
Molecular and Functional Analyses of Characterized Sesquiterpene Synthases in Mushroom-Forming Fungi

Abstract
:1. Introduction
2. Materials and Methods
2.1. Literature Search for Characterized STSs
2.2. Phylogenetic Tree Construction
2.3. Analysis of Protein Motifs and Domains
2.4. Plasmids and Strain Construction
2.5. Heterologous Expression of STSs in Yeast
2.6. GC–MS Analysis
3. Results
3.1. Database of Characterized STSs
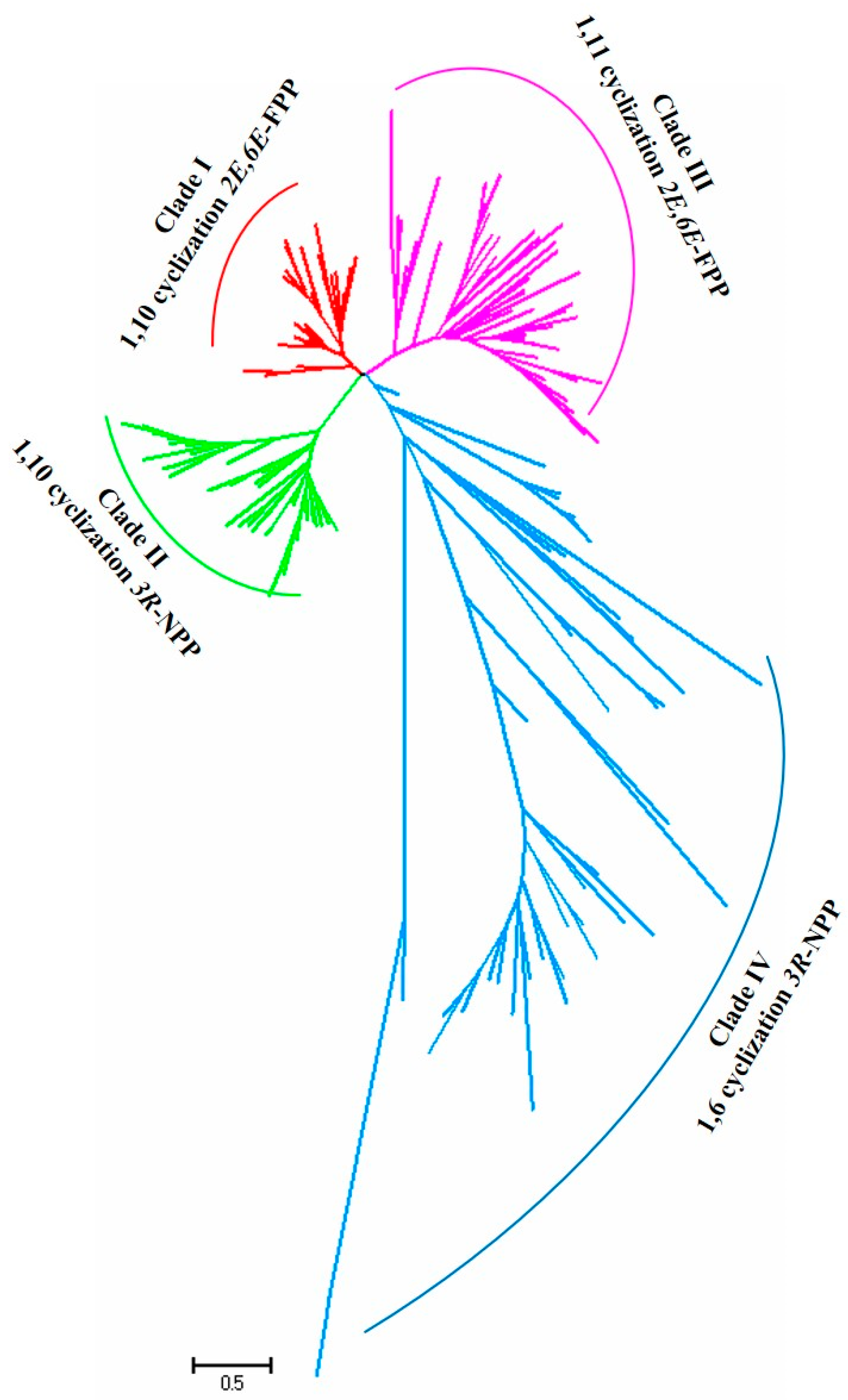
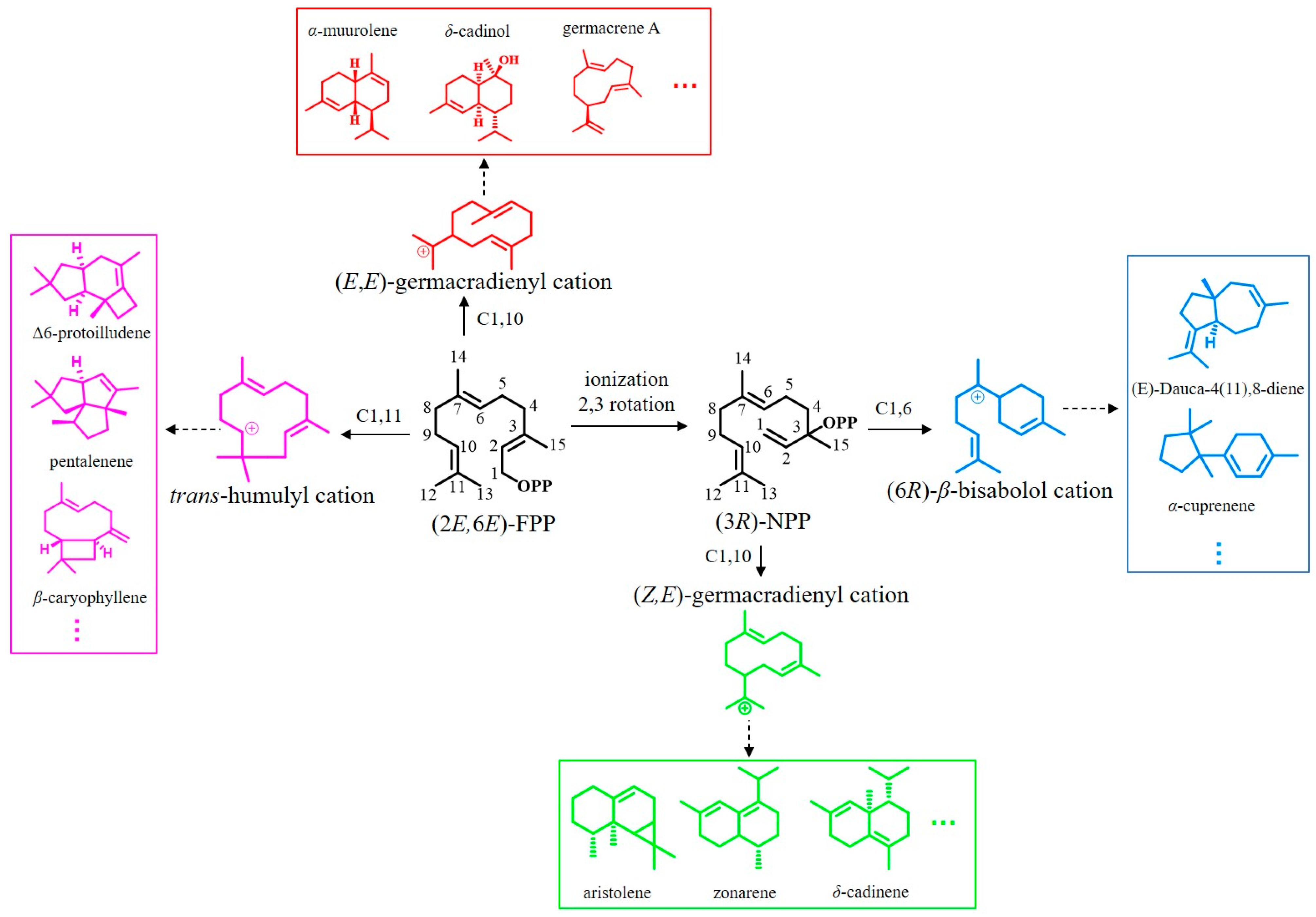
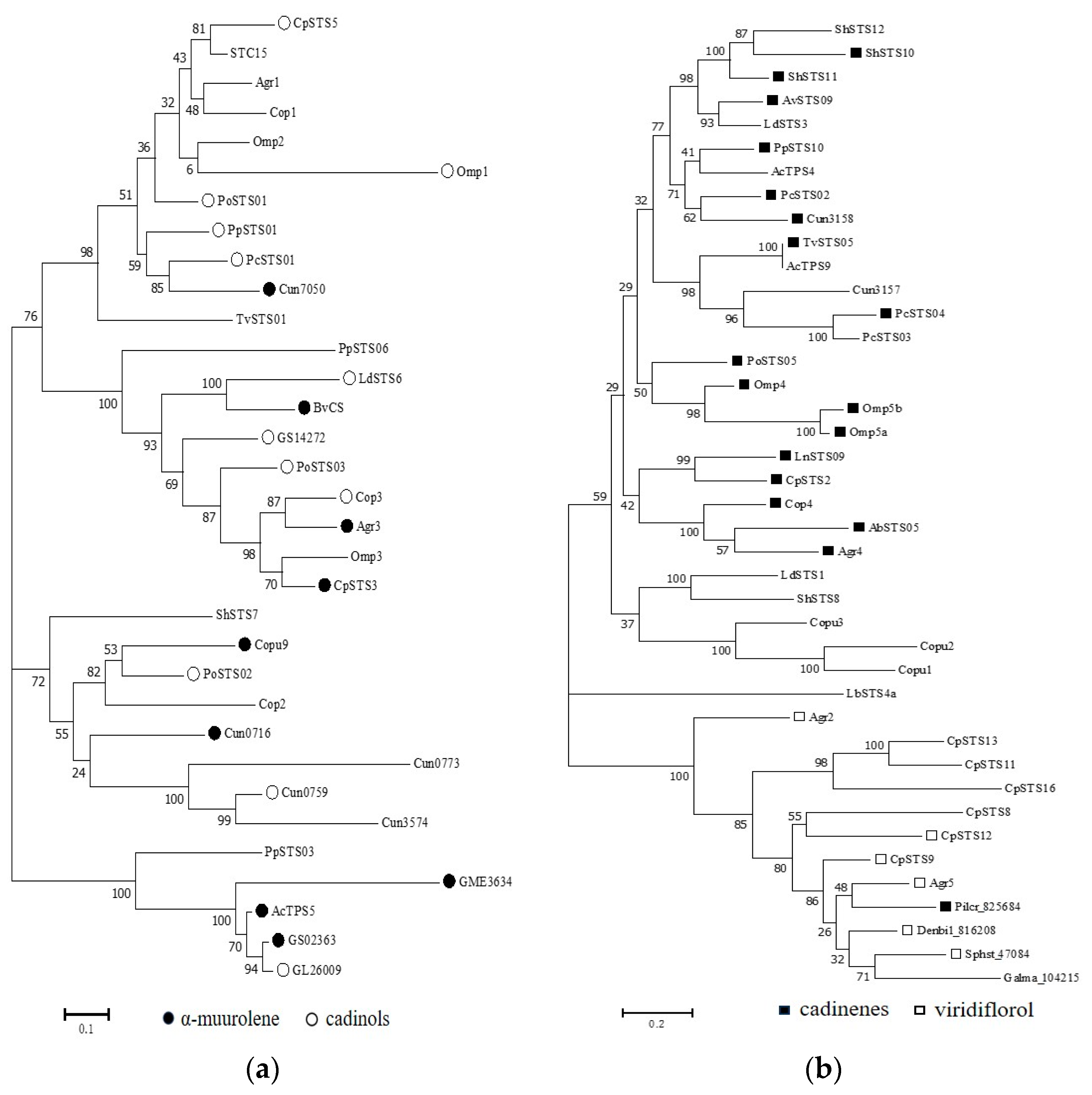
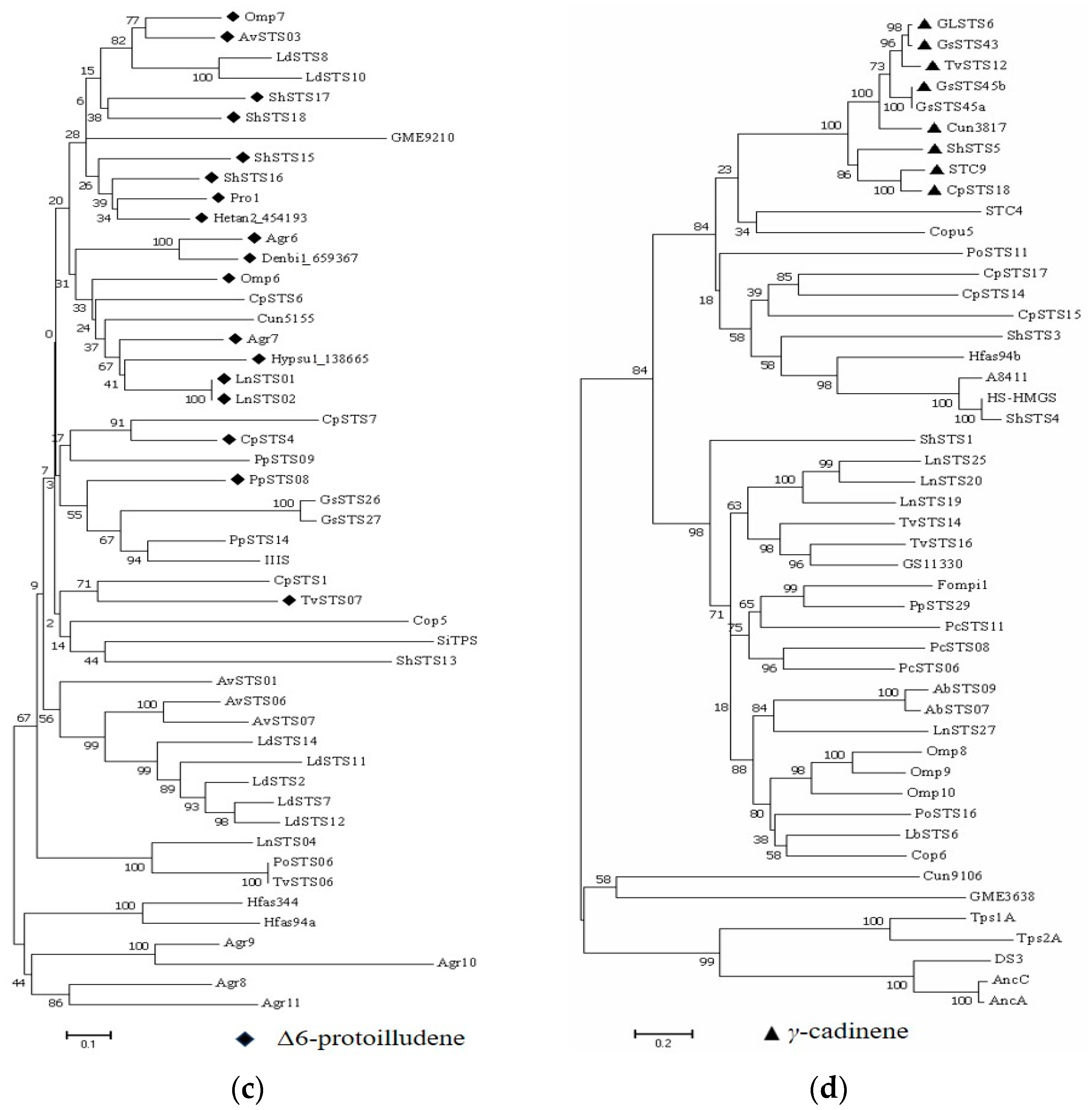
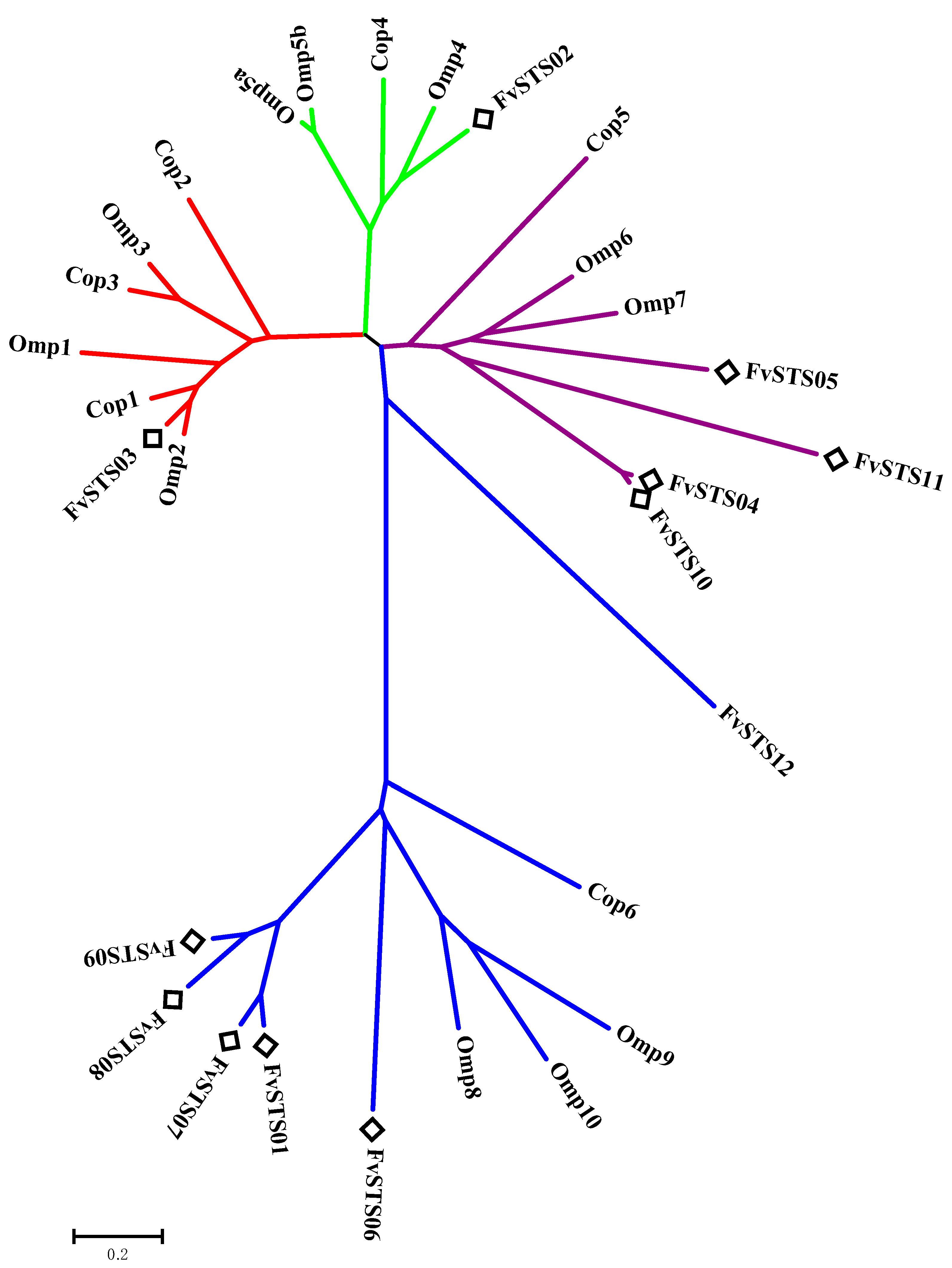
3.2. Phylogenetic Analysis
3.3. Analysis of Protein Conserved Motifs
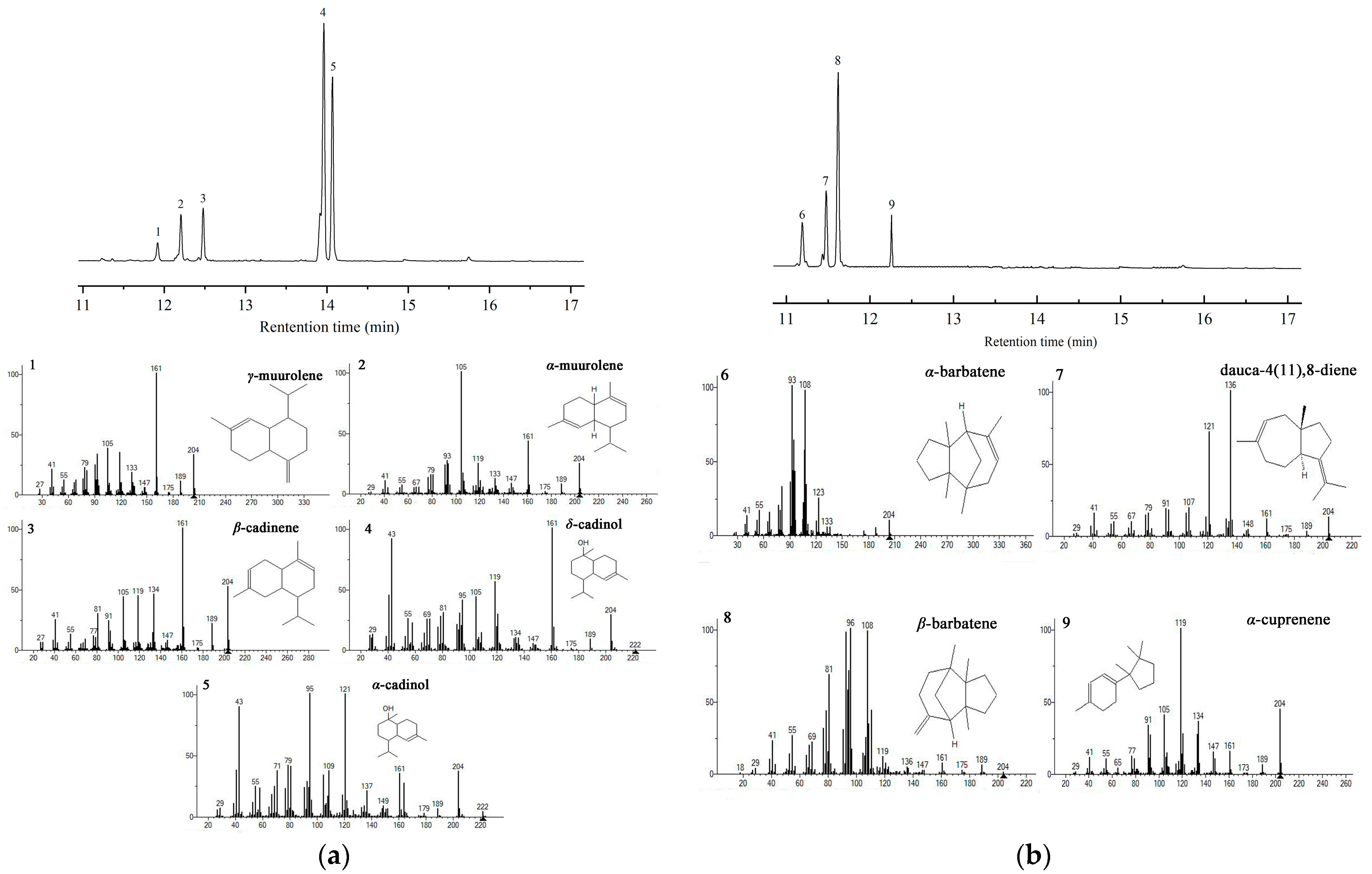
3.4. Characterization of FvSTSs
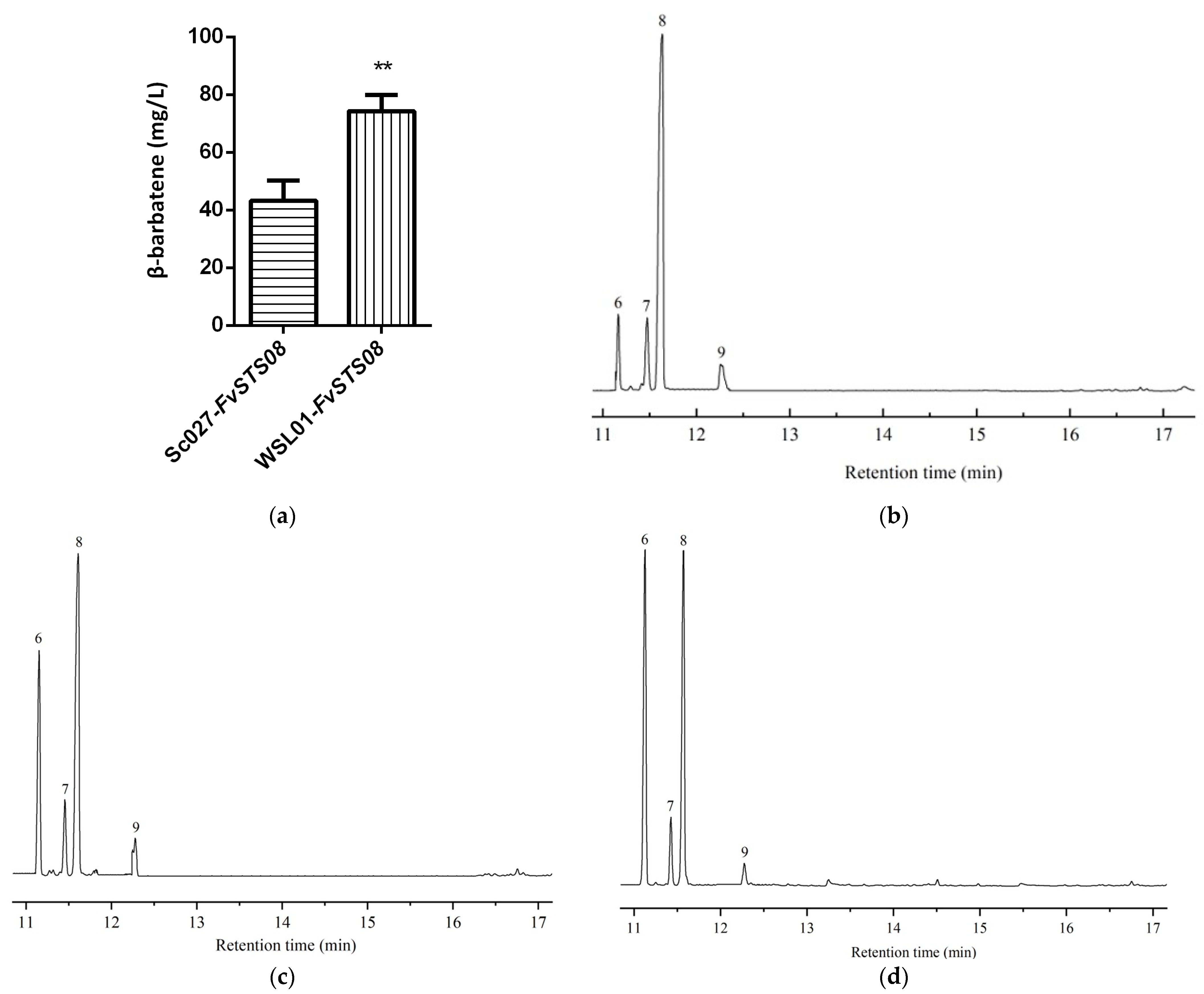
3.5. Heterologous Production of β-Barbatene in S. cerevisiae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fraga, B.M. Natural sesquiterpenoids. Nat. Prod. Rep. 2012, 29, 133. [Google Scholar]
- Kramer, R.; Abraham, W.R. Volatile sesquiterpenes from fungi: What are they good for? Phytochem. Rev. 2012, 11, 15–37. [Google Scholar]
- Bartikova, H.; Hanusova, V.; Skalova, L.; Ambroz, M.; Bousova, I. Antioxidant, pro-oxidant and other biological activities of ssesquiterpenes. Curr. Top. Med. Chem. 2014, 14, 2478–2494. [Google Scholar] [CrossRef]
- Abu-Izneid, T.; Rauf, A.; Shariati, M.A.; Khalil, A.A.; Imran, M.; Rebezov, M.; Uddin, M.S.; Mahomoodally, M.F.; Rengasamy, K.R.R. Sesquiterpenes and their derivatives-natural anticancer compounds: An update. Pharmacol. Res. 2020, 161, 105165. [Google Scholar]
- Chen, X.; Zhang, C.; Lindley, N.D. Metabolic engineering strategies for sustainable terpenoid flavor and fragrance synthesis. J. Agric. Food Chem. 2020, 68, 10252–11026. [Google Scholar] [CrossRef]
- Peralta-Yahya, P.P.; Ouellet, M.; Chan, R.; Mukhopadhyay, A.; Keasling, J.D.; Lee, T.S. Identification and microbial production of a terpene-based advanced biofuel. Nat. Commun. 2011, 2, 483. [Google Scholar] [CrossRef]
- Wang, S.; Zhan, C.; Chen, R.; Li, W.; Song, H.; Zhao, G.; Wen, M.; Liang, D.; Qiao, J. Achievements and perspectives of synthetic biology in botanical insecticides. J. Cell Physiol. 2022. [Google Scholar] [CrossRef]
- Lee, K.H.; Morris-Natschke, S.L.; Yang, X.; Huang, R.; Zhou, T.; Wu, S.F.; Shi, Q.; Itokawa, H. Recent progress of research on medicinal mushrooms, foods, and other herbal products used in traditional chinese medicine. J. Tradit. Complement. Med. 2012, 2, 1–12. [Google Scholar]
- Alves, M.; Ferreira, I.; Dias, J.; Teixeira, V.; Martins, A.; Pintado, M. A review on antifungal activity of mushroom (basidiomycetes) extracts and isolated compounds. Curr. Top. Med. Chem. 2013, 13, 2648–2659. [Google Scholar] [CrossRef] [PubMed]
- Muszyńska, B.; Grzywacz-Kisielewska, A.; Kała, K.; Gdula-Argasińska, J. Anti-inflammatory properties of edible mushrooms: A review. Food Chem. 2018, 243, 373–381. [Google Scholar] [PubMed]
- Fukushima-Sakuno, E. Bioactive small secondary metabolites from the mushrooms Lentinula edodes and Flammulina velutipes. J. Antibiot. 2020, 73, 687–696. [Google Scholar]
- Wirth, S.; Krause, K.; Kunert, M.; Broska, S.; Paetz, C.; Boland, W.; Kothe, E. Function of sesquiterpenes from Schizophyllum commune in interspecific interactions. PLoS ONE 2021, 16, e0245623. [Google Scholar]
- Abraham, W.R. Bioactive sesquiterpenes produced by fungi are they useful for humans as well? Curr. Med. Chem. 2001, 8, 583–606. [Google Scholar] [CrossRef]
- Luo, D.Q.; Wang, F.; Bian, X.Y.; Liu, J.K. Rufuslactone, a new antifungal sesquiterpene from the fruiting bodies of the basidiomycete Lactarius rufus. J. Antibiot. 2005, 58, 456–459. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.P.; Liu, J.K. Secondary metabolites from higher fungi. Prog. Chem. Org. Nat. Prod. 2017, 106, 1–201. [Google Scholar]
- Wu, J.; Yang, X.; Duan, Y.; Wang, P.; Qi, J.; Gao, J.M.; Liu, C. Biosynthesis of sesquiterpenes in basidiomycetes: A review. J. Fungi 2022, 8, 913. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, X.; Orban, A.; Shukal, S.; Birk, F.; Too, H.P.; Rühl, M. Agrocybe aegerita serves as a gateway for identifying sesquiterpene biosynthetic enzymes in higher fungi. ACS Chem. Biol. 2020, 15, 1268–1277. [Google Scholar]
- Ichinose, H.; Kitaoka, T. Insight into metabolic diversity of the brown-rot basidiomycete Postia placenta responsible for sesquiterpene biosynthesis: Semi-comprehensive screening of cytochrome P450 monooxygenase involved in protoilludene metabolism. Microb. Biotechnol. 2018, 11, 952–996. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, J.K.; Zhao, Q.; He, Q.L. Mechanistic investigations of hirsutene biosynthesis catalyzed by a chimeric sesquiterpene synthase from Steccherinum ochraceum. Fungal Genet. Biol. 2022, 161, 10370. [Google Scholar] [CrossRef]
- Asai, S.; Tsunematsu, Y.; Masuya, T.; Otaka, J.; Osada, H.; Watanabe, K. Uncovering hidden sesquiterpene biosynthetic pathway through expression boost area-mediated productivity enhancement in basidiomycete. J. Antibiot. 2020, 73, 721–728. [Google Scholar] [CrossRef]
