
The Plant Defensin Ppdef1 Is a Novel Topical Treatment for Onychomycosis
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequence Alignment
2.2. Strains and Culture Conditions
2.3. Recombinant Expression of Ppdef1 Using Pichia pastoris
2.4. NMR
2.5. Structural Analysis
2.6. Antifungal Activity Assays
2.7. Propidium Iodide Uptake in Response to Antifungal Treatment
2.8. Antifungal Activity in the Presence of Keratin
2.9. Cell Killing of T. rubrum Growing on Nail Fragments
2.10. Nail Penetration Assays
2.11. Infected Nail Model
2.12. Statistical Analysis
3. Results
3.1. Ppdef1 Is a Member of the Plant Defenisn Family
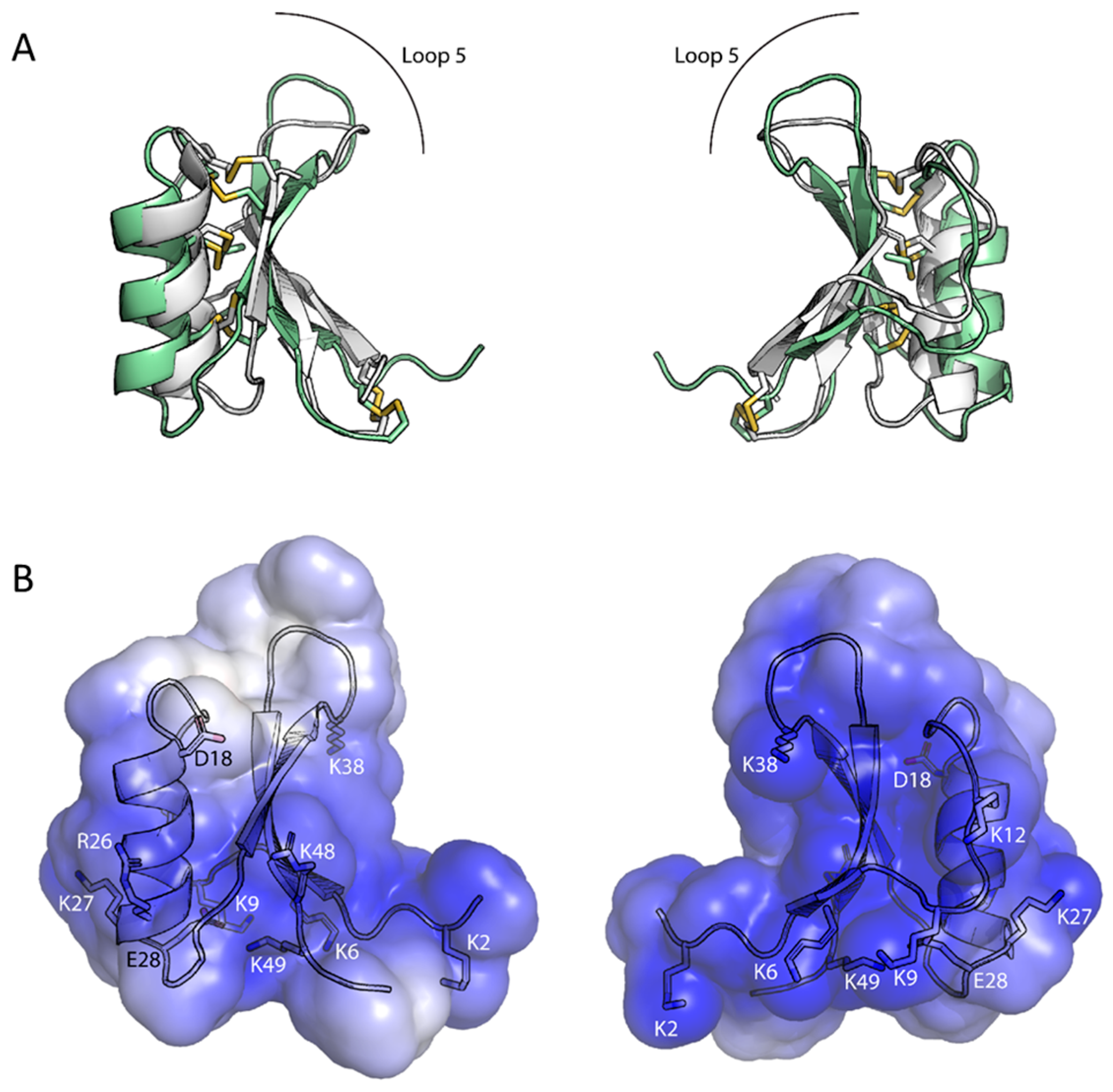
3.2. Ppdef1 Forms the Conserved Cysteine Stabilised αβ Motif Typical of Plant Defensins
3.3. Ppdef1 Is Active against a Broad Range of Human Fungal Pathogens That Cause Fungal Nail Infections
3.4. Ppdef1 Kills C. albicans Cells within 30 min
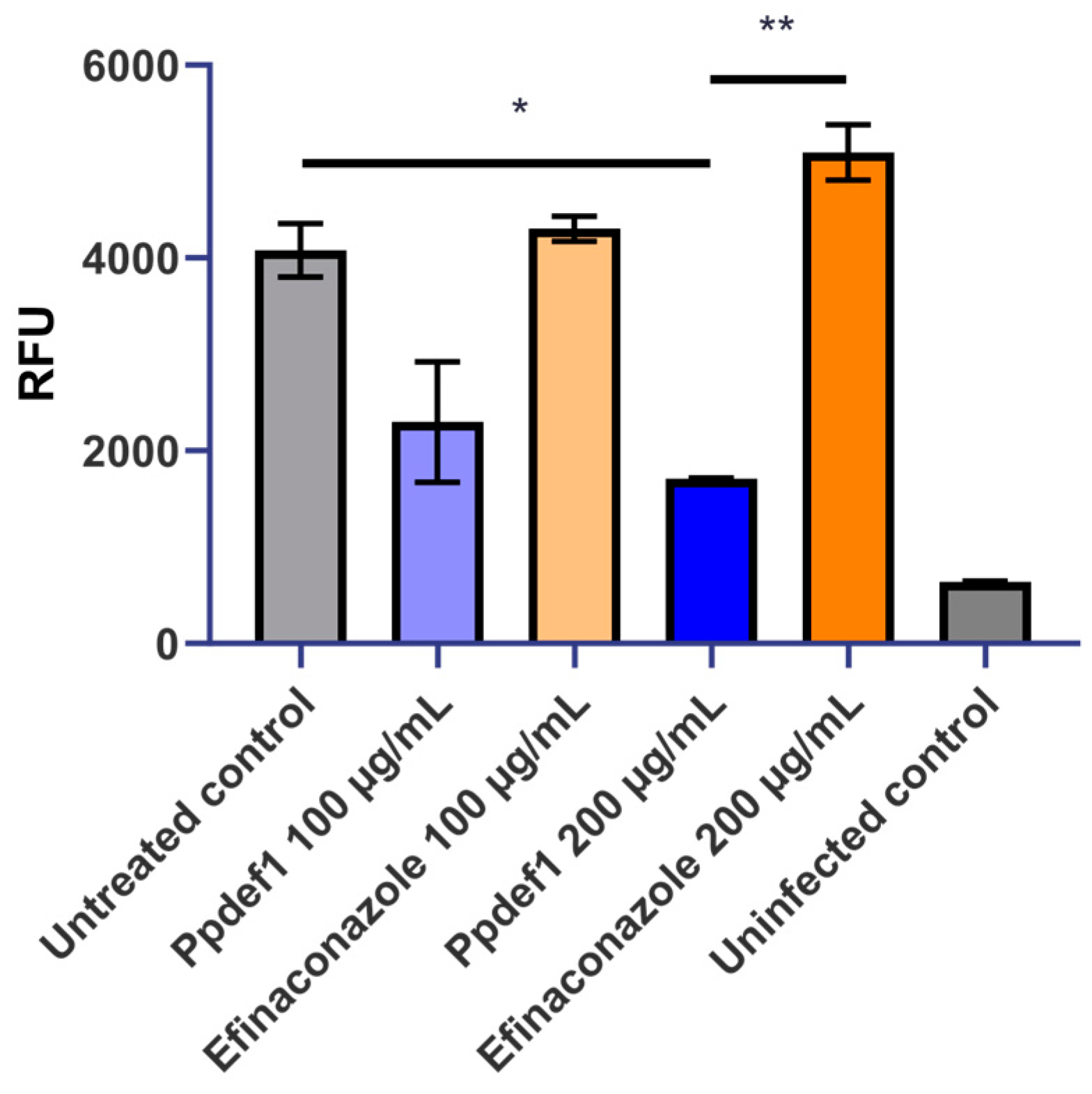
3.5. Ppdef1 Retains Activity against T. rubrum in the Presence of Keratin
3.6. Ppdef1 Kills T. rubrum Growing on Keratin as a Sole Nutrient Source
3.7. Ppdef1 Penetrates Human Nail Fragments
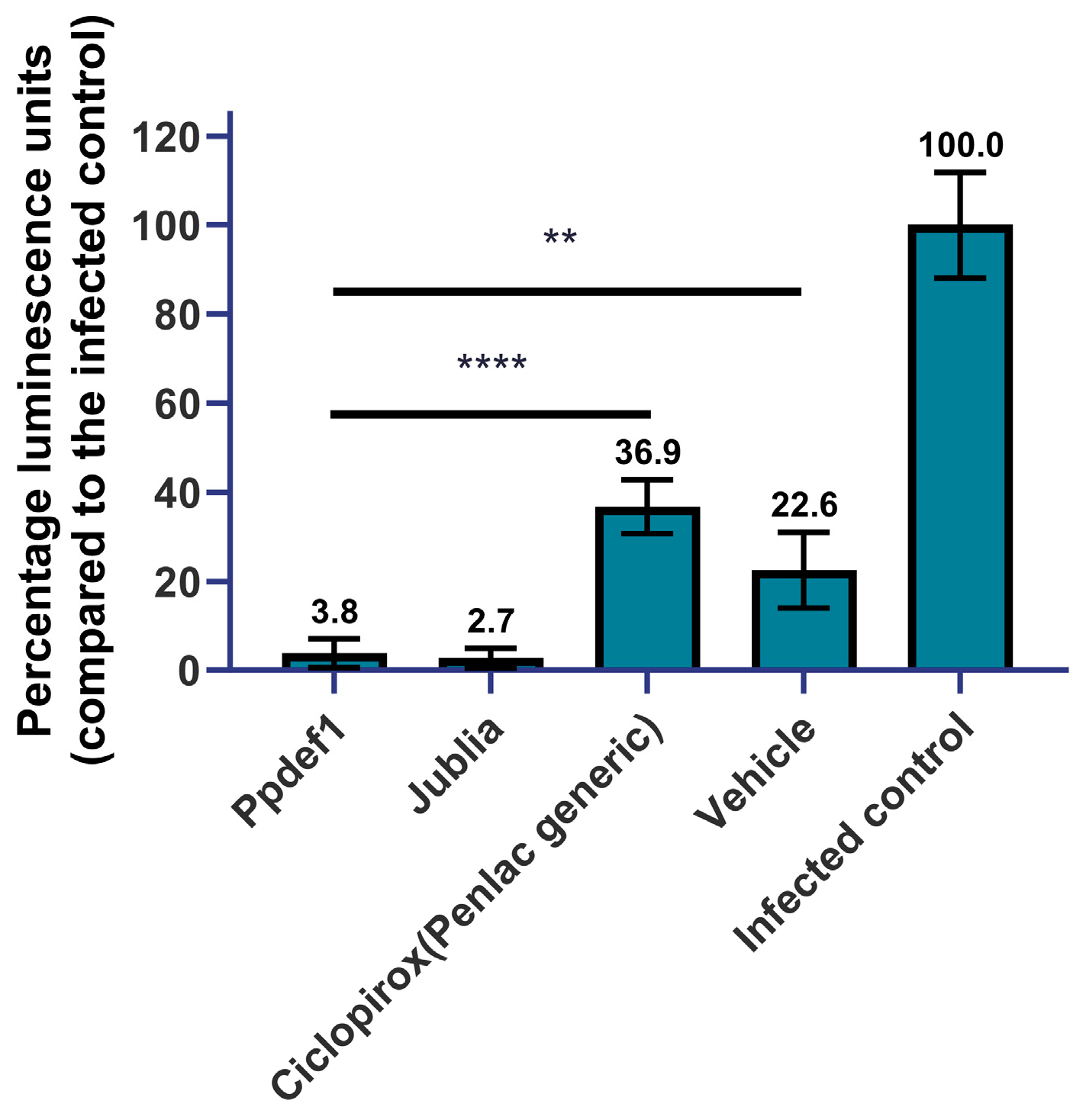
3.8. Infected Nail Model
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cañete-Gibas, C.; Wiederhold, N.P. Mycology of Onychomycosis. Clin. Microbiol. Newsl. 2023, 45, 11–17. [Google Scholar] [CrossRef]
- Gupta, A.K.; Polla Ravi, S.; Choi, S.Y.; Konda, A.; Cooper, E.A. Strategies for the enhancement of nail plate permeation of drugs to treat onychomycosis. J. Eur. Acad. Dermatol. Venereol. 2023, 37, 243–255. [Google Scholar] [CrossRef]
- Maskan Bermudez, N.; Rodríguez-Tamez, G.; Perez, S.; Tosti, A. Onychomycosis: Old and New. J. Fungi 2023, 9, 559. [Google Scholar] [CrossRef]
- Gupta, A.K.; Fleckman, P.; Baran, R. Ciclopirox nail lacquer topical solution 8% in the treatment of toenail onychomycosis. J. Am. Acad. Dermatol. 2000, 43, S70–S80. [Google Scholar] [CrossRef]
