AM Fungal Communities
4.2. Influence of Soil Physico-Chemical Variables on the Composition of AM Fungal Communities
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | Crop Age (Years) | Sample | Altitude | Latitude, Longitude |
|---|---|---|---|---|
| Lamas-Paucarpata | 1 | LAPAAP1 | 462 | S 06°26′26.44″, O 76°31′39.11″ |
| LAPAAP3 | 443 | S 06°26′36.90″, O 76°31′37.86″ | ||
| LAPAAP4 | 439 | S 06°26′37.50″, O 76°31′39.18″ | ||
| LAPAAP5 | 437 | S 06°26′36.84″, O 76°31′39.84″ | ||
| 4 | LAPABP1 | 502 | S 06°26′8.88″, O 76°31′51.84″ | |
| LAPABP2 | 503 | S 06°26′9.06″, O 76°31′52.20″ | ||
| LAPABP3 | 501 | S 06°26′8.58″, O 76°31′52.08″ | ||
| LAPABP4 | 502 | S 06°26′8.82″, O 76°31′52.14″ | ||
| Picota-San Antonio | 0.5 | DOCAAP1 | 250 | S 06°59′45.12″, O 76°12′32.58″ |
| DOCAAP2 | 241 | S 06°59′46.32″, O 76°12′33.36″ | ||
| DOCAAP3 | 234 | S 06°59′45.36″, O 76°12′30.72″ | ||
| DOCAAP4 | 234 | S 06°59′45.60″, O 76°12′29.22″ | ||
| 3.5 | DOCABP2 | 284 | S 06°59′39.12″, O 76°12′29.70″ | |
| DOCABP3 | 286 | S 06°59′38.22″, O 76°12′30.06″ | ||
| DOCABP4 | 283 | S 06°59′38.34″, O 76°12′28.80″ | ||
| DOCABP5 | 284 | S 06°59′38.04″, O 76°12′27.36″ | ||
| El Dorado-Cashnahuasi | 1 | PISAAP1 | 900 | S 06°26′51.60″, O 76°51′37.02″ |
| PISAAP2 | 897 | S 06°26′05.52″, O 76°51′39.72″ | ||
| PISAAP3 | 904 | S 06°26′05.64″, O 76°51′36.66″ | ||
| PISAAP5 | 914 | S 06°26′05.64″, O 76°51′36.12″ | ||
| 6 | PISABP1 | 891 | S 06°26′3.90″, O 76°51′37.62″ | |
| PISABP2 | 893 | S 06°26′4.14″, O 76°51′37.08″ | ||
| PISABP3 | 898 | S 06°26′4.08″, O 76°51′36.66″ | ||
| PISABP4 | 898 | S 06°26′4.32″, O 76°51′36.66″ | ||
| Bellavista-Nuevo Progreso | 0.25 | BEPOAP1 | 305 | S 07°10′01.88″, O 76°22′15.18″ |
| BEPOAP2 | 304 | S 07°10′01.91″, O 76°22′15.51″ | ||
| BEPOAP3 | 306 | S 07°10′01.87″, O 76°22′15.60″ | ||
| BEPOAP4 | 309 | S 07°10′02.27″, O 76°22′15.85″ | ||
| 5 | BEPOBP1 | 286 | S 07°10′05.29″, O 76°22′14.83″ | |
| BEPOBP3 | 285 | S 07°10′05.68″, O 76°22′15.55″ | ||
| BEPOBP4 | 282 | S 07°10′06.24″, O 76°22′14.67″ | ||
| BEPOBP5 | 286 | S 07°10′07.18″, O 76°22′14.51″ |
| Locality | Paucarpata-Lamas | Cashnahuasi-El Dorado | San Antonio-Picota | Nuevo Progreso-Bellavista | |
|---|---|---|---|---|---|
| pH | 7.7 | 7.05 | 7.85 | 7.8 | |
| Electrical conductivity (dS/m) | 0.4 | 0.3 | 0.45 | 0.45 | |
| CaCO3 (%) | 16.1 | 1.25 | 14.15 | 27.05 | |
| Organic matter (%) | 4.7 | 8.9 | 4.15 | 5 | |
| P (ppm) | 6.75 | 9.2 | 5.45 | 3.7 | |
| K (ppm) | 305.05 | 440.8 | 873.75 | 465.65 | |
| Texture | Sand (%) | 34 | 29.25 | 27.75 | 26.25 |
| Silt (%) | 31 | 33.75 | 33.75 | 32.75 | |
| Clay (%) | 35 | 37 | 38.5 | 41 | |
| CIC (meq/100 g) | 31.3 | 35.25 | 27.4 | 30.9 | |
| Exchangeable cations | Ca+2 | 27.6 | 26.6 | 20.95 | 25.2 |
| Mg+2 | 2.85 | 2.6 | 4.35 | 4.6 | |
