
Trichoderma pubescens Elicit Induced Systemic Resistance in Tomato Challenged by Rhizoctonia solani

 ,
, 

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of Pathogen and Bioagents
2.2. Effect of Trichoderma Isolates on Growth of Rhizoctonia
2.3. Effect of Fungicides on R. solani In Vitro
2.4. Identification of R. solani and Trichoderma Isolates
2.5. Biocontrol Potential and Plant Growth-Promoting Abilities of Trichoderma Isolate against R. solani on Tomato Plants
2.6. Analysis of Defense-Related-Genes
2.6.1. Extraction of RNA and Synthesis of cDNA
2.6.2. Assay of qRT-PCR
2.7. Oxidative Stress Markers
2.7.1. Analysis of Malondialdehyde
2.7.2. Determination of Hydrogen Peroxide
2.8. Antioxidant Enzymatic Activities Measurement
2.8.1. Polyphenol Oxidase Enzyme
2.8.2. Peroxidase Enzyme
2.8.3. Catalase Enzyme
2.8.4. Superoxide Dismutase Enzyme
2.9. Polyphenolic Components in Tomato Leaves
2.9.1. Preparation of Tomato Samples for Phenolic Analysis
2.9.2. HPLC Analysis of Tomato-Collected Leaves Extract
2.10. Statistical Analysis
3. Results
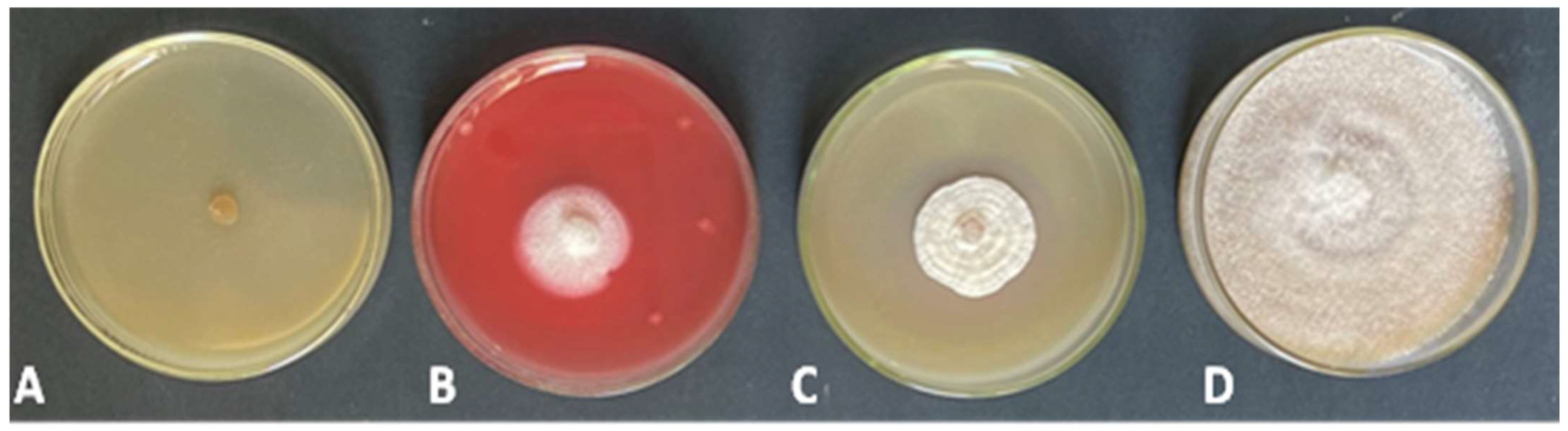
3.1. Rhizoctonia, Trichoderma Isolation and Identification
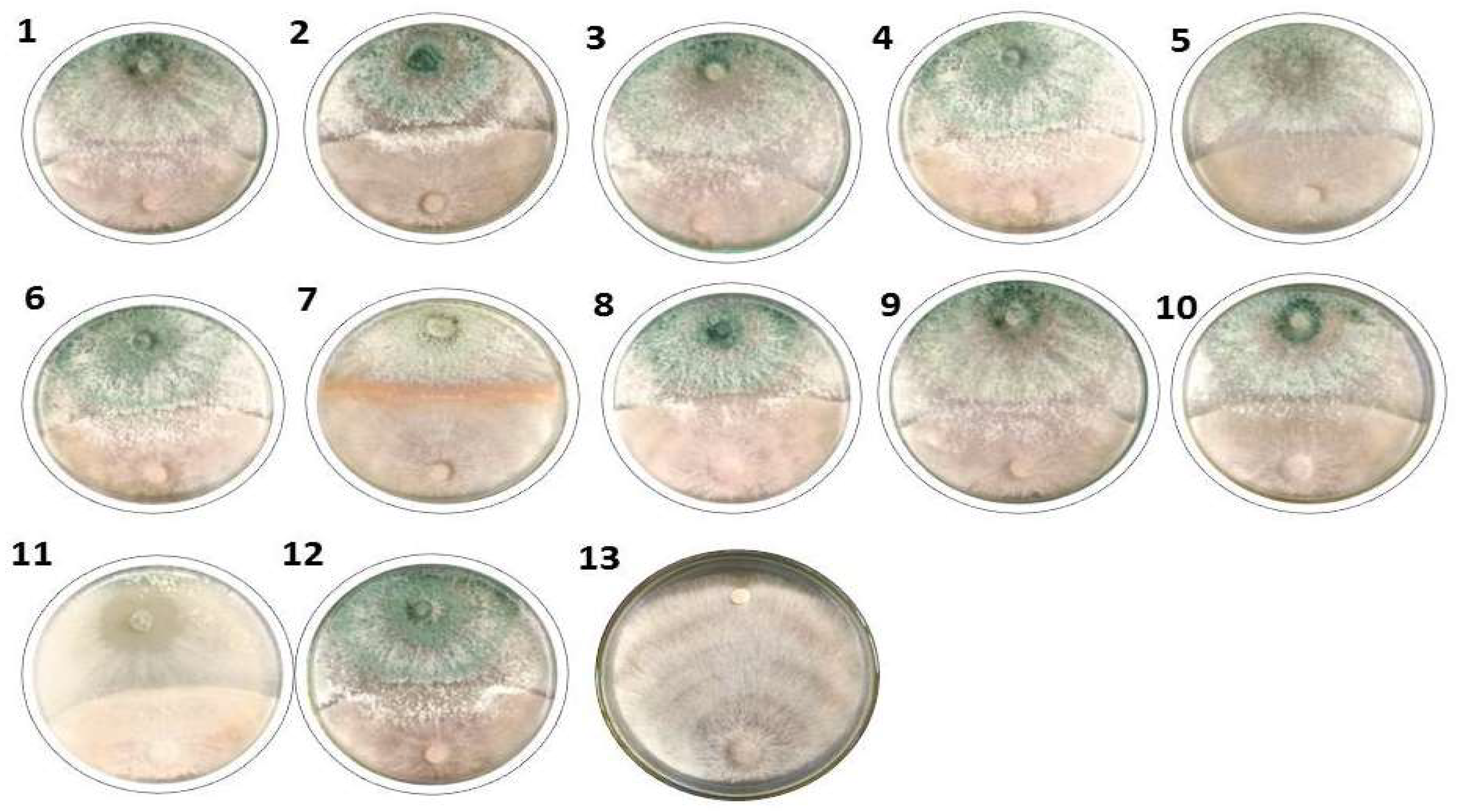
3.2. Effect of Trichoderma Isolates on R. solani In Vitro
3.3. Effect of Fungicides on R. solani In Vitro
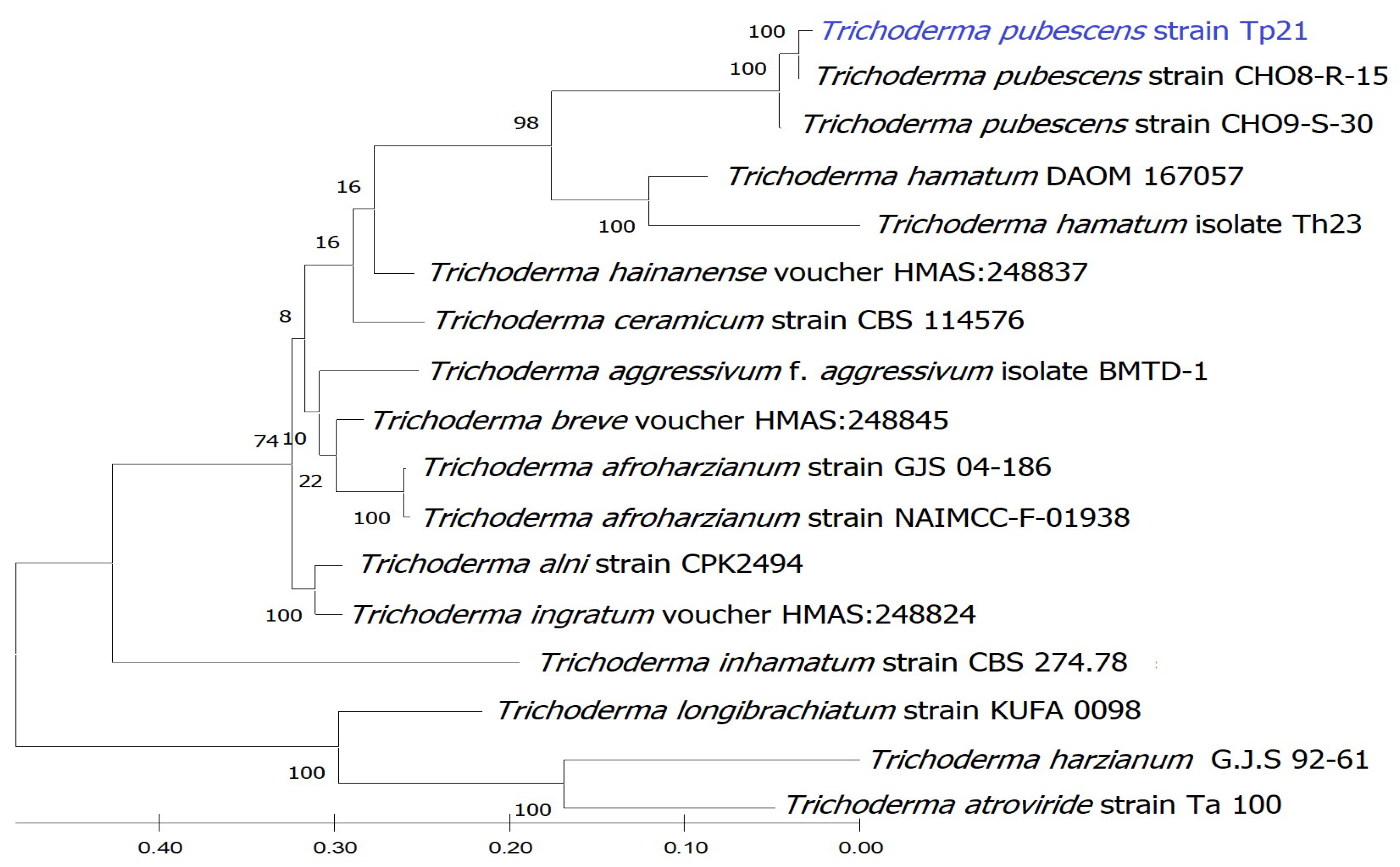
3.4. Molecular Identification of R. solani and Trichoderma spp.
3.5. Effect of T. pubescens on R. solani Root Rot In Vivo
3.6. Efficacy of T. pubescens on Tomato Growth Parameters
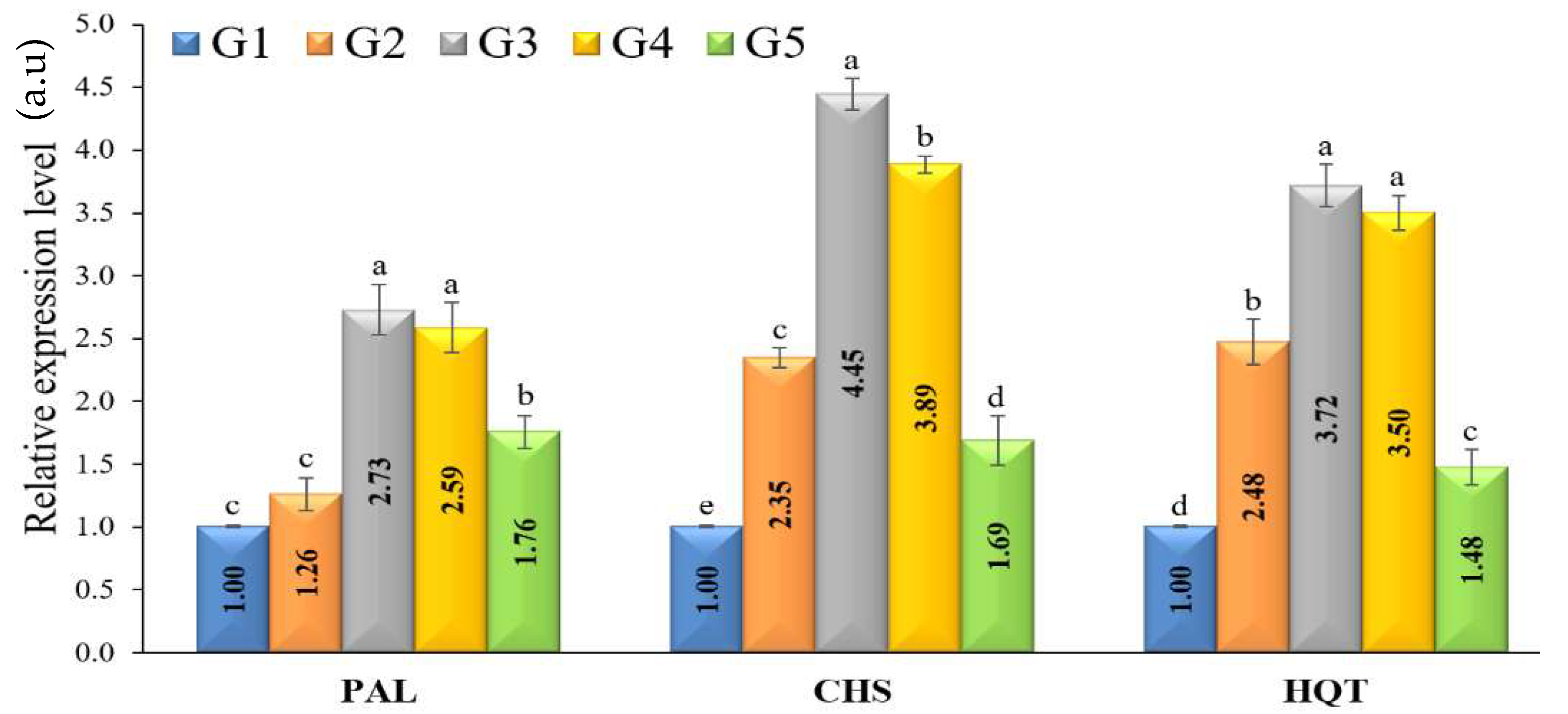
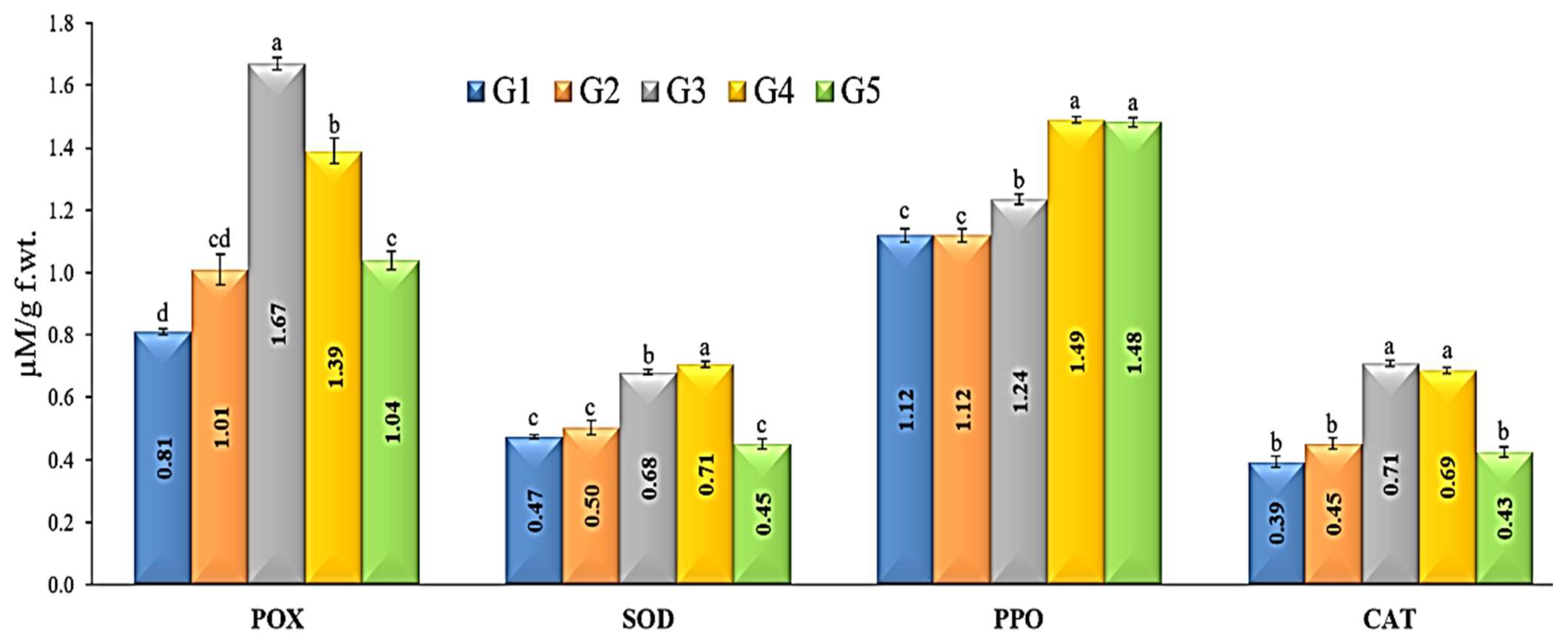
3.7. Defense-Related Enzymes Activity
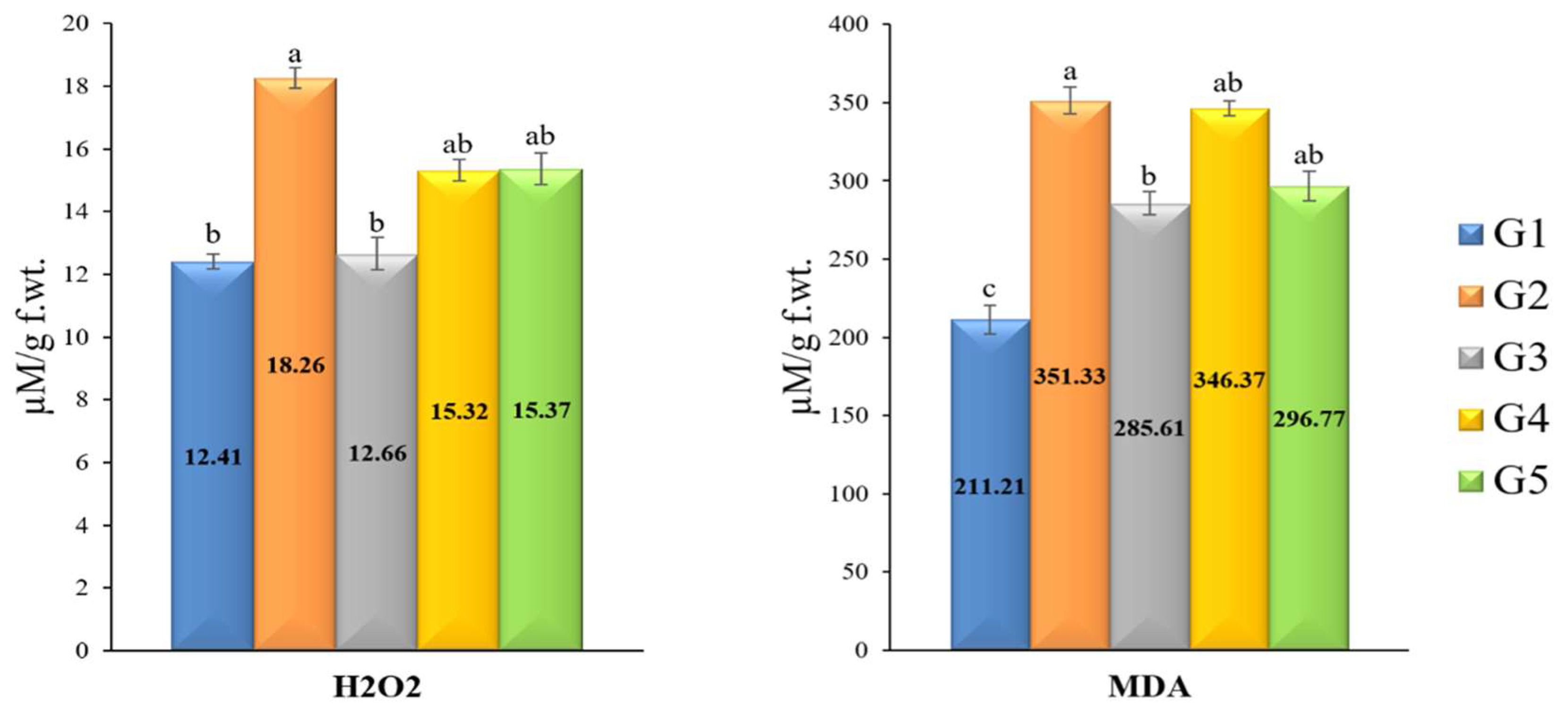
3.8. H2O2 and MDA Kinetics
3.9. Antioxidant Enzymes Activities
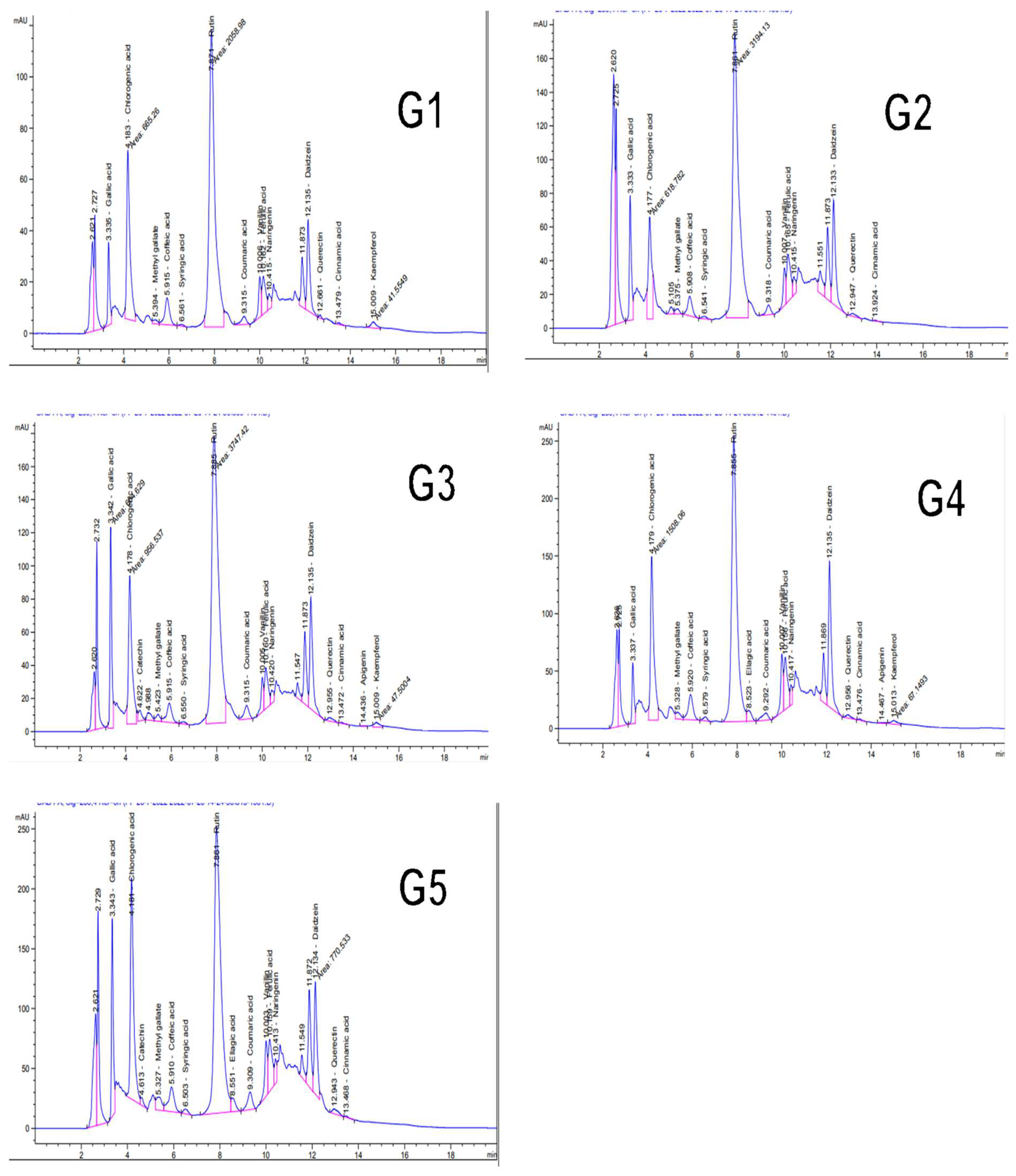
3.10. Phytochemical Analysis of Tomato Leaf Extract
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Georgé, S.; Tourniaire, F.; Gautier, H.; Goupy, P.; Rock, E.; Caris-Veyrat, C. Changes in the contents of carotenoids, phenolic compounds and vitamin C during technical processing and lyophilisation of red and yellow tomatoes. Food Chem. 2011, 124, 1603–1611. [Google Scholar] [CrossRef]
- Mickelbart, M.V.; Hasegawa, P.M.; Bailey-Serres, J. Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability. Nat. Rev. Genet. 2015, 16, 237–251. [Google Scholar] [CrossRef] [PubMed]
- Abdelkhalek, A.; Hafez, E. Plant viral diseases in Egypt and their control. In Cottage Industry of Biocontrol Agents and Their Applications; Springer: Berlin/Heidelberg, Germany, 2020; pp. 403–421. [Google Scholar]
