
Complement, but Not Platelets, Plays a Pivotal Role in the Outcome of Mucormycosis In Vivo

 , ,
, ,  , , and
, , and 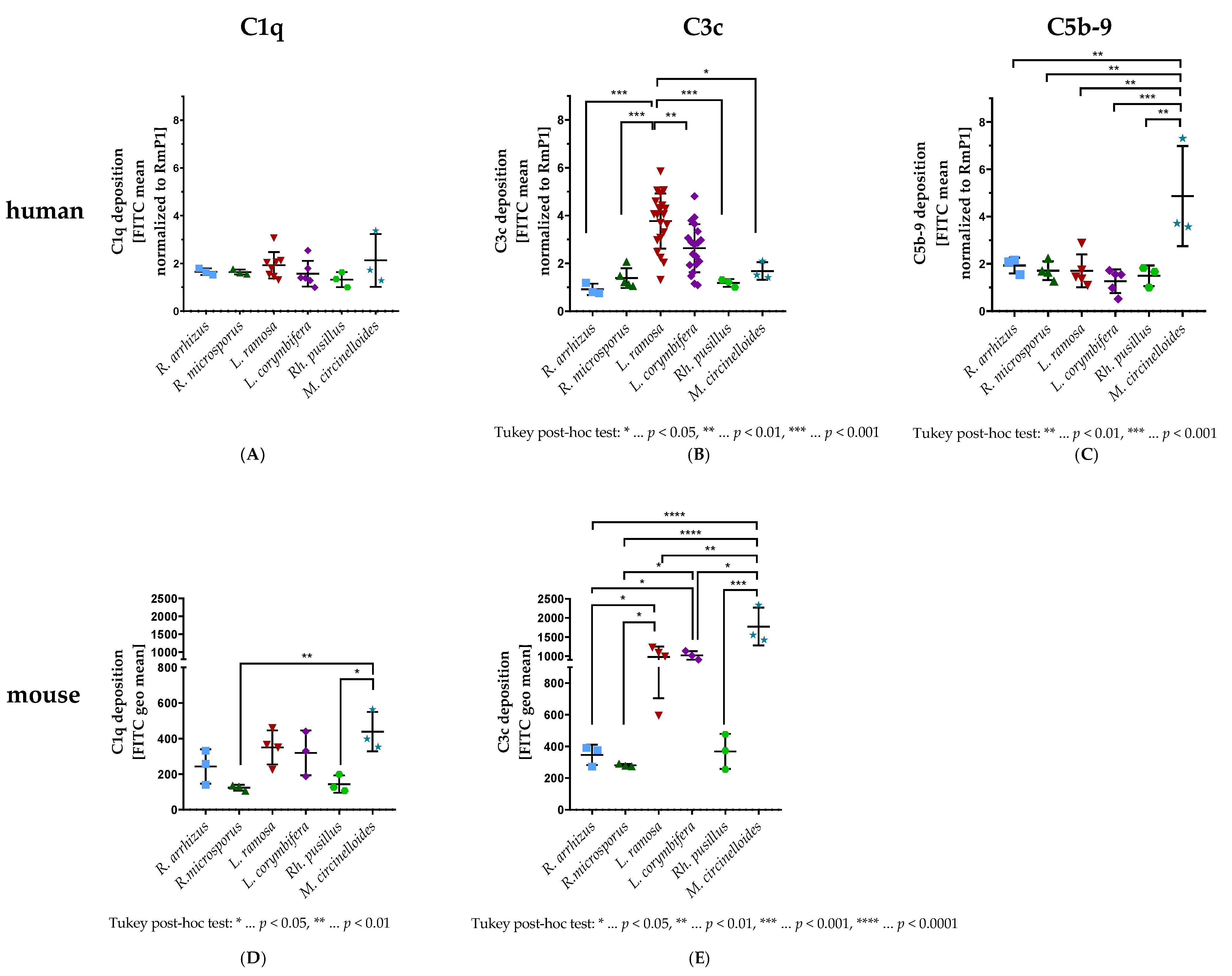{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Media
2.2. Fungal Isolates and Cultivation
2.3. Human and Murine C1q and C3c Deposition and Human C5b-9 Deposition on Mucormycete Spores
2.4. Animal Experiments
2.4.1. Ethics Statement
2.4.2. Mouse Lines, Experimental Groups, Humane Endpoints
2.4.3. Performance of Animal Experiments
2.4.4. Analysis of Blood and Organ Samples
2.5. Detection of Mucormycetes in the Kidney by Polymerase Chain Reaction
2.6. DuoSet Sandwich Enzyme-Linked Immunosorbent Assay (ELISA) to Detect Murine Soluble P-Selectin/sCD62P in Plasma
2.6.1. Plate Preparation
2.6.2. Assay Procedure
2.7. Statistical Analyses
3. Results
3.1. C1q, C3c, and C5b-9 Deposition on Spores
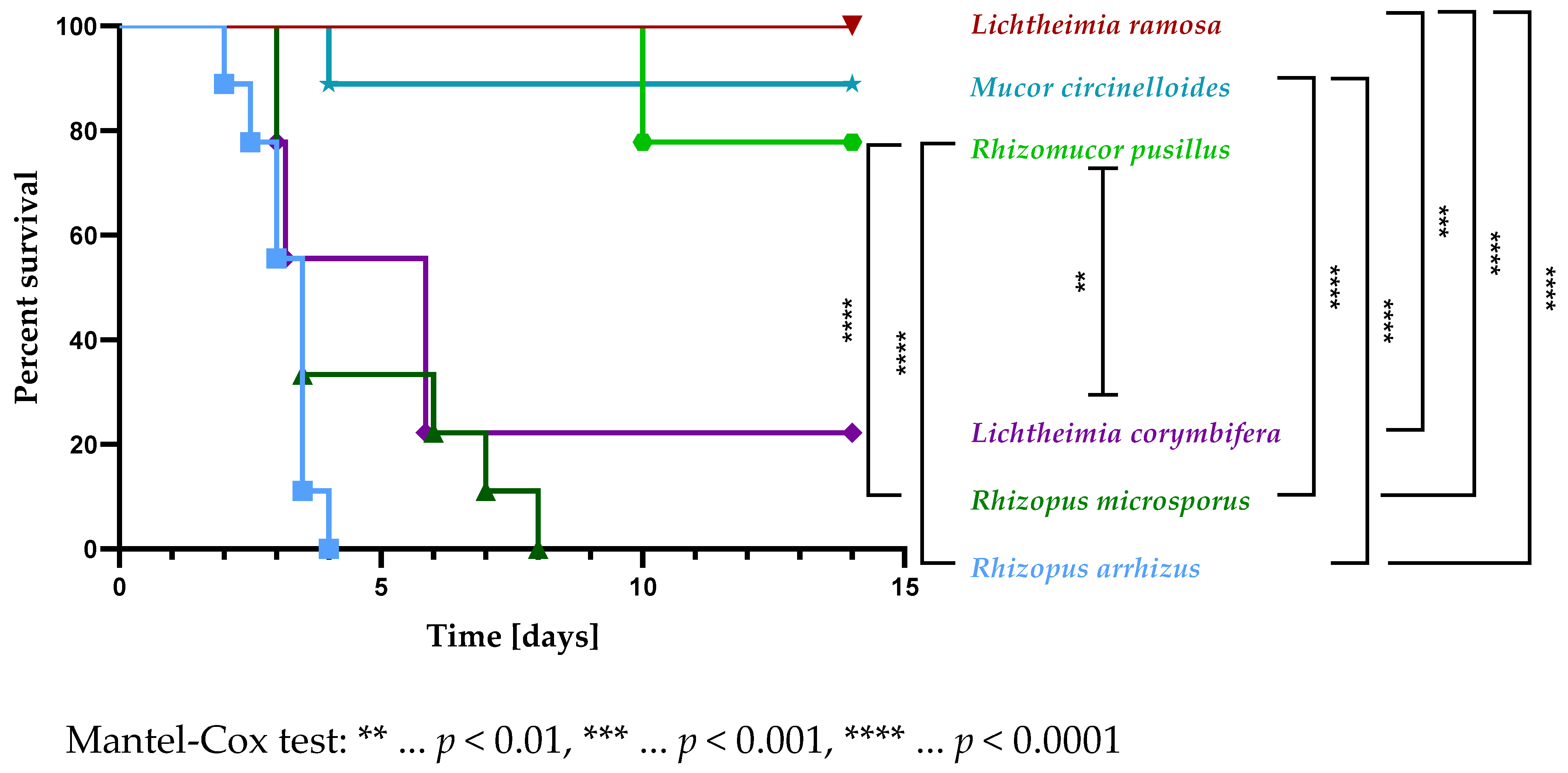
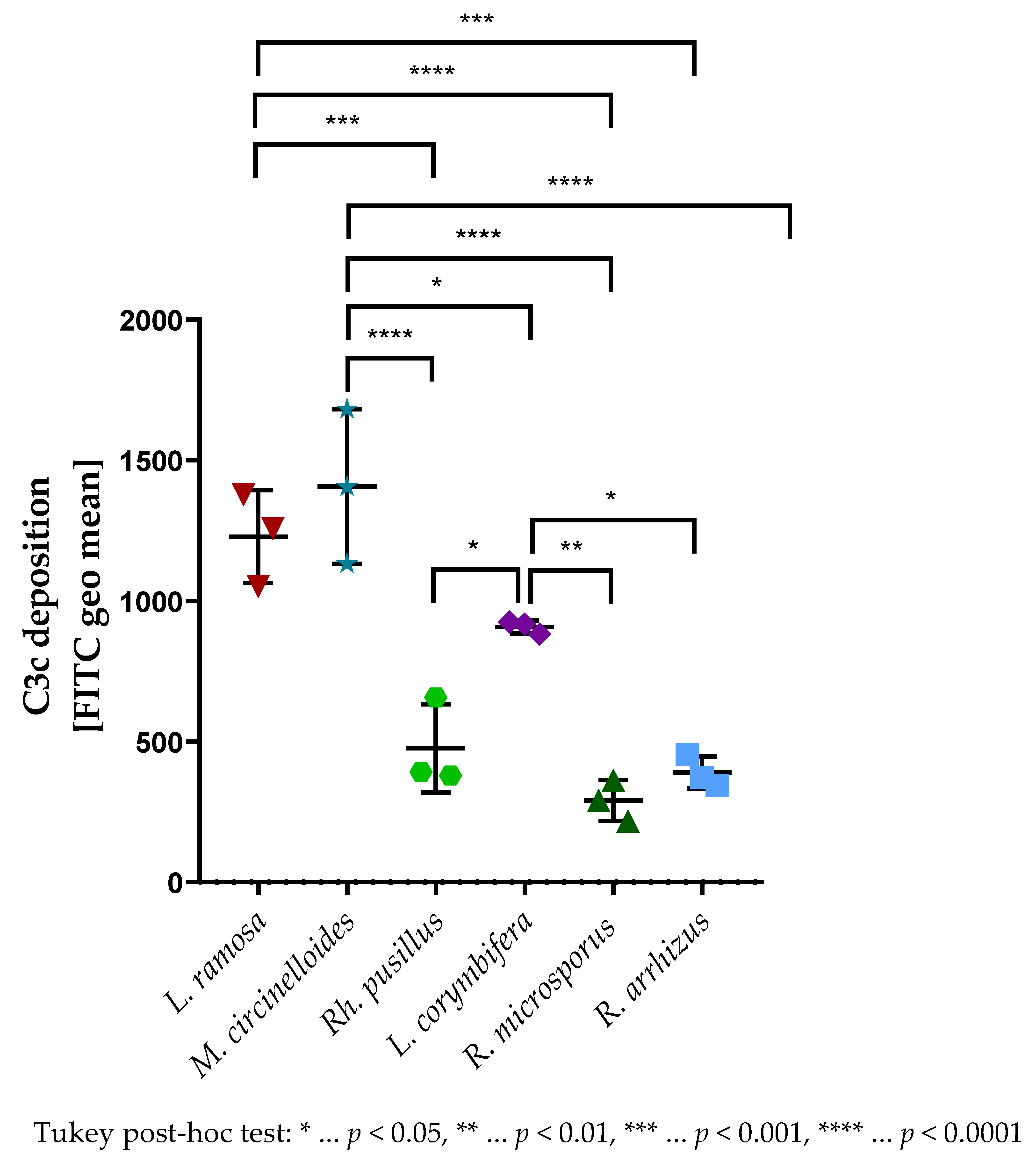
3.2. The Virulence of Selected Mucormycetes Negatively Correlated with the C3c Deposition on Spores
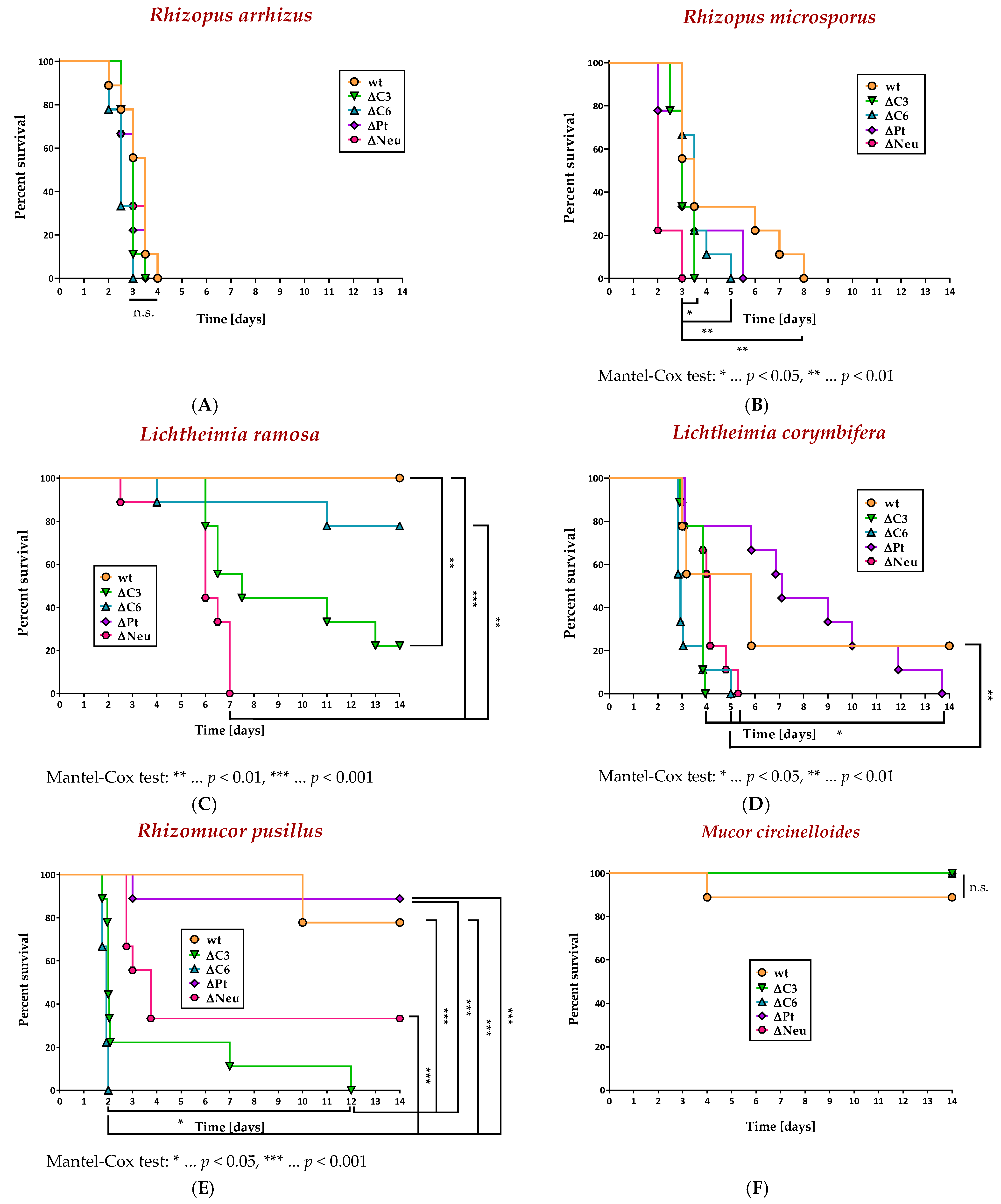
3.3. Absence of Neutrophils, C3, or C6, but Not Platelets, Increased the Lethality in a Murine Model of Disseminated Mucormycosis
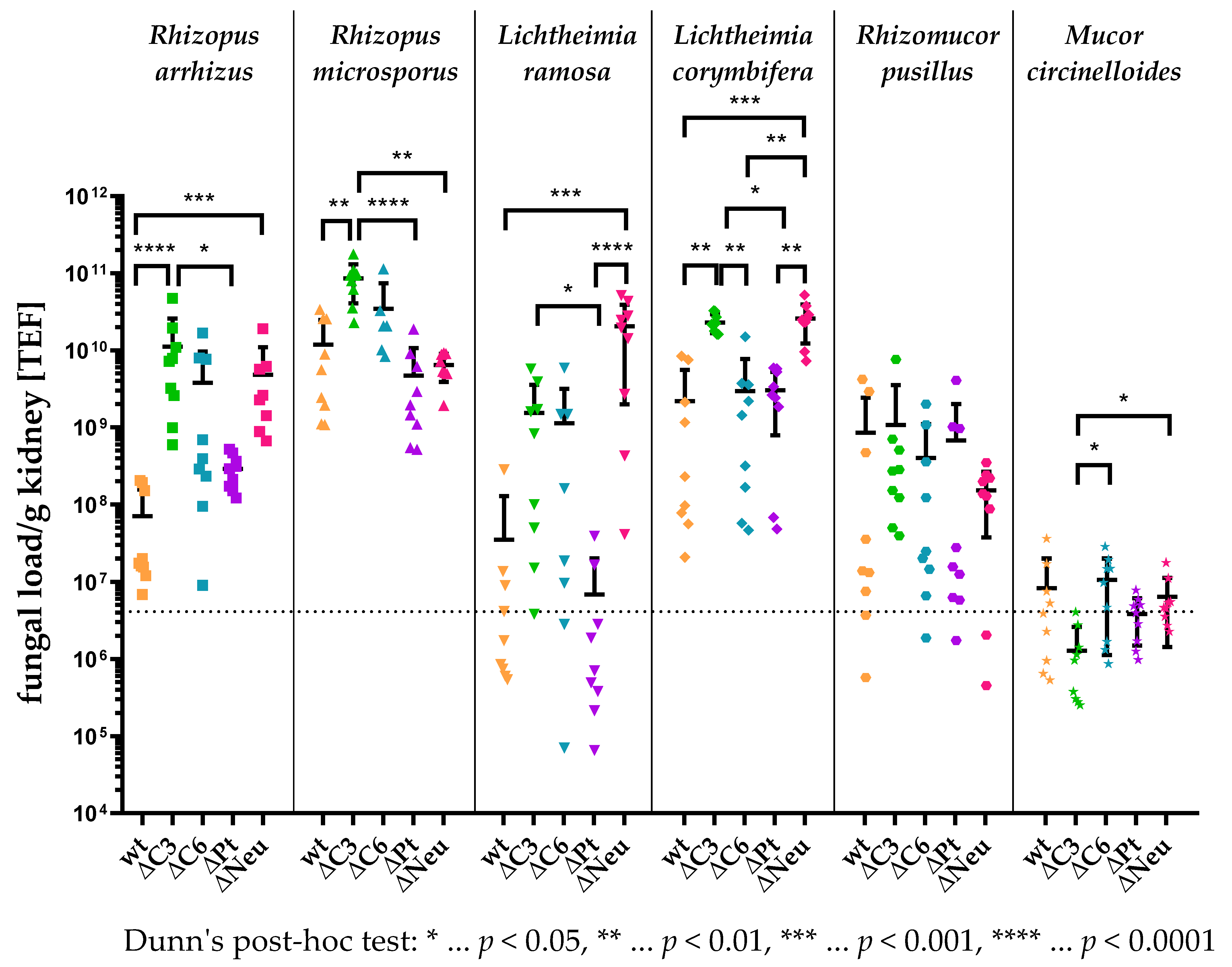
3.4. High Kidney Fungal Load in Complement-Deficient and Neutropenic Animals
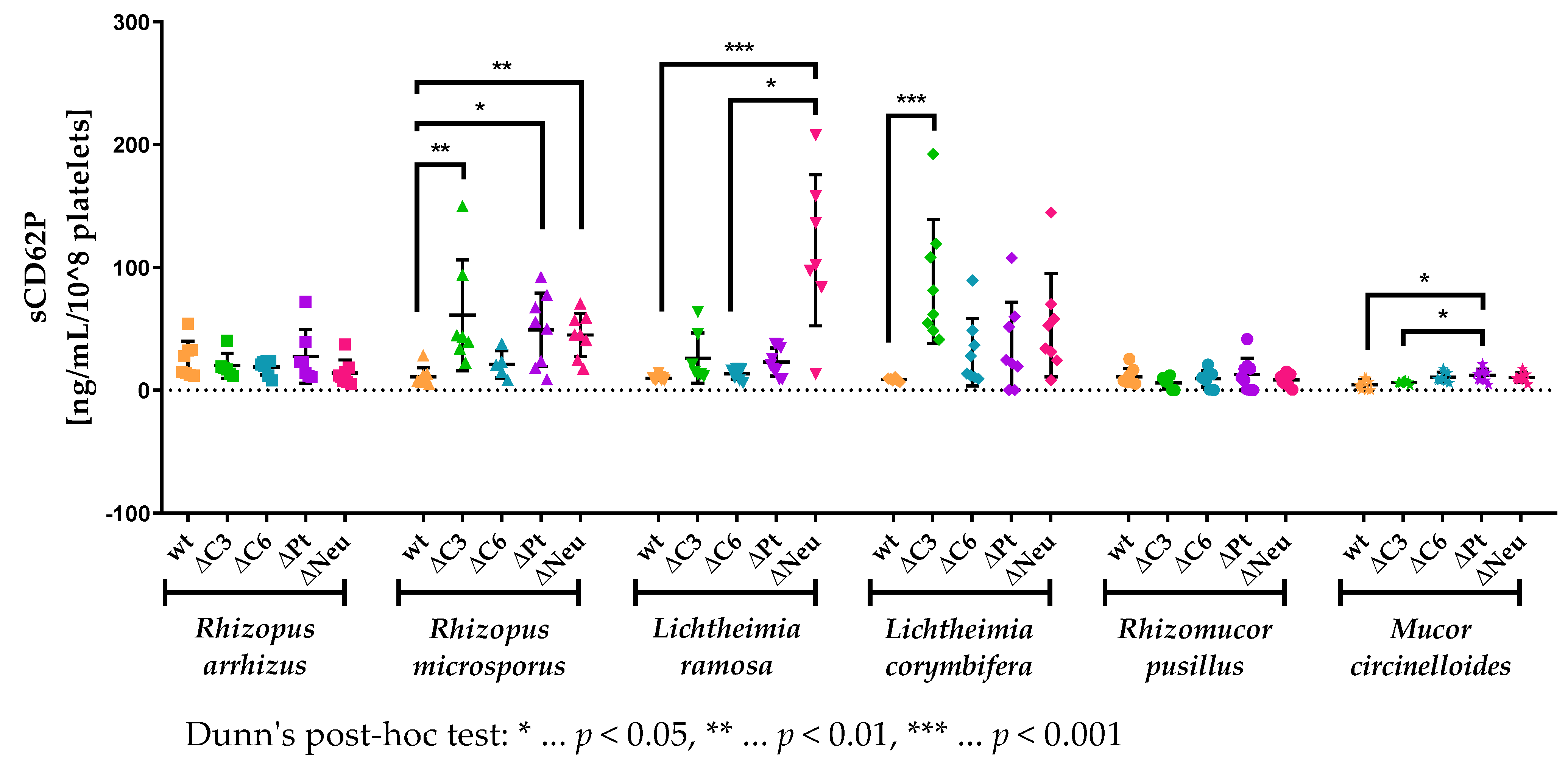
3.5. sCD62P Levels Are Increased in Neutropenic and Complement-Deficient Animals
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Binder, U.; Maurer, E.; Lass-Florl, C. Mucormycosis—From the pathogens to the disease. Clin. Microbiol. Infect. 2014, 20 (Suppl. S6), 60–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walther, G.; Wagner, L.; Kurzai, O. Outbreaks of Mucorales and the Species Involved. Mycopathologia 2020, 185, 765–781. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Vidal, C.; Salavert Lleti, M. Immunopathogenesis of invasive mould infections. Rev. Iberoam. Micol. 2014, 31, 219–228. [Google Scholar] [CrossRef]
