
The Monothiol Glutaredoxin Grx4 Influences Iron Homeostasis and Virulence in Ustilago maydis
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions
2.2. Construction of Pcrg1::grx4 in Strains FB1 and FB2
2.3. Grx4 Depletion
2.4. Low Iron Minimal Medium (MM)
2.5. Cell Size Measurements
2.6. RNA Isolation, RNA-Seq Analysis, and Quantitative PCR
2.7. Siderophore Measurement
2.8. Assays for Cellular Pigmentation and Extracellular Melanin
2.9. Virulence
2.10. Data Availability, Genomic and Bioinformatic Resources
2.11. Statistics
3. Results
3.1. Identification of the Grx4 Monothiol Glutaredoxin in U. maydis
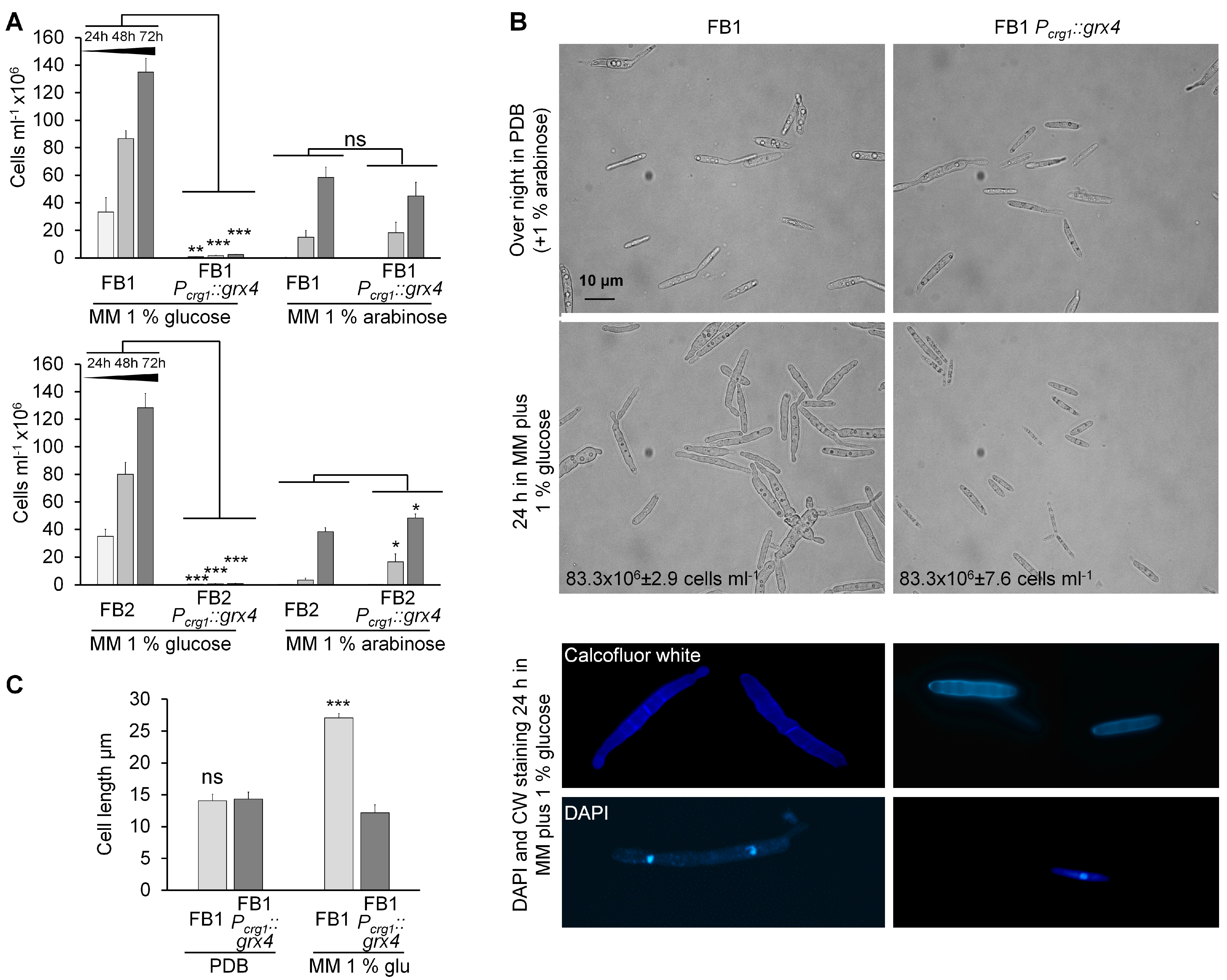
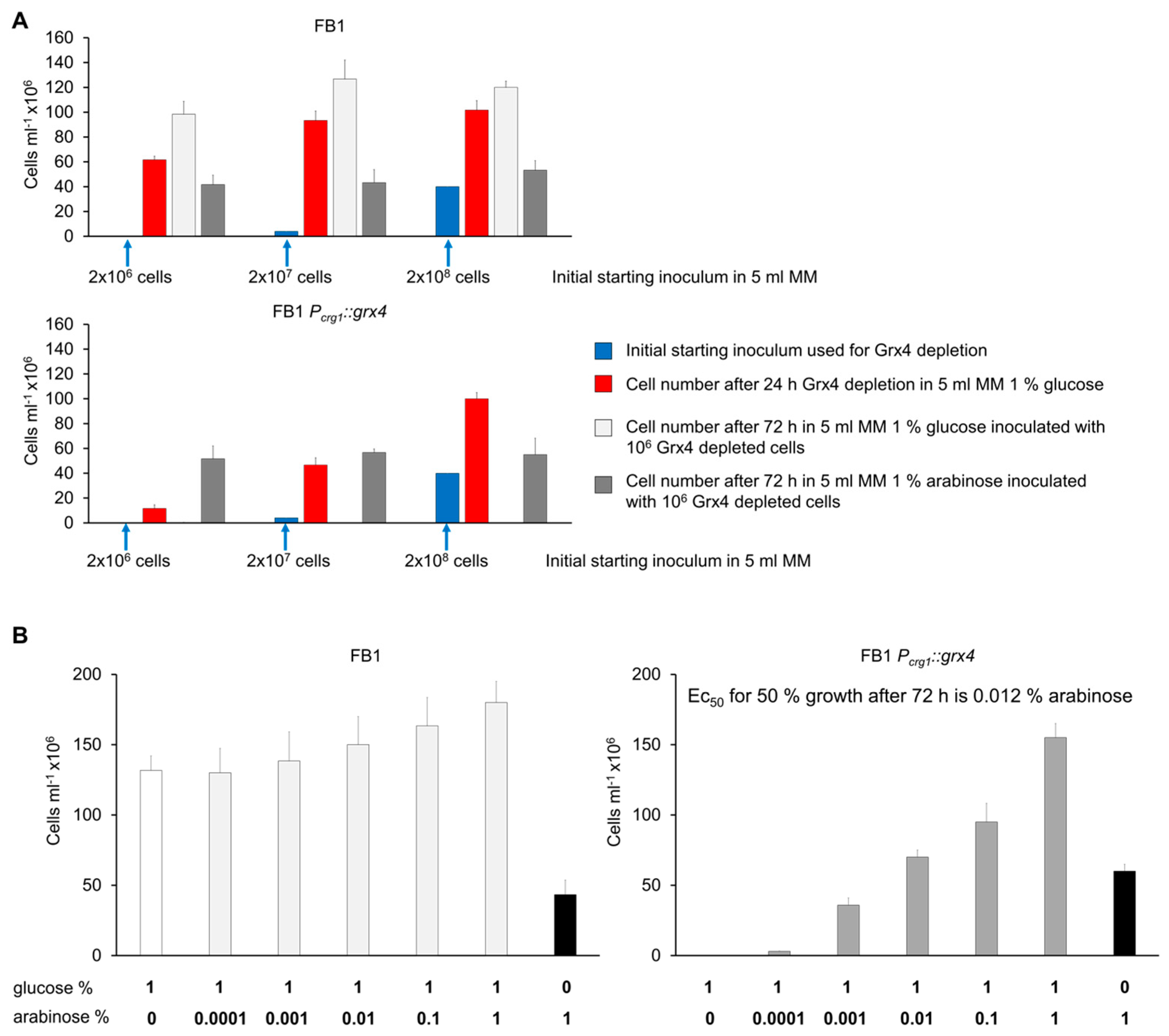
3.2. Growth of Strains with Conditional Grx4 Expression
3.3. Identification of Grx4-Regulated Gene Functions
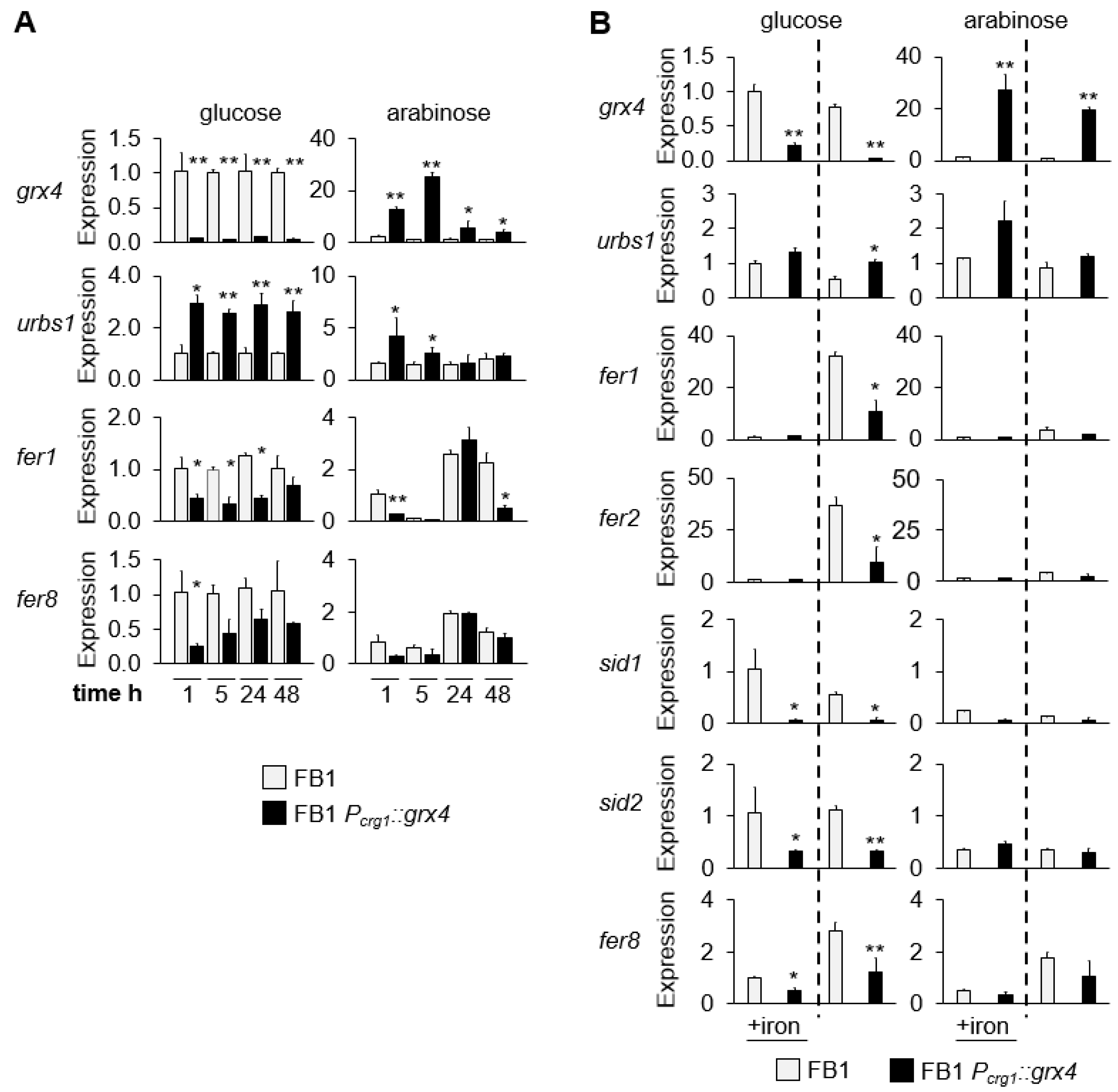
3.4. Expression of Grx4 and Iron-Regulated Genes at Different Timepoints and in Low and High Iron
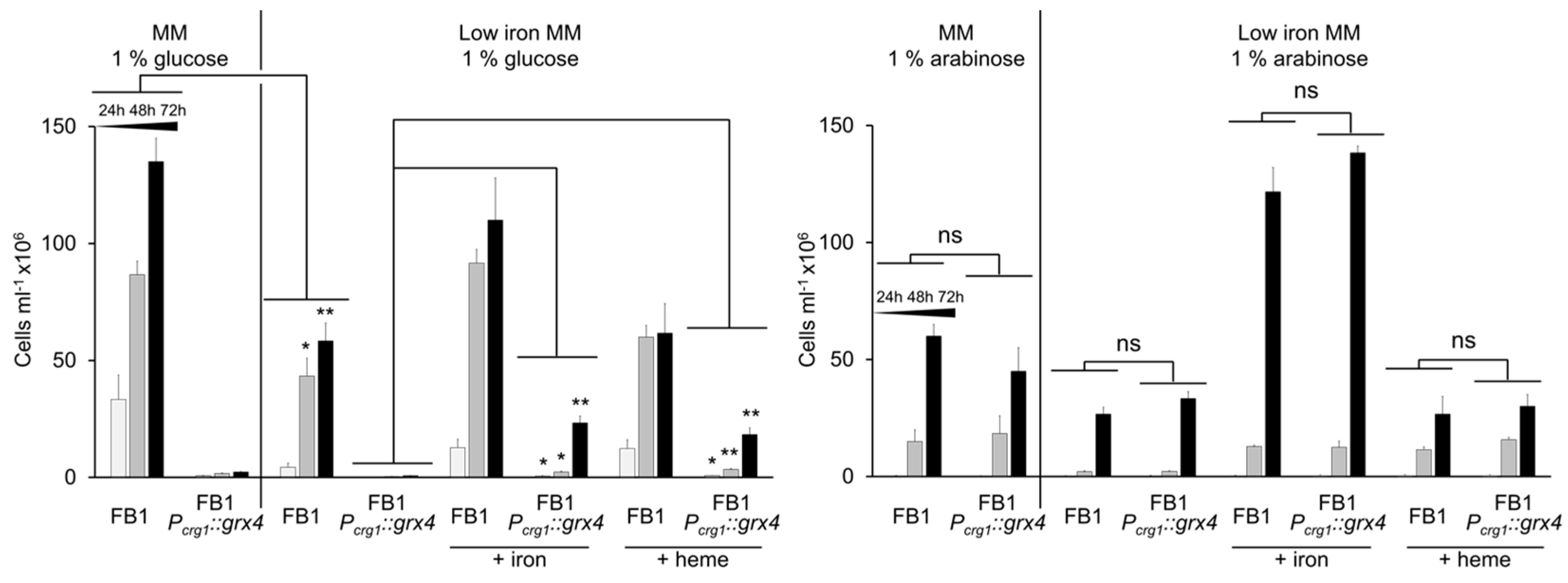
3.5. Addition of Iron Partially Restores Growth in the Context of Grx4 Depletion
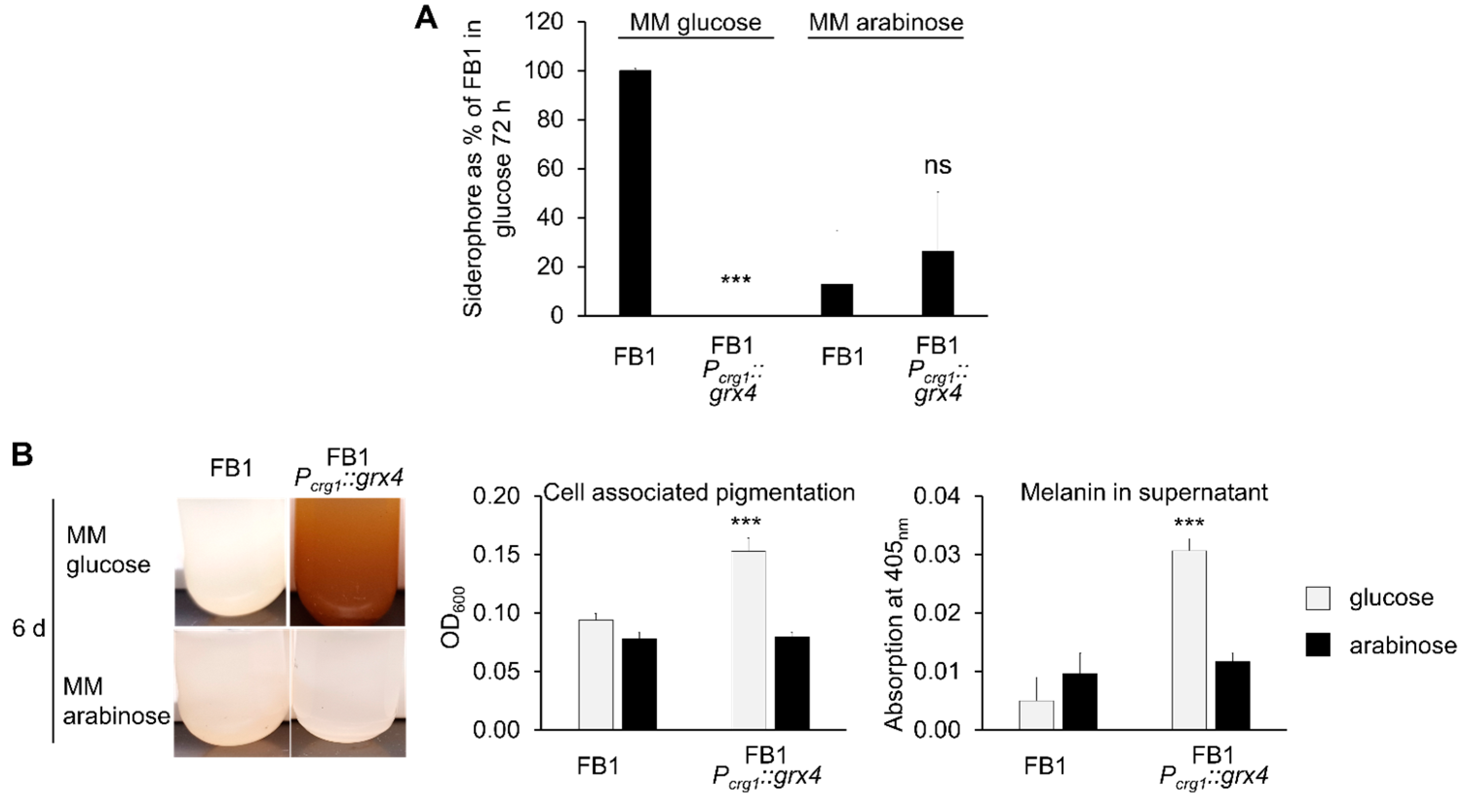
3.6. Siderophore Formation Is Regulated by Grx4
3.7. Pigment/Melanin Formation Is Repressed by Grx4
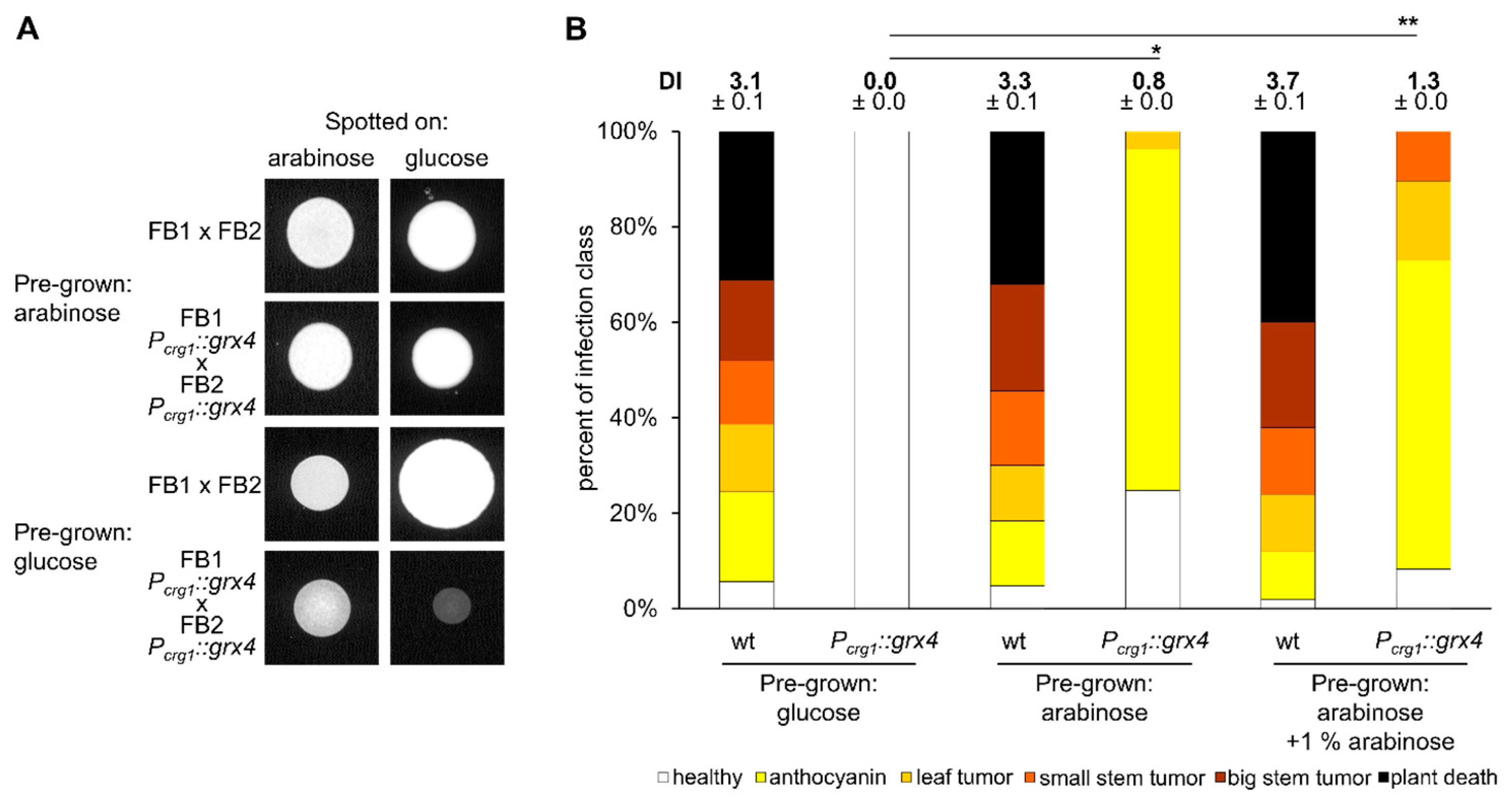
3.8. Appropriate Grx4 Levels Are Important for Mating
3.9. Grx4 Is Needed for Virulence and Pathogenic Development