- Singh, N.; Raina, S.; Singh, D.; Ghosh, M.; Heflish, A. Exploitation of promising native strains of Bacillus subtilis with antagonistic properties against fungal pathogens and their PGPR characteristics. J. Plant Pathol. 2017, 99, 27–35. [Google Scholar]
- Heflish, A.A.; Abdelkhalek, A.; Al-Askar, A.A.; Behiry, S.I. Protective and curative effects of Trichoderma asperelloides Ta41 on tomato root rot caused by Rhizoctonia solani Rs33. Agronomy 2021, 11, 1162. [Google Scholar] [CrossRef]
- Channa, M.Y.; Pathan, M.A.; Solangi, G.R.; Wondiar, M. Studies on Rhizoctonia solani (Kuhn) causing root rot of lentil. Sarhad J. Agric. 1995, 11, 495–499. [Google Scholar]
- Halifu, S.; Deng, X.; Song, X.; Song, R. Effects of two Trichoderma strains on plant growth, rhizosphere soil nutrients, and fungal community of Pinus sylvestris var. mongolica annual seedlings. Forests 2019, 10, 758. [Google Scholar]
- Hahn, M. The rising threat of fungicide resistance in plant pathogenic fungi: Botrytis as a case study. J. Chem. Biol. 2014, 7, 133–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakade, R.R.; Sundaresha, S.; Lal, M. Management strategies and alternatives for fungicidal resistance in potato. In Pesticides-Updates on Toxicity, Efficacy and Risk Assessment; IntechOpen: London, UK, 2022; ISBN 1803560398. [Google Scholar]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Barka, E.A. Biological control of plant pathogens: A global perspective. Microorganisms 2022, 10, 596. [Google Scholar] [CrossRef]
- Punja, Z.K.; Utkhede, R.S. Using fungi and yeasts to manage vegetable crop diseases. Trends Biotechnol. 2003, 21, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Shoresh, M.; Harman, G.E. Differential expression of maize chitinases in the presence or absence of Trichoderma harzianum strain T22 and indications of a novel exo-endo-heterodimeric chitinase activity. BMC Plant Biol. 2010, 10, 136. [Google Scholar] [CrossRef] [Green Version]
- Vinale, F.; Marra, R.; Scala, F.; Ghisalberti, E.L.; Lorito, M.; Sivasithamparam, K. Major secondary metabolites produced by two commercial Trichoderma strains active against different phytopathogens. Lett. Appl. Microbiol. 2006, 43, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Montero-Barrientos, M.; Hermosa, R.; Cardoza, R.E.; Gutiérrez, S.; Monte, E. Functional analysis of the Trichoderma harzianum nox1 gene, encoding an NADPH oxidase, relates production of reactive oxygen species to specific biocontrol activity against Pythium ultimum. Appl. Environ. Microbiol. 2011, 77, 3009–3016. [Google Scholar] [CrossRef] [Green Version]
