
Mucormycosis and COVID-19-Associated Mucormycosis: Insights of a Deadly but Neglected Mycosis


{kind=link}
Abstract
:1. Introduction
2. Mucormycosis
2.1. Risk Factors
- -
- -
- -
- -
- -
- -
- -
2.2. Pathogenesis
2.3. Immune Response during Mucormycosis
2.4. Treatment
3. COVID-19-Associated Mucormycosis
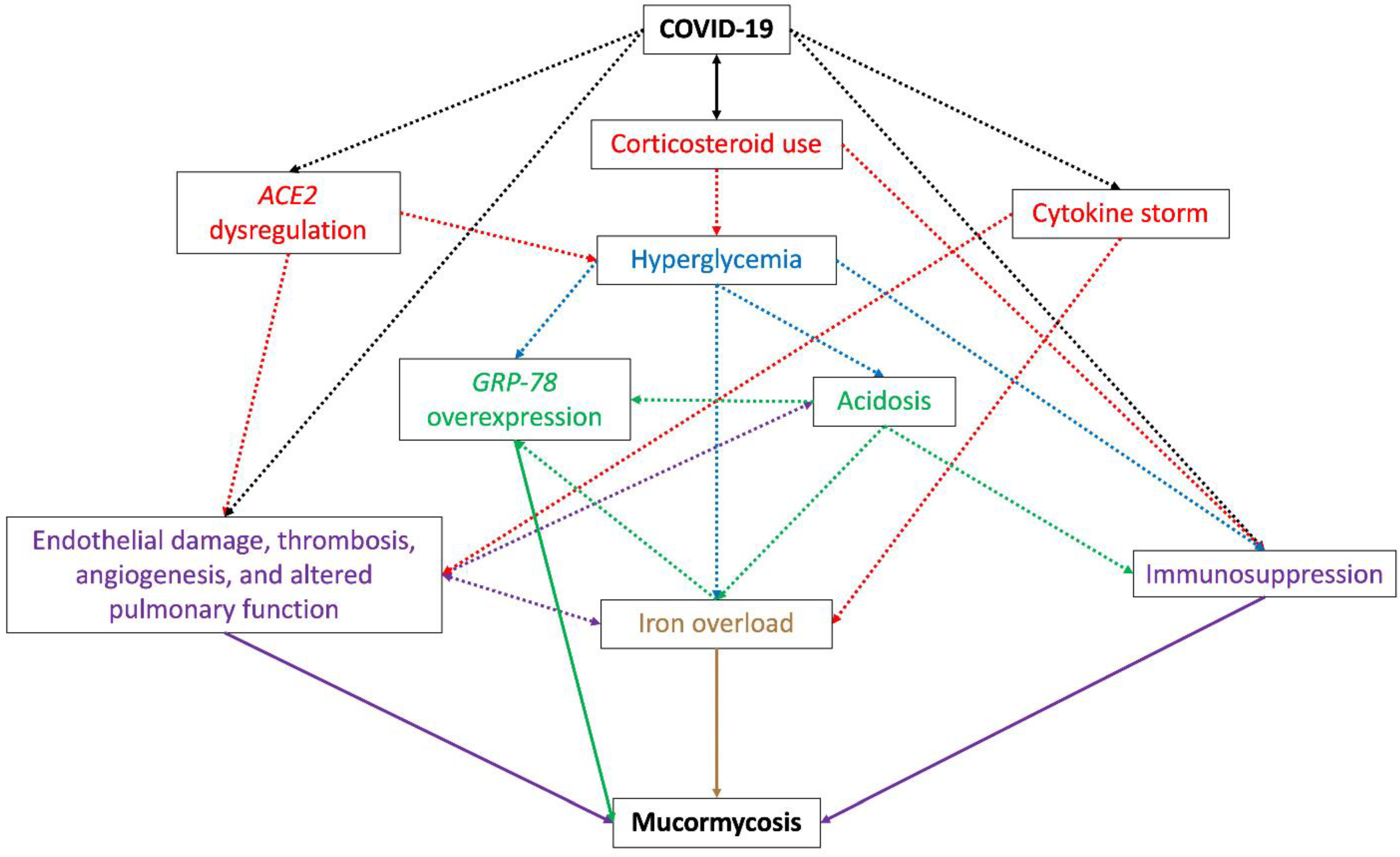
- Hyperglycemia, caused by the use of corticosteroids to treat COVID-19 [143] and dysregulation of the host receptor ACE2 (angiotensin-converting enzyme 2) observed during the viral infection [144,145] (explained in vii). Pre-existing diabetes is the main risk factor in most CAM cases [12] and is also related to an increase in the severity of SARS-CoV-2 infection [146];
- Free iron availability, caused by hyperglycemia, the COVID-19 cytokine storm, or by acidosis. Hyperglycemia and diabetic ketoacidosis cause dissociation of iron from ferritin and lactoferrin, thus elevating free iron concentrations [152], and some cytokines, mainly IL-6, stimulate ferritin synthesis and decrease iron export, increasing intracellular iron storage [153,154,155] and causing tissue damage and thus the release of iron into circulation [156]. Therapeutic intervention with lactoferrin has been suggested to revert iron availability [157,158,159];
- Overexpression of endothelial cells GRP-78, caused by hyperglycemia, acidosis, and iron availability, which enables angioinvasion, hematogenous dissemination, and tissue necrosis [58];
- Dysregulation of ACE2 by SARS-CoV-2. By being present in many organs and tissues, the alteration of the receptor ACE2 expression causes a suitable environment for the development of mucormycosis. During COVID-19, a downregulation of ACE2 has been observed in the lungs [161], which causes inflammation, leukocytes exudation, and altered pulmonary function and therefore poor oxygenation [144]. Additionally, the effect of the virus on ACE2 in the pancreatic beta cells causes hyperglycemia, while dysregulation of the receptor in the vascular endothelium causes endothelial damage and vascular thrombosis, leading to vascular endothelial injury and venous stasis, causing an increase in serum iron due to hemolysis [144,145];
3.1. Clinical Manifestations
3.2. CAM Diagnosis and Treatment
3.3. CAM Prevention and Outcome
- -
- During COVID-19 management, hyperglycemia should be strictly controlled;
- -
- Corticosteroid therapy should only be used in patients with severe SARS-CoV-2 infection, for short periods and at low doses, just like other drugs that target the immune response, such as tocilizumab [210];
- -
- The patient’s exposition to Mucorales should be limited as much as possible by using face masks and avoiding being outdoors;
- -
- Mass vaccination against COVID-19 can also help to reduce the chance to develop severe secondary complications [210].
4. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rodriguez-Morales, A.J.; Bonilla-Aldana, D.K.; Balbin-Ramon, G.J.; Rabaan, A.A.; Sah, R.; Paniz-Mondolfi, A.; Pagliano, P.; Esposito, S. History is repeating itself: Probable zoonotic spillover as the cause of the 2019 novel Coronavirus epidemic. Le Infez. Med. 2020, 28, 3–5. [Google Scholar]
- Krishnan, A.; Hamilton, J.P.; Alqahtani, S.A.; Woreta, T.A. COVID-19: An overview and a clinical update. World J. Clin. Cases 2021, 9, 8–23. [Google Scholar] [CrossRef] [PubMed]
- Gralinski, L.E.; Menachery, V.D. Return of the Coronavirus: 2019-nCoV. Viruses 2020, 12, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdoli, A.; Falahi, S.; Kenarkoohi, A. COVID-19-associated opportunistic infections: A snapshot on the current reports. Clin. Exp. Med. 2021, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Agarwal, R.; Rudramurthy, S.M.; Shevkani, M.; Xess, I.; Sharma, R.; Savio, J.; Sethuraman, N.; Madan, S.; Shastri, P.; et al. Multicenter Epidemiologic Study of Coronavirus Disease–Associated Mucormycosis, India. Emerg. Infect. Dis. 2021, 27, 2349–2359. [Google Scholar] [CrossRef]
- Song, G.; Liang, G.; Liu, W. Fungal Co-infections Associated with Global COVID-19 Pandemic: A Clinical and Diagnostic Perspective from China. Mycopathologia 2020, 185, 599–606. [Google Scholar] [CrossRef]
- Bhatt, K.; Agolli, A.; Patel, M.H.; Garimella, R.; Devi, M.; Garcia, E.; Amin, H.; Domingue, C.; Del Castillo, R.G.; Sanchez-Gonzalez, M. High mortality co-infections of COVID-19 patients: Mucormycosis and other fungal infections. Discoveries 2021, 9, e126. [Google Scholar] [CrossRef]
- Rodriguez-Morales, A.J.; Sah, R.; Millan-Oñate, J.; Gonzalez, A.; Montenegro-Idrogo, J.J.; Scherger, S.; Franco-Paredes, C.; Henao-Martínez, A.F. COVID-19 associated mucormycosis: The urgent need to reconsider the indiscriminate use of immunosuppressive drugs. Ther. Adv. Infect. Dis. 2021, 8, 20499361211027065. [Google Scholar] [CrossRef]
- Arora, S.; Hemmige, V.S.; Mandke, C.; Chansoria, M.; Rawat, S.K.; Dravid, A.; Sethi, Y.; Medikeri, G.; Jariwala, S.P.; Puius, Y.A. Online Registry of COVID-19–Associated Mucormycosis Cases, India, 2021. Emerg. Infect. Dis. 2021, 27, 2963–2965. [Google Scholar] [CrossRef]
- Bonifaz, A.; Tirado-Sánchez, A.; Hernández-Medel, M.L.; Araiza, J.; Kassack, J.J.; del Angel-Arenas, T.; Moisés-Hernández, J.F.; Paredes-Farrera, F.; Gómez-Apo, E.; Treviño-Rangel, R.D.J.; et al. Mucormycosis at a tertiary-care center in Mexico. A 35-year retrospective study of 214 cases. Mycoses 2020, 64, 372–380. [Google Scholar] [CrossRef]
- Bhatia, M. The rise of mucormycosis in COVID-19 patients in India. Expert Rev. Anti-infective Ther. 2021, 20, 137–138. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Singh, R.; Joshi, S.R.; Misra, A. Mucormycosis in COVID-19: A systematic review of cases reported worldwide and in India. Diabetes Metab. Syndr. Clin. Res. Rev. 2021, 15, 102146. [Google Scholar] [CrossRef] [PubMed]
- Junior, E.S.D.M.; Dos Santos, M.E.L.; Ribeiro, I.B.; Luz, G.D.O.; Baba, E.R.; Hirsch, B.S.; Funari, M.P.; De Moura, E.G.H. Rare and Fatal Gastrointestinal Mucormycosis (Zygomycosis) in a COVID-19 Patient: A Case Report. Clin. Endosc. 2020, 53, 746–749. [Google Scholar] [CrossRef]
- Waizel-Haiat, S.; Guerrero-Paz, J.A.; Sanchez-Hurtado, L.; Calleja-Alarcon, S.; Romero-Gutierrez, L. A Case of Fatal Rhino-Orbital Mucormycosis Associated With New Onset Diabetic Ketoacidosis and COVID-19. Cureus 2021, 13, e13163. [Google Scholar] [CrossRef] [PubMed]
- Khatri, A.; Chang, K.-M.; Berlinrut, I.; Wallach, F. Mucormycosis after Coronavirus disease 2019 infection in a heart transplant recipient–case report and review of literature. J. Mycol. Med. 2021, 31, 101125. [Google Scholar] [CrossRef] [PubMed]
- Placik, D.A.; Taylor, W.L.; Wnuk, N.M. Bronchopleural fistula development in the setting of novel therapies for acute respiratory distress syndrome in SARS-CoV-2 pneumonia. Radiol. Case Rep. 2020, 15, 2378–2381. [Google Scholar] [CrossRef]
- Werthman-Ehrenreich, A. Mucormycosis with orbital compartment syndrome in a patient with COVID-19. Am. J. Emerg. Med. 2020, 42, 264.e5–264.e8. [Google Scholar] [CrossRef]
- Mekonnen, Z.K.; Ashraf, D.C.; Jankowski, T.; Grob, S.R.; Vagefi, M.R.; Kersten, R.C.; Simko, J.P.; Winn, B.J. Acute Invasive Rhino-Orbital Mucormycosis in a Patient With COVID-19-Associated Acute Respiratory Distress Syndrome. Ophthalmic Plast. Reconstr. Surg. 2020, 37, e40–e80. [Google Scholar] [CrossRef]
- Kanwar, A.; Jordan, A.; Olewiler, S.; Wehberg, K.; Cortes, M.; Jackson, B. A Fatal Case of Rhizopus azygosporus Pneumonia Following COVID-19. J. Fungi 2021, 7, 174. [Google Scholar] [CrossRef]
- Dallalzadeh, L.O.; Ozzello, D.J.; Liu, C.Y.; Kikkawa, D.O.; Korn, B.S. Secondary infection with rhino-orbital cerebral mucormycosis associated with COVID-19. Orbit 2021, 2021, 1–4. [Google Scholar] [CrossRef]
- Johnson, A.K.; Ghazarian, Z.; Cendrowski, K.D.; Persichino, J.G. Pulmonary aspergillosis and mucormycosis in a patient with COVID-19. Med. Mycol. Case Rep. 2021, 32, 64–67. [Google Scholar] [CrossRef] [PubMed]
- Hanley, B.; Naresh, K.; Roufosse, C.; Nicholson, A.G.; Weir, J.; Cooke, G.S.; Thursz, M.; Manousou, P.; Corbett, R.; Goldin, R.; et al. Histopathological findings and viral tropism in UK patients with severe fatal COVID-19: A post-mortem study. Lancet Microbe 2020, 1, e245–e253. [Google Scholar] [CrossRef]