- Skiada, A.; Pagano, L.; Groll, A.; Zimmerli, S.; Dupont, B.; Lagrou, K.; Lass-Florl, C.; Bouza, E.; Klimko, N.; Gaustad, P.; et al. Zygomycosis in Europe: Analysis of 230 cases accrued by the registry of the European Confederation of Medical Mycology (ECMM) Working Group on Zygomycosis between 2005 and 2007. Clin. Microbiol. Infect. 2011, 17, 1859–1867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakash, H.; Chakrabarti, A. Epidemiology of Mucormycosis in India. Microorganisms 2021, 9, 523. [Google Scholar] [CrossRef]
- Spellberg, B.; Edwards, J., Jr.; Ibrahim, A. Novel perspectives on mucormycosis: Pathophysiology, presentation, and management. Clin. Microbiol. Rev. 2005, 18, 556–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldin, C.; Ibrahim, A.S. Molecular mechanisms of mucormycosis-The bitter and the sweet. PLoS Pathog. 2017, 13, e1006408. [Google Scholar] [CrossRef] [Green Version]
- John, T.M.; Jacob, C.N.; Kontoyiannis, D.P. When Uncontrolled Diabetes Mellitus and Severe COVID-19 Converge: The Perfect Storm for Mucormycosis. J. Fungi 2021, 7, 298. [Google Scholar] [CrossRef] [PubMed]
- Skiada, A.; Lass-Floerl, C.; Klimko, N.; Ibrahim, A.; Roilides, E.; Petrikkos, G. Challenges in the diagnosis and treatment of mucormycosis. Med. Mycol. 2018, 56, 93–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caramalho, R.; Tyndall, J.D.A.; Monk, B.C.; Larentis, T.; Lass-Florl, C.; Lackner, M. Intrinsic short-tailed azole resistance in mucormycetes is due to an evolutionary conserved aminoacid substitution of the lanosterol 14alpha-demethylase. Sci. Rep. 2017, 7, 15898. [Google Scholar] [CrossRef]
