
Transcriptome Analysis Reveals Mycelial and Fruiting Responses to Lithium Chloride in Coprinopsis cinerea

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Cultivation Conditions
2.2. RNA Isolation, Library Preparation, and Sequencing
2.3. Read Alignment and Transcript Assembly
2.4. Differential Expression Analysis and Functional Gene Annotation
2.5. GO and KEGG Enrichment Analysis
2.6. cDNA Synthesis and qRT-PCR Validation
2.7. Statistical Analysis
3. Results and Discussion
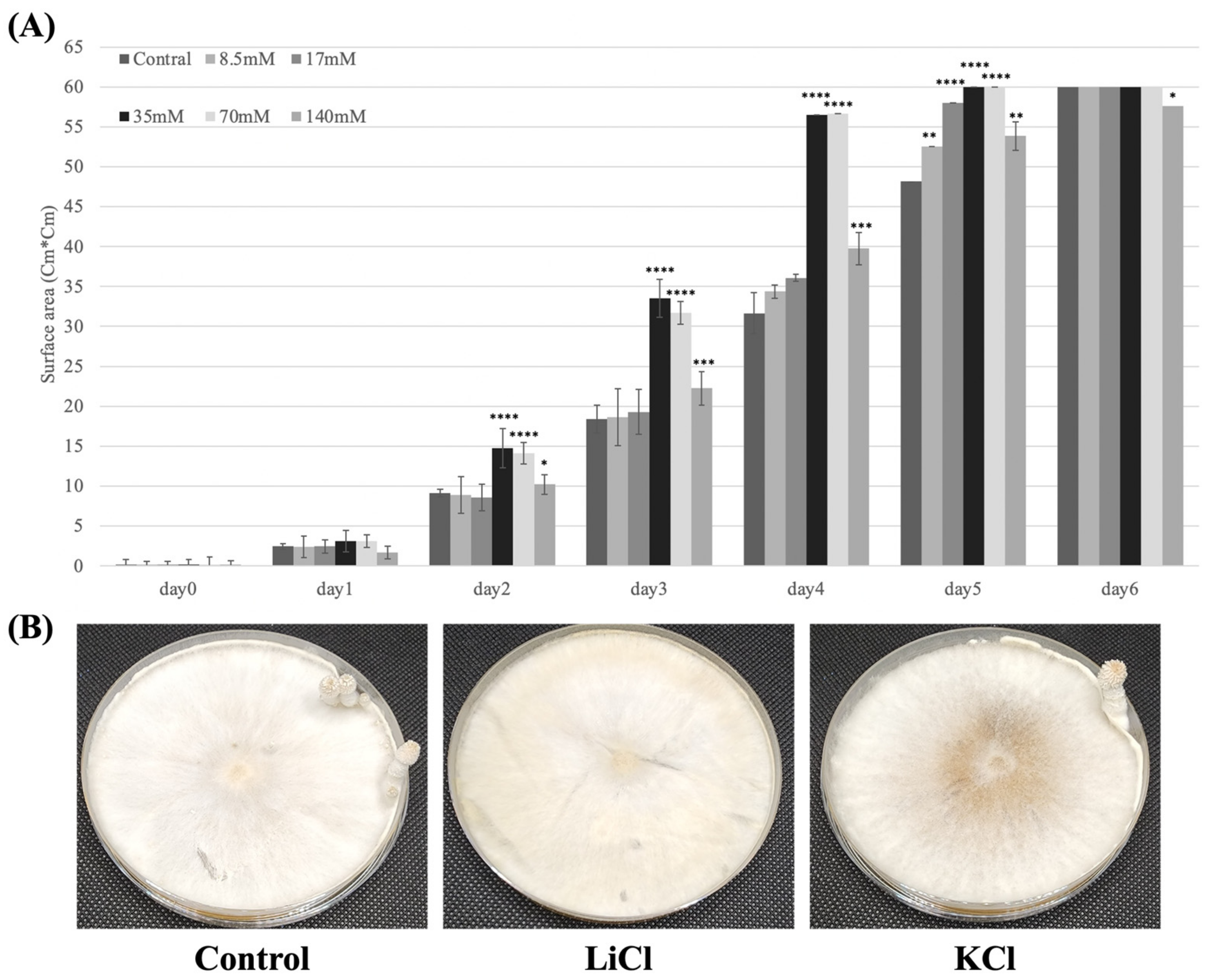
3.1. Effects of LiCl on Mycelium Growth
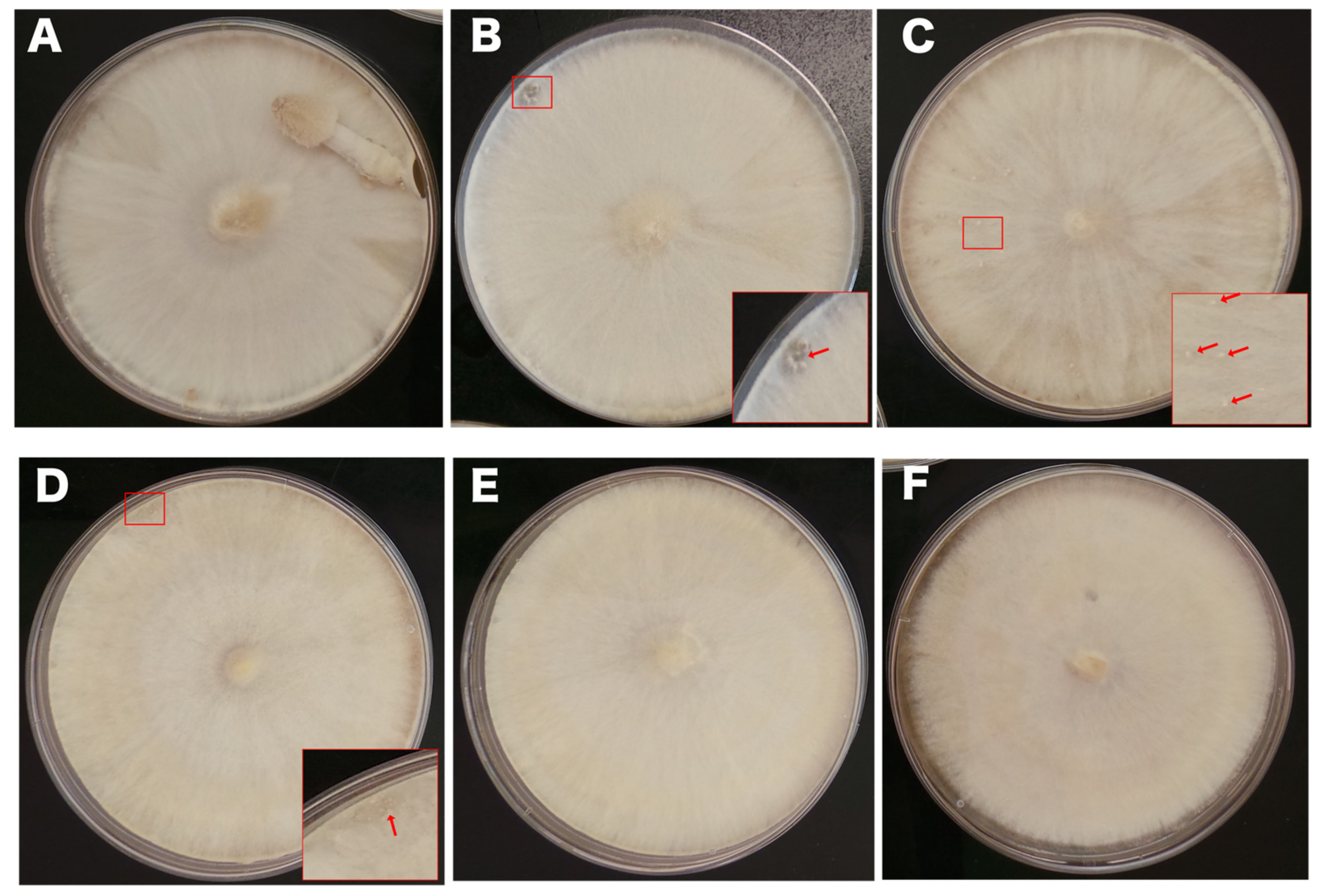
3.2. Effects of LiCl on Fruiting Body Initiation
3.3. Sequencing of the C. cinerea Transcriptome
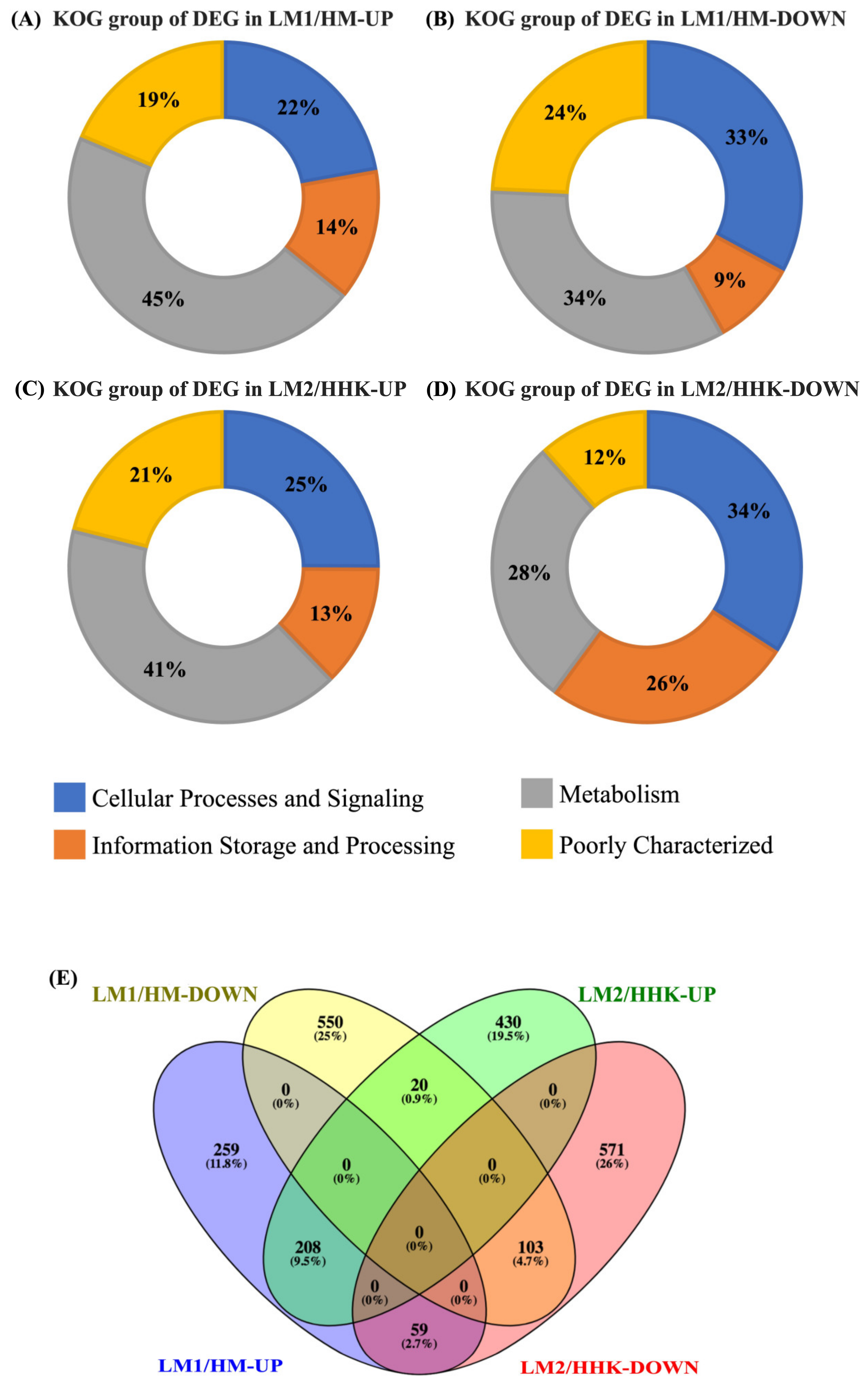
3.4. Differential Expression of Genes in Mycelium under the Influence of LiCl
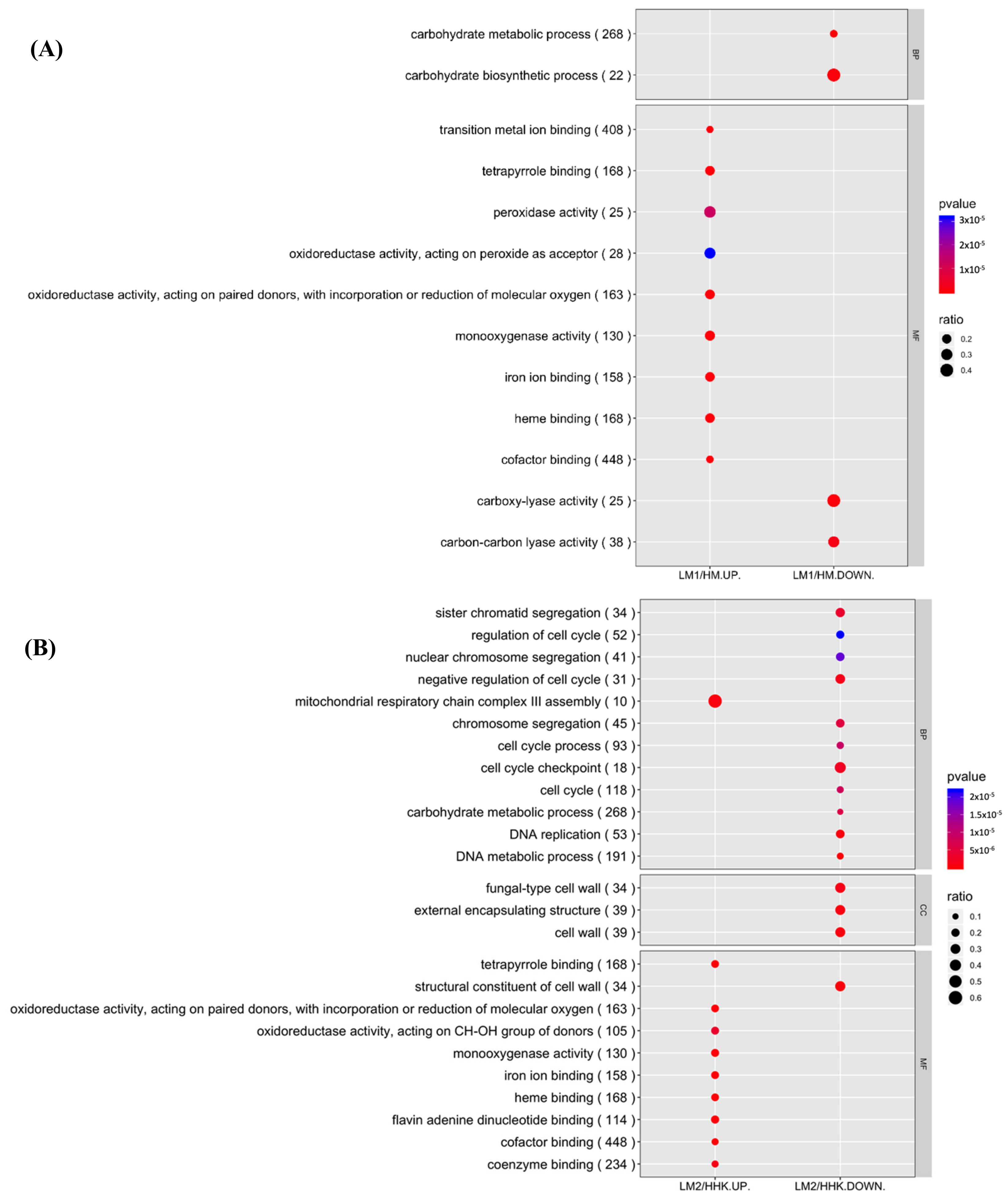
3.5. GO Functional Enrichment Analysis Explained the Fast Mycelial Growth under the Influence of LiCl by Increased Oxidative Activities
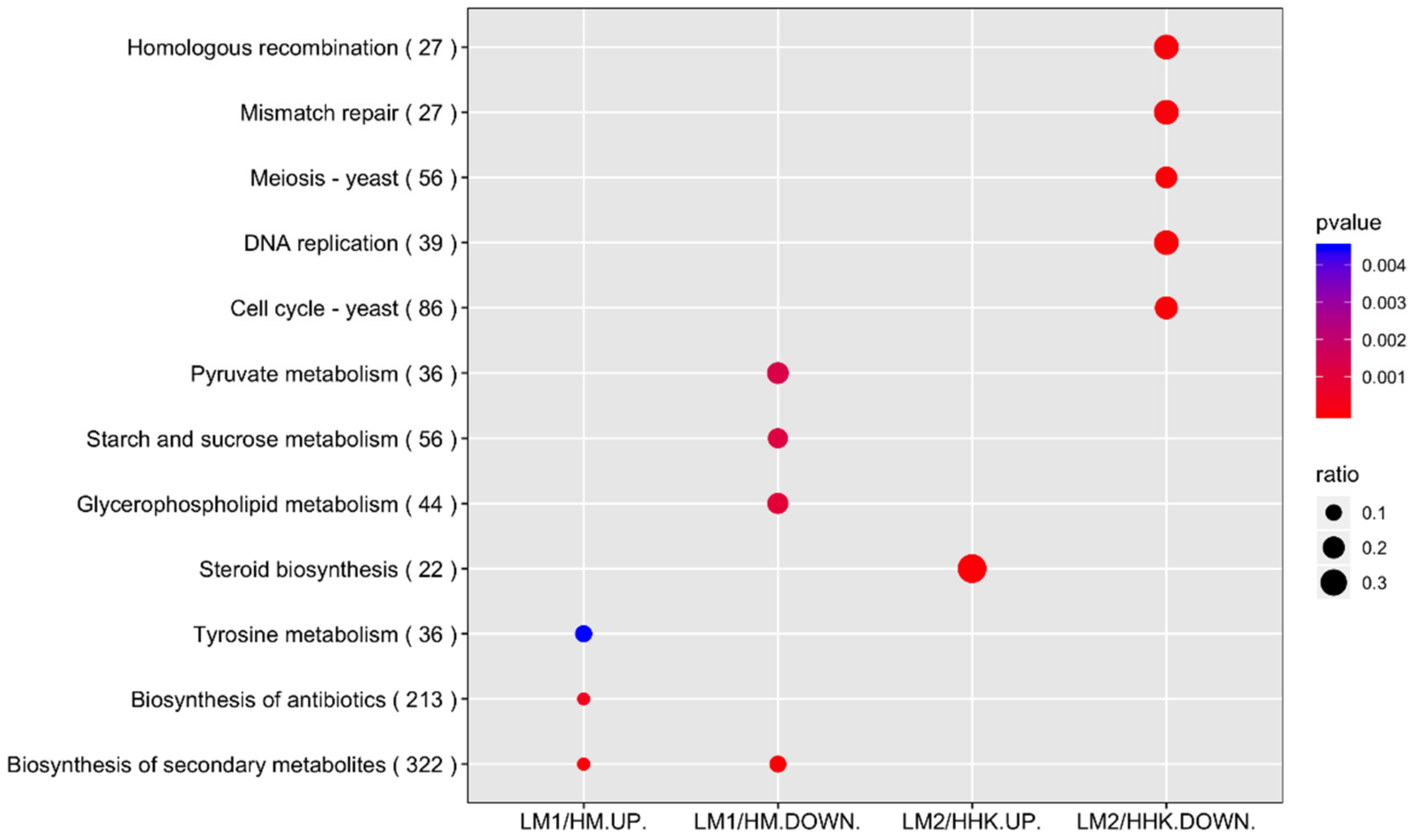
3.6. KEGG Pathway Enrichment Analysis Revealed an Association between Metabolism Pathways and Enriched Mycelial Growth under LiCl Influence