- Pasero, D.; Sanna, S.; Liperi, C.; Piredda, D.; Branca, G.P.; Casadio, L.; Simeo, R.; Buselli, A.; Rizzo, D.; Bussu, F.; et al. A challenging complication following SARS-CoV-2 infection: A case of pulmonary mucormycosis. Infection 2020, 49, 1055–1060. [Google Scholar] [CrossRef]
- Bellanger, A.-P.; Navellou, J.-C.; Lepiller, Q.; Brion, A.; Brunel, A.-S.; Millon, L.; Berceanu, A. Mixed mold infection with Aspergillus fumigatus and Rhizopus microsporus in a severe acute respiratory syndrome Coronavirus 2 (SARS-CoV-2) patient. Infect. Dis. Now 2021, 51, 633–635. [Google Scholar] [CrossRef] [PubMed]
- Zurl, C.; Hoenigl, M.; Schulz, E.; Hatzl, S.; Gorkiewicz, G.; Krause, R.; Eller, P.; Prattes, J. Autopsy Proven Pulmonary Mucormycosis Due to Rhizopus microsporus in a Critically Ill COVID-19 Patient with Underlying Hematological Malignancy. J. Fungi 2021, 7, 88. [Google Scholar] [CrossRef]
- Ahmadikia, K.; Hashemi, S.J.; Khodavaisy, S.; Getso, M.I.; Alijani, N.; Badali, H.; Mirhendi, H.; Salehi, M.; Tabari, A.; Ardehali, M.M.; et al. The double-edged sword of systemic corticosteroid therapy in viral pneumonia: A case report and comparative review of influenza-associated mucormycosis versus COVID-19 associated mucormycosis. Mycoses 2021, 64, 798–808. [Google Scholar] [CrossRef]
- Karimi-Galougahi, M.; Arastou, S.; Haseli, S. Fulminant mucormycosis complicating coronavirus disease 2019 (COVID-19). Int. Forum Allergy Rhinol. 2021, 11, 1029–1030. [Google Scholar] [CrossRef]
- Veisi, A.; Bagheri, A.; Eshaghi, M.; Rikhtehgar, M.H.; Kanavi, M.R.; Farjad, R. Rhino-orbital mucormycosis during steroid therapy in COVID-19 patients: A case report. Eur. J. Ophthalmol. 2021, 2021, 1–6. [Google Scholar] [CrossRef]
- Baldin, C.; Ibrahim, A.S. Molecular mechanisms of mucormycosis—The bitter and the sweet. PLOS Pathog. 2017, 13, e1006408. [Google Scholar] [CrossRef] [Green Version]
- Gleissner, B.; Schilling, A.; Anagnostopolous, I.; Siehl, I.; Thiel, E. Improved Outcome of Zygomycosis in Patients with Hematological Diseases? Leuk. Lymphoma 2004, 45, 1351–1360. [Google Scholar] [CrossRef]
- Dignani, M.C. Epidemiology of invasive fungal diseases on the basis of autopsy reports. F1000Prime Rep. 2014, 6, 81. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, A.; Singh, R. The emerging epidemiology of mould infections in developing countries. Curr. Opin. Infect. Dis. 2011, 24, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Skiada, A.; Pagano, L.; Groll, A.; Zimmerli, S.; Dupont, B.; Lagrou, K.; Lass-Florl, C.; Bouza, E.; Klimko, N.; Gaustad, P.; et al. Zygomycosis in Europe: Analysis of 230 cases accrued by the registry of the European Confederation of Medical Mycology (ECMM) Working Group on Zygomycosis between 2005 and 2007. Clin. Microbiol. Infect. 2011, 17, 1859–1867. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, A.S.; Spellberg, B.; Walsh, T.J.; Kontoyiannis, D.P. Pathogenesis of Mucormycosis. Clin. Infect. Dis. 2012, 54, S16–S22. [Google Scholar] [CrossRef]
- Park, H.-R.; Voigt, K. Interaction of Zygomycetes with innate immune cells reconsidered with respect to ecology, morphology, evolution and infection biology: A mini-review. Mycoses 2014, 57, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Challa, S. Mucormycosis: Pathogenesis and Pathology. Curr. Fungal Infect. Rep. 2019, 13, 11–20. [Google Scholar] [CrossRef]
- Spellberg, B.; Edwards, J., Jr.; Ibrahim, A. Novel Perspectives on Mucormycosis: Pathophysiology, Presentation, and Management. Clin. Microbiol. Rev. 2005, 18, 556–569. [Google Scholar] [CrossRef] [Green Version]
- Hassan, M.A.; Voigt, K. Pathogenicity patterns of mucormycosis: Epidemiology, interaction with immune cells and virulence factors. Med Mycol. 2019, 57, S245–S256. [Google Scholar] [CrossRef] [Green Version]
- Kwon-Chung, K.J. Taxonomy of Fungi Causing Mucormycosis and Entomophthoramycosis (Zygomycosis) and Nomenclature of the Disease: Molecular Mycologic Perspectives. Clin. Infect. Dis. 2012, 54, S8–S15. [Google Scholar] [CrossRef] [Green Version]
- Petrikkos, G.; Skiada, A.; Lortholary, O.; Roilides, E.; Walsh, T.J.; Kontoyiannis, D.P. Epidemiology and Clinical Manifestations of Mucormycosis. Clin. Infect. Dis. 2012, 54 (Suppl. 1), S23–S34. [Google Scholar] [CrossRef]
- Farmakiotis, D.; Kontoyiannis, D.P. Mucormycoses. Infect. Dis. Clin. North Am. 2016, 30, 143–163. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.-Y.; Singh, N. Mucormycosis: Its contemporary face and management strategies. Lancet Infect. Dis. 2011, 11, 301–311. [Google Scholar] [CrossRef]
- Richardson, M. The ecology of the Zygomycetes and its impact on environmental exposure. Clin. Microbiol. Infect. 2009, 15, 2–9. [Google Scholar] [CrossRef]
- Idnurm, A. Sex Determination in the First-Described Sexual Fungus. Eukaryot. Cell 2011, 10, 1485–1491. [Google Scholar] [CrossRef] [Green Version]
- Katragkou, A.; Walsh, T.J.; Roilides, E. Why is mucormycosis more difficult to cure than more common mycoses? Clin. Microbiol. Infect. 2014, 20, 74–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spellberg, B. Mucormycosis pathogenesis: Beyond Rhizopus. Virulence 2017, 8, 1481–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze, B.; Rambach, G.; Schwartze, V.U.; Voigt, K.; Schubert, K.; Speth, C.; Jacobsen, I.D. Ketoacidosis alone does not predispose to mucormycosis by Lichtheimia in a murine pulmonary infection model. Virulence 2017, 8, 1657–1667. [Google Scholar] [CrossRef] [Green Version]
- Chandra, S.; Sharma, S.; Vats, R.; Pandey, S. Isolated cerebral mucormycosis masquerading as a tumor in an immunocompetent patient. Autops. Case Rep. 2020, 11, e2020233. [Google Scholar] [CrossRef]
- Roden, M.M.; Zaoutis, T.E.; Buchanan, W.L.; Knudsen, T.A.; Sarkisova, T.A.; Schaufele, R.L.; Sein, M.; Sein, T.; Chiou, C.C.; Chu, J.H.; et al. Epidemiology and Outcome of Zygomycosis: A Review of 929 Reported Cases. Clin. Infect. Dis. 2005, 41, 634–653. [Google Scholar] [CrossRef] [Green Version]
- Petrikkos, G.; Tsioutis, C. Recent Advances in the Pathogenesis of Mucormycoses. Clin. Ther. 2018, 40, 894–902. [Google Scholar] [CrossRef] [Green Version]
- Artis, W.M.; Fountain, J.A.; Delcher, H.K.; Jones, H.E. A Mechanism of Susceptibility to Mucormycosis in Diabetic Ketoacidosis Transferrin and Iron Availability. Diabetes 1982, 31, 1109–1114. [Google Scholar] [CrossRef] [PubMed]
- Lamaris, G.A.; Ben-Ami, R.; Lewis, R.; Chamilos, G.; Samonis, G.; Kontoyiannis, D.P. Increased Virulence of Zygomycetes Organisms Following Exposure to Voriconazole: A Study Involving Fly and Murine Models of Zygomycosis. J. Infect. Dis. 2009, 199, 1399–1406. [Google Scholar] [CrossRef] [PubMed]
- Benedict, K.; Park, B.J. Invasive Fungal Infections after Natural Disasters. Emerg. Infect. Dis. 2014, 20, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Sugar, A.M. Mucormycosis. Clin. Infect. Dis. 1992, 14, S126–S129. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.E.; Kontoyiannis, D.P. Epidemiology and treatment of mucormycosis. Futur. Microbiol. 2013, 8, 1163–1175. [Google Scholar] [CrossRef]
- Ma, L.-J.; Ibrahim, A.S.; Skory, C.; Grabherr, M.G.; Burger, G.; Butler, M.; Elias, M.; Idnurm, A.; Lang, B.F.; Sone, T.; et al. Genomic Analysis of the Basal Lineage Fungus Rhizopus oryzae Reveals a Whole-Genome Duplication. PLoS Genet. 2009, 5, e1000549. [Google Scholar] [CrossRef]
- Ibrahim, A.S.; Gebremariam, T.; Lin, L.; Luo, G.; Husseiny, M.I.; Skory, C.D.; Fu, Y.; French, S.W.; Edwards, J.E., Jr.; Spellberg, B. The high affinity iron permease is a key virulence factor required for Rhizopus oryzae pathogenesis. Mol. Microbiol. 2010, 77, 587–604. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Spellberg, B.; Phan, Q.T.; Fu, Y.; Fu, Y.; Lee, A.; Edwards, J.E.; Filler, S.G.; Ibrahim, A.S. The endothelial cell receptor GRP78 is required for mucormycosis pathogenesis in diabetic mice. J. Clin. Investig. 2010, 120, 1914–1924. [Google Scholar] [CrossRef] [Green Version]