- Tapwal, A.; Pandey, H. In vitro evaluation of Trichoderma species for virulence efficacy on Botryodiplodia palmarum. Curr. Life Sci. 2016, 2, 86–91. [Google Scholar]
- Halifu, S.; Deng, X.; Song, X.; Song, R.; Liang, X. Inhibitory mechanism of Trichoderma virens ZT05 on Rhizoctonia solani. Plants 2020, 9, 912. [Google Scholar] [CrossRef] [PubMed]
- Kredics, L.; Chen, L.; Kedves, O.; Büchner, R.; Hatvani, L.; Allaga, H.; Nagy, V.D.; Khaled, J.M.; Alharbi, N.S.; Vágvölgyi, C. Molecular tools for monitoring Trichoderma in agricultural environments. Front. Microbiol. 2018, 9, 1599. [Google Scholar] [CrossRef] [PubMed]
- Berini, F.; Caccia, S.; Franzetti, E.; Congiu, T.; Marinelli, F.; Casartelli, M.; Tettamanti, G. Effects of Trichoderma viride chitinases on the peritrophic matrix of Lepidoptera. Pest Manag. Sci. 2016, 72, 980–989. [Google Scholar] [CrossRef]
- Xiong, H.; Xue, K.; Qin, W.; Chen, X.; Wang, H.; Shi, X.; Ma, T.; Sun, Z.; Chen, W.; Tian, X. Does soil treated with conidial formulations of Trichoderma spp. attract or repel subterranean termites? J. Econ. Entomol. 2018, 111, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.L.; Lorito, M. Exploiting the interactions between fungal antagonists, pathogens and the plant for biocontrol. In Novel Biotechnologies for Biocontrol Agent Enhancement and Management; Springer: Berlin/Heidelberg, Germany, 2007; pp. 107–130. [Google Scholar]
- Malmierca, M.G.; Barua, J.; McCormick, S.P.; Izquierdo-Bueno, I.; Cardoza, R.E.; Alexander, N.J.; Hermosa, R.; Collado, I.G.; Monte, E.; Gutiérrez, S. Novel aspinolide production by T richoderma arundinaceum with a potential role in Botrytis cinerea antagonistic activity and plant defence priming. Environ. Microbiol. 2015, 17, 1103–1118. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Horwitz, B.A.; Herrera-Estrella, A.; Schmoll, M.; Kenerley, C.M. Trichoderma research in the genome era. Annu. Rev. Phytopathol. 2013, 51, 105–129. [Google Scholar] [CrossRef] [PubMed]
- Heflish, A.I.A.I.; Singh, N.; Raina, S.; Buttar, D.S. Evaluation of Trichoderma isolates against Rhizoctonia solani and Rhizoctonia oryzae causing sheath blight of rice. Plant Dis. Res. 2017, 32, 36–46. [Google Scholar]
- Elad, Y.; Chet, I.; Katan, J. Trichoderma harzianum: A biocontrol agent effective against Sclerotium rolfsii and Rhizoctonia solani. Phytopathology 1980, 70, 119–121. [Google Scholar] [CrossRef] [Green Version]
- Fahmi, A.I.; Al-Talhi, A.D.; Hassan, M.M. Protoplast fusion enhances antagonistic activity in Trichoderma spp. Nat. Sci. 2012, 10, 100–106. [Google Scholar]