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holliday, R. Ustilago maydis. In Handbook of Genetics; King, R.C., Ed.; Plenum: New York, NY, USA, 1974; Volume 1, pp. 575–595. [Google Scholar]
- Zuo, W.; Ökmen, B.; Depotter, J.R.L.; Ebert, M.K.; Redkar, A.; Misas Villamil, J.; Doehlemann, G. Molecular Interactions Between Smut Fungi and Their Host Plants. Annu. Rev. Phytopathol. 2019, 57, 411–430. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 Fungal Pathogens in Molecular Plant Pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Kämper, J.; Kahmann, R.; Bölker, M.; Ma, L.J.; Brefort, T.; Saville, B.J.; Banuett, F.; Kronstad, J.W.; Gold, S.E.; Müller, O.; et al. Insights from the Genome of the Biotrophic Fungal Plant Pathogen Ustilago maydis. Nature 2006, 444, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, M.; Damoo, D.; Sun, S.; Lee, C.W.J.; Croll, D.; Brumer, H.; Kronstad, J. Organic Acids and Glucose Prime Late-Stage Fungal Biotrophy in Maize. Science 2022, 376, 1187–1191. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Outten, C.E. Iron-Sulfur Cluster Signaling: The Common Thread in Fungal Iron Tegulation. Curr. Opin. Chem. Biol. 2020, 55, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Pujol-Carrion, N.; Belli, G.; Herrero, E.; Nogues, A.; de la Torre-Ruiz, M.A. Glutaredoxins Grx3 and Grx4 Regulate Nuclear Localisation of Aft1 and the Oxidative Stress Response in Saccharomyces cerevisiae. J. Cell Sci. 2006, 119, 4554–4564. [Google Scholar] [CrossRef]