- Binder, U.; Maurer, E.; Lass-Flörl, C. Mucormycosis–from the pathogens to the disease. Clin. Microbiol. Infect. 2014, 20, 60–66. [Google Scholar] [CrossRef] [Green Version]
- Tissot, F.; Agrawal, S.; Pagano, L.; Petrikkos, G.; Groll, A.H.; Skiada, A.; Lass-Flörl, C.; Calandra, T.; Viscoli, C.; Herbrecht, R. ECIL-6 guidelines for the treatment of invasive candidiasis, aspergillosis and mucormycosis in leukemia and hematopoietic stem cell transplant patients. Haematologica 2016, 102, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.L.; Hesseltine, C.W. Studies on the extracellular proteolytic enzymes of rhizopus oligosporus. Can. J. Microbiol. 1965, 11, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Staib, F. Extracellular proteolysis by Mucoraceae in serum-albumin agar tested by the agar block method. Mycoses 2009, 34, 231–233. [Google Scholar] [CrossRef]
- Schoen, C.; Reichard, U.; Monod, M.; Kratzin, H.D.; Rüchel, R. Molecular cloning of an extracellular aspartic proteinase fromRhizopus microsporusand evidence for its expression during infection. Med Mycol. 2002, 40, 61–71. [Google Scholar] [CrossRef] [Green Version]
- Spreer, A.; Rüchel, R.; Reichard, U. Characterization of an extracellular subtilisin protease ofRhizopus microsporusand evidence for its expression during invasive rhinoorbital mycosis. Med Mycol. 2006, 44, 723–731. [Google Scholar] [CrossRef] [Green Version]
- Ribes, J.A.; Vanover-Sams, C.L.; Baker, D.J. Zygomycetes in Human Disease. Clin. Microbiol. Rev. 2000, 13, 236–301. [Google Scholar] [CrossRef] [PubMed]
- Gebremariam, T.; Liu, M.; Luo, G.; Bruno, V.; Phan, Q.T.; Waring, A.J.; Edwards, J.E.; Filler, S.G.; Yeaman, M.R.; Ibrahim, A.S. CotH3 mediates fungal invasion of host cells during mucormycosis. J. Clin. Investig. 2013, 124, 237–250. [Google Scholar] [CrossRef] [Green Version]
- Chibucos, M.C.; Soliman, S.; Gebremariam, T.; Lee, H.; Daugherty, S.; Orvis, J.; Shetty, A.; Crabtree, J.; Hazen, T.H.; Etienne, K.A.; et al. An integrated genomic and transcriptomic survey of mucormycosis-causing fungi. Nat. Commun. 2016, 7, 12218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantinieaux, B.; Janssens, A.; Boelaert, J.; Lejeune, M.; Vermylen, C.; Kerrels, V.; Cornu, G.; Winand, J.; Fondu, P. Ferritin-associated iron induces neutrophil dysfunction in hemosiderosis. J. Lab. Clin. Med. 1999, 133, 353–361. [Google Scholar] [CrossRef]
- Shirazi, F.; Kontoyiannis, D.P.; Ibrahim, A.S. Iron starvation induces apoptosis in rhizopus oryzae in vitro. Virulence 2015, 6, 121–126. [Google Scholar] [CrossRef] [Green Version]
- Gebremariam, T.; Lin, L.; Liu, M.; Kontoyiannis, D.P.; French, S.; Edwards, J.E.; Filler, S.G.; Ibrahim, A.S. Bicarbonate correction of ketoacidosis alters host-pathogen interactions and alleviates mucormycosis. J. Clin. Investig. 2016, 126, 2280–2294. [Google Scholar] [CrossRef] [Green Version]
- Thieken, A.; Winkelmann, G.; Winkelmann, G. Rhizoferrin: A complexone type siderophore of the mocorales and entomophthorales (Zygomycetes). FEMS Microbiol. Lett. 1992, 94, 37–41. [Google Scholar] [CrossRef]
- De Locht, M.; Boelaert, J.R.; Schneider, Y.-J. Iron uptake from ferrioxamine and from ferrirhizoferrin by germinating spores of rhizopus microsporus. Biochem. Pharmacol. 1994, 47, 1843–1850. [Google Scholar] [CrossRef]
- Boelaert, J.R.; De Locht, M.; Van Cutsem, J.; Kerrels, V.; Cantinieaux, B.; Verdonck, A.; Van Landuyt, H.W.; Schneider, Y.-J. Mucormycosis during deferoxamine therapy is a siderophore-mediated infection. In vitro and in vivo animal studies. J. Clin. Investig. 1993, 91, 1979–1986. [Google Scholar] [CrossRef]
- Liu, M.; Lin, L.; Gebremariam, T.; Luo, G.; Skory, C.D.; French, S.W.; Chou, T.-F.; Edwards, J.E., Jr.; Ibrahim, A.S. Fob1 and Fob2 Proteins Are Virulence Determinants of Rhizopus oryzae via Facilitating Iron Uptake from Ferrioxamine. PLoS Pathog. 2015, 11, e1004842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roilides, E.; Kontoyiannis, D.P.; Walsh, T.J. Host Defenses Against Zygomycetes. Clin. Infect. Dis. 2012, 54, S61–S66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamilos, G.; Lewis, R.E.; Lamaris, G.; Walsh, T.J.; Kontoyiannis, D.P. Zygomycetes Hyphae Trigger an Early, Robust Proinflammatory Response in Human Polymorphonuclear Neutrophils through Toll-Like Receptor 2 Induction but Display Relative Resistance to Oxidative Damage. Antimicrob. Agents Chemother. 2008, 52, 722–724. [Google Scholar] [CrossRef] [Green Version]
- .Ibrahim, A.S.; Voelz, K. The mucormycete–host interface. Curr. Opin. Microbiol. 2017, 40, 40–45. [Google Scholar] [CrossRef]
- Andrianaki, A.M.; Kyrmizi, I.; Thanopoulou, K.; Baldin, C.; Drakos, E.; Soliman, S.S.M.; Shetty, A.C.; McCracken, C.; Akoumianaki, T.; Stylianou, K.; et al. Iron restriction inside macrophages regulates pulmonary host defense against Rhizopus species. Nat. Commun. 2018, 9, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Kraibooj, K.; Park, H.-R.; Dahse, H.-M.; Skerka, C.; Voigt, K.; Figge, M.T. Virulent strain ofLichtheimia corymbiferashows increased phagocytosis by macrophages as revealed by automated microscopy image analysis. Mycoses 2014, 57, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Ghuman, H.; Voelz, K. Innate and Adaptive Immunity to Mucorales. J. Fungi 2017, 3, 48. [Google Scholar] [CrossRef] [Green Version]
- Perkhofer, S.; Kainzner, B.; Kehrel, B.E.; Dierich, M.P.; Nussbaumer, W.; Lass-Flörl, C. Potential Antifungal Effects of Human Platelets against Zygomycetes In Vitro. J. Infect. Dis. 2009, 200, 1176–1179. [Google Scholar] [CrossRef] [PubMed]
- Simitsopoulou, M.; Roilides, E.; Maloukou, A.; Gil-Lamaignere, C.; Walsh, T.J. Interaction of amphotericin B lipid formulations and triazoles with human polymorphonuclear leucocytes for antifungal activity against Zygomycetes. Mycoses 2008, 51, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Speth, C.; Lass-Flörl, C.; Rambach, G. Platelet immunology in fungal infections. Thromb. Haemost. 2014, 112, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Tramsen, L.; Perkhofer, S.; Lass-Flörl, C.; Hanisch, M.; Röger, F.; Klingebiel, T.; Koehl, U.; Lehrnbecher, T. Rhizopus oryzae hyphae are damaged by human natural killer (NK) cells, but suppress NK cell mediated immunity. Immunobiology 2013, 218, 939–944. [Google Scholar] [CrossRef]
- Schmidt, S.; Tramsen, L.; Lehrnbecher, T. Natural Killer Cells in Antifungal Immunity. Front. Immunol. 2017, 8, 1623. [Google Scholar] [CrossRef]
- Schmidt, S.; Schneider, A.; Demir, A.; Lass-Flörl, C.; Lehrnbecher, T. Natural killer cell-mediated damage of clinical isolates of mucormycetes. Mycoses 2015, 59, 34–38. [Google Scholar] [CrossRef]
- Chamilos, G.; Lewis, R.E.; Hu, J.; Xiao, L.; Zal, T.; Gilliet, M.; Halder, G.; Kontoyiannis, D.P. Drosophila melanogaster as a model host to dissect the immunopathogenesis of zygomycosis. Proc. Natl. Acad. Sci. USA 2008, 105, 9367–9372. [Google Scholar] [CrossRef] [Green Version]
- dos Santos, A.R.; Fraga-Silva, T.F.; Almeida-Donanzam, D.D.F.; dos Santos, R.F.; Finato, A.C.; Soares, C.T.; Lara, V.S.; Almeida, N.L.M.; Andrade, M.I.; de Arruda, O.S.; et al. IFN-γ Mediated Signaling Improves Fungal Clearance in Experimental Pulmonary Mucormycosis. Mycopathologia 2022, 187, 15–30. [Google Scholar] [CrossRef]
- Chinn, R.Y.; Diamond, R.D. Generation of chemotactic factors by Rhizopus oryzae in the presence and absence of serum: Relationship to hyphal damage mediated by human neutrophils and effects of hyperglycemia and ketoacidosis. Infect. Immun. 1982, 38, 1123–1129. [Google Scholar] [CrossRef] [Green Version]
- Borman, A.; Fraser, M.; Patterson, Z.; Palmer, M.; Johnson, E. In Vitro Antifungal Drug Resistance Profiles of Clinically Relevant Members of the Mucorales (Mucoromycota) Especially with the Newer Triazoles. J. Fungi 2021, 7, 271. [Google Scholar] [CrossRef]