- Christianson, D.W. Structural and chemical biology of terpenoid cyclases. Chem. Rev. 2017, 117, 11570–11648. [Google Scholar] [PubMed]
- Yuan, Y.; Litzenburger, M.; Cheng, S.; Bian, G.; Hu, B.; Yan, P.; Cai, Y.; Deng, Z.; Bernhardt, R.; Liu, T. Sesquiterpenoids produced by combining two sesquiterpene cyclases with promiscuous myxobacterial CYP260B1. ChemBioChem 2019, 20, 677–682. [Google Scholar] [CrossRef]
- Ichinose, H.; Ukeba, S.; Kitaoka, T. Latent potentials of the white-rot basidiomycete Phanerochaete chrysosporium responsible for sesquiterpene metabolism: CYP5158A1 and CYP5144C8 decorate (E)-α-Bisabolene. Enzyme Microb. Technol. 2022, 158, 110037. [Google Scholar] [CrossRef] [PubMed]
- Cao, R.; Wu, X.; Wang, Q.; Qi, P.; Zhang, Y.; Wang, L.; Sun, C. Characterization of γ-cadinene enzymes in Ganoderma lucidum and Ganoderma sinensis from basidiomycetes provides insight into the identification of terpenoid synthases. ACS Omega 2022, 7, 7229–7723. [Google Scholar] [CrossRef] [PubMed]
- Cheong, C.B.; Peh, G.; Wei, Y.; T, R.; Ang, E.L.; Zhao, H.; Zhang, C.; Lim, Y.H. A spirobicyclo [3.1.0]terpene from the investigation of sesquiterpene synthases from Lactarius deliciosus. ACS Chem. Biol. 2023, 18, 134–140. [Google Scholar] [CrossRef] [PubMed]
- López-Gallego, F.; Wawrzyn, G.T.; Schmidt-Dannert, C. Selectivity of fungal sesquiterpene synthases: Role of the active site’s H-1α Loop in catalysis. Appl. Environ. Microbiol. 2010, 76, 7723–7773. [Google Scholar] [CrossRef] [PubMed]
- Lauchli, R.; Pitzer, J.; Kitto, R.Z.; Kalbarczyk, K.Z.; Rabe, K.S. Improved selectivity of an engineered multi-product terpene synthase. Org. Biomol. Chem. 2014, 12, 4013–4402. [Google Scholar]
- Quin, M.B.; Michel, S.N.; Schmidt-Dannert, C. Moonlighting metals: Insights into regulation of cyclization pathways in fungal Δ6 -protoilludene sesquiterpene synthases. ChemBioChem 2015, 16, 2191–2219. [Google Scholar] [CrossRef]
- Quin, M.B.; Wawrzyn, G.; Schmidt-Dannert, C. Purification, crystallization and preliminary X-ray diffraction analysis of Omp6, a protoilludene synthase from Omphalotus olearius. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2013, 69, 574–577. [Google Scholar] [CrossRef]
- Ringel, M.; Dimos, N.; Himpich, S.; Haack, M.; Huber, C.; Eisenreich, W.; Schenk, G.; Loll, B.; Brück, T. Biotechnological potential and initial characterization of two novel sesquiterpene synthases from basidiomycota Coniophora puteana for heterologous production of Δ-Cadinol. Microb. Cell Fact. 2022, 21, 6. [Google Scholar]
- Wawrzyn, G.T.; Quin, M.B.; Choudhary, S.; López-Gallego, F.; Schmidt-Dannert, C. Draft genome of Omphalotus olearius provides a predictive framework for sesquiterpenoid natural product biosynthesis in basidiomycota. Chem. Biol. 2012, 19, 772–778. [Google Scholar] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yan, X.; Zhang, Y.; Liang, D.; Caiyin, Q.; Qiao, J. Characterization of trans-nerolidol synthase from Celastrus angulatus maxim and production of trans-nerolidol in engineered Saccharomyces cerevisiae. J. Agric. Food Chem. 2021, 69, 2236–2244. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 31–33. [Google Scholar] [CrossRef]
- Zeng, B.X.; Yao, M.D.; Wang, Y.; Xiao, W.H.; Yuan, Y.J. Metabolic engineering of Saccharomyces cerevisiae for enhanced dihydroartemisinic acid production. Front. Bioeng. Biotechnol. 2020, 8, 15. [Google Scholar] [CrossRef]
- Rodriguez, S.; Kirby, J.; Denby, C.M.; Keasling, J.D. Production and quantification of sesquiterpenes in Saccharomyces cerevisiae, including extraction, detection and quantification of terpene products and key related metabolites. Nat. Protoc. 2014, 9, 1980–1996. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Zhou, J.; Ge, J.; Li, W.; Liang, D.; Singh, W.; Black, G.; Nie, S.; Liu, J.; Sun, M.; et al. Computer-informed engineering: A new class I sesquiterpene synthase JeSTS4 for the synthesis of an unusual C10-(S)-bicyclogermacrene. ACS Catal. 2022, 12, 4037–4404. [Google Scholar] [CrossRef]
- Agger, S.; Lopez-Gallego, F.; Schmidt-Dannert, C. Diversity of sesquiterpene synthases in the basidiomycete Coprinus cinereus. Mol. Microbiol. 2009, 72, 1181–1195. [Google Scholar] [CrossRef]
- Quin, M.B.; Flynn, C.M.; Wawrzyn, G.T.; Choudhary, S.; Schmidt-Dannert, C. Mushroom hunting by using bioinformatics: Application of a predictive framework facilitates the selective identification of sesquiterpene synthases in basidiomycota. ChemBioChem 2013, 14, 2480–2491. [Google Scholar] [CrossRef]
- Flynn, C.M.; Schmidt-Dannert, C. Sesquiterpene synthase–3-hydroxy-3-methylglutaryl coenzyme A synthase fusion protein responsible for hirsutene biosynthesis in Stereum hirsutum. Appl. Environ. Microbiol. 2018, 84, e00036-1. [Google Scholar] [CrossRef]