- Singh, G.; Milne, K.S. Laboratory evaluation of fungicides against fungi causing flower blight of chrysanthemums. N. Z. J. Exp. Agric. 1974, 2, 181–183. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Samuels, G.J.; Dodd, S.L.; Gams, W.; Castlebury, L.A.; Petrini, O. Trichoderma species associated with the green mold epidemic of commercially grown Agaricus bisporus. Mycologia 2002, 94, 146–170. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Behiry, S.I.; Al-Askar, A.A. Bacillus velezensis PEA1 inhibits Fusarium oxysporum growth and induces systemic resistance to Cucumber mosaic virus. Agronomy 2020, 10, 1312. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guid. Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Hatvani, L.; Antal, Z.; Manczinger, L.; Szekeres, A.; Druzhinina, I.S.; Kubicek, C.P.; Nagy, A.; Nagy, E.; Vágvölgyi, C.; Kredics, L. Green mold diseases of Agaricus and Pleurotus spp. are caused by related but phylogenetically different Trichoderma species. Phytopathology 2007, 97, 532–537. [Google Scholar] [CrossRef] [Green Version]
- Wurz, R.P.; Charette, A.B. Doubly activated cyclopropanes as synthetic precursors for the preparation of 4-nitro-and 4-cyano-dihydropyrroles and pyrroles. Org. Lett. 2005, 7, 2313–2316. [Google Scholar] [CrossRef]
- André, C.M.; Schafleitner, R.; Legay, S.; Lefèvre, I.; Aliaga, C.A.A.; Nomberto, G.; Hoffmann, L.; Hausman, J.-F.; Larondelle, Y.; Evers, D. Gene expression changes related to the production of phenolic compounds in potato tubers grown under drought stress. Phytochemistry 2009, 70, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Abdeljalil, N.O.-B.; Vallance, J.; Gerbore, J.; Bruez, E.; Martins, G.; Rey, P.; Daami-Remadi, M. Biocontrol of Rhizoctonia root rot in tomato and enhancement of plant growth using rhizobacteria naturally associated to tomato. J. Plant Pathol. Microbiol. 2016, 7, 1000356. [Google Scholar] [CrossRef] [Green Version]
- Abdelkhalek, A.; Sanan-Mishra, N. Differential expression profiles of tomato miRNAs induced by Tobacco mosaic virus. J. Agric. Sci. Technol. 2019, 21, 475–485. [Google Scholar]
- Behiry, S.I.; Ashmawy, N.A.; Abdelkhalek, A.A.; Younes, H.A.; Khaled, A.E.; Hafez, E.E. Compatible- and incompatible-type interactions related to defense genes in potato elucidation by Pectobacterium carotovorum. J. Plant Dis. Prot. 2018, 125, 197–204. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Dessoky, E.S.; Hafez, E. Polyphenolic genes expression pattern and their role in viral resistance in tomato plant infected with Tobacco mosaic virus. Biosci. Res. 2018, 15, 3349–3356. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Junglee, S.; Urban, L.; Sallanon, H.; Lopez-Lauri, F. Optimized assay for hydrogen peroxide determination in plant tissue using potassium iodide. Am. J. Anal. Chem. 2014, 5, 730–736. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.L.; Zhong, H.W.; Zhang, A.Q. Plant Physiology Experimentation; Peking University Press: Beijing, China, 1990. [Google Scholar]