3.7. Differential Expression of Genes in Fruiting Body Initiation under the Influence of LiCl
3.8. GO Functional Enrichment Analysis Revealed Most Downregulated Biological Processes in Fruiting Body Initiation in LiCl-Treated Samples
3.9. KEGG Pathway Enrichment Analysis Revealed an Association between Cellular Process Pathways and Inhibited Fruiting Body Initiation under LiCl Influence
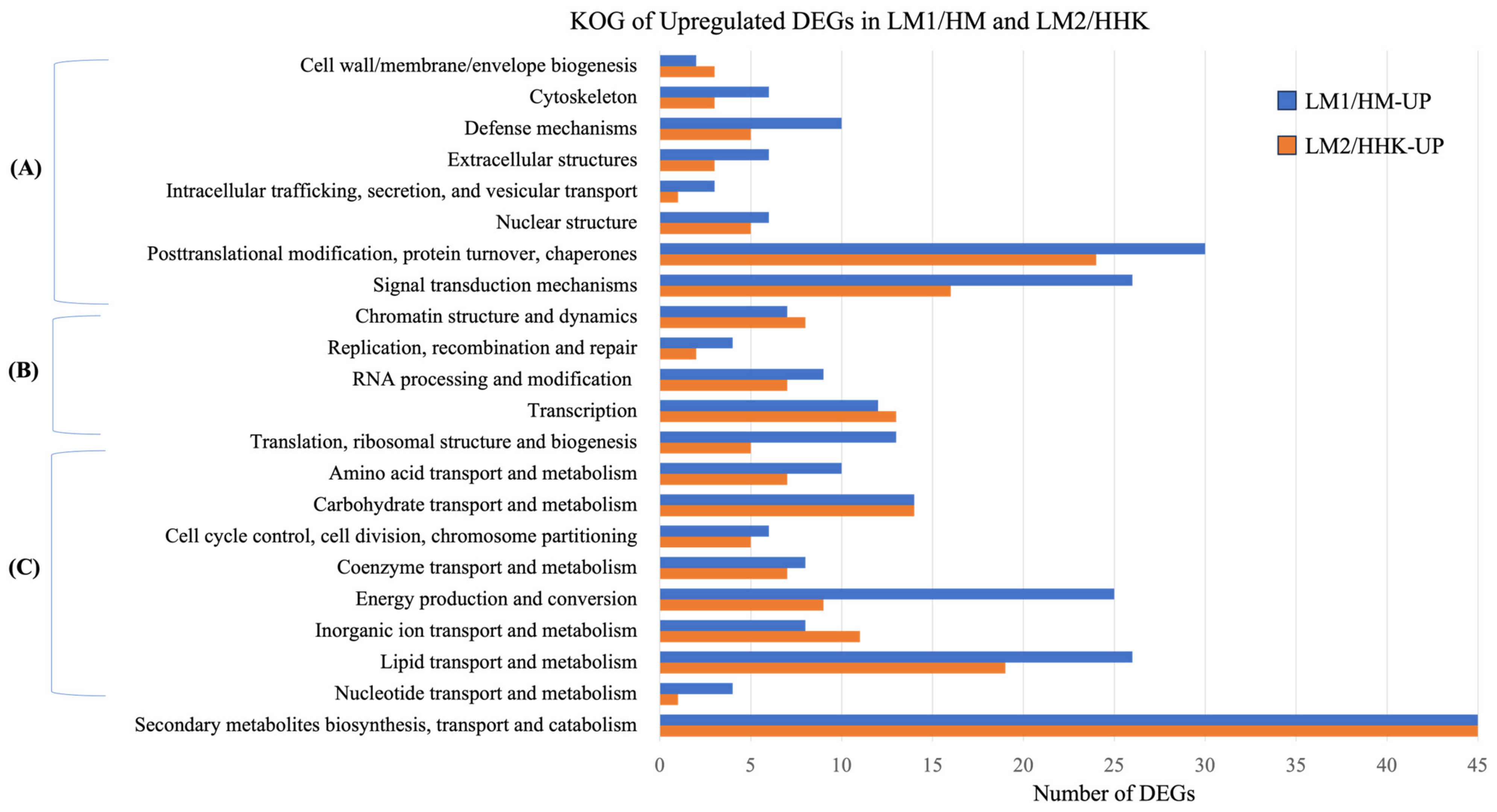
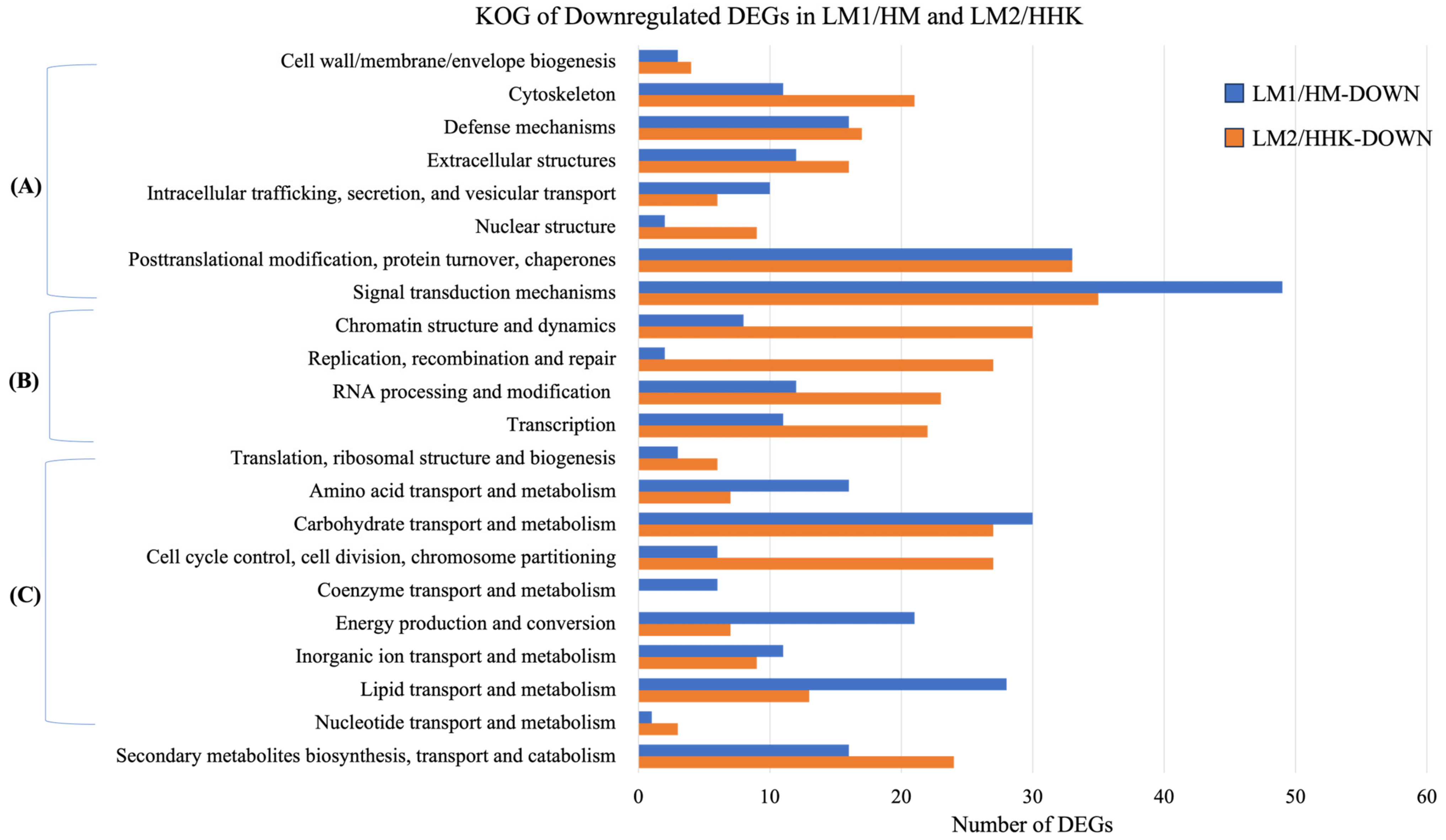
3.10. Downregulated DEGs Were Associated with the Inhibition of Fruiting Body Development
3.11. LiCl Inhibited Signal Transduction Responding to Environmental Factors
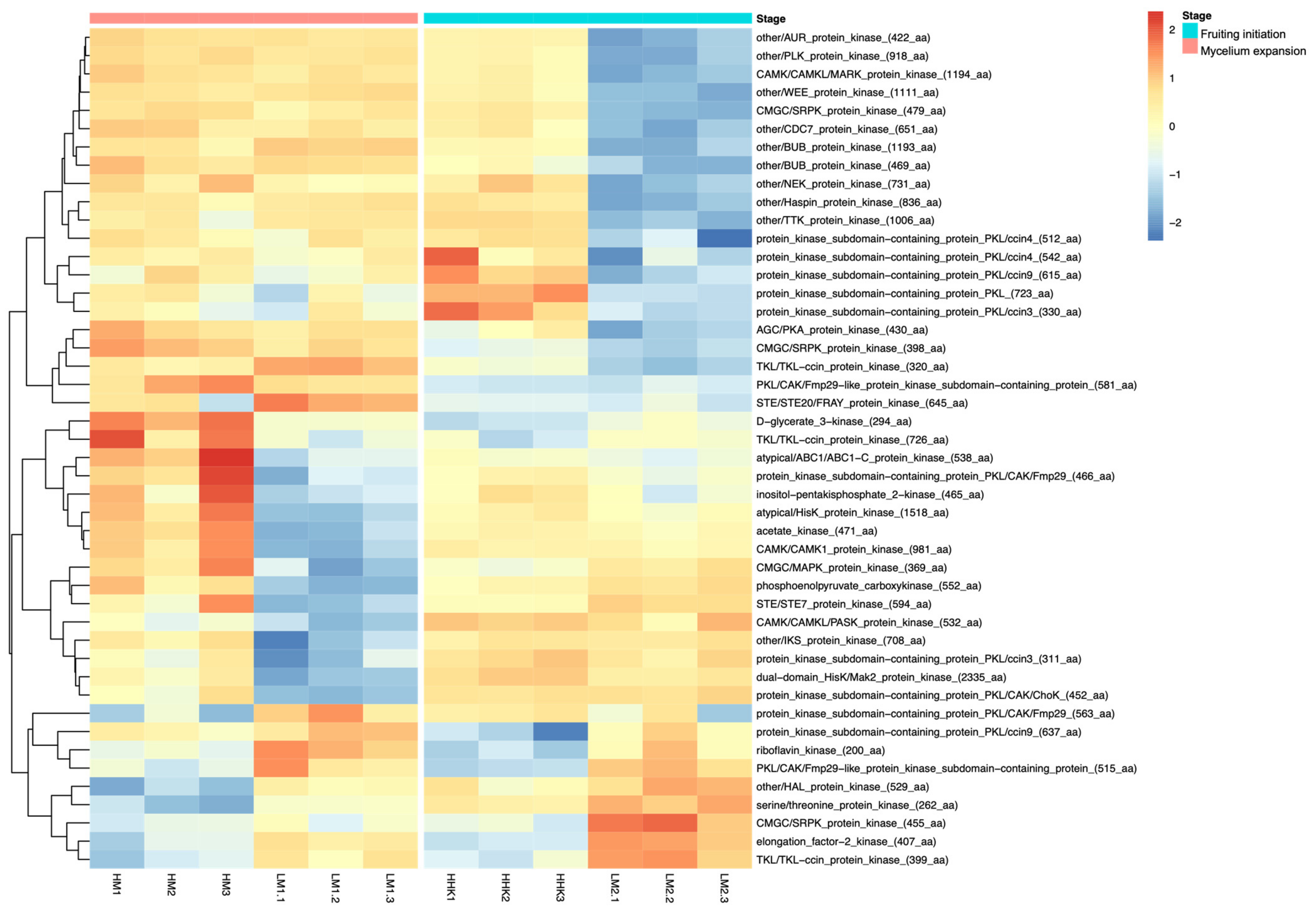
3.12. Kinases Were Downregulated in Mycelium and Fruiting Body Initiation
3.13. The Effect of LiCl Revealed That the Activity of GSK3 Could Be an Important Channel in Fruiting Body Development
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kues, U. Life History and Developmental Processes in the Basidiomycete Coprinus cinereus. Microbiol. Mol. Biol. Rev. 2000, 64, 316–353. [Google Scholar] [CrossRef]