- Elewski, B.E.; Rich, P.; Pollak, R.; Pariser, D.M.; Watanabe, S.; Senda, H.; Ieda, C.; Smith, K.; Pillai, R.; Ramakrishna, T.; et al. Efinaconazole 10% solution in the treatment of toenail onychomycosis: Two phase III multicenter, randomized, double-blind studies. J. Am. Acad. Dermatol. 2013, 68, 600–608. [Google Scholar] [CrossRef]
- Elewski, B.E.; Aly, R.; Baldwin, S.L.; Gonzalez Soto, R.F.; Rich, P.; Weisfeld, M.; Wiltz, H.; Zane, L.T.; Pollak, R. Efficacy and safety of tavaborole topical solution, 5%, a novel boron-based antifungal agent, for the treatment of toenail onychomycosis: Results from 2 randomized phase-III studies. J. Am. Acad. Dermatol. 2015, 73, 62–69. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Miyamoto, M.; Sugibayashi, K.; Morimoto, Y. Drug permeation through the three layers of the human nail plate. J. Pharm. Pharmacol. 1999, 51, 271–278. [Google Scholar] [CrossRef]
- McAuley, W.; Jones, S.; Traynor, M.; Guesné, S.; Murdan, S.; Brown, M. An investigation of how fungal infection influences drug penetration through onychomycosis patient’s nail plates. Eur. J. Pharm. Biopharm. 2016, 102, 178–184. [Google Scholar] [CrossRef]
- Mertin, D.; Lippold, B.C. In-vitro permeability of the human nail and of a keratin membrane from bovine hooves: Influence of the partition coefficient octanol/water and the water solubility of drugs on their permeability and maximum flux. J. Pharm. Pharmacol. 1997, 49, 30–34. [Google Scholar]
- Darkes, M.J.M.; Scott, L.J.; Goa, K.L. Terbinafine. Am. J. Clin. Dermatol. 2003, 4, 39–65. [Google Scholar] [CrossRef]
- Rosen, T.; Friedlander, S.F.; Kircik, L.; Zirwas, M.J.; Stein, L.G.; Bhatia, N.; Gupta, A.K. Onychomycosis: Epidemiology, diagnosis, and treatment in a changing landscape. J. Drugs Dermatol. JDD 2015, 14, 223–233. [Google Scholar]
- Parisi, K.; Shafee, T.M.A.; Quimbar, P.; van der Weerden, N.L.; Bleackley, M.R.; Anderson, M.A. The evolution, function and mechanisms of action for plant defensins. Semin. Cell Dev. Biol. 2018, 88, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Gelinski, J.M.L.N.; de Melo Franco, B.D.G.; Fonseca, G.G. Plant-derived antimicrobial peptides. In Antimicrobial Peptides; Elsevier: London, UK; Academic Press: Cambridge, MA, USA, 2023; pp. 157–169. [Google Scholar] [CrossRef]