| K+ | 0.75 | 1 | 1.95 | 0.95 | |
| Na+ | 0.1 | 0.1 | 0.15 | 0.15 | |
| Base Sum | 31.3 | 30.35 | 27.4 | 30.9 | |
| % Bases Saturation | 100 | 86.4 | 100 | 100 | |
| N (%) | 0.3 | 0.4 | 0.3 | 0.4 | |
| OTU | Scientific Name | Query Cover | Per. Ident. | Accession | Family |
|---|---|---|---|---|---|
| OTU1 | Uncultured Diversispora | 95% | 96.61% | LC416144.1 | Diversisporaceae |
| OTU2 | Uncultured Glomeromycotina | 94% | 97.64% | LR656581.1 | |
| Diversispora trimurales | 89% | 95.05% | FJ461854.1 | Diversisporaceae | |
| OTU3 | Glomeromycotina sp. | 100% | 94.68% | KY114635.1 | |
| Acaulospora kentinensis | 100% | 94.15% | FN547520.1 | Acaulosporaceae | |
| OTU4 | Uncultured Glomus | 100% | 95.67% | JQ218218.1 | Glomeraceae |
| OTU5 | Uncultured Glomus | 100% | 94.44% | KF849703.1 | Glomeraceae |
| OTU6 | Uncultured Glomeromycotina | 100% | 88.78% | MH469403.1 | |
| Oehlia diaphana | 100% | 88.83% | MG836662.1 | Glomeraceae | |
| OTU7 | Uncultured Claroideoglomus | 95% | 97.38% | LC416147.1 | Glomerales |
| OTU8 | Kamienskia sp. | 100% | 89.93% | MT765700.1 | Glomeraceae |
| OTU9 | Uncultured Glomerales | 91% | 97.35% | KC411100.1 | Glomeraceae |
| OTU10 | Rhizophagus irregularis | 100% | 96.81% | FM992381.1 | Glomeraceae |
| OTU11 | Uncultured Funneliformis | 95% | 97.18% | LC416202.1 | Glomeraceae |
| OTU12 | Uncultured Glomus | 100% | 95.11% | KJ701497.1 | Glomeraceae |
| OTU13 | Acaulospora scrobiculata | 100% | 98.91% | FR692351.1 | Acaulosporaceae |
References
- Hamaker, B.R.; Valles, C.; Gilman, R.; Hardmeier, R.M.; Clark, D.; García, H.H.; Gonzales, A.E.; Kohlstad, I.; Castro, M.; Valdivia, R.; et al. Amino acid and fatty acid profiles of the Inca peanut (Plukenetia volubilis). Cereal Chem. 1992, 69, 461–463. [Google Scholar]
- Gillespie, L.J. A synopsis of neotropical Plukenetia (Euphorbiaceae) including two new species. Syst. Bot. 1993, 18, 575–592. [Google Scholar] [CrossRef]
- Del-Castillo, A.M.R.; Gonzalez-Aspajo, G.; Sánchez-Márquez, M.F.; Kodahl, N. Ethnobotanical Knowledge in the Peruvian Amazon of the Neglected and Underutilized Crop Sacha Inchi (Plukenetia volubilis L.). Econ. Bot. 2019, 73, 281–287. [Google Scholar] [CrossRef]
- Kodahl, N. Sacha inchi (Plukenetia volubilis L.)—From lost crop of the Incas to part of the solution to global challenges? Planta 2020, 251, 80. [Google Scholar] [CrossRef]
- Chirinos, R.; Zuloeta, G.; Pedreschi, R.; Mignolet, E.; Larondelle, Y.; Campos, D. Sacha inchi (Plukenetia volubilis): A seed source of polyunsaturated fatty acids, tocopherols, phytosterols, phenolic compounds and antioxidant capacity. Food Chem. 2013, 141, 1732–1739. [Google Scholar] [CrossRef] [PubMed]
