
Functional Expression of Recombinant Candida auris Proteins in Saccharomyces cerevisiae Enables Azole Susceptibility Evaluation and Drug Discovery



Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Cell Culture Reagents
2.2. Chemicals and Drugs
2.3. Construction of Recombinant Strains
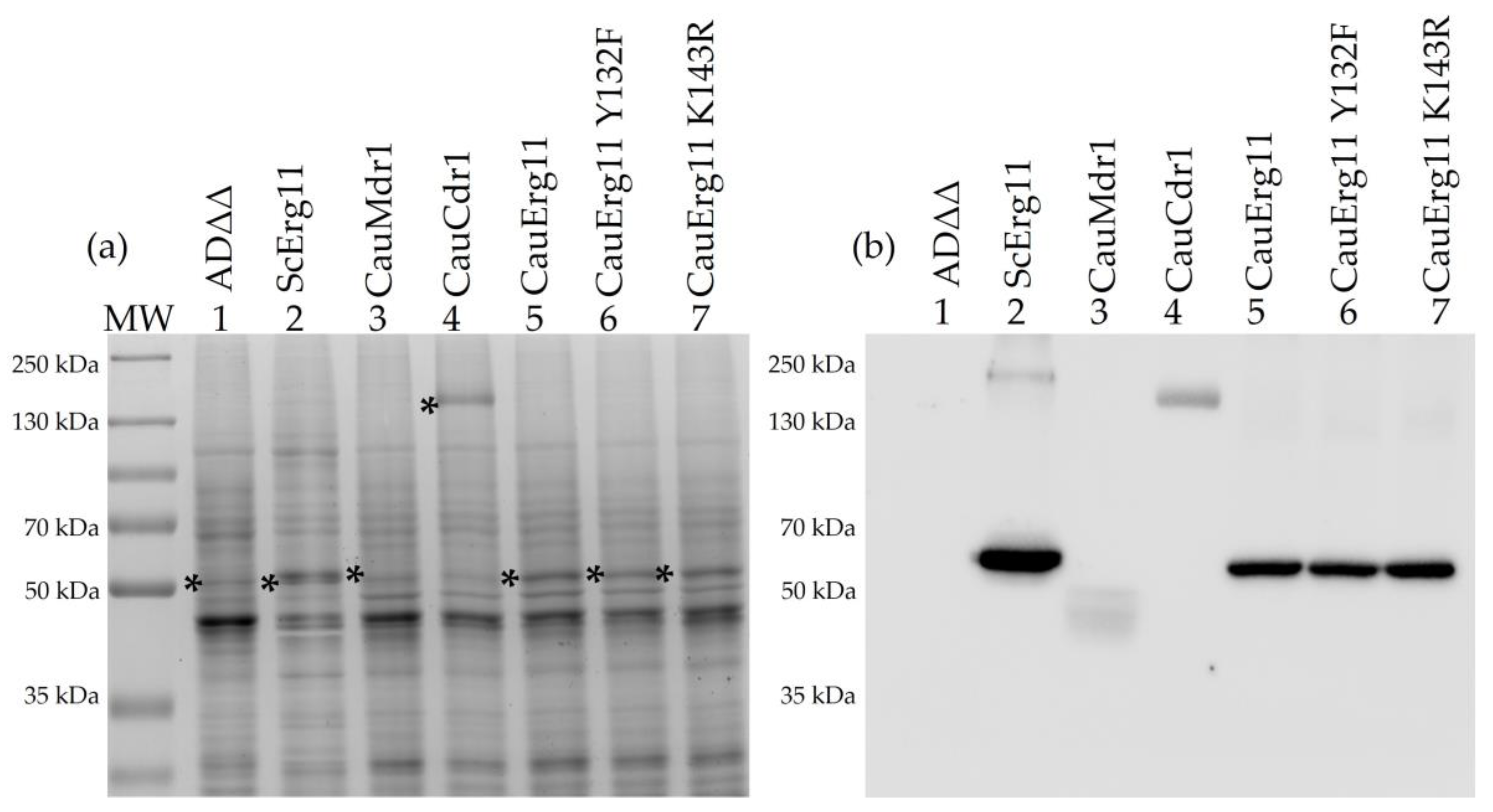
2.4. Crude Membrane Preparation and Western Blot Analysis
2.5. Partial Purification of Hexahistidine-Tagged Proteins from Recombinant Strains
2.6. Antifungal Susceptibility Testing
2.7. Nile Red Efflux Assay
2.8. ATPase Activity of the Recombinant Strain Overexpressing CauCDR1-6×His
2.9. Type II Azole Binding
2.10. Statistical Analysis
3. Results
3.1. Biochemical Characterization of Recombinant Strains
3.2. Mass Spectrometry of Tryptic Fragments Confirms Identity of Heterologously Expressed Proteins
3.3. Susceptibility Profiles of Recombinant Strains Confirm Phenotypes
3.4. Nile Red Confirms Efflux Pump Activity
3.5. ATPase Activity
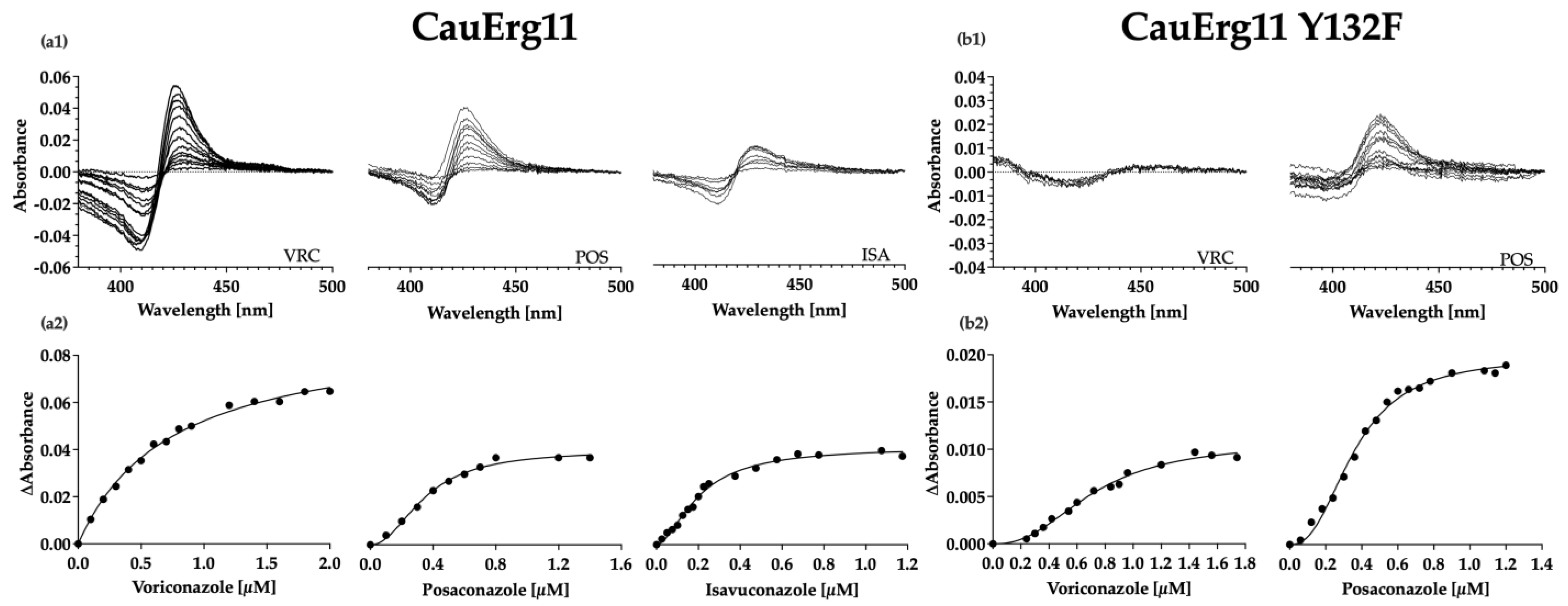
3.6. Type II Azole-Binding Studies
4. Discussion
4.1. Functional Expression of the C. Auris Azole Target and Drug Efflux Pumps in S. cerevisiae
4.2. Azole Susceptibility Wild-Type CauErg11 and the Y132F and K143R Mutants
4.3. Susceptibility Efflux Pumps Mdr1 and Cdr1