- Sakamoto, Y. Influences of environmental factors on fruiting body induction, development and maturation in mushroom-forming fungi. Fungal Biol. Rev. 2018, 32, 236–248. [Google Scholar] [CrossRef]
- Muraguchi, H.; Umezawa, K.; Niikura, M.; Yoshida, M.; Kozaki, T.; Ishii, K.; Sakai, K.; Shimizu, M.; Nakahori, K.; Sakamoto, Y.; et al. Strand-specific RNA-seq analyses of fruiting body development in Coprinopsis cinerea. PLoS ONE 2015, 10, e0141586. [Google Scholar] [CrossRef]
- Cheng, C.K.; Au, C.H.; Wilke, S.K.; Stajich, J.E.; Zolan, M.E.; Pukkila, P.J.; Kwan, H.S. 5′-Serial Analysis of Gene Expression studies reveal a transcriptomic switch during fruiting body development in Coprinopsis cinerea. BMC Genom. 2013, 14, 195. [Google Scholar] [CrossRef]
- Lu, B.C. The control of meiosis progression in the fungus Coprinus cinereus by light/dark cycles. Fungal Genet. Biol. 2000, 31, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Stajich, J.E.; Wilke, S.K.; Ahren, D.; Au, C.H.; Birren, B.W.; Borodovsky, M.; Burns, C.; Canbäck, B.; Casselton, L.A.; Cheng, C.K.; et al. Insights into evolution of multicellular fungi from the assembled chromosomes of the mushroom Coprinopsis cinerea (Coprinus cinereus). Proc. Natl. Acad. Sci. USA 2010, 107, 11889–11894. [Google Scholar] [CrossRef] [PubMed]