- Štěrbová, L.; Hlásná Čepková, P.; Viehmannová, I.; Huansi, D.C. Effect of Thermal Processing on Phenolic Content, Tocopherols and Antioxidant Activity of Sacha Inchi Kernels. J. Food Process. Preserv. 2017, 41, e12848. [Google Scholar] [CrossRef]
- Srichamnong, W.; Ting, P.; Pitchakarn, P.; Nuchuchua, O.; Temviriyanukul, P. Safety assessment of Plukenetia volubilis (Inca peanut) seeds, leaves, and their products. Food Sci. Nutr. 2018, 6, 962–969. [Google Scholar] [CrossRef]
- Wang, S.; Zhu, F.; Kakuda, Y. Sacha inchi (Plukenetia volubilis L.): Nutritional composition, biological activity, and uses. Food Chem. 2018, 265, 316–328. [Google Scholar] [CrossRef]
- Dostert, N.; Brokamp, G.; Cano, A.; La Torre, M.I.; Weigend, M. Factsheet: Datos botánicos de Sacha Inchi—Plukenetia volubilis L. Proyecto Perú Biodiverso—PBD. 2009. Available online: https://repositorio.promperu.gob.pe/bitstream/handle/20.500.14152/1337/Factsheet_datos_botanicos_sacha_inchi_2009_keyword_principal.pdf?sequence=1 (accessed on 1 December 2022).
- Kodahl, N.; Sørensen, M. Sacha Inchi (Plukenetia volubilis L.) Is an Underutilized Crop with a Great Potential. Agronomy 2021, 11, 1066. [Google Scholar] [CrossRef]
- Manco Céspedes, E.I. Sacha Inchi (Plukenetia volubilis L.) Cultivo Promisorio Para la Amazonía Peruana; Instituto Nacional de Innovación Agraria: Puno, Peru, 2008. [Google Scholar]
- Álvarez Gómez, L.; Ríos Torres, S. Estudio de Viabilidad Económica del Cultivo de Plukenetia volubilis L., Sacha Inchi, en el Departamento de San Martín; Instituto de Investigaciones de la Amazonía Peruana: Iquitos, Peru, 2009. [Google Scholar]
- Santín, C.; Doerr, S.H. Fire effects on soils: The human dimension. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150171. [Google Scholar] [CrossRef]
- Neary, D.G.; Klopatek, C.C.; DeBano, L.F.; Ffolliott, P.F. Fire effects on belowground sustainability: A review and synthesis. For. Ecol. Manag. 1999, 122, 51–71. [Google Scholar] [CrossRef]
- Gongalsky, K.B.; Persson, T. Recovery of soil macrofauna after wildfires in boreal forests. Soil Biol. Biochem. 2013, 57, 182–191. [Google Scholar] [CrossRef]
- Moody, J.A.; Shakesby, R.A.; Robichaud, P.R.; Cannon, S.H.; Martin, D.A. Current research issues related to post-wildfire runoff and erosion processes. Earth-Sci. Rev. 2013, 122, 10–37. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef]
- Frąc, M.; Hannula, S.E.; Bełka, M.; Jędryczka, M. Fungal Biodiversity and Their Role in Soil Health. Front. Microbiol. 2022, 9, 707. [Google Scholar] [CrossRef]
- Brundrett, M.C. Mycorrhizal associations and other means of nutrition of vascular plants: Understanding the global diversity of host plants by resolving conflicting information and developing reliable means of diagnosis. Plant Soil 2009, 320, 37–77. [Google Scholar]