4.4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guinea, J. Global trends in the distribution of Candida species causing candidemia. Clin. Microbiol. Infect. 2014, 20 (Suppl. S6), 5–10. [Google Scholar] [CrossRef] [Green Version]
- Cleveland, A.; Harrison, L.H.; Farley, M.M.; Hollick, R.; Stein, B.; Chiller, T.M.; Lockhart, S.R.; Park, B.J. Declining Incidence of Candidemia and the Shifting Epidemiology of Candida Resistance in Two US Metropolitan Areas, 2008–2013: Results from Population-Based Surveillance. PLoS ONE 2015, 10, e0120452. [Google Scholar] [CrossRef] [Green Version]
- Hesstvedt, L.; Gaustad, P.; Andersen, C.T.; Haarr, E.; Hannula, R.; Haukland, H.H.; Hermansen, N.-O.; Larssen, K.W.; Mylvaganam, H.; Ranheim, T.E.; et al. Twenty-two years of candidaemia surveillance: Results from a Norwegian national study. Clin. Microbiol. Infect. 2015, 21, 938–945. [Google Scholar] [CrossRef] [Green Version]
- Schelenz, S.; Hagen, F.; Rhodes, J.L.; Abdolrasouli, A.; Chowdhary, A.; Hall, A.; Ryan, L.; Shackleton, J.; Trimlett, R.; Meis, J.F.; et al. First hospital outbreak of the globally emerging Candida auris in a European hospital. Antimicrob. Resist. Infect. Control. 2016, 5, 35. [Google Scholar] [CrossRef] [Green Version]
- WHO. WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action; World Health Organization: Geneva, Switzerland, 2022.