- Morin-Sardin, S.; Nodet, P.; Coton, E.; Jany, J.L. Mucor: A Janus-faced fungal genus with human health impact and industrial applications. Fungal. Biology. Rev. 2017, 31, 12–32. [Google Scholar] [CrossRef]
- Barnum, S.R. Complement: A primer for the coming therapeutic revolution. Pharmacol. Ther. 2017, 172, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Kopp, A.; Hebecker, M.; Svobodova, E.; Jozsi, M. Factor h: A complement regulator in health and disease, and a mediator of cellular interactions. Biomolecules 2012, 2, 46–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambach, G.; Wurzner, R.; Speth, C. Complement: An efficient sword of innate immunity. Contrib. Microbiol. 2008, 15, 78–100. [Google Scholar] [CrossRef]
- Koupenova, M.; Livada, A.C.; Morrell, C.N. Platelet and Megakaryocyte Roles in Innate and Adaptive Immunity. Circ. Res. 2022, 130, 288–308. [Google Scholar] [CrossRef]
- Eriksson, O.; Mohlin, C.; Nilsson, B.; Ekdahl, K.N. The Human Platelet as an Innate Immune Cell: Interactions Between Activated Platelets and the Complement System. Front. Immunol. 2019, 10, 1590. [Google Scholar] [CrossRef] [Green Version]
- Broadley, S.P.; Plaumann, A.; Coletti, R.; Lehmann, C.; Wanisch, A.; Seidlmeier, A.; Esser, K.; Luo, S.; Ramer, P.C.; Massberg, S.; et al. Dual-Track Clearance of Circulating Bacteria Balances Rapid Restoration of Blood Sterility with Induction of Adaptive Immunity. Cell Host. Microbe. 2016, 20, 36–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauter, R.J.; Sauter, M.; Obrich, M.; Emschermann, F.N.; Nording, H.; Patzelt, J.; Wendel, H.P.; Reil, J.C.; Edlich, F.; Langer, H.F. Anaphylatoxin Receptor C3aR Contributes to Platelet Function, Thrombus Formation and In Vivo Haemostasis. Thromb. Haemost. 2019, 119, 179–182. [Google Scholar] [CrossRef]