- Guinea, J.; Peláez, T.; Recio, S.; Torres-Narbona, M.; Bouza, E. In Vitro Antifungal Activities of Isavuconazole (BAL4815), Voriconazole, and Fluconazole against 1,007 Isolates of Zygomycete, Candida, Aspergillus, Fusarium, and Scedosporium Species. Antimicrob. Agents Chemother. 2008, 52, 1396–1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alastruey-Izquierdo, A.; Castelli, M.; Cuesta, I.; Zaragoza, O.; Monzón, A.; Mellado, E.; Rodríguez-Tudela, J. In vitro activity of antifungals against Zygomycetes. Clin. Microbiol. Infect. 2009, 15, 71–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alastruey-Izquierdo, A.; Cuesta, I.; Walther, G.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L. Antifungal Susceptibility Profile of Human-Pathogenic Species of Lichtheimia. Antimicrob. Agents Chemother. 2010, 54, 3058–3060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhary, A.; Singh, P.K.; Kathuria, S.; Hagen, F.; Meis, J.F. Comparison of the EUCAST and CLSI Broth Microdilution Methods for Testing Isavuconazole, Posaconazole, and Amphotericin B against Molecularly Identified Mucorales Species. Antimicrob. Agents Chemother. 2015, 59, 7882–7887. [Google Scholar] [CrossRef] [Green Version]
- Maurer, E.; Binder, U.; Sparber, M.; Lackner, M.; Caramalho, R.; Lass-Flörl, C. Susceptibility Profiles of Amphotericin B and Posaconazole against Clinically Relevant Mucorales Species under Hypoxic Conditions. Antimicrob. Agents Chemother. 2015, 59, 1344–1346. [Google Scholar] [CrossRef] [Green Version]
- Borman, A.M.; Fraser, M.; Palmer, M.D.; Szekely, A.; Houldsworth, M.; Patterson, Z.; Johnson, E.M. MIC Distributions and Evaluation of Fungicidal Activity for Amphotericin B, Itraconazole, Voriconazole, Posaconazole and Caspofungin and 20 Species of Pathogenic Filamentous Fungi Determined Using the CLSI Broth Microdilution Method. J. Fungi 2017, 3, 27. [Google Scholar] [CrossRef] [Green Version]
- Perkhofer, S.; Lechner, V.; Lass-Flörl, C. In Vitro Activity of Isavuconazole against Aspergillus Species and Zygomycetes According to the Methodology of the European Committee on Antimicrobial Susceptibility Testing. Antimicrob. Agents Chemother. 2009, 53, 1645–1647. [Google Scholar] [CrossRef] [Green Version]
- Arendrup, M.C.; Jensen, R.H.; Meletiadis, J. In Vitro Activity of Isavuconazole and Comparators against Clinical Isolates of the Mucorales Order. Antimicrob. Agents Chemother. 2015, 59, 7735–7742. [Google Scholar] [CrossRef] [Green Version]
- Van Sickels, N.; Hoffman, J.; Stuke, L.; Kempe, K. Survival of a Patient With Trauma-Induced Mucormycosis Using an Aggressive Surgical and Medical Approach. J. Trauma: Inj. Infect. Crit. Care 2011, 70, 507–509. [Google Scholar] [CrossRef]
- Morace, G.; Borghi, E. Invasive Mold Infections: Virulence and Pathogenesis of Mucorales. Int. J. Microbiol. 2012, 2012, 349278. [Google Scholar] [CrossRef] [Green Version]
- Ravi, P.; Ramadorai, A.; Narayanan, V. Rhinocerebral mucormycosis: A prospective analysis of an effective treatment protocol. Ann. Maxillofac. Surg. 2019, 9, 192–196. [Google Scholar] [CrossRef]
- Wei, L.-W.; Zhu, P.-Q.; Chen, X.-Q.; Yu, J. Mucormycosis in Mainland China: A Systematic Review of Case Reports. Mycopathologia 2021, 187, 1–14. [Google Scholar] [CrossRef]
- Mahalaxmi, I.; Jayaramayya, K.; Venkatesan, D.; Subramaniam, M.D.; Renu, K.; Vijayakumar, P.; Narayanasamy, A.; Gopalakrishnan, A.V.; Kumar, N.S.; Sivaprakash, P.; et al. Mucormycosis: An opportunistic pathogen during COVID-19. Environ. Res. 2021, 201, 111643. [Google Scholar] [CrossRef]
- Cornely, O.A.; Alastruey-Izquierdo, A.; Arenz, D.; Chen, S.C.A.; Dannaoui, E.; Hochhegger, B.; Hoenigl, M.; Jensen, H.E.; Lagrou, K.; Lewis, R.E.; et al. Global guideline for the diagnosis and management of mucormycosis: An initiative of the European Confederation of Medical Mycology in cooperation with the Mycoses Study Group Education and Research Consortium. Lancet Infect. Dis. 2019, 19, e405–e421. [Google Scholar] [CrossRef]
- Walsh, T.J.; Finberg, R.W.; Arndt, C.; Hiemenz, J.; Schwartz, C.; Bodensteiner, D.; Pappas, P.; Seibel, N.; Greenberg, R.N.; Dummer, S.; et al. Liposomal amphotericin B for empirical therapy in patients with persistent fever and neutropenia. N. Engl. J. Med. 1999, 340, 764–771. [Google Scholar] [CrossRef]
- Ullmann, A.J.; Sanz, M.A.; Tramarin, A.; Barnes, R.A.; Wu, W.; Gerlach, B.A.; Krobot, K.J.; Gerth, W.C.; Longitudinal Evaluation of Antifungal Drugs (LEAD I) Investigators. Prospective Study of Amphotericin B Formulations in Immunocompromised Patients in 4 European Countries. Clin. Infect. Dis. 2006, 43, e29–e38. [Google Scholar] [CrossRef] [Green Version]
- Marty, F.M.; Ostrosky-Zeichner, L.; Cornely, O.A.; Mullane, K.M.; Perfect, J.R.; Thompson, G.R.; Alangaden, G.J.; Brown, J.M.; Fredricks, D.N.; Heinz, W.J.; et al. Isavuconazole treatment for mucormycosis: A single-arm open-label trial and case-control analysis. Lancet Infect. Dis. 2016, 16, 828–837. [Google Scholar] [CrossRef]
- Roilides, E.; Antachopoulos, C. Isavuconazole: An azole active against mucormycosis. Lancet Infect. Dis. 2016, 16, 761–762. [Google Scholar] [CrossRef]
- Maertens, J.A.; Raad, I.I.; Marr, K.A.; Patterson, T.; Kontoyiannis, D.P.; Cornely, O.; Bow, E.J.; Rahav, G.; Neofytos, D.; Aoun, M.; et al. Isavuconazole versus voriconazole for primary treatment of invasive mould disease caused by Aspergillus and other filamentous fungi (SECURE): A phase 3, randomised-controlled, non-inferiority trial. Lancet 2015, 387, 760–769. [Google Scholar] [CrossRef]
- Mellinghoff, S.C.; Bassetti, M.; Dörfel, D.; Hagel, S.; Lehners, N.; Plis, A.; Schalk, E.; Vena, A.; Cornely, O.A. Isavuconazole shortens the QTc interval. Mycoses 2017, 61, 256–260. [Google Scholar] [CrossRef] [Green Version]
- Ruping, M.J.G.T.; Heinz, W.J.; Kindo, A.J.; Rickerts, V.; Lass-Florl, C.; Beisel, C.; Herbrecht, R.; Roth, Y.; Silling, G.; Ullmann, A.J.; et al. Forty-one recent cases of invasive zygomycosis from a global clinical registry. J. Antimicrob. Chemother. 2009, 65, 296–302. [Google Scholar] [CrossRef] [Green Version]
- Maertens, J.; Cornely, O.A.; Ullmann, A.J.; Heinz, W.J.; Krishna, G.; Patino, H.; Caceres, M.; Kartsonis, N.; Waskin, H.; Robertson, M.N. Phase 1B Study of the Pharmacokinetics and Safety of Posaconazole Intravenous Solution in Patients at Risk for Invasive Fungal Disease. Antimicrob. Agents Chemother. 2014, 58, 3610–3617. [Google Scholar] [CrossRef] [Green Version]
- Duarte, R.F.; López-Jiménez, J.; Cornely, O.A.; Laverdiere, M.; Helfgott, D.; Haider, S.; Chandrasekar, P.; Langston, A.; Perfect, J.; Ma, L.; et al. Phase 1b Study of New Posaconazole Tablet for Prevention of Invasive Fungal Infections in High-Risk Patients with Neutropenia. Antimicrob. Agents Chemother. 2014, 58, 5758–5765. [Google Scholar] [CrossRef] [Green Version]
- Cornely, O.A.; Duarte, R.F.; Haider, S.; Chandrasekar, P.; Helfgott, D.; Jiménez, J.L.; Candoni, A.; Raad, I.; Laverdiere, M.; Langston, A.; et al. Phase 3 pharmacokinetics and safety study of a posaconazole tablet formulation in patients at risk for invasive fungal disease. J. Antimicrob. Chemother. 2015, 71, 718–726. [Google Scholar] [CrossRef] [Green Version]
- Cornely, O.A.; Robertson, M.N.; Haider, S.; Grigg, A.; Geddes, M.; Aoun, M.; Heinz, W.J.; Raad, I.; Schanz, U.; Meyer, R.G.; et al. Pharmacokinetics and safety results from the Phase 3 randomized, open-label, study of intravenous posaconazole in patients at risk of invasive fungal disease. J. Antimicrob. Chemother. 2017, 72, 3406–3413. [Google Scholar] [CrossRef] [Green Version]
- Perkhofer, S.; Locher, M.; Cuenca-Estrella, M.; Rüchel, R.; Würzner, R.; Dierich, M.P.; Lass-Flörl, C. Posaconazole Enhances the Activity of Amphotericin B against Hyphae of Zygomycetes In Vitro. Antimicrob. Agents Chemother. 2008, 52, 2636–2638. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Qiao, J.; Giovanni, G.; Liu, G.; Yang, H.; Wu, J.; Chen, J. Mucormycosis in renal transplant recipients: Review of 174 reported cases. BMC Infect. Dis. 2017, 17, 283. [Google Scholar] [CrossRef]
- Jeong, W.; Keighley, C.; Wolfe, R.; Lee, W.L.; Slavin, M.A.; Chen, S.C.-A.; Kong, D.C. Contemporary management and clinical outcomes of mucormycosis: A systematic review and meta-analysis of case reports. Int. J. Antimicrob. Agents 2019, 53, 589–597. [Google Scholar] [CrossRef]