- Satoh, K.; Makimura, K.; Hasumi, Y.; Nishiyama, Y.; Uchida, K.; Yamaguchi, H. Candida auris sp. nov., a novel ascomycetous yeast isolated from the external ear canal of an inpatient in a Japanese hospital. Microbiol. Immunol. 2009, 53, 41–44. [Google Scholar] [CrossRef]
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.L.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A.; et al. Simultaneous Emergence of Multidrug-Resistant Candida auris on 3 Continents Confirmed by Whole-Genome Sequencing and Epidemiological Analyses. Clin. Infect. Dis. 2017, 64, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Mariita, R.M.; Davis, J.H.; Lottridge, M.M.; Randive, R.V. Shining light on multi-drug resistant Candida auris: Ultraviolet-C disinfection, wavelength sensitivity, and prevention of biofilm formation of an emerging yeast pathogen. Microbiologyopen 2022, 11, e1261. [Google Scholar] [CrossRef]
- Sekizuka, T.; Iguchi, S.; Umeyama, T.; Inamine, Y.; Makimura, K.; Kuroda, M.; Miyazaki, Y.; Kikuchi, K. Clade II Candida auris possess genomic structural variations related to an ancestral strain. PLoS ONE 2019, 14, e0223433. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, J.F.; Gade, L.; Chow, N.A.; Loparev, V.N.; Juieng, P.; Berkow, E.L.; Farrer, R.A.; Litvintseva, A.P.; Cuomo, C.A. Genomic insights into multidrug-resistance, mating and virulence in Candida auris and related emerging species. Nat. Commun. 2018, 9, 5346. [Google Scholar] [CrossRef] [Green Version]
- Chow, N.A.; de Groot, T.; Badali, H.; Abastabar, M.; Chiller, T.M.; Meis, J.F. Potential Fifth Clade of Candida auris, Iran, 2018. Emerg. Infect. Dis. 2019, 25, 1780–1781. [Google Scholar] [CrossRef] [Green Version]
- Spruijtenburg, B.; Badali, H.; Abastabar, M.; Mirhendi, H.; Khodavaisy, S.; Sharifisooraki, J.; Armaki, M.T.; de Groot, T.; Meis, J.F. Confirmation of fifth Candida auris clade by whole genome sequencing. Emerg. Microbes Infect. 2022, 11, 2405–2411. [Google Scholar] [CrossRef]
- Forgács, L.; Borman, A.M.; Prépost, E.; Tóth, Z.; Kardos, G.; Kovács, R.; Szekely, A.; Nagy, F.; Kovacs, I.; Majoros, L. Comparison of in vivo pathogenicity of four Candida auris clades in a neutropenic bloodstream infection murine model. Emerg. Microbes Infect. 2020, 9, 1160–1169. [Google Scholar] [CrossRef]
- CDC. Candida auris . Available online: https://www.cdc.gov/fungal/candida-auris/index.html (accessed on 27 November 2022).