- Jbel, M.; Mercier, A.; Labbé, S. Grx4 Monothiol Glutaredoxin is Required for Iron Limitation-Dependent Inhibition of Fep1. Euk. Cell 2011, 10, 629–645. [Google Scholar] [CrossRef]
- Attarian, R.; Hu, G.; Sánchez-León, E.; Caza, M.; Croll, D.; Do, E.; Bach, H.; Missall, T.; Lodge, J.; Jung, W.H.; et al. The Monothiol Glutaredoxin Grx4 Regulates Iron Homeostasis and Virulence in Cryptococcus neoformans. mBio 2018, 9, e02377-18. [Google Scholar] [CrossRef]
- Misslinger, M.; Scheven, M.T.; Hortschansky, P.; López-Berges, M.S.; Heiss, K.; Beckmann, N.; Heigl, T.; Hermann, M.; Krüger, T.; Kniemeyer, O.; et al. The Monothiol Glutaredoxin GrxD is Essential for Sensing Iron Starvation in Aspergillus fumigatus. PLoS Genet. 2019, 15, e1008379. [Google Scholar] [CrossRef]
- Hu, G.; Horianopoulos, L.; Sánchez-León, E.; Caza, M.; Jung, W.; Kronstad, J.W. The Monothiol Glutaredoxin Grx4 Influences Thermotolerance, Cell Wall Integrity, and Mpk1 Signaling in Cryptococcus neoformans. G3 2021, 11, jkab322. [Google Scholar] [CrossRef]
- Mei, B.; Budde, A.D.; Leong, S.A. sid1, a Gene Initiating Siderophore Biosynthesis in Ustilago maydis: Molecular Characterization, Regulation by Iron, and Role in Phytopathogenicity. Proc. Natl. Acad. Sci. USA 1993, 90, 903–907. [Google Scholar] [CrossRef] [PubMed]