- Moerschbacher, B.M.; Noll, U.M.; Flott, B.E.; Reisener, H.-J. Lignin biosynthetic enzymes in stem rust infected, resistant and susceptible near-isogenic wheat lines. Physiol. Mol. Plant Pathol. 1988, 33, 33–46. [Google Scholar] [CrossRef]
- Cakmak, I.; Marschner, H. Magnesium deficiency and high light intensity enhance activities of superoxide dismutase, ascorbate peroxidase, and glutathione reductase in bean leaves. Plant Physiol. 1992, 98, 1222–1227. [Google Scholar] [CrossRef] [Green Version]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Kumar, A.; Dutt, S.; Bagler, G.; Ahuja, P.S.; Kumar, S. Engineering a thermo-stable superoxide dismutase functional at sub-zero to >50 °C, which also tolerates autoclaving. Sci. Rep. 2012, 2, 387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelkhalek, A.; Al-Askar, A.A.; Alsubaie, M.M.; Behiry, S.I. First Report of protective activity of Paronychia argentea extract against Tobacco mosaic virus infection. Plants 2021, 10, 2435. [Google Scholar] [CrossRef] [PubMed]
- El-Kazzaz, M.K.; Ghoneim, K.E.; Agha, M.K.M.; Helmy, A.; Behiry, S.I.; Abdelkhalek, A.; Saleem, M.H.; Al-Askar, A.A.; Arishi, A.A.; Elsharkawy, M.M. Suppression of pepper root rot and wilt diseases caused by Rhizoctonia solani and Fusarium oxysporum. Life 2022, 12, 587. [Google Scholar] [CrossRef]
- Sharma, A. Fungi as Biological Control Agents. In Biofertilizers for Sustainable Agriculture and Environment; Springer: Berlin/Heidelberg, Germany, 2019; pp. 395–411. [Google Scholar]
- Abdelkhalek, A.; Aseel, D.G.; Király, L.; Künstler, A.; Moawad, H.; Al-Askar, A.A. Induction of systemic resistance to Tobacco mosaic virus in tomato through foliar application of Bacillus amyloliquefaciens Strain TBorg1 culture filtrate. Viruses 2022, 14, 1830. [Google Scholar] [CrossRef]
- Lamenew, F.; Mekonnen, H.; Gashaw, T. Biocontrol potential of Trichoderma and yeast against post harvest fruit fungal diseases: A review. World News Nat. Sci. 2019, 27, 153–173. [Google Scholar]
- Siameto, E.N.; Okoth, S.; Amugune, N.O.; Chege, N.C. Antagonism of Trichoderma harzianum isolates on soil borne plant pathogenic fungi from Embu District, Kenya. J. Yeast Fungal Res. 2010, 1, 47–54. [Google Scholar]
- Guédez, C.; Cañizalez, L.; Castillo, C.; Olivar, R. Evaluación in vitro de aislamientos de Trichoderma harzianum para el control de Rhizoctonia solani, Sclerotium rolfsii y Fusarium oxysporum en plantas de tomate. Rev. Soc. Venez. Microbiol. 2012, 32, 44–49. [Google Scholar]