- Chow, N.A.; Muñoz, J.F.; Gade, L.; Berkow, E.L.; Li, X.; Welsh, R.M.; Forsberg, K.; Lockhart, S.R.; Adam, R.; Alanio, A.; et al. Tracing the Evolutionary History and Global Expansion of Candida auris Using Population Genomic Analyses. Mbio 2020, 11, e03364-19. [Google Scholar] [CrossRef]
- Healey, K.R.; Kordalewska, M.; Ortigosa, C.J.; Singh, A.; Berrío, I.; Chowdhary, A.; Perlin, D.S. Limited ERG11 Mutations Identified in Isolates of Candida auris Directly Contribute to Reduced Azole Susceptibility. Antimicrob. Agents Chemother. 2018, 62, e01427-18. [Google Scholar] [CrossRef] [Green Version]
- Chowdhary, A.; Prakash, A.; Sharma, C.; Kordalewska, M.; Kumar, A.; Sarma, S.; Tarai, B.; Singh, A.; Upadhyaya, G.; Upadhyay, S.; et al. A multicentre study of antifungal susceptibility patterns among 350 Candida auris isolates (2009–17) in India: Role of the ERG11 and FKS1 genes in azole and echinocandin resistance. J. Antimicrob. Chemother. 2018, 73, 891–899. [Google Scholar] [CrossRef]
- Williamson, B.; Wilk, A.; Guerrero, K.D.; Mikulski, T.D.; Elias, T.N.; Sawh, I.; Cancino-Prado, G.; Gardam, D.; Heath, C.H.; Govender, N.P.; et al. Impact of Erg11 Amino Acid Substitutions Identified in Candida auris Clade III Isolates on Triazole Drug Susceptibility. Antimicrob. Agents Chemother. 2022, 66, e01624-21. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Holowka, T.; Orner, E.P.; Fries, B.C. Gene Duplication Associated with Increased Fluconazole Tolerance in Candida auris cells of Advanced Generational Age. Sci. Rep. 2019, 9, 5052. [Google Scholar] [CrossRef] [Green Version]
- Rybak, J.M.; Doorley, L.A.; Nishimoto, A.T.; Barker, K.S.; Palmer, G.E.; Rogers, P.D. Abrogation of Triazole Resistance upon Deletion of CDR1 in a Clinical Isolate of Candida auris. Antimicrob. Agents Chemother. 2019, 63, e00057-19. [Google Scholar] [CrossRef] [Green Version]
- Ben-Ami, R.; Berman, J.; Novikov, A.; Bash, E.; Shachor-Meyouhas, Y.; Zakin, S.; Maor, Y.; Tarabia, J.; Schechner, V.; Adler, A.; et al. Multidrug-Resistant Candida haemulonii and C. auris, Tel Aviv, Israel. Emerg. Infect. Dis. 2017, 23, 195–203. [Google Scholar] [CrossRef]
- Rybak, J.M.; Muñoz, J.F.; Barker, K.S.; Parker, J.E.; Esquivel, B.D.; Berkow, E.L.; Lockhart, S.R.; Gade, L.; Palmer, G.E.; White, T.C.; et al. Mutations in TAC1B: A Novel Genetic Determinant of Clinical Fluconazole Resistance in Candida auris. Mbio 2020, 11, e00365-20. [Google Scholar] [CrossRef]
- Li, J.; Coste, A.T.; Liechti, M.; Bachmann, D.; Sanglard, D.; Lamoth, F. Novel ERG11 and TAC1b Mutations Associated with Azole Resistance in Candida auris. Antimicrob. Agents Chemother. 2021, 65, e02663-20. [Google Scholar] [CrossRef]
- Li, J.; Coste, A.T.; Bachmann, D.; Sanglard, D.; Lamoth, F. Deciphering the Mrr1/Mdr1 Pathway in Azole Resistance of Candida auris. Antimicrob. Agents Chemother. 2022, 66, e00067-22. [Google Scholar] [CrossRef]
- Kordalewska, M.; Lee, A.; Park, S.; Berrio, I.; Chowdhary, A.; Zhao, Y.; Perlin, D.S. Understanding Echinocandin Resistance in the Emerging Pathogen Candida auris. Antimicrob. Agents Chemother. 2018, 62, e00238-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berkow, E.L.; Lockhart, S.R. Activity of CD101, a long-acting echinocandin, against clinical isolates of Candida auris. Diagn. Microbiol. Infect. Dis. 2018, 90, 196–197. [Google Scholar] [CrossRef]