- Budavari, S.; O’Neil, M.J.; Smith, A.; Heckelman, P.E. (Eds.) The Merck Index: An Encyclopedia of Chemicals, Drugs and Biologicals, 11th ed.; Merck: Rahway, NJ, USA, 1989. [Google Scholar]
- Faria, M.G.I.; do Valle, J.S.; Lopes, A.D.; Gonçalves, A.C., Jr.; Dragunski, D.C.; Colauto, N.B.; Linde, G.A. Bioaccumulation of lithium (Li2CO3) in mycelia of the culinary-medicinal oyster mushroom, Pleurotus ostreatus (Agaricomycetes). Int. J. Med. Mushrooms 2018, 20, 901–907. [Google Scholar] [CrossRef]
- Mateus, D.N.; Willian, L.C.; Jos, M.D.L.; Maria, C.K. Lithium chloride affects mycelial growth of white rot fungi: Fungal screening for Li-enrichment. Afr. J. Microbiol. Res. 2014, 8, 2111–2123. [Google Scholar] [CrossRef]
- Richter, D.L.; Robinson, S.C.; Beardslee, M.P.; Habarth, M.L. Differential sensitivity of fungi to lithium chloride in culture media. Mycol. Res. 2008, 112, 717–724. [Google Scholar] [CrossRef]
- Martins, L.F.; Montero-Lomelí, M.; Masuda, C.A.; Fortes, F.S.; Previato, J.O.; Mendonça-Previato, L. Lithium-mediated suppression of morphogenesis and growth in Candida albicans. FEMS Yeast Res. 2008, 8, 615–621. [Google Scholar] [CrossRef]