- Nagamine, S.; Liu, C.; Nishishita, J.; Kozaki, T.; Sogahata, K.; Sato, Y.; Minami, A.; Ozaki, T.; Schmidt-Dannert, C.; Maruyama, J.; et al. Ascomycete Aspergillus oryzae is an efficient expression host for production of basidiomycete terpenes by using genomic DNA sequences. Appl. Environ. Microbiol. 2019, 85, e00409-1. [Google Scholar] [PubMed]
- Lin, Y.L.; Ma, L.T.; Lee, Y.R.; Shaw, J.F.; Wang, S.Y.; Chu, F.H. Differential gene expression network in terpenoid synthesis of Antrodia cinnamomea in mycelia and fruiting bodies. J. Agric. Food Chem. 2017, 65, 1874–1886. [Google Scholar] [CrossRef] [PubMed]
- Hewage, R.T.; Tseng, C.C.; Liang, S.Y.; Lin, H.C. Genome mining of cryptic bisabolenes that were biosynthesized by intramembrane terpene synthases from Antrodia cinnamomea. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2023, 378, 20220033. [Google Scholar] [CrossRef]
- Chen, T.; Chen, C.; Lee, C.; Huang, R.; Chen, K.; Lu, Y.; Liang, S.; Pham, M.; Rao, Y.K.; Wu, S.; et al. The biosynthetic gene cluster of mushroom-derived antrocin encodes two dual-functional haloacid dehalogenase-like terpene cyclases. Angew. Chem. Int. Ed. 2023, 62, e202215566. [Google Scholar] [CrossRef]
- Zhou, H.; Yang, Y.L.; Zeng, J.; Zhang, L.; Ding, Z.H.; Zeng, Y. Identification and characterization of a Δ-cadinol synthase potentially involved in the formation of boreovibrins in Boreostereum vibrans of basidiomycota. Nat. Prod. Bioprospect. 2016, 6, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Yap, H.Y.Y.; Muria-Gonzalez, M.J.; Kong, B.H.; Stubbs, K.A.; Tan, C.S.; Ng, S.T.; Tan, N.H.; Solomon, P.S.; Fung, S.Y.; Chooi, Y.H. Heterologous expression of cytotoxic sesquiterpenoids from the medicinal mushroom Lignosus rhinocerotis in Yeast. Microb. Cell Fact. 2017, 16, 10. [Google Scholar] [CrossRef]
- Mischko, W.; Hirte, M.; Fuchs, M.; Mehlmer, N.; Brück, T.B. Identification of sesquiterpene synthases from the basidiomycota Coniophora puteana for the efficient and highly selective β-copaene and cubebol production in E. coli. Microb. Cell Fact. 2018, 17, 164. [Google Scholar] [CrossRef]
- Engels, B.; Heinig, U.; Grothe, T.; Stadler, M.; Jennewein, S. Cloning and characterization of an Armillaria gallica cDNA encoding protoilludene synthase, which catalyzes the first committed step in the synthesis of antimicrobial melleolides. J. Biol. Chem. 2011, 286, 6871–6878. [Google Scholar] [CrossRef]
- Al-Salihi, S.A.A.; Dao, T.T.; Williams, K.; Bailey, A.M.; Foster, G.D. The biogenetic origin of the biologically active naematolin of Hypholoma species involves an unusual sesquiterpene synthase. Mol. Biotechnol. 2019, 61, 754–776. [Google Scholar] [CrossRef]
- Püth, N.; Ersoy, F.; Krings, U.; Berger, R.G. Sesquiterpene cyclases from the basidiomycete Cerrena unicolor. Catalysts 2021, 11, 136. [Google Scholar] [CrossRef]
- Wang, Q.; Qi, P.; Zhao, C.; Zhang, Y.; Wang, L.; Yu, H. Tandem expression of Ganoderma sinense sesquiterpene synthase and IDI promotes the production of gleenol in E. coli. Appl. Microbiol. Biotechnol. 2022, 106, 7779–7791. [Google Scholar] [PubMed]
- Chu, L.H.; Wang, L.Z.; Chen, S.L.; Zeng, X.Y.; Xu, J.; Li, Y.; Sun, C. Functional identification of a multi-product sesquiterpene synthase from Ganoderma sinense. Chin. J. Exp. Tradit. Med. Formulae 2019, 25, 151–157. [Google Scholar]
- Wei, J.; Pu, X.; Wang, L.; Sun, S.; Sun, C.; Wang, H. Cloning and characterization of sesquiterpene synthase genes from the Ganoderma sinense genome. Sci. Sin. Vitae 2018, 48, 447–454. [Google Scholar] [CrossRef]
- Lizhi, W.; Xiangdong, P.; Shiqiang, T.; Sijie, S.; Yutong, B.; Chao, S.; Shilin, C.; Haiying, W. Cloning and expression of the sesquiterpene synthase gene from Ganoderma lucidum. J. Agric. Univ. Hebei 2017, 40, 67–72. [Google Scholar]
- Burkhardt, I.; Kreuzenbeck, N.B.; Beemelmanns, C.; Dickschat, J.S. Mechanistic characterization of three sesquiterpene synthases from the termite-associated fungus Termitomyces. Org. Biomol. Chem. 2019, 17, 3348–3355. [Google Scholar] [CrossRef]
- Masunaga, N.; Kitaoka, T.; Ichinose, H. Biocatalyst collection and heterologous expression of sesquiterpene synthases from basidiomycetous fungi: Discovery of a novel sesquiterpene hydrocarbon. Microb. Biotechnol. 2023, 16, 632–664. [Google Scholar] [CrossRef]
- Chen, R.; Feng, T.; Li, M.; Zhang, X.; He, J.; Hu, B.; Deng, Z.; Liu, T.; Liu, J.K.; Wang, X.; et al. Characterization of tremulane sesquiterpene synthase from the basidiomycete Irpex lacteus. Org. Lett. 2022, 24, 5669–5673. [Google Scholar] [CrossRef]
- Ntana, F.; Bhat, W.W.; Johnson, S.R.; Jørgensen, H.J.L.; Collinge, D.B.; Jensen, B.; Hamberger, B. A sesquiterpene synthase from the endophytic fungus serendipita indica catalyzes formation of viridiflorol. Biomolecules 2021, 11, 89. [Google Scholar] [CrossRef]
- Kreuzenbeck, N.B.; Dhiman, S.; Roman, D.; Burkhardt, I.; Conlon, B.H.; Fricke, J.; Guo, H.; Blume, J.; Görls, H.; Poulsen, M.; et al. Isolation, (bio)synthetic studies and evaluation of antimicrobial properties of drimenol-type sesquiterpenes of Termitomyces fungi. Commun. Chem. 2023, 6, 7. [Google Scholar]