- Eichhorn, H.; Lessing, F.; Winterberg, B.; Schirawski, J.; Kämper, J.; Müller, P.; Kahmann, R. A Ferroxidation/Permeation Iron Uptake System is Required for Virulence in Ustilago maydis. Plant Cell 2006, 18, 3332–3345. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.M.; Gentil, G.D.; Budde, A.D.; Leong, S.A. Characterization of the Ustilago maydis sid2 Gene, Encoding a Multidomain Peptide Synthetase in the Ferrichrome Biosynthetic Gene Cluster. J. Bacteriol. 2001, 183, 4040–4051. [Google Scholar] [CrossRef] [PubMed]
- Winterberg, B.; Uhlmann, S.; Linne, U.; Lessing, F.; Marahiel, M.A.; Eichhorn, H.; Kahmann, R.; Schirawski, J. Elucidation of the Complete Ferrichrome A Biosynthetic Pathway in Ustilago maydis. Mol. Microbiol. 2010, 75, 1260–1271. [Google Scholar] [CrossRef] [PubMed]
- Bottin, A.; Kämper, J.; Kahmann, R. Isolation of a Carbon Source-Regulated Gene from Ustilago maydis. Mol. Gen. Genet. 1996, 253, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wen, A.; Shen, B.; Lu, J.; Huang, Y.; Chang, Y. FastCloning: A Highly Simplified, Purification-Free, Sequence- and Ligation-Independent PCR Cloning Method. BMC Biotechnol. 2011, 11, 92. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Holden, D.W.; Leong, S.A. Gene Transfer System for the Phytopathogenic Fungus Ustilago maydis. Proc. Natl. Acad. Sci. USA 1988, 85, 865–869. [Google Scholar] [CrossRef]