- Masuda, C.A.; Ramírez, J.; Peña, A.; Montero-Lomelí, M. Regulation of monovalent ion homeostasis and pH by the SerThr protein phosphatase SIT4 in Saccharomyces cerevisiae. J. Biol. Chem. 2000, 275, 30957–30961. [Google Scholar] [CrossRef]
- Chang, J.; Chan, P.L.; Xie, Y.; Ma, K.L.; Cheung, M.K.; Kwan, H.S. Modified recipe to inhibit fruiting body formation for living fungal biomaterial manufacture. PLoS ONE 2019, 14, e0209812. [Google Scholar] [CrossRef]
- Li, L.; Song, H.; Zhong, L.; Yang, R.; Yang, X.Q.; Jiang, K.L.; Liu, B.Z. Lithium chloride promotes apoptosis in human leukemia NB4 cells by inhibiting glycogen synthase kinase-3 beta. Int. J. Med. Sci. 2015, 12, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, R.; Jiang, C.; Zhao, S.; Li, W.; Tang, X. The neuroprotective effect of lithium chloride on cognitive impairment through glycogen synthase kinase-3β inhibition in intracerebral hemorrhage rats. Eur. J. Pharmacol. 2018, 840, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.T.; Chuang, D.M. Molecular actions and therapeutic potential of lithium in preclinical and clinical studies of CNS disorders. Pharmacol. Ther. 2010, 128, 281–304. [Google Scholar] [CrossRef]