- Ramírez-Cariño, H.F.; Guadarrama-Mendoza, P.C.; Sánchez-López, V.; Cuervo-Parra, J.A.; Ramírez-Reyes, T.; Dunlap, C.A.; Valadez-Blanco, R. Biocontrol of Alternaria alternata and Fusarium oxysporum by Trichoderma asperelloides and Bacillus paralicheniformis in tomato plants. Antonie Van Leeuwenhoek 2020, 113, 1247–1261. [Google Scholar] [CrossRef]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “secrets” of a multitalented biocontrol agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Yedidia, I.; Srivastva, A.K.; Kapulnik, Y.; Chet, I. Effect of Trichoderma harzianum on microelement concentrations and increased growth of cucumber plants. Plant Soil 2001, 235, 235–242. [Google Scholar] [CrossRef]
- Tchameni, S.N.; Sameza, M.L.; O’Donovan, A.; Fokom, R.; Mangaptche Ngonkeu, E.L.; Wakam Nana, L.; Etoa, F.-X.; Nwaga, D. Antagonism of Trichoderma asperellum against Phytophthora megakarya and its potential to promote cacao growth and induce biochemical defence. Mycology 2017, 8, 84–92. [Google Scholar] [CrossRef] [Green Version]
- Sundaramoorthy, S.; Balabaskar, P. Biocontrol efficacy of Trichoderma spp. against wilt of tomato caused by Fusarium oxysporum f. sp. lycopersici. J. Appl. Biol. Biotechnol. 2013, 1, 036–040. [Google Scholar]
- Casimiro, I.; Marchant, A.; Bhalerao, R.P.; Beeckman, T.; Dhooge, S.; Swarup, R.; Graham, N.; Inzé, D.; Sandberg, G.; Casero, P.J. Auxin transport promotes Arabidopsis lateral root initiation. Plant Cell 2001, 13, 843–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alexandria J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Zehra, A.; Meena, M.; Dubey, M.K.; Aamir, M.; Upadhyay, R.S. Synergistic effects of plant defense elicitors and Trichoderma harzianum on enhanced induction of antioxidant defense system in tomato against Fusarium wilt disease. Bot. Stud. 2017, 58, 44. [Google Scholar] [CrossRef] [Green Version]
- Garg, N.; Manchanda, G. ROS generation in plants: Boon or bane? Plant Biosyst. 2009, 143, 81–96. [Google Scholar] [CrossRef]
- Masuta, C.; Inaba, J.; Shimura, H. The 2b proteins of Cucumber mosaic virus generally have the potential to differentially induce necrosis on Arabidopsis. Plant Signal. Behav. 2012, 7, 43–45. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.S.; Na, H.; Kwack, Y.; Chun, C. Secondary metabolite profiling in various parts of tomato plants. Hortic. Sci. Technol. 2014, 32, 252–260. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.S.; Kwack, Y.; Lee, J.H.; Chun, C. Antimicrobial activity of various parts of tomato plants varied with different solvent extracts. Plant Pathol. J. 2019, 35, 149. [Google Scholar] [CrossRef] [PubMed]