- Bucher, M. Functional biology of plant phosphate uptake at root and mycorrhiza interfaces. New Phytol. 2007, 173, 11–26. [Google Scholar] [CrossRef]
- Bücking, H.; Kafle, A. Role of Arbuscular Mycorrhizal Fungi in the Nitrogen Uptake of Plants: Current Knowledge and Research Gaps. Agronomy 2015, 5, 587–612. [Google Scholar] [CrossRef]
- Saboor, A.; Ali, M.A.; Danish, S.; Ahmed, N.; Fahad, S.; Datta, R.; Ansari, M.J.; Nasif, O.; Rahman, M.H.U.; Glick, B.R. Effect of arbuscular mycorrhizal fungi on the physiological functioning of maize under zinc-deficient soils. Sci. Rep. 2021, 11, 18468. [Google Scholar] [CrossRef] [PubMed]
- Azcón-Aguilar, C.; Jaizme-Vega, M.C.; Calvet, C. The contribution of arbuscular mycorrhizal fungi to the control of soil-borne plant pathogens. In Mycorrhizal Technology in Agriculture: From Genes to Bioproducts; Gianinazzi, S., Schüepp, H., Barea, J.M., Haselwandter, K., Eds.; Birkhäuser: Basel, Switzerland, 2002; pp. 187–197. [Google Scholar]
- Ruiz-Lozano, J.M. Arbuscular mycorrhizal symbiosis and alleviation of osmotic stress. New perspectives for molecular studies. Mycorrhiza 2003, 13, 309–317. [Google Scholar] [CrossRef]
- Khade, S.W.; Adholeya, A. Feasible Bioremediation through Arbuscular Mycorrhizal Fungi Imparting Heavy Metal Tolerance: A Retrospective. Bioremediation J. 2007, 11, 33–43. [Google Scholar] [CrossRef]
- Berruti, A.; Lumini, E.; Balestrini, R.; Bianciotto, V. Arbuscular Mycorrhizal Fungi as Natural Biofertilizers: Let’s Benefit from Past Successes. Front. Microbiol. 2016, 6, 1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Lei, Y.; Zheng, Y.; Cai, Z. Synergistic effect of colonization with arbuscular mycorrhizal fungi improves growth and drought tolerance of Plukenetia volubilis seedlings. Acta Physiol. Plant. 2013, 35, 687–696. [Google Scholar] [CrossRef]
- Wiriya, J.; Rangjaroen, C.; Teaumroong, N.; Sungthong, R.; Lumyong, S. Rhizobacteria and Arbuscular Mycorrhizal Fungi of Oil Crops (Physic Nut and Sacha Inchi): A Cultivable-Based Assessment for Abundance, Diversity, and Plant Growth-Promoting Potentials. Plants 2020, 9, 1773. [Google Scholar] [CrossRef]
- Corazon-Guivin, M.A.; Mendoza, A.C.; Guerrero-Abad, J.C.; Vallejos-Tapullima, A.; Carballar-Hernández, S.; da Silva, G.A.; Oehl, F. Funneliglomus, gen. nov., and Funneliglomus sanmartinensis, a new arbuscular mycorrhizal fungus from the Amazonia region in Peru. Sydowia-Horn 2019, 71, 17–24. [Google Scholar]
- Corazon-Guivin, M.A.; Cerna-Mendoza, A.; Guerrero-Abad, J.C.; Vallejos-Tapullima, A.; Silva, A.; Oehl, F. Acaulospora aspera, a new fungal species in the Glomeromycetes from rhizosphere soils of the inka nut (Plukenetia volubilis L.) in Peru. J. Appl. Bot. Food Qual. 2019, 92, 250–257. [Google Scholar]