- Nording, H.; Baron, L.; Haberthur, D.; Emschermann, F.; Mezger, M.; Sauter, M.; Sauter, R.; Patzelt, J.; Knoepp, K.; Nording, A.; et al. The C5a/C5a receptor 1 axis controls tissue neovascularization through CXCL4 release from platelets. Nat. Commun. 2021, 12, 3352. [Google Scholar] [CrossRef]
- Wiedmer, T.; Sims, P.J. Participation of protein kinases in complement C5b-9-induced shedding of platelet plasma membrane vesicles. Blood 1991, 78, 2880–2886. [Google Scholar] [CrossRef]
- Perkhofer, S.; Kainzner, B.; Kehrel, B.E.; Dierich, M.P.; Nussbaumer, W.; Lass-Florl, C. Potential antifungal effects of human platelets against zygomycetes in vitro. J. Infect. Dis. 2009, 200, 1176–1179. [Google Scholar] [CrossRef]
- Maurer, E.; Hortnagl, C.; Lackner, M.; Grassle, D.; Naschberger, V.; Moser, P.; Segal, E.; Semis, M.; Lass-Florl, C.; Binder, U. Galleria mellonella as a model system to study virulence potential of mucormycetes and evaluation of antifungal treatment. Med. Mycol. 2019, 57, 351–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wostemeyer, J. Strain-dependent variation in ribosomal DNA arrangement in Absidia glauca. Eur. J. Biochem. 1985, 146, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Bruhn, K.W.; Dekitani, K.; Nielsen, T.B.; Pantapalangkoor, P.; Spellberg, B. Ly6G-mediated depletion of neutrophils is dependent on macrophages. Results Immunol. 2016, 6, 5–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamps, L.W.; Talley, L.R.; Nolen, C.T.; Womack, J.; Scott, M.A. A Combined Hematoxylin and EosinIMethnamine Silver Stain for the Histological Diagnosis of Fungi in Tissue Sections. J. Histotechnol. 2013, 23, 341–345. [Google Scholar] [CrossRef]