- Rhodes, J.; Abdolrasouli, A.; Farrer, R.A.; Cuomo, C.A.; Aanensen, D.M.; Armstrong-James, D.; Fisher, M.C.; Schelenz, S. Genomic epidemiology of the UK outbreak of the emerging human fungal pathogen Candida auris. Emerg. Microbes Infect. 2018, 7, 43. [Google Scholar] [CrossRef] [Green Version]
- Sharma, D.; Paul, R.A.; Rudramurthy, S.M.; Kashyap, N.; Bhattacharya, S.; Soman, R.; Shankarnarayan, S.A.; Chavan, D.; Singh, S.; Das, P.; et al. Impact of FKS1 Genotype on Echinocandin In Vitro Susceptibility in Candida auris and In Vivo Response in a Murine Model of Infection. Antimicrob. Agents Chemother. 2022, 66, Barantsevich NEe01652-21. [Google Scholar] [CrossRef]
- Escandón, P.; Chow, N.A.; Caceres, D.H.; Gade, L.; Berkow, E.L.; Armstrong, P.; Rivera, S.; Misas, E.; Duarte, C.; Moulton-Meissner, H.; et al. Molecular Epidemiology of Candida auris in Colombia Reveals a Highly Related, Countrywide Colonization with Regional Patterns in Amphotericin B Resistance. Clin. Infect. Dis. 2019, 68, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Rybak, J.M.; Barker, K.S.; Muñoz, J.F.; Parker, J.E.; Ahmad, S.; Mokaddas, E.; Abdullah, A.; Elhagracy, R.S.; Kelly, S.L.; Cuomo, C.A.; et al. In vivo emergence of high-level resistance during treatment reveals the first identified mechanism of amphotericin B resistance in Candida auris. Clin. Microbiol. Infect. 2022, 28, 838–843. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, S.E.; Jacobs, J.L.; Dennis, E.K.; Taimur, S.; Rana, M.; Patel, D.; Gitman, M.; Patel, G.; Schaefer, S.; Iyer, K.; et al. Candida auris Pan-Drug-Resistant to Four Classes of Antifungal Agents. Antimicrob. Agents Chemother. 2022, 66, e00053-22. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tian, S.; Han, X.; Chu, Y.; Wang, Q.; Zhou, B.; Shang, H. Is the superbug fungus really so scary? A systematic review and meta-analysis of global epidemiology and mortality of Candida auris. BMC Infect. Dis. 2020, 20, 827. [Google Scholar] [CrossRef] [PubMed]
- Barantsevich, N.E.; Vetokhina, A.V.; Ayushinova, N.I.; Orlova, O.E.; Barantsevich, E.P. Candida auris Bloodstream Infections in Russia. Antibiotics 2020, 9, 557. [Google Scholar] [CrossRef]
- Al-Rashdi, A.; Al-Maani, A.; Al-Wahaibi, A.; Alqayoudhi, A.; Al-Jardani, A.; Al-Abri, S. Characteristics, Risk Factors, and Survival Analysis of Candida auris Cases: Results of One-Year National Surveillance Data from Oman. J. Fungi 2021, 7, 31. [Google Scholar] [CrossRef]
- Monk, B.C.; Tomasiak, T.M.; Keniya, M.V.; Huschmann, F.U.; Tyndall, J.D.A.; O’Connell, J.D., III; Cannon, R.D.; McDonald, J.G.; Rodriguez, A.; Finer-Moore, J.S.; et al. Architecture of a single membrane spanning cytochrome P450 suggests constraints that orient the catalytic domain relative to a bilayer. Proc. Natl. Acad. Sci. USA 2014, 111, 3865–3870. [Google Scholar] [CrossRef] [Green Version]
- Ruma, Y.N.; Keniya, M.V.; Tyndall, J.D.A.; Monk, B.C. Characterisation of Candida parapsilosis CYP51 as a Drug Target Using Saccharomyces cerevisiae as Host. J. Fungi 2022, 8, 69. [Google Scholar] [CrossRef] [PubMed]