- Nosenko, T.; Zimmer, I.; Ghirardo, A.; Köllner, T.G.; Weber, B.; Polle, A.; Rosenkranz, M.; Schnitzler, J.P. Predicting functions of putative fungal sesquiterpene synthase genes based on multiomics data analysis. Fungal Genet. Biol. 2023, 165, 103779. [Google Scholar] [CrossRef]
- Gressler, M.; Löhr, N.A.; Schäfer, T.; Lawrinowitz, S.; Seibold, P.S.; Hoffmeister, D. Mind the mushroom: Natural product biosynthetic genes and enzymes of basidiomycota. Nat. Prod. Rep. 2021, 38, 702–772. [Google Scholar] [PubMed]
- Quin, M.B.; Flynn, C.M.; Schmidt-Dannert, C. Traversing the fungal terpenome. Nat. Prod. Rep. 2014, 31, 1449–1473. [Google Scholar] [PubMed]
- Aaron, J.A.; Christianson, D.W. Trinuclear metal clusters in catalysis by terpenoid synthases. Pure Appl. Chem. 2010, 82, 1585–1597. [Google Scholar] [CrossRef] [PubMed]
- Rynkiewicz, M.J.; Cane, D.E.; Christianson, D.W. Structure of trichodiene synthase from Fusarium sporotrichioides provides mechanistic inferences on the terpene cyclization cascade. Proc. Natl. Acad. Sci. USA 2001, 98, 13543–13548. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, A.; Fukushima-Sakuno, E.; Osaki-Oka, K.; Futamura, Y.; Motoyama, T.; Osada, H.; Ishikawa, N.K.; Nagasawa, E.; Tokimoto, K. Productivity and bioactivity of enokipodins A–D of Flammulina rossica and Flammulina velutipes. Biosci. Biotechnol. Biochem. 2020, 84, 876–888. [Google Scholar] [CrossRef]
- Ishikawa, N.K.; Yamaji, K.; Tahara, S.; Fukushi, Y.; Takahashi, K. Highly oxidized cuparene-type sesquiterpenes from a mycelial culture of Flammulina velutipes. Phytochemistry 2000, 54, 777–778. [Google Scholar] [CrossRef]
- Ishikawa, N.K.; Fukushi, Y.; Yamaji, K.; Tahara, S.; Takahashi, K. Antimicrobial cuparene-type sesquiterpenes, enokipodins C and D, from a mycelial culture of Flammulina velutipes. J. Nat. Prod. 2001, 64, 932–993. [Google Scholar] [CrossRef]
- Li, H.P.; Yang, W.J.; Qu, S.X.; Pei, F.; Luo, X.; Mariga, A.M.; Ma, L. Variation of volatile terpenes in the edible fungi mycelia Flammulina velutipes and communications in fungus-mite interactions. Food Res. Int. 2018, 103, 150–155. [Google Scholar] [CrossRef]
- Fu, H.; Liang, Y.; Zhong, X.; Pan, Z.; Huang, L.; Zhang, H.; Xu, Y.; Zhou, W.; Liu, Z. Codon optimization with deep learning to enhance protein expression. Sci. Rep. 2020, 10, 1–9. [Google Scholar]
- Chen, X.; Köllner, T.G.; Jia, Q.; Norris, A.; Santhanam, B.; Rabe, P.; Dickschat, J.S.; Shaulsky, G.; Gershenzon, J.; Chen, F. Terpene synthase genes in eukaryotes beyond plants and fungi: Occurrence in social amoebae. Proc. Natl. Acad. Sci. USA 2016, 113, 12132–12137. [Google Scholar] [CrossRef]
- Petek, M.; Rotter, A.; Kogovšek, P.; Baebler, Š.; Mithöfer, A.; Gruden, K. Potato virus Y infection hinders potato defence response and renders plants more vulnerable to colorado potato beetle attack. Mol. Ecol. 2014, 23, 5378–5539. [Google Scholar] [CrossRef] [PubMed]
- Faldt, J.; Jonsell, M.; Nordlander, G.; Borg-Karlson, A.K. Volatiles of bracket fungi fomitopsis pinicola and fomes fomentarius and their functions as insect attractants. J Chem Ecol 1999, 25, 567–590. [Google Scholar] [CrossRef]
- Faulkner, A.; Chen, X.; Rush, J.; Horazdovsky, B.; Waechter, C.J.; Carman, G.M.; Sternweis, P.C. The LPP1 and DPP1 gene products account for most of the isoprenoid phosphate phosphatase activities in Saccharomyces Cerevisiae. J. Biol. Chem. 1999, 274, 14831–14837. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Ye, Z.; Wan, X.; Yao, G.; Duan, J.; Liu, J.; Yao, M.; Sun, X.; Deng, Z.; Shen, K.; et al. Systematic mining and evaluation of the sesquiterpene skeletons as high energy aviation fuel molecules. Adv. Sci. 2023, 10, 2300889. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.J.; Tantillo, D.J. Branching out from the bisabolyl cation. Unifying mechanistic pathways to barbatene, bazzanene, chamigrene, chamipinene, cumacrene, cuprenene, dunniene, isobazzanene, iso-γ-Bisabolene, isochamigrene, laurene, microbiotene, sesquithujene, sesquisabinene, thujopsene, trichodiene, and widdradiene sesquiterpenes. J. Am. Chem. Soc. 2014, 136, 2450–2463. [Google Scholar]
- Hong, Y.J.; Tantillo, D.J. Is a 1,4-alkyl shift involved in the biosynthesis of ledol and viridiflorol? J. Org. Chem. 2017, 82, 3957–3959. [Google Scholar] [CrossRef]
- Durairaj, J.; Di Girolamo, A.; Bouwmeester, H.J.; de Ridder, D.; Beekwilder, J.; van Dijk, A.D.J. An analysis of characterized plant sesquiterpene synthases. Phytochemistry 2019, 158, 157–165. [Google Scholar] [CrossRef]
- Yoshikuni, Y.; Ferrin, T.E.; Keasling, J.D. Designed divergent evolution of enzyme function. Nature 2006, 440, 1078–1082. [Google Scholar] [CrossRef]
- Huang, J.Q.; Li, D.M.; Li, J.X.; Lin, J.L.; Tian, X.; Wang, L.J.; Chen, X.Y.; Fang, X. 1,10/1,11-Cyclization catalyzed by diverged plant sesquiterpene synthases is dependent on a single residue. Org. Biomol. Chem. 2021, 19, 6650–6656. [Google Scholar] [CrossRef]
- Wang, Y.; Bao, L.; Yang, X.; Li, L.; Li, S.; Gao, H.; Yao, X.S.; Wen, H.; Liu, H.W. Bioactive sesquiterpenoids from the solid culture of the edible mushroom Flammulina velutipes growing on cooked rice. Food Chem. 2012, 132, 1346–1353. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Gene Name | GenBank or JGI Protein ID | Products | Clade | Reference |