- Rambach, G.; Fleischer, V.; Harpf, V.; Lackner, M.; Meinitzer, A.; Maier, H.; Engesser, J.; Lass-Florl, C.; Speth, C. Comparative immunopathogenesis in a murine model of inhalative infection with the mucormycetes Lichtheimia corymbifera and Rhizopus arrhizus. PLoS ONE 2020, 15, e0234063. [Google Scholar] [CrossRef]
- Petrikkos, G.; Skiada, A.; Drogari-Apiranthitou, M. Epidemiology of mucormycosis in Europe. Clin. Microbiol. Infect. 2014, 20 (Suppl. S6), 67–73. [Google Scholar] [CrossRef] [Green Version]
- Granja, L.F.; Pinto, L.; Almeida, C.A.; Alviano, D.S.; Da Silva, M.H.; Ejzemberg, R.; Alviano, C.S. Spores of Mucor ramosissimus, Mucor plumbeus and Mucor circinelloides and their ability to activate human complement system in vitro. Med. Mycol. 2010, 48, 278–284. [Google Scholar] [CrossRef] [Green Version]
- Wagner, L.; de Hoog, S.; Alastruey-Izquierdo, A.; Voigt, K.; Kurzai, O.; Walther, G. A Revised Species Concept for Opportunistic Mucor Species Reveals Species-Specific Antifungal Susceptibility Profiles. Antimicrob. Agents Chemother. 2019, 63, e00653-19. [Google Scholar] [CrossRef] [Green Version]
- Prakash, H.; Chakrabarti, A. Global Epidemiology of Mucormycosis. J. Fungi 2019, 5, 26. [Google Scholar] [CrossRef]
- Ma, Y.J.; Garred, P. Pentraxins in Complement Activation and Regulation. Front. Immunol. 2018, 9, 3046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garlanda, C.; Hirsch, E.; Bozza, S.; Salustri, A.; De Acetis, M.; Nota, R.; Maccagno, A.; Riva, F.; Bottazzi, B.; Peri, G.; et al. Non-redundant role of the long pentraxin PTX3 in anti-fungal innate immune response. Nature 2002, 420, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Moalli, F.; Doni, A.; Deban, L.; Zelante, T.; Zagarella, S.; Bottazzi, B.; Romani, L.; Mantovani, A.; Garlanda, C. Role of complement and Fc{gamma} receptors