- Corazon Guivin, M.A.; Cerna-Mendoza, A.; Guerrero-Abad, J.C.; Vallejos-Tapullima, A.; Carballar-Hernández, S.; da Silva, G.A.; Oehl, F. Nanoglomus plukenetiae, a new fungus from Peru, and a key to small-spored Glomeraceae species, including three new genera in the “Dominikia complex/clades”. Mycol. Prog. 2019, 18, 1395–1409. [Google Scholar] [CrossRef]
- Corazon Guivin, M.A.; Cerna-Mendoza, A.; Guerrero-Abad, J.C.; Vallejos-Tapullima, A.; Carballar-Hernández, S.; da Silva, G.A.; Oehl, F. Microkamienskia gen. nov. and Microkamienskia peruviana, a new arbuscular mycorrhizal fungus from Western Amazonia. Nova Hedwig. 2019, 109, 355–368. [Google Scholar] [CrossRef]
- Corazon Guivin, M.A.; Cerna-Mendoza, A.; Guerrero-Abad, J.C.; Vallejos-Tapullima, A.; Carballar-Hernández, S.; da Silva, G.A.; Oehl, F. Acaulospora flava, a new arbuscular mycorrhizal fungus from Coffea arabica and Plukenetia volubilis plantations at the sources of the Amazon river in Peru. J. Appl. Bot. Food Qual. 2021, 94, 116–123. [Google Scholar]
- Brundrett, M.; Ashwath, N. Glomeromycotan mycorrhizal fungi from tropical Australia III. Measuring diversity in natural and disturbed habitats. Plant Soil 2013, 370, 419–443. [Google Scholar] [CrossRef]
- Jha, D.; Sharmah, D. Diversity of Arbuscular Mycorrhizal Fungi in Disturbed and Undisturbed Forests of Karbi Anglong Hills of Assam, India. Agric. Res. 2014, 3, 229–238. [Google Scholar]
- van der Heyde, M.; Ohsowski, B.; Abbott, L.K.; Hart, M.M. Arbuscular mycorrhizal fungus responses to disturbance are context-dependent. Mycorrhiza 2017, 27, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Ohsowski, B.; Zaitsoff, D.; Opik, M.; Hart, M.M. Where the wild things are: Looking for uncultured Glomeromycota. New Phytol. 2014, 204, 171–179. [Google Scholar] [CrossRef]
- Debnath, A. Arbuscular mycorrhiza fungal diversity in the open land adjacent to rubber plantation in Tripura, Northeast India. Mycorrhiza News 2014, 26, 4–9. [Google Scholar]
- García De León, D.; Davison, J.; Moora, M.; Öpik, M.; Feng, H.; Hiiesalu, I.; Jairus, T.; Koorem, K.; Liu, Y.; Phosri, C.; et al. Anthropogenic disturbance equalizes diversity levels in arbuscular mycorrhizal fungal communities. Global Chang. Biol. 2018, 24, 2649–2659. [Google Scholar] [CrossRef] [PubMed]
- Mori, A.S.; Isbell, F.; Seidl, R. β-Diversity, Community Assembly, and Ecosystem Functioning. Trends Ecol. Evol. 2018, 33, 549–564. [Google Scholar] [CrossRef]
- Castro, A.; Davila, C.; Laura, W.; Cubas, F.; Avalos, G.; López-Ocaña, C.; Villena, D.; Valdez, M.; Urbiola, J.; Trebejo, I.; et al. Climas del Perú—Mapa de Clasificación Climática Nacional; Servicio Nacional de Meteorología e Hidrología del Perú–SENAMHI: Lima, Perú, 2021. [Google Scholar]
- Olsen, S.R. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; US Department of Agriculture: Washington, DC, USA, 1954.