|---|---|---|---|---|---|
| Lactarius deliciosus | LdSTS1 | KAH9077233.1 | 1,10-di-epi-cubenol | II | [25] |
| LdSTS2 | KAH9033224.1 | ND | III | ||
| LdSTS3 | KAH9035467.1 | myrcene, trans-β-ocimene, etc. | II | ||
| LdSTS6 | KAH9079282.1 | α-muurolene, γ-gurjunene, α-selinene, etc. | I | ||
| LdSTS7 | KAH9033225.1 | aristolene | III | ||
| LdSTS8 | KAH9071064.1 | ND | III | ||
| LdSTS10 | KAH9071064.1 | ND | III | ||
| LdSTS11 | KAH9053987.1 | aristolene | III | ||
| LdSTS12 | KAH9033225.1 | ND | III | ||
| LdSTS14 | KAH9033227.1 | ND | III | ||
| Coprinus cinereus | Cop1 | XP_001832573.1 | germacrene A; Δ-cadinene; α-muurolene; germacrene D | I | [38] |
| Cop2 | XP_001836556.1 | germacrene A; Δ-cadinene; α-muurolene | I | ||
| Cop3 | XP_001832925.1 | α-muurolene; germacrene A; γ-muurolene; germacrene D; Δ-cadinene; α-copaene | I | ||
| Cop4 | XP_001836356.1 | Δ-cadinene; β-copaene; β-cubebene; sativene; germacrene D; cubebol | II | ||
| Cop5 | XP_001834007.1 | ND | III | ||
| Cop6 | XP_001832549.1 | α-cuprenene | IV | ||
| Omphalotus olearius | Omp1 | JGI ID: 1311 | α-muurolene | I | [31] |
| Omp2 | not available | ND | I | ||
| Omp3 | JGI ID: 4636 | germacrene A; α-muurolene; elina-4,7-diene; Δ-cadinene | I | ||
| Omp4 | JGI ID: 1447 | Δ-cadinene | II | ||
| Omp5a | JGI ID: 2392 | γ-cadinene; epi-zonarene; germacrene A | II | ||
| Omp5b | JGI ID: 2393 | γ-cadinene; germacrene A | II | ||
| Omp6 | JGI ID: 4774 | Δ6-protoilludene | III | ||
| Omp7 | JGI ID: 2271 | Δ6-protoilludene; pentalenene | III | ||
| Omp8 | not available | ND | IV | ||
| Omp9 | JGI ID: 3258 | α-barbatene; β-barbatene | IV | ||
| Omp10 | JGI ID: 3981 | (E)-dauca-4(11),8-diene; daucene | IV | ||
| Stereum hirsutum | ShSTS1 | JGI ID: 159379 | β-barbatene; α-barbatene | IV | [39] |
| ShSTS11 | JGI ID: 128017 | Δ-cadinene; α-cubebene | II | ||
| ShSTS18 | JGI ID: 25180 | Δ6-protoilludene | III | ||
| ShSTS15 | JGI ID: 64702 | Δ6-protoilludene | III | ||
| ShSTS16 | JGI ID: 73029 | Δ6-protoilludene | III | ||
| ShSTS3 | JGI ID: 122776 | α-farnesene; β-farnesene | IV | ||
| ShSTS4 | JGI ID: 52743 | hirsutene | IV | ||
| ShSTS5 | JGI ID: 161672 | γ-cadinene | IV | ||
| ShSTS7 | JGI ID: 167646 | Δ-cadinene | I | ||
| ShSTS8 | JGI ID: 146390 | 1-epi-cubenol; α-cubebene | II | ||
| ShSTS10 | JGI ID: 111121 | Δ-cadinene; germacrene D | II | ||
| ShSTS12 | JGI ID: 111127 | α-cubebene; β-cubebene | II | ||
| ShSTS13 | JGI ID: 50042 | β-caryophyllene | III | ||
| ShSTS17 | JGI ID: 69906 | Δ6-protoilludene | III | ||
| HS-HMGS | not available | hirsutene; β-caryophyllene | IV | [40] | |
| Clitopillus pseudo-pinsitus | CpSTS1 | BBH51498.1 | sterpurene | III | [41] |
| CpSTS2 | BBH51499.1 | Δ-cadinene; α-cubebene | II | ||
| CpSTS3 | BBH51500.1 | Δ-cadinol; α-muurolene; γ-muurolene; unknown sesquiterpene | I | ||
| CpSTS4 | BBH51501.1 | Δ6-protoilludene | III | ||
| CpSTS5 | BBH51502.1 | α-muurolene; γ-muurolene | I | ||
| CpSTS6 | BBH51503.1 | pentalenene | III | ||
| CpSTS7 | BBH51504.1 | α-farnesene | III | ||
| CpSTS8 | BBH51505.1 | alloaromadendrene; unknown sesquiterpene | II | ||
| CpSTS9 | BBH51506.1 | virifloridol; ledene | II | ||
| CpSTS11 | BBH51508.1 | 9-alloaromadendrene | II | ||
| CpSTS12 | BBH51509.1 | virifloridol; β-elemene; ledene | II | ||
| CpSTS13 | BBH51510.1 | ledene; unknown sesquiterpene | II | ||
| CpSTS14 | BBH51511.1 | β-elemene; β-farnesene; α-farnesene | IV | ||
| CpSTS15 | BBH51512.1 | ND | IV | ||
| CpSTS16 | BBH51513.1 | aristolene; unknown sesquiterpene | II | ||
| CpSTS17 | BBH51514.1 | β-caryophyllene | IV | ||
| CpSTS18 | BBH51515.1 | γ-cadinene | IV | ||
| Antrodia cinnamomea | AcTPS4 | JGI ID: 40411 | zonarene; α-cubebene; sibirene; γ-cadinene | II | [42] |
| AcTPS5 | JGI ID: 40579 | T-cadinol; γ-cadinene | I | ||
| AcTPS7 | JGI ID: 36944 | nerolidol; α-farnesol | \ | ||
| AcTPS9 | JGI ID: 47706 | 1-epi-cubenol; sibirene; cubebol; α-cubebene; α-farnesol; γ-muurolene | II | ||
| Tps1A | KAI0942648.1 | (+)-(S,Z)-α-bisabolene | IV | [43] | |
| Tps2A | KAI0928020.1 | ||||
| AncA | ACg006372 | (R)-trans-γ-monocyclofarnesol | IV | [44] | |
| AncC | ACg006375 | drimane-type sesquiterpene (+)-albicanol | |||
| Boreostereum vibrans | BvCS | KU668561.1 | Δ-cadinol; α-muurolene; γ-muurolene | I | [45] |
| Lignosus rhinocerotis | GME3634 | KX281943 | α-cadinol; germacrene D-4-ol | I | [46] |
| GME3638 | KX281944 | torreyol; germacrene D-4-ol; β-cubebene | IV | ||
| GME9210 | KX281945 | 1,3,4,5,6,7-hexahydro-2,5,5-trimethyl-2H-2,4a-ethanonaphthalene | III | ||
| Agrocybe aegerita | Agr1 | MN146024 | Δ-cadinene; α-cadinol; Δ-cadinol; α-muurolene | I | [17] |
| Agr2 | MN146025 | viridiflorene | II | ||
| Agr3 | MN146026 | Δ-cadinol; Δ-cadinene; α-muurolene; γ-muurolene | I | ||
| Agr4 | MN146027 | Δ-cadinene; epicubenol; cadina-1(6),4-diene; β- myrcene | II | ||
| Agr5 | MN146028 | viridiflorol; viridiflorene | II | ||