- Montesinos, E. Functional Peptides for Plant Disease Control. Annu. Rev. Phytopathol. 2023, 61, 301–324. [Google Scholar] [CrossRef] [PubMed]
- Mookherjee, N.; Anderson, M.A.; Haagsman, H.P.; Davidson, D.J. Antimicrobial host defence peptides: Functions and clinical potential. Nat. Rev. Drug Discov. 2020, 19, 311–332. [Google Scholar] [PubMed]
- Lay, F.T.; Brugliera, F.; Anderson, M.A. Isolation and properties of floral defensins from ornamental tobacco and petunia. Plant Physiol. 2003, 131, 1283–1293. [Google Scholar] [CrossRef]
- Kovaleva, V.; Bukhteeva, I.; Kit, O.Y.; Nesmelova, I.V. Plant defensins from a structural perspective. Int. J. Mol. Sci. 2020, 21, 5307. [Google Scholar] [CrossRef]
- Shafee, T.; Anderson, M.A. A quantitative map of protein sequence space for the cis-defensin superfamily. Bioinformatics 2018, 35, 743–752. [Google Scholar] [CrossRef]
- Anderson, N.L.V.D.W.M.A. A method of treatment of fungal infections. Patent number EP3209319B1, 2015. [Google Scholar]
- Parisi, K.; McKenna, J.A.; Lowe, R.G.T.; Harris, K.S.; Thomas, S.; Guarino, R.; Lee, E.; van der Weerden, N.L.; Bleackley, M.R.; Anderson, M.A. Hyperpolarisation of mitochondrial membranes is a critical component of the antifungal mechanism of the plant defensin Ppdef1. J. Fungi 2023. [Google Scholar]
- Chang, T.; Schroder, L.A.; Thomson, J.M.; Klocman, A.S.; Tomasini, A.J.; Stromhaug, P.E.; Dunn, W.A., Jr. PpATG9 encodes a novel membrane protein that traffics to vacuolar membranes, which sequester peroxisomes during pexophagy in Pichia pastoris. Mol. Biol. Cell 2005, 16, 4941–4953. [Google Scholar] [CrossRef]
- Lay, F.T.; Mills, G.D.; Poon, I.K.; Cowieson, N.P.; Kirby, N.; Baxter, A.A.; van der Weerden, N.L.; Dogovski, C.; Perugini, M.A.; Anderson, M.A.; et al. Dimerization of plant defensin NaD1 enhances its antifungal activity. J. Biol. Chem. 2012, 287, 19961–19972. [Google Scholar] [CrossRef]