- McCotter, S.W. Grx4: A Key Regulator of Secondary Metabolism, Nitrogen Uptake, and Iron Homeostasis in the Corn Smut Fungus, Ustilago maydis. Ph.D. Thesis, University of British Columbia, Vancouver, BC, Canada, 2022. [Google Scholar] [CrossRef]
- Kretschmer, M.; Lambie, S.; Croll, D.; Kronstad, J.W. Acetate Provokes Mitochondrial Stress and Cell Death in Ustilago maydis. Mol. Microbiol. 2018, 107, 488–507. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal Chemical Assay for the Detection and Determination of Siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Mayer, F.L.; Kronstad, J.W. Disarming Fungal Pathogens: Bacillus safensis Inhibits Virulence Factor Production and Biofilm Formation by Cryptococcus neoformans and Candida albicans. mBio 2017, 8, e01537-17. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, M.; Klose, J.; Kronstad, J.W. Defects in Mitochondrial and Peroxisomal β-Oxidation Influence Virulence in the Maize Pathogen Ustilago maydis. Eukaryot. Cell 2012, 11, 1055–1066. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, M.; Croll, D.; Kronstad, J.W. Maize Susceptibility to Ustilago maydis is Influenced by Genetic and Chemical Perturbation of Carbohydrate Allocation. Mol. Plant Pathol. 2017, 18, 1222–1237. [Google Scholar] [CrossRef] [PubMed]