- Walkley, A.; Black, I.A. An examination of the Degtjardreff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Bouyoucos, G.J. Directions for Making Mechanical Analyses of Soils by the Hydrometer Method. Soil Sci. 1936, 42, 225–230. [Google Scholar] [CrossRef]
- Thomas, G.W. Exchangeable Cations. Methods of Soil Analysis, Part 2, Chemical and Microbiological Properties, 2nd ed.; Page, A.L., Ed.; Agronomy American Society of Agronomy, Soil Science Society of America: Madison, WL, USA, 1982; pp. 159–165. [Google Scholar]
- Kjeldahl, J. Neue Methode zur Bestimmung des Stickstoffs in organischen Körpern. Z. Für Anal. Chem. 1883, 22, 366–382. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. (Eds.) A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Dickie, I.; FitzJohn, R. Using terminal restriction fragment length polymorphism (T-RFLP) to identify mycorrhizal fungi: A methods review. Mycorrhiza 2007, 17, 259–270. [Google Scholar] [CrossRef] [PubMed]
- López-García, A. Analysis of Arbuscular Mycorrhizal Fungal Communities by Terminal Restriction Fragment Length Polymorphism (TRFLP). Methods Mol. Biol. 2020, 2146, 117–136. [Google Scholar] [PubMed]
- Krüger, M.; Stockinger, H.; Krüger, C.; Schüßler, A. DNA-based species level detection of Glomeromycota: One PCR primer set for all arbuscular mycorrhizal fungi. New Phytol. 2009, 183, 212–223. [Google Scholar] [CrossRef]
- Gollotte, A.; van Tuinen, D.; Atkinson, D. Diversity of arbuscular mycorrhizal fungi colonising roots of the grass species Agrostis capillaris and Lolium perenne in a field experiment. Mycorrhiza 2004, 14, 111–117. [Google Scholar] [CrossRef] [PubMed]
- van Tuinen, D.; Jacquot, E.; Zhao, B.; Gollotte, A.; Gianinazzi-Pearson, V. Characterization of root colonization profiles by a microcosm community of arbuscular mycorrhizal fungi using 25S rDNA-targeted nested PCR. Mol. Ecol. 1998, 7, 879–887. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Lekberg, Y.; Gibbons, S.M.; Rosendahl, S. Will different OTU delineation methods change interpretation of arbuscular mycorrhizal fungal community patterns? New Phytol. 2014, 202, 1101–1104. [Google Scholar] [CrossRef]
- Schloss, P.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Huson, D.H.; Bryant, D. Application of Phylogenetic Networks in Evolutionary Studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) Software Version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Collins, R.; Rocap, G. REPK: An analytical web server to select restriction endonucleases for terminal restriction fragment length polymorphism analysis. Nucleic Acids Res. 2007, 35, W58–W62. [Google Scholar] [CrossRef] [PubMed]
- Junier, P.; Junier, T.; Witzel, K.-P. TRiFLe, a Program for In Silico Terminal Restriction Fragment Length Polymorphism Analysis with User-Defined Sequence Sets. Appl. Environ. Microbiol. 2008, 74, 6452–6456. [Google Scholar] [CrossRef]
- Fitzjohn, R.G.; Dickie, I.A. TRAMPR: An R package for analysis and matching of terminal-restriction fragment length polymorphism (TRFLP) profiles. Mol. Ecol. Notes 2007, 7, 583–587. [Google Scholar] [CrossRef]
- López-García, A.; Azcón-Aguilar, C.; Barea, J.M. The interactions between plant life form and fungal traits of arbuscular mycorrhizal fungi determine the symbiotic community. Oecologia 2014, 176, 1075–1086. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Meth. Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinforma. Oxf. Engl. 2010, 26, 1463–1464. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 2003. [Google Scholar]
- Kil, Y.-J.; Eo, J.-K.; Eom, A.H. Root Age-Dependent Changes in Arbuscular Mycorrhizal Fungal Communities Colonizing Roots of Panax ginseng. Mycobiology 2014, 42, 416–421. [Google Scholar] [CrossRef]