- Rao, P.S.; Niederpruem, D.J. Carbohydrate metabolism during morphogenesis of Coprinus lagopus (sensu Buller). J. Bacteriol. 1969, 100, 1222–1228. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Pachter, L.; Salzberg, S.L.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; et al. Transcript assembly and abundance estimation from RNA-Seq reveals thousands of new transcripts and switching among isoforms. Nat. Biotechnol. 2011, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L.; et al. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R Package for Comparing Biological Themes among Gene Clusters. Omics J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Li, Q.; Wang, Q.; Lu, D.; Zhang, H.; Wang, J.; Fu, R. Transcriptional profiling provides new insights into the role of nitric oxide in enhancing Ganoderma oregonense resistance to heat stress. Sci. Rep. 2017, 7, 15694. [Google Scholar] [CrossRef]
- Yang, D.-D.; François, J.M.; de Billerbeck, G.M. Cloning, expression and characterization of an aryl-alcohol dehydrogenase from the white-rot fungus Phanerochaete chrysosporium strain BKM-F-1767. BMC Microbiol. 2012, 12, 126. [Google Scholar] [CrossRef] [PubMed]
- Ichinose, H.; Wariishi, H.; Tanaka, H. Molecular analysis of arylalcohol dehydrogenase of Coriolus versicolor expressed against exogenous addition of dibenzothiophene derivatives. J. Basic Microbiol. 2002, 42, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.D.; de Billerbeck, G.M.; Zhang, J.J.; Rosenzweig, F.; Francois, J.M. Deciphering the origin, evolution, and physiological function of the subtelomeric aryl-alcohol dehydrogenase gene family in the yeast Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2018, 84, e01553-17. [Google Scholar] [CrossRef] [PubMed]
- De Paula, R.; Azzariti de Pinho, C.; Terenzi, H.; Bertolini, M. Molecular and biochemical characterisation of the Neurospora crassa glycogen synthase encoded by the gsn cDNA. Mol. Genet. Genom. 2002, 267, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Douglas, C. Fungal b (1,3)-D-glucan synthesis. Med. Mycol. 2001, 39, 55–66. [Google Scholar] [CrossRef]