| Agr6 | MN146029 | Δ6-protoilludene | III | ||
| Agr7 | MN146030 | Δ6-protoilludene | III | ||
| Agr8 | MN146031 | γ-muurolene; β-cadinene; Δ-cadinol | III | ||
| Agr9 | MN146032 | γ-muurolene; Δ-cadinene; unknown sesquiterpenol | III | ||
| Agr10 | MN146033 | ND | III | ||
| Agr11 | MN146034 | ND | III | ||
| Coniophora puteana | Copu1 | XP_007772164.1 | ND | II | [47] |
| Copu2 | XP_007771895.1 | β-copaene; germacrene D; cubebol; germacrene D-4-ol | II | ||
| Copu3 | XP_007765978.1 | cubebol; germacrene D-4-ol; Δ-cadinene | II | ||
| Copu5 | XP_007765330.1 | Δ-cadinol; Δ-cadinene; cubebol; α-cadinol | IV | [30] | |
| Copu9 | XP_007765560.1 | I | |||
| Piloderma croceum | Pilcr_825684 | JGI ID: 825684 | γ-cadinene; viridiflorene; β-elemene | II | [17] |
| Galerina marginata | Galma_104215 | JGI ID: 104215 | β-gurjunene | II | |
| Sphaerobolus stellatus | Sphst_47084 | JGI ID: 47084 | viridiflorol; viridiflorene | II | |
| Dendrothele bispora | Denbi1_816208 | JGI ID: 816208 | viridiflorol; viridiflorene | II | |
| Denbi1_659367 | JGI ID: 659367 | Δ6-protoilludene | III | ||
| Heterobasidion annosum | HEtan2_454193 | XP_009550163.1 | Δ6-protoilludene | III | |
| Hypholoma sublateritium | Hypsu1_138665 | A0A0D2L718.1 | Δ6-protoilludene | III | |
| Armillaria gallica | Pro1 | MT277003.1 | Δ6-protoilludene | III | [48] |
| Hypholoma fasciculare | Hfas94a | MK287936.1 | α-humulene; β-caryophyllene | III | [49] |
| Hfas94b | MK287937.1 | α-humulene; β-caryophyllene | IV | ||
| Hfas255 | not available | ND | \ | ||
| Hfas344 | MK287938.1 | unknown sesquiterpene | III | ||
| Cerrena unicolor | Cun3817 | JGI ID: 3817 | γ-cadinene | IV | [50] |
| Cun5155 | JGI ID: 5155 | aromadendrene | III | ||
| Cun3157 | JGI ID: 3157 | β-cubebene; germacrene D; epicubenol; Δ-cadinene | II | ||
| Cun3158 | JGI ID: 3158 | Δ-cadinene; germacrene D; β-cubebene; γ-amorphene | II | ||
| Cun0773 | JGI ID: 0773 | germacrene D | I | ||
| Cun7050 | JGI ID: 7050 | Δ-cadinol | I | ||
| Cun0716 | JGI ID: 0716 | Δ-cadinol; α-muurolene | I | ||
| Cun0759 | JGI ID: 0759 | α-muurolene | I | ||
| Cun3574 | JGI ID: 3574 | α-copaene | I | ||
| Cun9106 | JGI ID: 9106 | unknown sesquiterpene | IV | ||
| Postia placenta | PpSTS01 | XP_024337827.1 | α-muurolene; Δ-cadinene; β-elemene | I | [18] |
| PpSTS03 | A0A348B781.1 | γ-cadinene; α-cadinene; Δ-cadinene; β-elemene | I | ||
| PpSTS06 | A0A348B782.1 | α-gurjunene; bicycloelemene; bicyclogermacrene | I | ||
| PpSTS08 | A0A348B784.1 | Δ6-protoilludene | III | ||
| PpSTS09 | A0A348B785.1 | unknown sesquiterpene | III | ||
| PpSTS10 | XP_024334632.1 | Δ-cadinene; β-copaene; sativene; sesquisabinene | II | ||
| PpSTS14 | A0A348B788.1 | pentalenene; caryophyllene | III | ||
| PpSTS29 | A0A348B794.1 | unknown sesquiterpene | IV | ||
| Fomitopsis pinicola | Fompi1 | JGI ID: 84944 | α-cuprenene | IV | [31] |
| Phanerodontia chrysosporium | PcSTS01 | BCX55496.1 | α-muurolene; Δ-cadinene; γ-muurolene; α-muurolol | I | [23] |
| PcSTS02 | BCX55497.1 | Δ-cadinene; β-copaene; β-farnesene; cadina-1(6),4-diene | II | ||
| PcSTS03 | BCX55498.1 | epicubenol | II | ||
| PcSTS04 | BCX55499.1 | Δ-cadinene; β-farnesene; β-copaene; epicubenol | II | ||
| PcSTS06 | BCX55500.1 | β-barbatene; α-barbatene | IV | ||
| PcSTS08 | BCX55502.1 | (E)-α-bisabolene | IV | ||
| PcSTS11 | BCX55504.1 | α-santalene | IV | ||
| Steccherinum ochraceum | A8411 | not available | hirsutene | IV | [19] |
| Ganoderma sinensis | GsSTS43 | PIL26225 | γ-cadinene | IV | [24] |
| GsSTS45a | UDP19925 | ND | IV | ||
| GsSTS45b | UDP19925 | γ-cadinene | IV | ||
| GsSTS26 | MT584777.1 | gleenol; di-epi-1,10-cubenol; Ʈ-muurolol | III | [51] | |
| GsSTS27 | OP094045 | III | |||
| GS02363 | PIL35634 | α-cadinol; Δ-cadinene; γ-cadinene; T-cadinol | I | [52] | |
| GS14272 | PIL24516 | α-muurolene | I | [53] | |
| GS11330 | not available | α-cuprenene | IV | ||
| Ganoderma lucidum | GL26009 | not available | α-muurolene; γ-muurolene | I | [54] |
| GLSTS6 | UDP19923 | γ-cadinene | IV | [24] | |
| Termitomyces sp. J132 | STC4 | KNZ72568.1 | (+)-intermedeol; α-selinene; β-selinene | IV | [55] |
| STC9 | KAG5341349 | γ-cadinene | IV | ||
| STC15 | KNZ74377.1 | (+)-germacrene D-4-ol; γ-cadinene; Δ-cadinene; α-cadinene; β-elemene | I | ||
| Agaricus bisporus | AbSTS05 | LC712879 | cadina-1,4-diene; cadina-1(6),4-diene; Δ-cadinene; zonarene; epicubenol; cadin-4-en-10-ol | II | [56] |
| AbSTS07 | LC712880 | Δ-cadinene; epizonarene | IV | ||
| AbSTS09 | LC712881 | (Z)-α-bisabolene | IV | ||
| Auriscalpium vulgare | AvSTS01 | LC712882 | unknown sesquiterpene | III | |
| AvSTS03 | LC712883 | Δ6-protoilludene | III | ||
| AvSTS06 | LC712885 | (E)-nerolidol | III | ||
| AvSTS07 | LC712886 | (E)-nerolidol | III | ||
| AvSTS09 | LC712887 | cadina-1,4-diene; cadina-1(6),4-diene; Δ-cadinene; β-copaene; zonarene; epicubenol; cadin-4-en-10-ol | II | ||
| Lepista nuda | LnSTS01 | LC712891 | Δ6-protoilludene | III | |
| LnSTS02 | LC712892 | Δ6-protoilludene | III | ||
| LnSTS04 | LC719126 | pleostene; isobazzanene | III | ||