- Luikenhuis, S.; Perrone, G.; Dawes, I.W.; Grant, C.M. The Yeast Saccharomyces cerevisiae Contains Two Glutaredoxin Genes that are Required for Protection Against Reactive Oxygen Species. Mol. Biol. Cell 1998, 9, 1081–1091. [Google Scholar] [CrossRef] [PubMed]
- Ueta, R.; Fujiwara, N.; Iwai, K.; Yamaguchi-Iwai, Y. Iron-Induced Dissociation of the Aft1p Transcriptional Regulator from Target Gene Promoters is an Initial Event in Iron-Dependent Gene Suppression. Mol. Biol. Cell 2012, 32, 4998–5008. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Manzaneque, M.T.; Ros, J.; Cabiscol, E.; Sorribas, A.; Herrero, E. Grx5 Glutaredoxin Plays a Central Role in Protection Against Protein Oxidative Damage in Saccharomyces cerevisiae. Mol. Biol. Cell 1999, 19, 8180–8190. [Google Scholar] [CrossRef]
- Rodríguez-Manzaneque, M.T.; Tamarit, J.; Bellí, G.; Ros, J.; Herrero, E. Grx5 is a Mitochondrial Glutaredoxin Required for the Activity of Iron/Sulfur Enzymes. Mol. Biol. Cell 2002, 13, 1109–1121. [Google Scholar] [CrossRef]
- Gonzalez Porqué, P.; Baldesten, A.; Reichard, P. The Involvement of the Thioredoxin System in the Reduction of Methionine Sulfoxide and Sulfate. J. Biol. Chem. 1970, 245, 2371–2374. [Google Scholar] [CrossRef]
- Trotter, E.W.; Grant, C.M. Thioredoxins are Required for Protection Against a Reductive Stress in the Yeast Saccharomyces cerevisiae. Mol. Microbiol. 