- Vehreschild, J.J.; Birtel, A.; Vehreschild, M.J.G.T.; Liss, B.J.; Farowski, F.; Kochanek, M.; Sieniawski, M.; Steinbach, A.; Wahlers, K.; Fätkenheuer, G.; et al. Mucormycosis treated with posaconazole: Review of 96 case reports. Crit. Rev. Microbiol. 2012, 39, 310–324. [Google Scholar] [CrossRef]
- Lebeau, O.; Van Delden, C.; Garbino, J.; Robert, J.; Lamoth, F.; Passweg, J.; Chalandon, Y. Disseminated Rhizopus microsporus infection cured by salvage allogeneic hematopoietic stem cell transplantation, antifungal combination therapy, and surgical resection. Transpl. Infect. Dis. 2010, 12, 269–272. [Google Scholar] [CrossRef]
- Roux, B.G.-L.; Méchinaud, F.; Gay-Andrieu, F.; Lortholary, O.; Dannaoui, E.; Hoinard, D.; Corradini, N. Successful Triple Combination Therapy of Disseminated Absidia corymbifera Infection in an Adolescent With Osteosarcoma. J. Pediatr. Hematol. 2010, 32, 131–133. [Google Scholar] [CrossRef]
- Warkentien, T.; Rodriguez, C.; Lloyd, B.; Wells, J.; Weintrob, A.; Dunne, J.R.; Ganesan, A.; Li, P.; Bradley, W.; Gaskins, L.J.; et al. Invasive Mold Infections Following Combat-related Injuries. Clin. Infect. Dis. 2012, 55, 1441–1449. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, C.J.; Tribble, D.R.; Malone, D.L.; Murray, C.K.; Jessie, E.M.; Khan, M.; Fleming, M.E.; Potter, B.K.; Gordon, W.T.; Shackelford, S.A. Treatment of Suspected Invasive Fungal Infection in War Wounds. Mil. Med. 2018, 183, 142–146. [Google Scholar] [CrossRef] [Green Version]
- Symeonidis, A.S. The role of iron and iron chelators in zygomycosis. Clin. Microbiol. Infect. 2009, 15, 26–32. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, S.; Tramsen, L.; Perkhofer, S.; Lass-Flörl, C.; Röger, F.; Schubert, R.; Lehrnbecher, T. Characterization of the Cellular Immune Responses to Rhizopus oryzae With Potential Impact on Immunotherapeutic Strategies in Hematopoietic Stem Cell Transplantation. J. Infect. Dis. 2012, 206, 135–139. [Google Scholar] [CrossRef] [Green Version]
- Castillo, P.; Wright, K.E.; Kontoyiannis, D.P.; Walsh, T.; Patel, S.; Chorvinsky, E.; Bose, S.; Hazrat, Y.; Omer, B.; Albert, N.; et al. A New Method for Reactivating and Expanding T Cells Specific for Rhizopus oryzae. Mol. Ther. Methods Clin. Dev. 2018, 9, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Gil-Lamaignere, C.; Simitsopoulou, M.; Roilides, E.; Maloukou, A.; Winn, R.M.; Walsh, T.J. Interferon-γ and Granulocyte-Macrophage Colony-Stimulating Factor Augment the Activity of Polymorphonuclear Leukocytes against Medically Important Zygomycetes. J. Infect. Dis. 2005, 191, 1180–1187. [Google Scholar] [CrossRef] [Green Version]
- Grimaldi, D.; Pradier, O.; Hotchkiss, R.S.; Vincent, J.-L. Nivolumab plus interferon-γ in the treatment of intractable mucormycosis. Lancet Infect. Dis. 2017, 17, 18. [Google Scholar] [CrossRef] [Green Version]
- Paltauf, A. Mycosis mucorina. Virchows Arch. Pathol. Anat. Physiol. Klin. Med. 2005, 102, 543–564. [Google Scholar] [CrossRef]
- Al-Tawfiq, J.A.; Alhumaid, S.; Alshukairi, A.N.; Temsah, M.-H.; Barry, M.; Al Mutair, A.; Rabaan, A.A.; Al-Omari, A.; Tirupathi, R.; AlQahtani, M.; et al. COVID-19 and mucormycosis superinfection: The perfect storm. Infection 2021, 49, 833–853. [Google Scholar] [CrossRef]
- Chakrabarti, A. The recent mucormycosis storm over Indian sky. Indian J. Med Microbiol. 2021, 39, 269–270. [Google Scholar] [CrossRef]
- Suvvari, T.K.; Arigapudi, N.; Kandi, V.R.; Kutikuppala, L.S. Mucormycosis: A killer in the shadow of COVID-19. J. De Mycol. Med. 2021, 31, 101161. [Google Scholar] [CrossRef]
- Musuuza, J.S.; Watson, L.; Parmasad, V.; Putman-Buehler, N.; Christensen, L.; Safdar, N. Prevalence and outcomes of co-infection and superinfection with SARS-CoV-2 and other pathogens: A systematic review and meta-analysis. PLoS ONE 2021, 16, e0251170. [Google Scholar] [CrossRef]
- Arakeri, G.; Rao Us, V.; Mendes, R.A.; Oeppen, R.S.; Brennan, P.A. COVID-associated mucormycosis (CAM): Is the Delta variant a cause? Br. J. Oral Maxillofac. Surg. 2021, 59, 1095–1098. [Google Scholar] [CrossRef]
- Pal, R.; Singh, B.; Bhadada, S.K.; Banerjee, M.; Bhogal, R.S.; Hage, N.; Kumar, A. COVID-19-associated mucormycosis: An updated systematic review of literature. Mycoses 2021, 64, 1452–1459. [Google Scholar] [CrossRef]
- Sharma, S.; Grover, M.; Bhargava, S.; Samdani, S.; Kataria, T. Post coronavirus disease mucormycosis: A deadly addition to the pandemic spectrum. J. Laryngol. Otol. 2021, 135, 442–447. [Google Scholar] [CrossRef]
- Afzal, S.; Nasir, M. Aspergillosis and mucormycosis in COVID-19 patients; a systematic review and meta-analysis. Medrxiv 2021. [Google Scholar] [CrossRef]
- Ramaswami, A.; Sahu, A.K.; Kumar, A.; Suresh, S.; Nair, A.; Gupta, D.; Chouhan, R.; Bhat, R.; Mathew, R.; Majeed, J.A.; et al. COVID-19-associated mucormycosis presenting to the Emergency Department—an observational study of 70 patients. QJM: Int. J. Med. 2021, 114, 464–470. [Google Scholar] [CrossRef]
- Selarka, L.; Sharma, S.; Saini, D.; Sharma, S.; Batra, A.; Waghmare, V.T.; Dileep, P.; Patel, S.; Shah, M.; Parikh, T.; et al. Mucormycosis and COVID-19: An epidemic within a pandemic in India. Mycoses 2021, 64, 1253–1260. [Google Scholar] [CrossRef]
- Bhanuprasad, K.; Manesh, A.; Devasagayam, E.; Varghese, L.; Cherian, L.M.; Kurien, R.; Karthik, R.; Deodhar, D.; Vanjare, H.; Peter, J.; et al. Risk factors associated with the mucormycosis epidemic during the COVID-19 pandemic. Int. J. Infect. Dis. 2021, 111, 267–270. [Google Scholar] [CrossRef]
- Aranjani, J.M.; Manuel, A.; Razack, H.I.A.; Mathew, S.T. COVID-19–associated mucormycosis: Evidence-based critical review of an emerging infection burden during the pandemic’s second wave in India. PLOS Neglected Trop. Dis. 2021, 15, e0009921. [Google Scholar] [CrossRef] [PubMed]
- Honavar, S.G.; Sen, M.; Lahane, S.; Lahane, T.P.; Parekh, R. Mucor in a Viral Land: A Tale of Two Pathogens. Indian J. Ophthalmol. 2021, 69, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Freeland, B.; Funnell, M. Corticosteroid-induced hyperglycemia. Nursing 2012, 42, 68–69. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhong, L.; Deng, J.; Peng, J.; Dan, H.; Zeng, X.; Li, T.; Chen, Q. High expression of ACE2 receptor of 2019-nCoV on the epithelial cells of oral mucosa. Int. J. Oral Sci. 2020, 12, 8. [Google Scholar] [CrossRef]
- Agnihotri, A.K.; Vij, M.; Aruoma, O.I.; Yagnik, V.D.; Bahorun, T.; Villamil, M.E.; Menezes, G.A.; Gupta, V. The double trouble: COVID-19 associated mucormycosis a focused review and future perspectives. Glob. J. Medical, Pharm. Biomed. Updat. 2021, 16, 4. [Google Scholar] [CrossRef]
- Lim, S.; Bae, J.H.; Kwon, H.-S.; Nauck, M.A. COVID-19 and diabetes mellitus: From pathophysiology to clinical management. Nat. Rev. Endocrinol. 2020, 17, 11–30. [Google Scholar] [CrossRef]
- Hulter, H.N.; Licht, J.H.; Bonner, E.L.; Glynn, R.D.; Sebastián, A. Effects of glucocorticoid steroids on renal and systemic acid-base metabolism. Am. J. Physiol. Physiol. 1980, 239, F30–F43. [Google Scholar] [CrossRef]
- Damgaci, S.; Ibrahim-Hashim, A.; Enríquez-Navas, P.M.; Pilon-Thomas, S.; Guvenis, A.; Gillies, R.J. Hypoxia and acidosis: Immune suppressors and therapeutic targets. Immunology 2018, 154, 354–362. [Google Scholar] [CrossRef] [Green Version]
- Bhogireddy, R.; Krishnamurthy, V.; Jabaris S, S.L.; Pullaiah, C.P.; Manohar, S. Is Mucormycosis an inevitable complication of COVID-19 in India? Braz. J. Infect. Dis. 2021, 25, 101597. [Google Scholar] [CrossRef]
- Ruhl, L.; Pink, I.; Kühne, J.F.; Beushausen, K.; Keil, J.; Christoph, S.; Sauer, A.; Boblitz, L.; Schmidt, J.; David, S.; et al. Endothelial dysfunction contributes to severe COVID-19 in combination with dysregulated lymphocyte responses and cytokine networks. Signal Transduct. Target. Ther. 2021, 6, 418. [Google Scholar] [CrossRef]
- Elyaspour, Z.; Zibaeenezhad, M.J.; Razmkhah, M.; Razeghian-Jahromi, I. Is It All About Endothelial Dysfunction and Thrombosis Formation? The Secret of COVID-19. Clin. Appl. Thromb. 2021, 27, 10760296211042940. [Google Scholar] [CrossRef] [PubMed]