in the protective activity of the long pentraxin PTX3 against Aspergillus fumigatus. Blood 2010, 116, 5170–5180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Wang, G.; Griffin, L.; Banda, N.K.; Saba, L.M.; Groman, E.V.; Scheinman, R.; Moghimi, S.M.; Simberg, D. Complement opsonization of nanoparticles: Differences between humans and preclinical species. J. Control Release 2021, 338, 548–556. [Google Scholar] [CrossRef]
- Speth, C.; Hagleitner, M.; Ott, H.W.; Würzner, R.; Lass-Flörl, C.; Rambach, G. Aspergillus fumigatus Activates Thrombocytes by Secretion of Soluble Compounds. J. Infect. Dis. 2013, 207, 823–833. [Google Scholar] [CrossRef]
- Speth, C.; Rambach, G.; Lass-Florl, C. Platelet immunology in fungal infections. Thromb. Haemost. 2014, 112, 632–639. [Google Scholar] [CrossRef]
- Rambach, G.; Blum, G.; Latge, J.P.; Fontaine, T.; Heinekamp, T.; Hagleitner, M.; Jeckstrom, H.; Weigel, G.; Wurtinger, P.; Pfaller, K.; et al. Identification of Aspergillus fumigatus Surface Components That Mediate Interaction of Conidia and Hyphae with Human Platelets. J. Infect. Dis. 2015, 212, 1140–1149. [Google Scholar] [CrossRef] [Green Version]
- Tischler, B.Y.; Tosini, N.L.; Cramer, R.A.; Hohl, T.M. Platelets are critical for survival and tissue integrity during murine pulmonary Aspergillus fumigatus infection. PLoS Pathog. 2020, 16, e1008544. [Google Scholar] [CrossRef]
- Baldwin, W.M., 3rd; Kuo, H.H.; Morrell, C.N. Platelets: Versatile modifiers of innate and adaptive immune responses to transplants. Curr. Opin. Organ. Transplant. 2011, 16, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Semple, J.W.; Italiano, J.E., Jr.; Freedman, J. Platelets and the immune continuum. Nat. Rev. Immunol. 2011, 11, 264–274. [Google Scholar] [CrossRef]