2002, 46, 869–878. [Google Scholar] [CrossRef]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A Web Server for Functional Enrichment Analysis and Conversions of Gene Lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Nanjaraj Urs, A.N.; Dancis, A.; Zhang, Y. Genetic Suppressors of Δgrx3 Δgrx4, Lacking Redundant Multidomain Monothiol Yeast Glutaredoxins, Rescue Growth and Iron Homeostasis. Biosci. Rep. 2022, 42, BSR20212665. [Google Scholar] [CrossRef] [PubMed]
- Yap, A.; Volz, R.; Paul, S.; Moye-Rowley, W.S.; Haas, H. Regulation of High-Affinity Iron Acquisition, Including Acquisition Mediated by the Iron Permease FtrA, Is Coordinated by AtrR, SrbA, and SreA in Aspergillus fumigatus. mBio 2023, 14, e0075723. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Dong, Y.; Yu, Q.; Kai, Z.; Zhang, M.; Jia, C.; Xiao, C.; Zhang, B.; Zhang, B.; Li, M. Function of Glutaredoxin 3 (Grx3) in Oxidative Stress Response Caused by Iron Homeostasis Disorder in Candida albicans. Future Microbiol. 2017, 12, 1397–1412. [Google Scholar] [CrossRef]
- Pujol-Carrion, N.; Torre-Ruiz, M.A. Physical Interaction between the MAPK Slt2 of the PKC1-MAPK Pathway and Grx3/Grx4 Glutaredoxins is Required for the Oxidative Stress Response in Budding Yeast. Free Radic. Biol. Med. 2017, 103, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Alkafeef, S.S.; Lane, S.; Yu, C.; Zhou, T.; Solis, N.V.; Filler, S.G.; Huang, L.; Liu, H. Proteomic Profiling of the Monothiol Glutaredoxin Grx3 Reveals its Global Role in the Regulation of Iron Dependent Processes. PLoS Genet. 2020, 16, e1008881. [Google Scholar] [CrossRef] [PubMed]