- Dolinsky, T.J.; Nielsen, J.E.; McCammon, J.A.; Baker, N.A. PDB2PQR: An automated pipeline for the setup of Poisson–Boltzmann electrostatics calculations. Nucleic Acids Res. 2004, 32, W665–W667. [Google Scholar] [CrossRef]
- Baker, N.A.; Sept, D.; Joseph, S.; Holst, M.J.; McCammon, J.A. Electrostatics of nanosystems: Application to microtubules and the ribosome. Proc. Natl. Acad. Sci. USA 2001, 98, 10037–10041. [Google Scholar] [CrossRef]
- Schrodinger, L.L.C. The PyMOL Molecular Graphics System, Version 2.1.0. 2018. Available online: https://pymol.org/2/ (accessed on 16 October 2023).
- Broekaert, W.; Terras, F.; Cammue, B.; Vanderleyden, J. An automated quantitative assay for fungal growth inhibition. FEMS Microbiol. Lett. 1990, 69, 55–60. [Google Scholar] [CrossRef]
- Morace, G.; Drago, M.; Scaltrito, M.M.; Conti, S.; Fanti, F.; Polonelli, L.; Group, G.P. In vitro activity (MIC and MFC) of voriconazole, amphotericin B, and itraconazole against 192 filamentous fungi: The GISIA-2 study. J. Chemother. 2007, 19, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Hui, X.; Shainhouse, Z.; Tanojo, H.; Anigbogu, A.; Markus, G.E.; Maibach, H.I.; Wester, R.C. Enhanced human nail drug delivery: Nail inner drug content assayed by new unique method. J. Pharm. Sci. 2002, 91, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Stensen, W.; Turner, R.; Brown, M.; Kondori, N.; Svendsen, J.S.; Svenson, J. Short Cationic Antimicrobial Peptides Display Superior Antifungal Activities toward Candidiasis and Onychomycosis in Comparison with Terbinafine and Amorolfine. Mol. Pharm. 2016, 13, 3595–3600. [Google Scholar] [CrossRef] [PubMed]
- Traynor, M.; Turner, R.; Evans, C.; Khengar, R.; Jones, S.; Brown, M. Effect of a novel penetration enhancer on the ungual permeation of two antifungal agents. J. Pharm. Pharmacol. 2010, 62, 730–737. [Google Scholar] [CrossRef]