- Chakraborty, K.; Sinha, S.; Debnath, A.; Roy Das, A.; Saha, A.; Das, P. Arbuscular mycorrhizal fungal colonization in three different age groups of rubber plantations in Tripura, North-East India. Plant Pathol. Quar Antine 2016, 6, 122–131. [Google Scholar] [CrossRef]
- Chagnon, P.-L.; Bradley, R.; Maherali, H.; Klironomos, J.N. A trait-based framework to understand life history of mycorrhizal fungi. Trends Plant Sci. 2013, 18, 484–491. [Google Scholar] [CrossRef]
- Fichtner, A.; Oheimb, G.; Haerdtle, W.; Wilken, C.; Gutknecht, J. Effects of anthropogenic disturbances on soil microbial communities in oak forests persist for more than 100 years. Soil Biol. Biochem. 2014, 70, 79–87. [Google Scholar] [CrossRef]
- Johnson, N.C.; Zak, D.R.; Tilman, D.; Pfleger, F.L. Dynamics of vesicular-arbuscular mycorrhizae during old field succession. Oecologia 1991, 86, 349–358. [Google Scholar] [CrossRef]
- Jansa, J.; Mozafar, A.; Kuhn, G.; Anken, T.; Ruh, R.; Sanders, I.R.; Frossard, E.J.E.A. Soil Tillage Affects the Community Structure of Mycorrhizal Fungi in Maize Roots. Ecol. Appl. 2003, 13, 1164–1176. [Google Scholar] [CrossRef]
- Evans, D.G.; Miller, M.H. The role of the external mycelial network in the effect of soil disturbance upon vesicular—Arbuscular mycorrhizal colonization of maize. New Phytol. 1990, 114, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Treseder, K.K. A meta-analysis of mycorrhizal responses to nitrogen, phosphorus, and atmospheric CO2 in field studies. New Phytol. 2004, 164, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Ling, N.; Guo, J.; Wang, M.; Guo, S.; Shen, Q. Impacts of Fertilization Regimes on Arbuscular Mycorrhizal Fungal (AMF) Community Composition Were Correlated with Organic Matter Composition in Maize Rhizosphere Soil. Front. Microbiol. 2022, 7, 1840. [Google Scholar] [CrossRef] [PubMed]
- Labidi, S.; Ben Jeddi, F.; Tisserant, B.; Debiane, D.; Rezgui, S.; Grandmougin-Ferjani, A.; Sahraoui, A.L.-H. Role of arbuscular mycorrhizal symbiosis in root mineral uptake under CaCO3 stress. Mycorrhiza 2011, 22, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Altland, J.; Jeong, K. Dolomitic Lime Amendment Affects Pine Bark Substrate pH, Nutrient Availability, and Plant Growth: A Review. HortTechnology 2016, 26, 565–573. [Google Scholar] [CrossRef]
- Schreiber, M.J.; Nunez, G.H. Calcium Carbonate Can Be Used to Manage Soilless Substrate pH for Blueberry Production. Horticulturae 2021, 7, 74. [Google Scholar] [CrossRef]
- Coughlan, A.P.; Dalpé, Y.; Lapointe, L.; Piché, Y. Soil pH-induced changes in root colonization, diversity, and reproduction of symbiotic arbuscular mycorrhizal fungi from healthy and declining maple forests. Can. J. For. Res. 2000, 30, 1543–1554. [Google Scholar] [CrossRef]
- Aarle, I.; Olsson, P.; Soderstrom, B. Arbuscular mycorrhizal fungi respond to the substrate pH of their extraradical mycelium by altered growth and root colonization. New Phytol. 2002, 155, 173–182. [Google Scholar] [CrossRef]
- Diop, I.; Ndoye, F.; Diédhiou, A.G.; Krasova-Wade, T.; Dorego, F.; Noba, K.; Ambrosi, J.P.; Kane, A. Diversity and spore density of arbuscular mycorrhizal fungi in the rhizosphere of Cowpea (Vigna unguiculate (L.) Walp.) cultivated in different soils in Senegal. J. Anim. Plant Sci. 2021, 48, 8552–8565. [Google Scholar]
- Parniske, M. Arbuscular mycorrhiza: The mother of plant root endosymbiosis. Nat. Rev. Microbiol. 2008, 6, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wei, S.; Hu, W.; Xiao, L.; Tang, M. Arbuscular Mycorrhizal Fungus Rhizophagus irregularis Increased Potassium Content and Expression of Genes Encoding Potassium Channels in Lycium barbarum. Front. Plant Sci. 2017, 8, 440. [Google Scholar] [CrossRef] [PubMed]