| LnSTS09 | LC712895 | cadina-1,4-diene; cadina-1(6),4-diene; Δ-cadinene; β-copaene; zonarene; epicubenol; cadin-4-en-10-ol | II | ||
| LnSTS19 | LC712898 | (E)-nerolidol | IV | ||
| LnSTS20 | LC712899 | β-barbatene | IV | ||
| LnSTS25 | LC712901 | unknown sesquiterpene | IV | ||
| LnSTS27 | LC712902 | acora-3(7),14-diene | IV | ||
| Pleurotus ostreatus | PoSTS01 | LC712903 | α-muurolene; isobazzanene; Δ-cadinene; α-muurolol | I | |
| PoSTS02 | LC712904 | α-muurolene; Δ-cadinene; α-muurolol; zonarene | I | ||
| PoSTS03 | LC712905 | α-muurolene; Δ-cadinene; isobazzanene; α-muurolol | I | ||
| PoSTS05 | LC712906 | cadina-1,4-diene; cadina-1(6),4-diene; Δ-cadinene; β-farnesene; zonarene; epicubenol; cadin-4-en-10-ol | II | ||
| PoSTS06 | LC712907 | pleostene | III | ||
| PoSTS11 | LC712908 | (E)-nerolidol | IV | ||
| PoSTS16 | LC712909 | α-cuprenene | IV | ||
| Trametes versicolor | TvSTS01 | LC712910 | Δ-cadinene; cadin-4-en-10-ol; Ʈ-muurolol | I | |
| TvSTS05 | LC712912 | cadina-1,4-diene; cadina-1(6),4-diene; Δ-cadinene; β-copaene; azoarene; epicubenol; cadin-4-en-10-ol | II | ||
| TvSTS06 | LC712913 | cadina-1,4-diene; cadina-1(6),4-diene; Δ-cadinene; β-copaene; β-farnesene; zonarene; epicubenol | III | ||
| TvSTS07 | LC712914 | Δ6-protoilludene | III | ||
| TvSTS12 | LC712917 | γ-cadinene | IV | ||
| TvSTS14 | LC712919 | β-barbatene; α-barbatene | IV | ||
| TvSTS16 | LC712920 | dauca-4(11),8-diene; isobazzanene | IV | ||
| Irpex lacteus | IIIS | JGI ID: Il4946 | iltremulanol A | III | [57] |
| Serendipita indica | SiTPS | JGI ID: 77541 | viridiflorol | III | [58] |
| Termitomyces sp. T153 | DS3 | not available | unknown sesquiterpene | IV | [59] |
| Laccaria bicolor | LbSTS4a | XP_001887869.1 | (E)-nerolidol | II | [60] |
| LbSTS6 | XP_001885710.1 | α-cuprenene; α-cuparene | IV |
| Sequence Name | Motif I D(D/E/N)xx(D/E) | Motif II (NDxxSxxxE) | Domains (Pfam ID) |
|---|---|---|---|
| LdSTS1 | DELSD | NDLYSYNME | PF19086 |
| LdSTS2 | DEYTD | QDLYSYNNE | PF19086 |
| LdSTS3 | DEVSD | NDVYSYNME | PF19086 |
| LdSTS6 | DNVSD | NDIFSYNVE | PF19086; PF03936; PF06330 |
| LdSTS7 | DEYTD | NDLYSYNIE | PF19086 |
| LdSTS8 | DEFSD | NDIASYNVE | PF19086 |
| LdSTS10 | DEFSE | NDIASYNVE | PF19086 |
| LdSTS11 | DEYTD | NDLYSYNIE | PF19086 |
| LdSTS12 | DEYTD | NDLYSYNVE | PF19086 |
| LdSTS14 | DEFTD | NDMYSYNIE | PF19086 |
| Cop1 | DNLSD | NDIFSFNVE | PF19086 |
| Cop2 | DDWLD | NDIFSFNRE | PF19086 |
| Cop3 | DNISD | NDIFSYNVE | PF19086; PF03936; PF06330; PF19035 |
| Cop4 | DEISD | NDVYSYDME | PF19086; PF03936 |
| Cop5 | DYFFD | NDAYSWNVE | PF19086; PF03936 |
| Cop6 | DDAFQ | NDLLSFYKE | PF06330 |
| Omp1 | DNLTD | NDIYSFNIE | PF19086 |
| Omp2 | DNLSD | NDIFSYNVE | PF19086 |
| Omp3 | DEVSD | NDIFSYNVE | PF19086 |
| Omp4 | DEVSD | NDVYSYNKE | PF19086 |
| Omp5a | DELSD | NDVYSYNVE | PF19086 |
| Omp5b | DEVSD | NDVYSYNVE | PF19086 |
| Omp6 | DEYSD | NDLCSYNVE | PF19086 |
| Omp7 | DEYSD | NDTASYNYE | PF19086 |
| Omp8 | DDVFE | NDIMSFYKE | PF06330 |
| Omp9 | DDVFE | NDVLSFYKE | PF06330 |
| Omp10 | DDIFP | NDVLSFYKE | PF06330 |
| ShSTS1 | DDSLE | NDLMSFYKE | PF06330 |
| ShSTS11 | DEISD | NDVYSYNVE | PF19086 |
| ShSTS18 | DEYSD | QDICSYNVE | PF19086 |
| ShSTS15 | DEHSD | NDIVSYNIE | PF19086 |
| ShSTS16 | DEYSD | NDIVSYNLE | PF19086 |
| ShSTS3 | DDWVD | NEASSYVKE | PF19086 |
| ShSTS4 | DDYID | NDFFSYLKE | PF19086 |
| ShSTS5 | DDLSD | NDLCSFNKE | PF19086 |
| ShSTS7 | DDWTD | NDIFSYNVE | PF19086 |
| ShSTS8 | DEISD | NDIYSYDME | PF19086 |
| ShSTS10 | DEISD | NDVYSYKVE | PF19086 |
| ShSTS12 | DEISD | QDVYSYSME | PF19086 |
| ShSTS13 | DDILD | NDTFSYRRE | PF19086 |
| ShSTS17 | DEHSD | SDIVSWNLE | PF19086 |
| HS-HMGS | DDYID | NDFFSYLKE | PF19086; PF01154 |
| CpSTS1 | DEYTD | NDMCSYKKE | PF19086 |
| CpSTS2 | DELSD | NDVYSYDME | PF19086 |
| CpSTS3 | DNISD | NDIFSYNVE | PF19086 |
| CpSTS4 | DEYTD | NDLCSFRNE | PF19086 |
| CpSTS5 | DNLSD | NDIFSYNVE | PF19086 |
| CpSTS6 | DEYSD | NDLYSYNVE | PF19086 |
| CpSTS7 | DEITE | NDVFSFKVE | PF19086 |
| CpSTS8 | DEYTD | NDVYSYNME | PF19086 |
| CpSTS9 | DEYTD | NDLFSYNME | PF19086 |
| CpSTS11 | DEATD | NDIHSYNME | PF19086 |
| CpSTS12 | DEYTD | NDLYSYNME | PF19086 |
| CpSTS13 | DETTD | NDIQSYNME | PF19086 |
| CpSTS14 | DDYIL | NDIYSYKVE | PF19086 |
| CpSTS15 | DDLME | NDLFSYRKE | PF19086 |
| CpSTS16 | DESSD | NDIHSYNME | PF19086 |
| CpSTS17 | DDIIE | NDLFSYRVE | PF19086 |
| CpSTS18 | DDLSD | NDLCSFNKE | PF19086; PF03936; PF06330; |
| AcTPS4 | DEVSD | NDVYSYNME | PF19086 |
| AcTPS5 | DDWTD | NDVLSYNAE | PF19086; PF03936 |
| AcTPS9 | DEISD | NDLYSYNME | PF19086 |
| Tps1A | DIEGD | QDFPDIEFD | PF01040 |
| Tps2A | DVAGD | QDFPDIEFD | PF01040 |
| AncA | DDRIE | DDFTDD | PF13419 |
| AncC | DDKIE | DDFTDD | PF13419 |
| BvCS | DNISD | NDVFSYNVE | PF19086; PF03936 |
| GME3634 | DDWTD | NDVLSYNAE | PF19086 |
| GME3638 | DDWSD | NDLFSYNVE | na |
| GME9210 | DEYSD | NDIVSYNVE | PF19086 |
| Agr1 | DNLSD | NDIFSYSVE | PF19086; PF03936 |
| Agr2 | DEVTD | NDLYSYNME | PF19086 |
| Agr3 | DNISD | NDIFSYNVE | PF19086; PF03936 |
| Agr4 | DEVSD | NDVYSYDME | PF19086 |