- Ejike, C.E.C.C.; Gong, M.; Udenigwe, C.C. Phytoalexins from the Poaceae: Biosynthesis, function and prospects in food preservation. Food Res. Int. 2013, 52, 167–177. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Varoni, E.M.; Iriti, M.; Martorell, M.; Setzer, W.N.; del Mar Contreras, M.; Salehi, B.; Soltani-Nejad, A.; Rajabi, S.; Tajbakhsh, M. Carvacrol and human health: A comprehensive review. Phyther. Res. 2018, 32, 1675–1687. [Google Scholar] [CrossRef] [PubMed]
- Ortega-García, J.G.; Montes-Belmont, R.; Rodríguez-Monroy, M.; Ramírez-Trujillo, J.A.; Suárez-Rodríguez, R.; Sepúlveda-Jiménez, G. Effect of Trichoderma asperellum applications and mineral fertilization on growth promotion and the content of phenolic compounds and flavonoids in onions. Sci. Hortic. 2015, 195, 8–16. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Amplified Region | Primer Direction | Sequence (5’-3’) | References |
|---|---|---|---|---|
| ITS | Internal transcribed spacer | ITS1 | TCCGTAGGTGAACCTGCGG | [30] |
| ITS4 | TCCTCCGCTTATTGATATGC | |||
| Rpb2 | RNA polymerase II subunit 2 | fRPB2-5f | GAYGAYMGWGATCAYTTYGG | [31] |
| fRPB2-7cr | CCCATRGCTTGYTTRCCCAT | |||
| Tef-1 | Translation elongation factor 1 alpha | EF1-728F | CATCGAGAAGTTCGAGAAGG | [32] |
| TEF1LLErev | AACTTGCAGGCAATGTGG | |||
| PAL | Phenylalanine ammonia-lyase | PAL-f | ACGGGTTGCCATCTAATCTGACA | [33] |
| PAL-r | CGAGCAATAAGAAGCCATCGCAAT | |||
| HQT | Hydroxycinnamoyl Co A quinate hydroxycinnamoyl transferase | HQT-f | CCCAATGGCTGGAAGATTAGCTA | [34] |
| HQT-r | CATGAATCACTTTCAGCCTCAACAA | |||
| CHS | Chalcone synthase | CHS-f | CACCGTGGAGGAGTATCGTAAGGC | [34] |
| CHS-r | TGATCAACACAGTTGGAAGGCG |
| Treatments | Inhibition % ± SD * |
|---|---|
| T1 | 70.74 ± 2.57 c |
| T2 | 68.52 ± 3.39 c |
| T3 | 76.93 ± 3.21 a |
| T4 | 61.85 ± 4.49 d |
| T5 | 68.52 ± 3.21 c |
| T6 | 68.52 ± 3.21 c |
| T7 | 70.37 ± 3.21 c |
| T8 | 62.96 ± 3.21 d |
| T9 | 74.07 ± 3.21 b |
| T10 | 69.26 ± 0.64 c |
| T11 | 74.07 ± 1.70 b |
| T12 | 74.44 ± 2.94 b |
| Control | 00.00 ± 0.00 e |
| Treatment | Disease Index ± SD * | Total Chlorophyll Content (SPAD) ± SD |
|---|---|---|
| G1 | 00.00 ± 0.00 c | 35.27 ± 0.90 b |
| G2 | 78.67 ± 5.58 a | 23.63 ± 1.07 c |
| G3 | 00.00 ± 0.00 c | 37.70 ± 0.70 a |
| G4 | 16.00 ± 3.65 b | 36.70 ± 0.79 ab |
| G5 | 14.67 ± 5.54 b | 35.10 ± 1.0 ab |
| Treatments ** | Length (cm) ± SD * | Fresh Weight (g) ± SD | Dry Weight (g) ± SD | |||
|---|---|---|---|---|---|---|
| Shoot | Root | Shoot | Root | Shoot | Root | |