- Mühlenhoff, U.; Molik, S.; Godoy, J.R.; Uzarska, M.A.; Richter, N.; Seubert, A.; Zhang, Y.; Stubbe, J.; Pierrel, F.; Herrero, E.; et al. Cytosolic Monothiol Glutaredoxins Function in Intracellular Iron Sensing and Trafficking via Their Bound Iron-Sulfur Cluster. Cell Metabol. 2010, 12, 373–385. [Google Scholar] [CrossRef]
- Banuett, F.; Herskowitz, I. Different a Alleles of Ustilago maydis are Necessary for Maintenance of Filamentous Growth but not for Meiosis. Proc. Natl. Acad. Sci. USA 1989, 86, 5878–5882. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McCotter, S.W.; Kretschmer, M.; Lee, C.W.J.; Heimel, K.; Kronstad, J.W. The Monothiol Glutaredoxin Grx4 Influences Iron Homeostasis and Virulence in Ustilago maydis. J. Fungi 2023, 9, 1112. https://doi.org/10.3390/jof9111112
McCotter SW, Kretschmer M, Lee CWJ, Heimel K, Kronstad JW. The Monothiol Glutaredoxin Grx4 Influences Iron Homeostasis and Virulence in Ustilago maydis. Journal of Fungi. 2023; 9(11):1112. https://doi.org/10.3390/jof9111112
Chicago/Turabian StyleMcCotter, Sean W., Matthias Kretschmer, Christopher W. J. Lee, Kai Heimel, and James W. Kronstad. 2023. "The Monothiol Glutaredoxin Grx4 Influences Iron Homeostasis and Virulence in Ustilago maydis" Journal of Fungi 9, no. 11: 1112. https://doi.org/10.3390/jof9111112