- Lugito, N.P.H.; Cucunawangsih, C. How Does Mucorales Benefit from the Dysregulated Iron Homeostasis During SARS-CoV-2 Infection? Mycopathologia 2021, 186, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Kawasumi, H.; Gono, T.; Kawaguchi, Y.; Kaneko, H.; Katsumata, Y.; Hanaoka, M.; Kataoka, S.; Yamanaka, H. IL-6, IL-8, and IL-10 Are Associated with Hyperferritinemia in Rapidly Progressive Interstitial Lung Disease with Polymyositis/Dermatomyositis. BioMed Res. Int. 2014, 2014, 815245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perricone, C.; Bartoloni, E.; Bursi, R.; Cafaro, G.; Guidelli, G.M.; Shoenfeld, Y.; Gerli, R. COVID-19 as part of the hyperferritinemic syndromes: The role of iron depletion therapy. Immunol. Res. 2020, 68, 213–224. [Google Scholar] [CrossRef]
- John, T.; Jacob, C.; Kontoyiannis, D. When Uncontrolled Diabetes Mellitus and Severe COVID-19 Converge: The Perfect Storm for Mucormycosis. J. Fungi 2021, 7, 298. [Google Scholar] [CrossRef]
- Edeas, M.; Saleh, J.; Peyssonnaux, C. Iron: Innocent bystander or vicious culprit in COVID-19 pathogenesis? Int. J. Infect. Dis. 2020, 97, 303–305. [Google Scholar] [CrossRef]
- Campione, E.; Cosio, T.; Rosa, L.; Lanna, C.; Di Girolamo, S.; Gaziano, R.; Valenti, P.; Bianchi, L. Lactoferrin as Protective Natural Barrier of Respiratory and Intestinal Mucosa against Coronavirus Infection and Inflammation. Int. J. Mol. Sci. 2020, 21, 4903. [Google Scholar] [CrossRef]
- Campione, E.; Lanna, C.; Cosio, T.; Rosa, L.; Conte, M.P.; Iacovelli, F.; Romeo, A.; Falconi, M.; Del Vecchio, C.; Franchin, E.; et al. Lactoferrin as Antiviral Treatment in COVID-19 Management: Preliminary Evidence. Int. J. Environ. Res. Public Heal. 2021, 18, 10985. [Google Scholar] [CrossRef]
- Campione, E.; Lanna, C.; Cosio, T.; Rosa, L.; Conte, M.P.; Iacovelli, F.; Romeo, A.; Falconi, M.; Del Vecchio, C.; Franchin, E.; et al. Lactoferrin Against SARS-CoV-2: In Vitro and In Silico Evidences. Front. Pharmacol. 2021, 12, 666600. [Google Scholar] [CrossRef]
- Ackermann, M.; Verleden, S.E.; Kuehnel, M.; Haverich, A.; Welte, T.; Laenger, F.; Vanstapel, A.; Werlein, C.; Stark, H.; Tzankov, A.; et al. Pulmonary Vascular Endothelialitis, Thrombosis, and Angiogenesis in COVID-19. N. Engl. J. Med. 2020, 383, 120–128. [Google Scholar] [CrossRef]
- Glowacka, I.; Bertram, S.; Herzog, P.; Pfefferle, S.; Steffen, I.; Muench, M.O.; Simmons, G.; Hofmann, H.; Kuri, T.; Weber, F.; et al. Differential Downregulation of ACE2 by the Spike Proteins of Severe Acute Respiratory Syndrome Coronavirus and Human Coronavirus NL63. J. Virol. 2010, 84, 1198–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, J.J.; Chan, A.C.; Sharma, A.K.; Sharma, S.; Sharma, V.K. The mucormycosis epidemic within COVID-19 pandemic- lessons from India. Brain, Behav. Immun. 2021, 97, 4–5. [Google Scholar] [CrossRef]
- Honavar, S.; Sen, M.; Sengupta, S.; Rao, R.; Kim, U.; Sharma, M.; Sachdev, M.; Grover, A.; Surve, A.; Budharapu, A.; et al. EpImpact of the use of digital devices on e of COVID-19-associated rhino-orbital-cerebral mucormycosis in 2826 patients in India–Collaborative OPAI-IJO Study on Mucormycosis in COVID-19 (COSMIC), Report 1. Indian J. Ophthalmol. 2021, 69, 1670–1692. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.; Shetty, A.; Nagesh, C. Orbital infarction syndrome secondary to rhino-orbital mucormycosis in a case of COVID-19: Clinico-radiological features. Indian J. Ophthalmol. 2021, 69, 1627–1630. [Google Scholar] [CrossRef] [PubMed]
- Pakdel, F.; Ahmadikia, K.; Salehi, M.; Tabari, A.; Jafari, R.; Mehrparvar, G.; Rezaie, Y.; Rajaeih, S.; Alijani, N.; Barac, A.; et al. Mucormycosis in patients with COVID-19: A cross-sectional descriptive multicentre study from Iran. Mycoses 2021, 64, 1238–1252. [Google Scholar] [CrossRef]
- Ashour, M.M.; Abdelaziz, T.T.; Ashour, D.M.; Askoura, A.; Saleh, M.I.; Mahmoud, M.S. Imaging spectrum of acute invasive fungal rhino-orbital-cerebral sinusitis in COVID-19 patients: A case series and a review of literature. J. Neuroradiol. 2021, 48, 319–324. [Google Scholar] [CrossRef]
- Joshi, A.R.; Muthe, M.M.; Patankar, S.H.; Athawale, A.; Achhapalia, Y. CT and MRI Findings of Invasive Mucormycosis in the Setting of COVID-19: Experience From a Single Center in India. Am. J. Roentgenol. 2021, 217, 1431–1432. [Google Scholar] [CrossRef]
- Garg, D.; Muthu, V.; Sehgal, I.S.; Ramachandran, R.; Kaur, H.; Bhalla, A.; Puri, G.D.; Chakrabarti, A.; Agarwal, R. Coronavirus Disease (COVID-19) Associated Mucormycosis (CAM): Case Report and Systematic Review of Literature. Mycopathologia 2021, 186, 289–298. [Google Scholar] [CrossRef]
- Rudramurthy, S.M.; Hoenigl, M.; Meis, J.F.; Cornely, O.A.; Muthu, V.; Gangneux, J.P.; Perfect, J.; Chakrabarti, A.; Isham, E.A. ECMM/ISHAM recommendations for clinical management of COVID-19 associated mucormycosis in low- and middle-income countries. Mycoses 2021, 64, 1028–1037. [Google Scholar] [CrossRef]
- Reid, G.; Lynch, J.P.; Fishbein, M.C.; Clark, N.M. Mucormycosis. Semin. Respir. Crit. Care Med. 2020, 41, 099–114. [Google Scholar] [CrossRef]
- Machouart, M.; Larché, J.; Burton, K.; Collomb, J.; Maurer, P.; Cintrat, A.; Biava, M.F.; Greciano, S.; Kuijpers, A.F.A.; Contet-Audonneau, N.; et al. Genetic Identification of the Main Opportunistic Mucorales by PCR-Restriction Fragment Length Polymorphism. J. Clin. Microbiol. 2006, 44, 805–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lass-Flörl, C.; Mayr, A. Diagnosing invasive fungal diseases–limitations of microbiological diagnostic methods. Expert Opin. Med Diagn. 2009, 3, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.; Dharne, M. COVID-19 associated mucormycosis: Evolving technologies for early and rapid diagnosis. 3 Biotech 2022, 12, 6. [Google Scholar] [CrossRef] [PubMed]
- Lass-Flörl, C. Zygomycosis: Conventional laboratory diagnosis. Clin. Microbiol. Infect. 2009, 15, 60–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kung, V.L.; Chernock, R.; Burnham, C.-A.D. Diagnostic accuracy of fungal identification in histopathology and cytopathology specimens. Eur. J. Clin. Microbiol. 2017, 37, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Hamilos, G.; Samonis, G.; Kontoyiannis, D.P. Pulmonary Mucormycosis. Semin. Respir. Crit. Care Med. 2011, 32, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Hoenigl, M.; Seidel, D.; Carvalho, A.; Rudramurthy, S.M.; Arastehfar, A.; Gangneux, J.-P.; Nasir, N.; Bonifaz, A.; Araiza, J.; Klimko, N.; et al. The emergence of COVID-19 associated mucormycosis: A review of cases from 18 countries. Lancet Microbe 2022. [Google Scholar] [CrossRef]
- Koehler, P.; Bassetti, M.; Chakrabarti, A.; Chen, S.C.A.; Colombo, A.L.; Hoenigl, M.; Klimko, N.; Lass-Flörl, C.; Oladele, R.O.; Vinh, D.C.; et al. Defining and managing COVID-19-associated pulmonary aspergillosis: The 2020 ECMM/ISHAM consensus criteria for research and clinical guidance. Lancet Infect. Dis. 2021, 21, e149–e162. [Google Scholar] [CrossRef]
- Hsiao, C.R.; Huang, L.; Bouchara, J.-P.; Barton, R.; Li, H.C.; Chang, T.C. Identification of Medically Important Molds by an Oligonucleotide Array. J. Clin. Microbiol. 2005, 43, 3760–3768. [Google Scholar] [CrossRef] [Green Version]
- Gholinejad-Ghadi, N.; Shokohi, T.; Seifi, Z.; Aghili, S.R.; Roilides, E.; Nikkhah, M.; Pormosa, R.; Karami, H.; Larjani, L.V.; Ghasemi, M.; et al. Identification of Mucorales in patients with proven invasive mucormycosis by polymerase chain reaction in tissue samples. Mycoses 2018, 61, 909–915. [Google Scholar] [CrossRef]
- Larché, J.; Machouart, M.; Burton, K.; Collomb, J.; Biava, M.F.; Gérard, A.; Fortier, B. Diagnosis of Cutaneous Mucormycosis Due to Rhizopus microsporus by an Innovative PCR-Restriction Fragment-Length Polymorphism Method. Clin. Infect. Dis. 2005, 41, 1362–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rickerts, V.; Just-Nübling, G.; Konrad, F.; Kern, J.; Lambrecht, E.; Böhme, A.; Jacobi, V.; Bialek, R. Diagnosis of invasive aspergillosis and mucormycosis in immunocompromised patients by seminested PCR assay of tissue samples. Eur. J. Clin. Microbiol. 2006, 25, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Hata, D.J.; Buckwalter, S.P.; Pritt, B.S.; Roberts, G.D.; Wengenack, N.L. Real-Time PCR Method for Detection of Zygomycetes. J. Clin. Microbiol. 2008, 46, 2353–2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millon, L.; Larosa, F.; Lepiller, Q.; Legrand, F.; Rocchi, S.; Daguindau, E.; Scherer, E.; Bellanger, A.-P.; Leroy, J.; Grenouillet, F. Quantitative Polymerase Chain Reaction Detection of Circulating DNA in Serum for Early Diagnosis of Mucormycosis in Immunocompromised Patients. Clin. Infect. Dis. 2013, 56, e95–e101. [Google Scholar] [CrossRef] [Green Version]