- Lamping, E.; Monk, B.C.; Niimi, K.; Holmes, A.R.; Tsao, S.; Tanabe, K.; Niimi, M.; Uehara, Y.; Cannon, R.D. Characterization of Three Classes of Membrane Proteins Involved in Fungal Azole Resistance by Functional Hyperexpression in Saccharomyces cerevisiae. Eukaryot. Cell 2007, 6, 1150–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keniya, M.V.; Ruma, Y.N.; Tyndall, J.D.A.; Monk, B.C. Heterologous Expression of Full-Length Lanosterol 14α-Demethylases of Prominent Fungal Pathogens Candida albicans and Candida glabrata Provides Tools for Antifungal Discovery. Antimicrob. Agents Chemother. 2018, 62, e01131-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keniya, M.V.; Sabherwal, M.; Wilson, R.K.; Woods, M.A.; Sagatova, A.A.; Tyndall, J.D.A.; Monk, B.C. Crystal Structures of Full-Length Lanosterol 14α-Demethylases of Prominent Fungal Pathogens Candida albicans and Candida glabrata Provide Tools for Antifungal Discovery. Antimicrob. Agents Chemother. 2018, 62, e01134-18. [Google Scholar] [CrossRef] [Green Version]
- Langer, S.J.; Ghafoori, A.P.; Byrd, M.; Leinwand, L. A genetic screen identifies novel non-compatible loxP sites. Nucleic Acids Res. 2002, 30, 3067–3077. [Google Scholar] [CrossRef]
- Nagy, A. Cre recombinase: The universal reagent for genome tailoring. Genesis 2000, 26, 99–109. [Google Scholar] [CrossRef]
- Güldener, U.; Heck, S.; Fiedler, T.; Beinhauer, J.; Hegemann, J.H. A new efficient gene disruption cassette for repeated use in budding yeast. Nucleic Acids Res. 1996, 24, 2519–2524. [Google Scholar] [CrossRef] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Ivnitski-Steele, I.; Holmes, A.R.; Lamping, E.; Monk, B.C.; Cannon, R.D.; Sklar, L.A. Identification of Nile red as a fluorescent substrate of the Candida albicans ATP-binding cassette transporters Cdr1p and Cdr2p and the major facilitator superfamily transporter Mdr1p. Anal. Biochem. 2009, 394, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Keniya, M.V.; Fleischer, E.; Klinger, A.; Cannon, R.; Monk, B.C. Inhibitors of the Candida albicans Major Facilitator Superfamily Transporter Mdr1p Responsible for Fluconazole Resistance. PLoS ONE 2015, 10, e0126350. [Google Scholar] [CrossRef] [PubMed]
- Monk, B.C.; Kurtz, M.B.; Marrinan, J.A.; Perlin, D.S. Cloning and characterization of the plasma membrane H(+)-ATPase from Candida albicans. J. Bacteriol. 1991, 173, 6826–6836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madani, G.; Lamping, E.; Cannon, R.D. Engineering a Cysteine-Deficient Functional Candida albicans Cdr1 Molecule Reveals a Conserved Region at the Cytosolic Apex of ABCG Transporters Important for Correct Folding and Trafficking of Cdr1. Msphere 2021, 6, e01318-20. [Google Scholar] [CrossRef] [PubMed]
- Madani, G.; Lamping, E.; Lee, H.J.; Niimi, M.; Mitra, A.K.; Cannon, R.D. Small-Scale Plasma Membrane Preparation for the Analysis of Candida albicans Cdr1-mGFPHis. J. Vis. Exp. 2021, 172, e62592. [Google Scholar] [CrossRef]
- Niimi, K.; Harding, D.R.K.; Parshot, R.; King, A.; Lun, D.J.; Decottignies, A.; Niimi, M.; Lin, S.; Cannon, R.D.; Goffeau, A.; et al. Chemosensitization of Fluconazole Resistance in Saccharomyces cerevisiae and Pathogenic Fungi by a d-Octapeptide Derivative. Antimicrob. Agents Chemother. 2004, 48, 1256–1271. [Google Scholar] [CrossRef] [Green Version]
- Guengerich, F.P.; Martin, M.V.; Sohl, C.D.; Cheng, Q. Measurement of cytochrome P450 and NADPH–cytochrome P450 reductase. Nat. Protoc. 2009, 4, 1245–1251. [Google Scholar] [CrossRef] [Green Version]
- Omura, T.; Sato, R. The Carbon Monoxide-binding Pigment of Liver Microsomes. J. Biol. Chem. 1964, 239, 2370–2378. [Google Scholar] [CrossRef]
- Greenspan, P.; Fowler, S.D. Spectrofluorometric studies of the lipid probe, nile red. J. Lipid Res. 1985, 26, 781–789. [Google Scholar] [CrossRef]