- Kerenga, B.K.; McKenna, J.A.; Harvey, P.J.; Quimbar, P.; Garcia, D.; Lay, F.T.; Phan, T.K.; Veneer, P.K.; Vasa, S.; Parisi, K. Salt-tolerant antifungal and antibacterial activities of the corn defensin ZmD32. Front. Microbiol. 2019, 10, 795. [Google Scholar] [CrossRef]
- Sagaram, U.S.; El-Mounadi, K.; Buchko, G.W.; Berg, H.R.; Kaur, J.; Pandurangi, R.S.; Smith, T.J.; Shah, D.M. Structural and functional studies of a phosphatidic acid-binding antifungal plant defensin MtDef4: Identification of an RGFRRR motif governing fungal cell entry. PLoS ONE 2013, 8, e82485. [Google Scholar] [CrossRef]
- Poon, I.; Baxter, A.A.; Lay, F.T.; Mills, G.D.; Adda, C.G.; Payne, J.A.; Phan, T.K.; Ryan, G.F.; White, J.A.; Veneer, P.K.; et al. Phosphoinositide-mediated oligomerization of a defensin induces cell lysis. Elife 2014, 3, e01808. [Google Scholar] [CrossRef]
- Järvå, M.; Lay, F.T.; Phan, T.K.; Humble, C.; Poon, I.K.H.; Bleackley, M.R.; Anderson, M.A.; Hulett, M.D.; Kvansakul, M. X-ray structure of a carpet-like antimicrobial defensin–phospholipid membrane disruption complex. Nat. Commun. 2018, 9, 1962. [Google Scholar] [CrossRef] [PubMed]
- Baxter, A.A.; Richter, V.; Lay, F.T.; Poon, I.K.; Adda, C.G.; Veneer, P.K.; Phan, T.K.; Bleackley, M.R.; Anderson, M.A.; Kvansakul, M. The tomato defensin TPP3 binds phosphatidylinositol (4, 5)-bisphosphate via a conserved dimeric cationic grip conformation to mediate cell lysis. Mol. Cell. Biol. 2015, 35, 1964–1978. [Google Scholar] [CrossRef]
- Tatsumi, Y.; Nagashima, M.; Shibanushi, T.; Iwata, A.; Kangawa, Y.; Inui, F.; Siu, W.J.J.; Pillai, R.; Nishiyama, Y. Mechanism of action of efinaconazole, a novel triazole antifungal agent. Antimicrob. Agents Chemother. 2013, 57, 2405–2409. [Google Scholar] [CrossRef] [PubMed]
- Manavathu, E.K.; Cutright, J.L.; Chandrasekar, P.H. Organism-dependent fungicidal activities of azoles. Antimicrob. Agents Chemother. 1998, 42, 3018–3021. [Google Scholar] [CrossRef] [PubMed]
- Cowen, L.E.; Sanglard, D.; Howard, S.J.; Rogers, P.D.; Perlin, D.S. Mechanisms of antifungal drug resistance. Cold Spring Harb. Perspect. Med. 2015, 5, a019752. [Google Scholar] [CrossRef] [PubMed]