- Ricklin, D.; Hajishengallis, G.; Yang, K.; Lambris, J.D. Complement: A key system for immune surveillance and homeostasis. Nat. Immunol. 2010, 11, 785–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granja, L.F.Z.; Pinto, L.; Silvestre, J.S.; Almeida, C.A.; Alviano, D.S.; Takaki, G.M.C.; Silva, M.H.D.; Ejzemberg, R.; Alviano, C.S. Spores and Yeasts of Mucor Polymorphosporus and their ability to Activate Human Complement System in vitro. Arch. Clin. Microbiol. 2016, 7, 1, https://www.itmedicalteam.pl/articles/spores-and-yeasts-of-mucor-polymorphosporus-and-their-ability-toactivate-human-complement-system-in-vitro-102469.html. [Google Scholar]
- Bossi, F.; Fischetti, F.; Pellis, V.; Bulla, R.; Ferrero, E.; Mollnes, T.E.; Regoli, D.; Tedesco, F. Platelet-activating factor and kinin-dependent vascular leakage as a novel functional activity of the soluble terminal complement complex. J. Immunol. 2004, 173, 6921–6927. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, P.; Sahoo, R.; Vaidya, A.; Chorev, M.; Halperin, J.A. Role of complement and complement regulatory proteins in the complications of diabetes. Endocr. Rev. 2015, 36, 272–288. [Google Scholar] [CrossRef] [PubMed]
- Serna, M.; Giles, J.L.; Morgan, B.P.; Bubeck, D. Structural basis of complement membrane attack complex formation. Nat. Commun. 2016, 7, 10587. [Google Scholar] [CrossRef] [Green Version]
- Frater, J.L.; Hall, G.S.; Procop, G.W. Histologic features of zygomycosis: Emphasis on perineural invasion and fungal morphology. Arch. Pathol. Lab. Med. 2001, 125, 375–378. [Google Scholar] [CrossRef]
- Melida, H.; Sain, D.; Stajich, J.E.; Bulone, V. Deciphering the uniqueness of Mucoromycotina cell walls by combining biochemical and phylogenomic approaches. Environ. Microbiol. 2015, 17, 1649–1662. [Google Scholar] [CrossRef] [Green Version]