- Szczepaniak, J.; Łukaszewicz, M.; Krasowska, A. Detection of inhibitors of Candida albicans Cdr transporters using a diS-C3(3) fluorescence. Front. Microbiol. 2015, 6, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warrilow, A.G.S.; Martel, C.M.; Parker, J.E.; Melo, N.; Lamb, D.C.; Nes, W.D.; Kelly, D.E.; Kelly, S.L. Azole Binding Properties of Candida albicans Sterol 14-α Demethylase (CaCYP51). Antimicrob. Agents Chemother. 2010, 54, 4235–4245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepesheva, G.I.; Nes, W.D.; Zhou, W.; Hill, G.C.; Waterman, M.R. CYP51 from Trypanosoma brucei Is Obtusifoliol-Specific. Biochemistry 2004, 43, 10789–10799. [Google Scholar] [CrossRef] [PubMed]
- Sagatova, A.A.; Keniya, M.V.; Tyndall, J.D.A.; Monk, B.C. Impact of Homologous Resistance Mutations from Pathogenic Yeast on Saccharomyces cerevisiae Lanosterol 14α-Demethylase. Antimicrob. Agents Chemother. 2018, 62, e02242-17. [Google Scholar] [CrossRef] [Green Version]
- Warrilow, A.G.; Nishimoto, A.T.; Parker, J.E.; Price, C.L.; Flowers, S.A.; Kelly, D.E.; Rogers, P.D.; Kelly, S.L. The Evolution of Azole Resistance in Candida albicans Sterol 14α-Demethylase (CYP51) through Incremental Amino Acid Substitutions. Antimicrob. Agents Chemother. 2019, 63, e02586-18. [Google Scholar] [CrossRef] [Green Version]
- Riley, J.; Brand, S.; Voice, M.; Caballero, I.; Calvo, D.; Read, K.D. Development of a Fluorescence-based Trypanosoma cruzi CYP51 Inhibition Assay for Effective Compound Triaging in Drug Discovery Programmes for Chagas Disease. PLoS Negl. Trop. Dis. 2015, 9, e0004014. [Google Scholar] [CrossRef]
- Decottignies, A.; Kolaczkowski, M.; Balzi, E.; Goffeau, A. Solubilization and characterization of the overexpressed PDR5 multidrug resistance nucleotide triphosphatase of yeast. J. Biol. Chem. 1994, 269, 12797–12803. [Google Scholar] [CrossRef]
- Niimi, M.; Niimi, K.; Tanabe, K.; Cannon, R.D.; Lamping, E. Inhibitor-Resistant Mutants Give Important Insights into Candida albicans ABC Transporter Cdr1 Substrate Specificity and Help Elucidate Efflux Pump Inhibition. Antimicrob. Agents Chemother. 2022, 66, e01748-21. [Google Scholar] [CrossRef]
- Rath, A.; Glibowicka, M.; Nadeau, V.G.; Chen, G.; Deber, C.M. Detergent binding explains anomalous SDS-PAGE migration of membrane proteins. Proc. Natl. Acad. Sci. USA 2009, 106, 1760–1765. [Google Scholar] [CrossRef] [Green Version]
- Rath, A.; Deber, C.M. Correction factors for membrane protein molecular weight readouts on sodium dodecyl sulfate–polyacrylamide gel electrophoresis. Anal. Biochem. 2013, 434, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Flowers, S.A.; Colón, B.; Whaley, S.G.; Schuler, M.A.; Rogers, P.D. Contribution of Clinically Derived Mutations in ERG11 to Azole Resistance in Candida albicans. Antimicrob. Agents Chemother. 2015, 59, 450–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, A.R.; Lin, Y.-H.; Niimi, K.; Lamping, E.; Keniya, M.; Niimi, M.; Tanabe, K.; Monk, B.C.; Cannon, R.D. ABC Transporter Cdr1p Contributes More than Cdr2p Does to Fluconazole Efflux in Fluconazole-Resistant Candida albicans Clinical Isolates. Antimicrob. Agents Chemother. 2008, 52, 3851–3862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagatova, A.A.; Keniya, M.V.; Wilson, R.K.; Monk, B.C.; Tyndall, J.D.A. Structural Insights into Binding of the Antifungal Drug Fluconazole to Saccharomyces cerevisiae Lanosterol 14α-Demethylase. Antimicrob. Agents Chemother. 2015, 59, 4982–4989. [Google Scholar] [CrossRef] [Green Version]