- Giri, B.; Mukerji, K. Mycorrhizal inoculant alleviates salt stress in Sesbania aegyptiaca and Sesbania grandiflora under field conditions: Evidence for reduced sodium and improved magnesium uptake. Mycorrhiza 2004, 14, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Klinsukon, C.; Lumyong, S.; Kuyper, T.; Boonlue, S. Colonization by arbuscular mycorrhizal fungi improves salinity tolerance of eucalyptus (Eucalyptus camaldulensis) seedlings. Sci. Rep. 2021, 11, 4362. [Google Scholar] [CrossRef]
- St Clair, S.B.; Lynch, J.P. Base cation stimulation of mycorrhization and photosynthesis of sugar maple on acid soils are coupled by foliar nutrient dynamics. New Phytol. 2005, 165, 581–590. [Google Scholar] [CrossRef]
- Lin, X.; Feng, Y.; Zhang, H.; Chen, R.; Wang, J.; Zhang, J.; Chu, H. Long-term balanced fertilization decreases arbuscular mycorrhizal fungal diversity in an arable soil in North China revealed by 454 pyrosequencing. Environ. Sci. Technol. 2012, 46, 5764–5771. [Google Scholar] [CrossRef]
- Camenzind, T.; Hempel, S.; Homeier, J.; Horn, S.; Velescu, A.; Wilcke, W.; Rillig, M.C. Nitrogen and phosphorus additions impact arbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest. Glob. Chang. Biol. 2014, 20, 3646–3659. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, Y.; Jiang, S.; Deng, Y.; Christie, P.; Murray, P.J.; Li, X.; Zhang, J. Arbuscular mycorrhizal fungi in soil and roots respond differently to phosphorus inputs in an intensively managed calcareous agricultural soil. Sci. Rep. 2016, 6, 24902. [Google Scholar] [CrossRef] [Green Version]
- Kiers, E.T.; Duhamel, M.; Beesetty, Y.; Mensah, J.A.; Franken, O.; Verbruggen, E.; Fellbaum, C.R.; Kowalchuk, G.A.; Hart, M.M.; Bago, A.; et al. Reciprocal Rewards Stabilize Cooperation in the Mycorrhizal Symbiosis. Science 2011, 333, 880–882. [Google Scholar] [CrossRef]
- Yu, L.; Nicolaisen, M.; Larsen, J.; Ravnskov, S. Organic fertilization alters the community composition of root associated fungi in Pisum sativum. Soil Biol. Biochem. 2013, 58, 36–41. [Google Scholar] [CrossRef]
- Ai, C.; Liang, G.; Sun, J.; Wang, X.; He, P.; Zhou, W. Reduced dependence of rhizosphere microbiome on plant-derived carbon in 32-year long-term inorganic and organic fertilized soils. Soil Biol. Biochem. 2015, 80, 70–78. [Google Scholar] [CrossRef]




| Fixed Factor | Random Factor | ||
|---|---|---|---|
| Age | Site | ||
| Database | OTU richness | 0.7911,28 | 2.7401 |
| Beta diversity | 7.5911,45 ** | 2.0011 | |
| SES-MPD | 0.0581,28 | 1.3721 | |
| Peak profile | OTU richness | 4.0531,27 | 0.0001 |
| Beta diversity | 2.7031,44 | 15.4451 *** | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de la Sota Ricaldi, A.M.; Rengifo del Águila, S.; Blas Sevillano, R.; López-García, Á.; Corazon-Guivin, M.A. Beta Diversity of Arbuscular Mycorrhizal Communities Increases in Time after Crop Establishment of Peruvian Sacha Inchi (Plukenetia volubilis). J. Fungi 2023, 9, 194. https://doi.org/10.3390/jof9020194
de la Sota Ricaldi AM, Rengifo del Águila S, Blas Sevillano R, López-García Á, Corazon-Guivin MA. Beta Diversity of Arbuscular Mycorrhizal Communities Increases in Time after Crop Establishment of Peruvian Sacha Inchi (Plukenetia volubilis). Journal of Fungi. 2023; 9(2):194. https://doi.org/10.3390/jof9020194
Chicago/Turabian Stylede la Sota Ricaldi, Ana Maria, Sofía Rengifo del Águila, Raúl Blas Sevillano, Álvaro López-García, and Mike Anderson Corazon-Guivin. 2023. "Beta Diversity of Arbuscular Mycorrhizal Communities Increases in Time after Crop Establishment of Peruvian Sacha Inchi (Plukenetia volubilis)" Journal of Fungi 9, no. 2: 194. https://doi.org/10.3390/jof9020194