| G1 | 13.30 ± 0.79 b | 06.23 ± 0.06 d | 4.40 ± 0.10 b | 2.03 ± 0.32 b | 1.87 ± 0.15 ab | 0.93 ± 0.15 b |
| G2 | 11.10 ± 1.11 b | 04.93 ± 0.38 e | 2.63 ± 0.15 c | 1.10 ± 0.20 b | 1.57 ± 0.31 b | 0.40 ± 0.10 c |
| G3 | 21.27 ± 0.64 a | 10.37 ± 0.12 a | 6.10 ± 0.53 a | 3.40 ± 0.10 a | 2.47 ± 0.25 a | 1.47 ± 0.15 a |
| G4 | 19.23 ± 1.44 a | 09.23 ± 0.49 b | 5.38 ± 0.13 a | 3.23 ± 0.12 a | 2.37 ± 0.55 ab | 1.37 ± 0.06 a |
| G5 | 13.27 ± 0.40 b | 07.07 ± 0.21 c | 4.53 ± 0.32 b | 1.70 ± 0.10 b | 1.90 ± 0.10 ab | 0.83 ± 0.23 b |
| Compound | Concentration (µg/g) | ||||
|---|---|---|---|---|---|
| G1 * | G2 | G3 | G4 | G5 | |
| Gallic acid | 1367.53 | 2294.84 | 4254.70 | 1275.36 | 2776.91 |
| Chlorogenic acid | 7875.70 | 5158.17 | 9981.89 | 9613.27 | 9282.39 |
| Ferulic acid | 1136.84 | 1581.69 | 1619.09 | 1839.91 | 1473.40 |
| Methyl gallate | 133.15 | 168.65 | 282.46 | 291.74 | 489.70 |
| Coffeic acid | 1235.52 | 903.82 | 1319.00 | 1430.01 | 1231.44 |
| Syringic acid | 81.76 | 126.33 | 152.03 | 217.20 | 204.89 |
| Ellagic acid | 0.00 | 0.00 | 0.00 | 1190.75 | 1083.84 |
| Coumaric acid | 136.32 | 143.42 | 294.12 | 173.35 | 237.75 |
| Cinnamic acid | 17.00 | 9.22 | 15.46 | 17.07 | 11.73 |
| Vanillin | 619.55 | 586.14 | 709.76 | 1071.24 | 805.74 |
| Catechin | 0.00 | 0.00 | 1251.23 | 0.00 | 538.17 |
| Rutin | 19,975.11 | 21,819.77 | 32,046.74 | 21,580.30 | 20,416.05 |
| Naringenin | 500.19 | 606.62 | 598.33 | 598.06 | 548.08 |
| Daidzein | 1546.99 | 2003.50 | 2671.25 | 3040.66 | 1650.84 |
| Querectin | 68.89 | 189.53 | 298.64 | 211.84 | 287.75 |
| Apigenin | 0.00 | 0.00 | 22.68 | 35.33 | 0.00 |
| Kaempferol | 511.38 | 0.00 | 515.26 | 444.95 | 0.00 |
| Total | 35,205.91 | 35,591.70 | 56,032.64 | 43,031.03 | 41,038.68 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Behiry, S.; Soliman, S.A.; Massoud, M.A.; Abdelbary, M.; Kordy, A.M.; Abdelkhalek, A.; Heflish, A. Trichoderma pubescens Elicit Induced Systemic Resistance in Tomato Challenged by Rhizoctonia solani. J. Fungi 2023, 9, 167. https://doi.org/10.3390/jof9020167
Behiry S, Soliman SA, Massoud MA, Abdelbary M, Kordy AM, Abdelkhalek A, Heflish A. Trichoderma pubescens Elicit Induced Systemic Resistance in Tomato Challenged by Rhizoctonia solani. Journal of Fungi. 2023; 9(2):167. https://doi.org/10.3390/jof9020167
Chicago/Turabian StyleBehiry, Said, Seham A. Soliman, Magdy A. Massoud, Moawad Abdelbary, Ahmed M. Kordy, Ahmed Abdelkhalek, and Ahmed Heflish. 2023. "Trichoderma pubescens Elicit Induced Systemic Resistance in Tomato Challenged by Rhizoctonia solani" Journal of Fungi 9, no. 2: 167. https://doi.org/10.3390/jof9020167