- Ryder, N. The mechanism of action of terbinafine. Clin. Exp. Dermatol. 1989, 14, 98–100. [Google Scholar] [CrossRef]
- Rock, F.L.; Mao, W.; Yaremchuk, A.; Tukalo, M.; Crépin, T.; Zhou, H.; Zhang, Y.-K.; Hernandez, V.; Akama, T.; Baker, S.J. An antifungal agent inhibits an aminoacyl-tRNA synthetase by trapping tRNA in the editing site. Science 2007, 316, 1759–1761. [Google Scholar] [CrossRef]
- Jue, S.; Dawson, G.; Brogden, R. Ciclopirox olamine 1% cream. Drugs 1985, 29, 330–341. [Google Scholar] [CrossRef]
- Bleackley, M.R.; Payne, J.A.; Hayes, B.M.; Durek, T.; Craik, D.J.; Shafee, T.M.; Poon, I.K.; Hulett, M.D.; Van Der Weerden, N.L.; Anderson, M.A. Nicotiana alata defensin chimeras reveal differences in the mechanism of fungal and tumor cell killing and an enhanced antifungal variant. Antimicrob. Agents Chemother. 2016, 60, 6302–6312. [Google Scholar] [CrossRef]
- Vriens, K.; Cammue, B.P.; Thevissen, K. Antifungal plant defensins: Mechanisms of action and production. Molecules 2014, 19, 12280–12303. [Google Scholar] [CrossRef]
- McColl, A.I.; Bleackley, M.R.; Anderson, M.A.; Lowe, R.G.T. Resistance to the Plant Defensin NaD1 Features Modifications to the Cell Wall and Osmo-Regulation Pathways of Yeast. Front. Microbiol. 2018, 9, 1648. [Google Scholar] [CrossRef] [PubMed]
- Mercer, D.K.; Stewart, C.S. Keratin hydrolysis by dermatophytes. Med. Mycol. 2018, 57, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, H.; Kumagai, N.; Tatsumi, Y. Fungicidal Activity in the Presence of Keratin as an Important Factor Contributing to In Vivo Efficacy: A Comparison of Efinaconazole, Tavaborole, and Ciclopirox. J. Fungi 2017, 3, 58. [Google Scholar] [CrossRef] [PubMed]
- Bleackley, M.R.; Dawson, C.S.; McKenna, J.A.; Quimbar, P.; Hayes, B.M.; van der Weerden, N.L.; Anderson, M.A. Synergistic activity between two antifungal proteins, the plant Defensin NaD1 and the bovine pancreatic trypsin inhibitor. Msphere 2017, 2, e00390-17. [Google Scholar] [CrossRef]