- Millon, L.; Herbrecht, R.; Grenouillet, F.; Morio, F.; Alanio, A.; Letscher-Bru, V.; Cassaing, S.; Chouaki, T.; Kauffmann-Lacroix, C.; Poirier, P.; et al. Early diagnosis and monitoring of mucormycosis by detection of circulating DNA in serum: Retrospective analysis of 44 cases collected through the French Surveillance Network of Invasive Fungal Infections (RESSIF). Clin. Microbiol. Infect. 2016, 22, 810.e1–810.e8. [Google Scholar] [CrossRef] [Green Version]
- Nyilasi, I.; Papp, T.; Csernetics, A.; Krizsán, K.; Nagy, E.; Vágvölgyi, C. High-affinity iron permease (FTR1) gene sequence-based molecular identification of clinically important Zygomycetes. Clin. Microbiol. Infect. 2008, 14, 393–397. [Google Scholar] [CrossRef] [Green Version]
- Jensen, H.E.; Salonen, J.; Ekfors, T.O. The use of immunohistochemistry to improve sensitivity and specificity in the diagnosis of systemic mycoses in patients with haematological malignancies. J. Pathol. 1997, 181, 100–105. [Google Scholar] [CrossRef]
- Sunagawa, K.; Ishige, T.; Kusumi, Y.; Asano, M.; Nisihikawa, E.; Kato, M.; Yagasaki, H.; Nemoto, N. Renal abscess involving mucormycosis by immunohistochemical detection in a patient with acute lymphocytic leukemia: A case report and literature review. Jpn. J. Infect. Dis. 2013, 66, 345–347. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.; Park, Y.S.; Sung, H.; Song, J.S.; Lee, S.-O.; Choi, S.-H.; Kim, Y.S.; Woo, J.H.; Kim, S.-H. Using immunohistochemistry to assess the accuracy of histomorphologic diagnosis of aspergillosis and mucormycosis. Clin. Infect. Dis. 2015, 61, 1664–1670. [Google Scholar] [CrossRef] [Green Version]
- Hammond, S.P.; Bialek, R.; Milner, D.A.; Petschnigg, E.M.; Baden, L.R.; Marty, F.M. Molecular Methods To Improve Diagnosis and Identification of Mucormycosis. J. Clin. Microbiol. 2011, 49, 2151–2153. [Google Scholar] [CrossRef] [Green Version]
- Salehi, E.; Hedayati, M.T.; Zoll, J.; Rafati, H.; Ghasemi, M.; Doroudinia, A.; Abastabar, M.; Tolooe, A.; Snelders, E.; van der Lee, H.A.; et al. Discrimination of Aspergillosis, Mucormycosis, Fusariosis, and Scedosporiosis in Formalin-Fixed Paraffin-Embedded Tissue Specimens by Use of Multiple Real-Time Quantitative PCR Assays. J. Clin. Microbiol. 2016, 54, 2798–2803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, H.; Yuan, Z.; Shen, J.; Wang, Z.; Xu, Y. Accuracy of Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry for Identification of Clinical Pathogenic Fungi: A Meta-Analysis. J. Clin. Microbiol. 2014, 52, 2573–2582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spanu, T.; Posteraro, B.; Fiori, B.; D’Inzeo, T.; Campoli, S.; Ruggeri, A.; Tumbarello, M.; Canu, G.; Trecarichi, E.M.; Parisi, G.; et al. Direct MALDI-TOF Mass Spectrometry Assay of Blood Culture Broths for Rapid Identification of Candida Species Causing Bloodstream Infections: An Observational Study in Two Large Microbiology Laboratories. J. Clin. Microbiol. 2012, 50, 176–179. [Google Scholar] [CrossRef] [Green Version]
- Schrödl, W.; Heydel, T.; Schwartze, V.U.; Hoffmann, K.; Große-Herrenthey, A.; Walther, G.; Alastruey-Izquierdo, A.; Rodriguez-Tudela, J.L.; Olias, P.; Jacobsen, I.D.; et al. Direct Analysis and Identification of Pathogenic Lichtheimia Species by Matrix-Assisted Laser Desorption Ionization–Time of Flight Analyzer-Mediated Mass Spectrometry. J. Clin. Microbiol. 2012, 50, 419–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, A.F.; Walchak, R.C.; Miller, H.B.; Slechta, E.S.; Kamboj, K.; Riebe, K.; Robertson, A.E.; Gilbreath, J.J.; Mitchell, K.F.; Wallace, M.A.; et al. Multicenter Study Demonstrates Standardization Requirements for Mold Identification by MALDI-TOF MS. Front. Microbiol. 2019, 10, 2098. [Google Scholar] [CrossRef] [PubMed]
- Koshy, S.; Ismail, N.; Astudillo, C.L.; Haeger, C.M.; Aloum, O.; Acharige, M.T.; Farmakiotis, D.; Baden, L.R.; Marty, F.; Kontoyiannis, D.P.; et al. Breath-Based Diagnosis of Invasive Mucormycosis (IM). Open Forum Infect. Dis. 2017, 4, S53–S54. [Google Scholar] [CrossRef]
- Richardson, M.; Page, I. Role of Serological Tests in the Diagnosis of Mold Infections. Curr. Fungal Infect. Rep. 2018, 12, 127–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angebault, C.; Lanternier, F.; Dalle, F.; Schrimpf, C.; Roupie, A.-L.; Dupuis, A.; Agathine, A.; Scemla, A.; Paubelle, E.; Caillot, D.; et al. Prospective Evaluation of Serum β-Glucan Testing in Patients With Probable or Proven Fungal Diseases. Open Forum Infect. Dis. 2016, 3, ofw128. [Google Scholar] [CrossRef]
- Girdhar, D.; Manocha, E. A Comprehensive Review on the Management of COVID-19-Associated Mucormycosis (CAM): The New Basics. BioMed 2022, 2, 181–198. [Google Scholar] [CrossRef]
- Schwarz, P.; Lortholary, O.; Dromer, F.; Dannaoui, E. Carbon Assimilation Profiles as a Tool for Identification of Zygomycetes. J. Clin. Microbiol. 2007, 45, 1433–1439. [Google Scholar] [CrossRef] [Green Version]
- Berkow, E.L.; Lockhart, S.R.; Ostrosky-Zeichner, L. Antifungal Susceptibility Testing: Current Approaches. Clin. Microbiol. Rev. 2020, 33, e00069-19. [Google Scholar] [CrossRef] [PubMed]
- Honavar, S. Code Mucor: Guidelines for the Diagnosis, Staging and Management of Rhino-Orbito-Cerebral Mucormycosis in the Setting of COVID-19. Indian J. Ophthalmol. 2021, 69, 1361–1365. [Google Scholar] [CrossRef]
- Sahu, R.K.; Salem-Bekhit, M.M.; Bhattacharjee, B.; Almoshari, Y.; Ikbal, A.M.A.; Alshamrani, M.; Bharali, A.; Salawi, A.; Widyowati, R.; Alshammari, A.; et al. Mucormycosis in Indian COVID-19 Patients: Insight into Its Patho-Genesis, Clinical Manifestation, and Management Strategies. Antibiotics 2021, 10, 1079. [Google Scholar] [CrossRef] [PubMed]
- León-Buitimea, A.; Garza-Cervantes, J.A.; Gallegos-Alvarado, D.Y.; Osorio-Concepción, M.; Morones-Ramírez, J.R. Nanomaterial-Based Antifungal Therapies to Combat Fungal Diseases Aspergillosis, Coccidioidomycosis, Mucormycosis, and Candidiasis. Pathogens 2021, 10, 1303. [Google Scholar] [CrossRef] [PubMed]
- Vuorio, A.; Kovanen, P.T. Mucormycosis and glucose-regulated protein 78 in COVID-19: Amenable to statin treatment? J. Intern. Med. 2021, 290, 931–933. [Google Scholar] [CrossRef]
- Chatterjee, S.; Vardhan, B.; Singh, D.K.; Maitra, A.; Ojha, U.K. Should statins be considered for the management of mucormycosis in COVID-19? Diabetes Metab. Syndr. Clin. Res. Rev. 2021, 15, 102162. [Google Scholar] [CrossRef]
- Gebremariam, T.; Alkhazraji, S.; Soliman, S.S.M.; Gu, Y.; Jeon, H.H.; Zhang, L.; French, S.W.; Stevens, D.A.; Edwards, J.E.; Filler, S.G.; et al. Anti-CotH3 antibodies protect mice from mucormycosis by prevention of invasion and augmenting opsonophagocytosis. Sci. Adv. 2019, 5, eaaw1327. [Google Scholar] [CrossRef] [Green Version]
- Alqarihi, A.; Gebremariam, T.; Gu, Y.; Swidergall, M.; Alkhazraji, S.; Soliman, S.S.M.; Bruno, V.M.; Edwards, J.E.; Filler, S.G.; Uppuluri, P.; et al. GRP78 and Integrins Play Different Roles in Host Cell Invasion during Mucormycosis. mBio 2020, 11, e01087-20. [Google Scholar] [CrossRef]
- John, B.; Chamilos, G.; Kontoyiannis, D. Hyperbaric oxygen as an adjunctive treatment for zygomycosis. Clin. Microbiol. Infect. 2005, 11, 515–517. [Google Scholar] [CrossRef] [Green Version]
- Alshahawey, M.G.; El-Housseiny, G.S.; Elsayed, N.S.; Alshahrani, M.Y.; EL Wakeel, L.M.; Aboshanab, K.M. New insights on mucormycosis and its association with the COVID-19 pandemic. Futur. Sci. OA 2022, 8, FSO772. [Google Scholar] [CrossRef]
- Dalié, D.; Deschamps, A.; Richard-Forget, F. Lactic acid bacteria–Potential for control of mould growth and mycotoxins: A review. Food Control 2010, 21, 370–380. [Google Scholar] [CrossRef]
- Aiko, V.; Edamana, P.; Mehta, A. Decomposition and detoxification of aflatoxin B1by lactic acid. J. Sci. Food Agric. 2015, 96, 1959–1966. [Google Scholar] [CrossRef] [PubMed]