- Decottignies, A.; Grant, A.M.; Nichols, J.W.; de Wet, H.; McIntosh, D.B.; Goffeau, A. ATPase and Multidrug Transport Activities of the Overexpressed Yeast ABC Protein Yor1p. J. Biol. Chem. 1998, 273, 12612–12622. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antifungal Class | Drug Target | Resistance Mechanism in C. auris | Clade | References |
|---|---|---|---|---|
| Azoles | Erg11 | Erg11 point mutation | [7,12,15,16,17,18] | |
| F126L | III | |||
| Y132F | I, IV, V | |||
| K143R | I, IV | |||
| Erg11 copy number variants | I, III | [15,19] | ||
| Efflux pump gene overexpression MDR1 and CDR1 | I, III, IV | [20,21] | ||
| Mutation in Tac1b | [22,23,24] | |||
| A640V | I | |||
| A657V | I | |||
| F862_N866del | IV | |||
| Mutation in Mrr1 | ||||
| N647T | III | |||
| Echinocandins | β-1,3-glucan synthase | Mutation in Fks1 hot spot 1 | I, III, IV | [17,25,26,27,28] |
| S639F/P/Y | ||||
| F635Y/L | ||||
| Mutation in Fks1 hot spot 2 | I, III, IV | |||
| R1354S | ||||
| Polyenes | Ergosterol | Primary mechanism unknown ERG6 mutations | I, III, IV | [29,30] |
| Average MIC80 in µM/mg·L−1 | ||||||||
|---|---|---|---|---|---|---|---|---|
| Strain | FLC | VRC | POS | ITC | ISA | VT-1161 | MFG | AMB |
| ADΔΔ (host) | 2.58/0.8 | 0.08/0.03 | 0.22/0.15 | 0.21/0.15 | 0.015/0.007 | 0.018/0.009 | 0.23/0.29 | 3.31/3.06 |
| CauErg11 | 3.90/1.2 (1.5) | 0.22/0.08 (2.8) | 0.33/0.23 (1.5) | 0.21/0.15 (1) | 0.020/0.009 (1.3) | 0.05/0.03 (2.8) | 0.50/0.64 (2.2) | 5/4.62 (1.5) |
| CauErg11 Y132F | 250/76.6 (97) | 2/0.7 (25) | 0.20/0.14 (0.9) | 0.20/0.14 (0.95) | 0.015/0.007 (1) | 0.125/0.07 (7) | 0.34/0.43 (1.5) | 3.12/2.88 (0.9) |
| CauErg11 K143R | 56.5/17.3 (22) | 0.50/0.17 (6.25) | 0.18/0.12 (0.8) | 0.125/0.09 (0.6) | 0.020/0.009 (1.3) | 0.03/0.02 (1.7) | 0.34/0.43 (1.5) | 3.75/3.47 (1.1) |
| CauMdr1 | >400/>122 (>155) | 10.31/3.6 (129) | 0.20/0.14 (0.9) | 0.23/0.16 (1.1) | 0.10/0.04 (6.7) | 0.068/0.04 (3.8) | 0.24/0.3 (1) | 2.58/2.38 (0.8) |
| CauCdr1 | >400/>122 (>155) | 20.63/7.2 (258) | 1.08/0.76 (5) | >10/>7.06 (>47) | 12.5/5.5 (833) | >50/>26.4 (>2777) | 0.23/0.29 (1) | 1.90/1.76 (0.6) |
| Strain Designation | Azole | ΔAmax | Best Fit Equation | Kd (µM) |
|---|---|---|---|---|
| CauErg11 | Voriconazole | 0.09 | Michaelis-Menten | 0.74 (±0.09) |
| CauErg11 | Posaconazole | 0.04 | Hill | 0.34 (±0.04) |
| CauErg11 | Isavuconazole | 0.04 | Hill | 0.20 (±0.02) |
| CauErg11 Y132F | Voriconazole | 0.01 | Hill | 0.7 |
| CauErg11 Y132F | Posaconazole | 0.02 | Hill | 0.36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toepfer, S.; Lackner, M.; Keniya, M.V.; Monk, B.C. Functional Expression of Recombinant Candida auris Proteins in Saccharomyces cerevisiae Enables Azole Susceptibility Evaluation and Drug Discovery. J. Fungi 2023, 9, 168. https://doi.org/10.3390/jof9020168
Toepfer S, Lackner M, Keniya MV, Monk BC. Functional Expression of Recombinant Candida auris Proteins in Saccharomyces cerevisiae Enables Azole Susceptibility Evaluation and Drug Discovery. Journal of Fungi. 2023; 9(2):168. https://doi.org/10.3390/jof9020168
Chicago/Turabian StyleToepfer, Stephanie, Michaela Lackner, Mikhail V. Keniya, and Brian C. Monk. 2023. "Functional Expression of Recombinant Candida auris Proteins in Saccharomyces cerevisiae Enables Azole Susceptibility Evaluation and Drug Discovery" Journal of Fungi 9, no. 2: 168. https://doi.org/10.3390/jof9020168