- Vriens, K.; Cools, T.L.; Harvey, P.J.; Craik, D.J.; Spincemaille, P.; Cassiman, D.; Braem, A.; Vleugels, J.; Nibbering, P.H.; Drijfhout, J.W. Synergistic activity of the plant defensin HsAFP1 and caspofungin against Candida albicans biofilms and planktonic cultures. PLoS ONE 2015, 10, e0132701. [Google Scholar] [CrossRef]
- Su, S.; Yan, H.; Min, L.; Wang, H.; Chen, X.; Shi, J.; Sun, S. The antifungal activity of caspofungin in combination with antifungals or non-antifungals against Candida species in vitro and in clinical therapy. Expert Rev. Anti-Infect. Ther. 2022, 20, 161–178. [Google Scholar] [CrossRef]
- Zaikovska, O.; Kisis, J.; Pilmane, M. Defensins and cytokins in nail unit affected by onychomycosis; a pilot study. Pap. Anthropol. 2013, 22, 225–233. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Pathogen | MIC (µg/mL) | MFC (µg/mL) |
|---|---|---|
| Trichophyton rubrum | 20 ± 5 | 25 ± 5 |
| Trichophyton mentagrophytes | 13.3 ± 3.3 | 15 ± 5 |
| Candida albicans | 30 ± 0 | 30 ± 0 |
| Fusarium oxysporum | 15.8 ± 2.2 | 21.7 ± 1.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van der Weerden, N.L.; Parisi, K.; McKenna, J.A.; Hayes, B.M.; Harvey, P.J.; Quimbar, P.; Wevrett, S.R.; Veneer, P.K.; McCorkelle, O.; Vasa, S.; et al. The Plant Defensin Ppdef1 Is a Novel Topical Treatment for Onychomycosis. J. Fungi 2023, 9, 1111. https://doi.org/10.3390/jof9111111
van der Weerden NL, Parisi K, McKenna JA, Hayes BM, Harvey PJ, Quimbar P, Wevrett SR, Veneer PK, McCorkelle O, Vasa S, et al. The Plant Defensin Ppdef1 Is a Novel Topical Treatment for Onychomycosis. Journal of Fungi. 2023; 9(11):1111. https://doi.org/10.3390/jof9111111
Chicago/Turabian Stylevan der Weerden, Nicole L., Kathy Parisi, James A. McKenna, Brigitte M. Hayes, Peta J. Harvey, Pedro Quimbar, Sean R. Wevrett, Prem K. Veneer, Owen McCorkelle, Shaily Vasa, and et al. 2023. "The Plant Defensin Ppdef1 Is a Novel Topical Treatment for Onychomycosis" Journal of Fungi 9, no. 11: 1111. https://doi.org/10.3390/jof9111111