- Russo, P.; Arena, M.P.; Fiocco, D.; Capozzi, V.; Drider, D.; Spano, G. Lactobacillus plantarum with broad antifungal activity: A promising approach to increase safety and shelf-life of cereal-based products. Int. J. Food Microbiol. 2017, 247, 48–54. [Google Scholar] [CrossRef]
- Arasu, M.V.; Al-Dhabi, N.A. In vitro antifungal, probiotic, and antioxidant functional properties of a novel Lactobacillus paraplantarum isolated from fermented dates in Saudi Arabia. J. Sci. Food Agric. 2017, 97, 5287–5295. [Google Scholar] [CrossRef] [PubMed]
- Azizkhani, M.; Saris, P.E.J.; Baniasadi, M. An in-vitro assessment of antifungal and antibacterial activity of cow, camel, ewe, and goat milk kefir and probiotic yogurt. J. Food Meas. Charact. 2020, 15, 406–415. [Google Scholar] [CrossRef]
- Brzozowski, T.; Zwolinska-Wcislo, M.; Konturek, P.C.; Kwiecien, S.; Drozdowicz, D.; Konturek, S.J.; Stachura, J.; Budak, A.; Bogdal, J.; Pawlik, W.W.; et al. Influence of gastric colonization withCandida albicanson ulcer healing in rats: Effect of ranitidine, aspirin and probiotic therapy. Scand. J. Gastroenterol. 2005, 40, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Vicariotto, F.; Del Piano, M.; Mogna, L.; Mogna, G. Effectiveness of the Association of 2 Probiotic Strains Formulated in a Slow Release Vaginal Product, in Women Affected by Vulvovaginal Candidiasis. J. Clin. Gastroenterol. 2012, 46, S73–S80. [Google Scholar] [CrossRef] [Green Version]
- Demirel, G.; Celik, I.H.; Erdeve, O.; Saygan, S.; Dilmen, U.; Canpolat, F.E. Prophylactic Saccharomyces boulardii versus nystatin for the prevention of fungal colonization and invasive fungal infection in premature infants. Eur. J. Pediatr. 2013, 172, 1321–1326. [Google Scholar] [CrossRef]
- Matsubara, V.H.; Bandara, H.; Mayer, M.; Samaranayake, L.P. Probiotics as Antifungals in Mucosal Candidiasis. Clin. Infect. Dis. 2016, 62, 1143–1153. [Google Scholar] [CrossRef] [Green Version]
- He, L.-H.; Ren, L.-F.; Li, J.-F.; Wu, Y.-N.; Li, X.; Zhang, L. Intestinal Flora as a Potential Strategy to Fight SARS-CoV-2 Infection. Front. Microbiol. 2020, 11, 1388. [Google Scholar] [CrossRef]
- Morais, A.H.A.; Passos, T.S.; Maciel, B.L.L.; Da Silva-Maia, J.K. Can Probiotics and Diet Promote Beneficial Immune Modulation and Purine Control in Coronavirus Infection? Nutrients 2020, 12, 1737. [Google Scholar] [CrossRef] [PubMed]
- Olaimat, A.N.; Aolymat, I.; Al-Holy, M.; Ayyash, M.; Abu Ghoush, M.; Al-Nabulsi, A.A.; Osaili, T.; Apostolopoulos, V.; Liu, S.-Q.; Shah, N.P. The potential application of probiotics and prebiotics for the prevention and treatment of COVID-19. NPJ Sci. Food 2020, 4, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gohil, K.; Samson, R.; Dastager, S.; Dharne, M. Probiotics in the prophylaxis of COVID-19: Something is better than nothing. 3 Biotech 2020, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chhibber-Goel, J.; Gopinathan, S.; Sharma, A. Interplay between severities of COVID-19 and the gut microbiome: Implications of bacterial co-infections? Gut Pathog. 2021, 13, 1–6. [Google Scholar] [CrossRef]
- Oliveira, L.D.M.; Teixeira, F.M.E.; Sato, M.N. Impact of Retinoic Acid on Immune Cells and Inflammatory Diseases. Mediat. Inflamm. 2018, 2018, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Goodarzi, A.; Sadeghzadeh-Bazargan, A.; Behrangi, E. Systemic retinoids in COVID-19 era, are they helpful, safe or harmful? A comprehensive review study. Iran. J. Dermatol. 2020, 23, 9–12. [Google Scholar] [CrossRef]
- Cosio, T.; Gaziano, R.; Zuccari, G.; Costanza, G.; Grelli, S.; Di Francesco, P.; Bianchi, L.; Campione, E. Retinoids in Fungal Infections: From Bench to Bedside. Pharmaceuticals 2021, 14, 962. [Google Scholar] [CrossRef]
- Ross, A.C.; Stephensen, C.B. Vitamin A and retinoids in antiviral responses. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1996, 10, 979–985. [Google Scholar] [CrossRef] [Green Version]
- Cengiz, A.B.; Ozyilmaz, C.; Tabaru, A.; Kayabasi, S.; Ege, S.S.; Ruzgar, S.; Emre, F.; Oktay, M.F. Effects of oral ısotretinoin on normal and wounded nasal mucosa: An experimental study. Eur. Arch. Oto-Rhino-Laryngol. 2018, 275, 3025–3031. [Google Scholar] [CrossRef]
- Msc, A.A.; Vestita, M.; Msc, H.S.E.; Ayhan, E.; An, I.; Öztürk, M.; Goldust, M. Systemic isotretinoin therapy in the era of COVID-19. Dermatol. Ther. 2020, 33, e13482. [Google Scholar] [CrossRef]
- Oikawa, T.; Hirotani, K.; Nakamura, O.; Shudo, K.; Hiragun, A.; Iwaguchi, T. A highly potent antiangiogenic activity of retinoids. Cancer Lett. 1989, 48, 157–162. [Google Scholar] [CrossRef]
- Wu, C.; Liu, Y.; Yang, Y.; Zhang, P.; Zhong, W.; Wang, Y.; Wang, Q.; Xu, Y.; Li, M.; Li, X.; et al. Analysis of therapeutic targets for SARS-CoV-2 and discovery of potential drugs by computational methods. Acta Pharm. Sin. B 2020, 10, 766–788. [Google Scholar] [CrossRef] [PubMed]
- Midha, I.K.; Kumar, N.; Kumar, A.; Madan, T. Mega doses of retinol: A possible immunomodulation in COVID-19 illness in resource-limited settings. Rev. Med Virol. 2020, 31, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Campione, E.; Paterno, E.J.; Diluvio, L.; Costanza, G.; Bianchi, L.; Carboni, I.; Chimenti, S.; Orlandi, A.; Marino, D.; Favalli, C. Tazarotene as alternative topical treatment for onychomycosis. Drug Des. Dev. Ther. 2015, 9, 879–886. [Google Scholar] [CrossRef] [Green Version]
- Campione, E.; Gaziano, R.; Doldo, E.; Marino, D.; Falconi, M.; Iacovelli, F.; Tagliaferri, D.; Pacello, L.; Bianchi, L.; Lanna, C.; et al. Antifungal Effect of All- trans Retinoic Acid against Aspergillus fumigatus In Vitro and in a Pulmonary Aspergillosis In Vivo Model. Antimicrob. Agents Chemother. 2021, 65, e01874-20. [Google Scholar] [CrossRef]
- Brunet, K.; Rammaert, B. Mucormycosis treatment: Recommendations, latest advances, and perspectives. J. Mycol. Med. 2020, 30, 101007. [Google Scholar] [CrossRef]
- Gebremariam, T.; Wiederhold, N.; Fothergill, A.W.; Garvey, E.P.; Hoekstra, W.J.; Schotzinger, R.J.; Patterson, T.F.; Filler, S.G.; Ibrahim, A.S. VT-1161 Protects Immunosuppressed Mice from Rhizopus arrhizus var. arrhizus Infection. Antimicrob. Agents Chemother. 2015, 59, 7815–7817. [Google Scholar] [CrossRef] [Green Version]
- Macedo, D.; Leonardelli, F.; Dudiuk, C.; Theill, L.; Cabeza, M.S.; Gamarra, S.; Garcia-Effron, G. Molecular Confirmation of the Linkage between the Rhizopus oryzae CYP51A Gene Coding Region and Its Intrinsic Voriconazole and Fluconazole Resistance. Antimicrob. Agents Chemother. 2018, 62, e00224-18. [Google Scholar] [CrossRef] [Green Version]
- Gebremariam, T.; Alkhazraji, S.; Alqarihi, A.; Wiederhold, N.P.; Shaw, K.J.; Patterson, T.F.; Filler, S.G.; Ibrahim, A.S. Fosmanogepix (APX001) Is Effective in the Treatment of Pulmonary Murine Mucormycosis Due to Rhizopus arrhizus. Antimicrob. Agents Chemother. 2020, 64, e00178-20. [Google Scholar] [CrossRef] [Green Version]
- Ben Yaakov, D.; Rivkin, A.; Mircus, G.; Albert, N.; Dietl, A.-M.; Kovalerchick, D.; Carmeli, S.; Haas, H.; Kontoyiannis, D.P.; Osherov, N. Identification and characterization of haemofungin, a novel antifungal compound that inhibits the final step of haem biosynthesis. J. Antimicrob. Chemother. 2016, 71, 946–952. [Google Scholar] [CrossRef] [Green Version]
- Chao, C.-M.; Lai, C.-C.; Yu, W.-L. COVID-19 associated mucormycosis–An emerging threat. J. Microbiol. Immunol. Infect. 2022, 55, 183–190. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Carnero, L.C.; Mora-Montes, H.M. Mucormycosis and COVID-19-Associated Mucormycosis: Insights of a Deadly but Neglected Mycosis. J. Fungi 2022, 8, 445. https://doi.org/10.3390/jof8050445
García-Carnero LC, Mora-Montes HM. Mucormycosis and COVID-19-Associated Mucormycosis: Insights of a Deadly but Neglected Mycosis. Journal of Fungi. 2022; 8(5):445. https://doi.org/10.3390/jof8050445
Chicago/Turabian StyleGarcía-Carnero, Laura C., and Héctor M. Mora-Montes. 2022. "Mucormycosis and COVID-19-Associated Mucormycosis: Insights of a Deadly but Neglected Mycosis" Journal of Fungi 8, no. 5: 445. https://doi.org/10.3390/jof8050445