- Hawker, L.E.; Abbott, P.M. Fine structure of vegetative hyphae of Rhizopus. J. Gen. Microbiol. 1963, 30, 401–408. [Google Scholar] [CrossRef] [Green Version]
- Bartnicki-Garcia, S.; Nelson, N.; Cota-Robles, E. Electron microscopy of spore germination and cell wall formation in Mucor rouxii. Arch. Mikrobiol. 1968, 63, 242–255. [Google Scholar] [CrossRef]
- Schmalhorst, P.S.; Krappmann, S.; Vervecken, W.; Rohde, M.; Muller, M.; Braus, G.H.; Contreras, R.; Braun, A.; Bakker, H.; Routier, F.H. Contribution of galactofuranose to the virulence of the opportunistic pathogen Aspergillus fumigatus. Eukaryot. Cell 2008, 7, 1268–1277. [Google Scholar] [CrossRef]
- Sterba, G.; Sterba, Y. Parasitic and Fungal Triggers. In Cytokine Storm Syndrome, 1st ed.; Cron, R.Q., Begrens, E.M., Eds.; Springer International Publishing: Basel, Switzerland, 2019; pp. 331–345. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harpf, V.; Rambach, G.; Parth, N.; Neurauter, M.; Fleischer, V.; Lackner, M.; Lass-Flörl, C.; Würzner, R.; Speth, C. Complement, but Not Platelets, Plays a Pivotal Role in the Outcome of Mucormycosis In Vivo. J. Fungi 2023, 9, 162. https://doi.org/10.3390/jof9020162
Harpf V, Rambach G, Parth N, Neurauter M, Fleischer V, Lackner M, Lass-Flörl C, Würzner R, Speth C. Complement, but Not Platelets, Plays a Pivotal Role in the Outcome of Mucormycosis In Vivo. Journal of Fungi. 2023; 9(2):162. https://doi.org/10.3390/jof9020162
Chicago/Turabian StyleHarpf, Verena, Günter Rambach, Nadia Parth, Magdalena Neurauter, Verena Fleischer, Michaela Lackner, Cornelia Lass-Flörl, Reinhard Würzner, and Cornelia Speth. 2023. "Complement, but Not Platelets, Plays a Pivotal Role in the Outcome of Mucormycosis In Vivo" Journal of Fungi 9, no. 2: 162. https://doi.org/10.3390/jof9020162