| Agr5 | DEYTD | NDLVSYNME | PF19086 |
| Agr6 | DEHTD | NDLCSYNVE | PF19086 |
| Agr7 | DEWSD | NDLCSYNVE | PF19086 |
| Agr8 | DEYTD | NDMHSYVRE | PF19086; PF03936 |
| Agr9 | DEYTD | NDIDSYAME | PF19086 |
| Agr10 | DECAD | na | na |
| Agr11 | DEYTD | na | PF19086 |
| Copu1 | DELTD | NDVYSYNME | PF19086 |
| Copu2 | DDLTD | NDVFSYNRE | PF19086 |
| Copu3 | DELSD | NDVYSYNME | PF19086 |
| Copu5 | DDWSD | NDVFSYNKE | PF19086; PF03936 |
| Copu9 | DDWLD | NDIFSYNKE | PF19086 |
| Pilcr_825684 | DELTD | NDLFSYNRE | PF19086 |
| Galma_104215 | DEFTD | NDLFSYDME | PF19086 |
| Sphst_47084 | DEYTD | NDLFSYNS | PF19086 |
| Denbi1_816208 | DEFTD | NDLFSYNME | PF19086 |
| Denbi1_659367 | DEHSD | NDLCSYNVE | PF19086 |
| Hetan2_454193 | DEYSD | NDIASYNLE | PF19086 |
| Hypsu1_138665 | DEHTD | NDLCSYKVE | PF19086; PF03936 |
| Pro1 | DEYSD | NDVVSYNLE | PF19086; PF03936 |
| Hfas94a | DEYTD | NDMHSYGLE | PF19086 |
| Hfas94b | DEDLD | NDLISYTKE | PF19086 |
| Hfas344 | DEYTD | NDMHSYALE | PF19086 |
| Cun3817 | DDLSD | NDLCSFNKE | PF19086; PF03936 |
| Cun5155 | DEHSD | NDLFSYNVE | PF19086 |
| Cun3157 | DEISD | NDIYSYNME | PF19086 |
| Cun3158 | DEVSD | NDVYSYNME | PF19086 |
| Cun0773 | DDWSD | NDILSYSKE | PF19086 |
| Cun7050 | DNISD | NDIFSYNVE | PF19086; PF03936 |
| Cun0716 | DDWSD | NDIFSFNVE | PF19086 |
| Cun0759 | DDWSD | NDIFSYNKE | PF19086 |
| Cun3574 | DDWTD | NDIFSYNKE | PF19086 |
| Cun9106 | NDDYE | na | PF06148 |
| PpSTS01 | DNISD | NDIFSYNVE | PF19086 |
| PpSTS03 | DDWSD | NDILSYNRE | PF19086; PF03936 |
| PpSTS06 | DDITD | NDIYSFNNE | PF19086 |
| PpSTS08 | DEYTD | NDLVSYNRE | PF19086 |
| PpSTS09 | DEYSD | NDMLSWNVE | PF19086 |
| PpSTS10 | DEVSD | NDVYSYNME | PF19086 |
| PpSTS14 | DEYTD | NDIASYNKE | PF19086 |
| PpSTS29 | DEPDI | NDILSFYKE | PF06330 |
| Fompi1 | DDPDI | NDILSFYKE | PF06330 |
| PcSTS01 | DNISD | NDIFSYNVE | PF19086; PF03936 |
| PcSTS02 | DEVSD | NDVYSYKME | PF19086 |
| PcSTS03 | DEISD | NDVYSYDME | PF19086 |
| PcSTS04 | DEISD | NDVYSYDME | PF19086 |
| PcSTS06 | DDFEI | NDLLSFYKE | PF06330 |
| PcSTS08 | DDEAI | NDILSFYKE | PF06330 |
| PcSTS11 | DDCEI | NDIYSFHKE | PF06330 |
| A8411 | DDYID | NDLFSYAKE | PF19086 |
| GsSTS43 | DDLSD | NDLCSFNKE | PF19086; PF03936 |
| GsSTS45a | DDLSD | NDLCSFNKE | PF19086 |
| GsSTS45b | DDLSD | NDLCSFNKE | PF19086 |
| GsSTS26 | DEYTD | NDVASYNRE | PF19086 |
| GsSTS27 | DEYTD | NDVASYNRE | PF19086 |
| GS02363 | DDWTD | NDVLSYNAE | PF19086; PF03936 |
| GS14272 | DNISD | NDIFSYNVE | PF19086 |
| GS11330 | DDLGE | NDILSFYKE | PF06330 |
| GL26009 | DDWTD | NDVLSYNAE | PF19086; PF03936 |
| GLSTS6 | DDLSD | NDLCSFNKE | PF19086; PF03936 |
| STC4 | DRLTD | NDLYSYKKE | PF19086 |
| STC9 | DDLSD | NDLCSFNKE | PF19086; PF03936 |
| STC15 | DNLSD | NDIFSYNVE | PF19086; PF03936 |
| AbSTS05 | DEISD | NDVYSYNVE | PF19086 |
| AbSTS07 | DDNFD | NDITSFYKE | PF06330 |
| AbSTS09 | DDNYD | NDIASFYKE | PF06330 |
| AvSTS01 | DEYTD | NDLCSYNKE | PF19086; PF03936 |
| AvSTS03 | DEYSD | NDIASYNLE | PF19086; PF03936 |
| AvSTS06 | DEFTD | NDTYSYNIE | PF19086 |
| AvSTS07 | DEFTD | NDTYSYNIE | PF19086 |
| AvSTS09 | DEVSD | NDVYSYNME | PF19086 |
| LnSTS01 | DEYSD | NDLCSYNVE | PF19086 |
| LnSTS02 | DEHSD | NDLCSYNVE | PF19086 |
| LnSTS04 | DEYSD | NDVYSYNKE | PF19086; PF03936 |
| LnSTS09 | DELSD | NDVYSYDME | PF19086 |
| LnSTS19 | DDVDS | NDLLSYHKE | PF06330 |
| LnSTS20 | DDMSS | NDILSFHKE | PF06330 |
| LnSTS25 | DDTSP | NDLMSFPKE | PF19086; PF06330 |
| LnSTS27 | DDKYF | NDIMSFYKE | PF06330 |
| PoSTS01 | DNLSD | NDIFSYNVE | PF19086; PF03936 |
| PoSTS02 | DDWLD | NDLFSYNVE | PF19086; PF03936 |
| PoSTS03 | DNISD | NDIFSYNVE | PF19086; PF03936 |
| PoSTS05 | DEVSD | NDVYSYNME | PF19086 |
| PoSTS06 | DEFSD | NDVYSWNVE | PF19086 |
| PoSTS11 | EEITE | NDIYSYKKE | PF19086 |
| PoSTS16 | DDISS | NDVLSFYKE | PF06330 |
| TvSTS01 | DNICD | NDIFSYNVE | PF19086 |
| TvSTS05 | DEISD | NDLYSYNME | PF19086 |
| TvSTS06 | DEVSD | NDVYSYNME | PF19086 |
| TvSTS07 | DEQTD | NDLLSYRKE | PF19086; PF03936 |
| TvSTS12 | DDLSD | NDLCSFNKE | PF19086; PF03936 |
| TvSTS14 | DDLGG | NDILSFYKE | PF06330 |
| TvSTS16 | DDLPG | NDLLSFYKE | PF06330 |
| IIIS | DEYTD | NDIASYNKE | PF19086 |
| SiTPS | DDLMD | NDVYSFDNE | PF19086; PF03936 |
| DS3 | DDKLE | DLDTT | PF13419 |
| LbSTS4a | DDITD | NDVYSYGKE | PF19086 |
| LbSTS6 | DDVFQ | NDVLSFYKE | PF06330 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Chen, R.; Yuan, L.; Zhang, C.; Liang, D.; Qiao, J. Molecular and Functional Analyses of Characterized Sesquiterpene Synthases in Mushroom-Forming Fungi. J. Fungi 2023, 9, 1017. https://doi.org/10.3390/jof9101017
Wang S, Chen R, Yuan L, Zhang C, Liang D, Qiao J. Molecular and Functional Analyses of Characterized Sesquiterpene Synthases in Mushroom-Forming Fungi. Journal of Fungi. 2023; 9(10):1017. https://doi.org/10.3390/jof9101017
Chicago/Turabian StyleWang, Shengli, Ruiqi Chen, Lin Yuan, Chenyang Zhang, Dongmei Liang, and Jianjun Qiao. 2023. "Molecular and Functional Analyses of Characterized Sesquiterpene Synthases in Mushroom-Forming Fungi" Journal of Fungi 9, no. 10: 1017. https://doi.org/10.3390/jof9101017