- Foreman, P.K.; Brown, D.; Dankmeyer, L.; Dean, R.; Diener, S.; Dunn-Coleman, N.S.; Goedegebuur, F.; Houfek, T.D.; England, G.J.; Kelley, A.S.; et al. Transcriptional regulation of biomass-degrading enzymes in the filamentous fungus Coprinus lagopus. J. Biol. Chem. 2003, 278, 31988–31997. [Google Scholar] [CrossRef]
- Walser, P.J.; Haebel, P.W.; Künzler, M.; Sargent, D.; Kües, U.; Aebi, M.; Ban, N. Structure and functional analysis of the fungal galectin CGL2. Structure 2004, 12, 689–702. [Google Scholar] [CrossRef]
- Bertossa, R.C.; Kües, U.; Aebi, M.; Künzler, M. Promoter analysis of cgl2, a galectin encoding gene transcribed during fruiting body formation in Coprinopsis cinerea (Coprinus cinereus). Fungal Genet. Biol. 2004, 41, 1120–1131. [Google Scholar] [CrossRef]
- Boulianne, R.P.; Liu, Y.; Aebi, M.; Lu, B.C.; Kues, U. Fruiting body development in Coprinus cinereus: Regulated expression of two galectins secreted by a non-classical pathway. Microbiology 2000, 146, 1841–1853. [Google Scholar] [CrossRef]
- Nakazawa, T.; Ando, Y.; Hata, T.; Nakahori, K. A mutation in the Cc.arp9 gene encoding a putative actin-related protein causes defects in fruiting initiation and asexual development in the agaricomycete Coprinopsis cinerea. Curr. Genet. 2016, 62, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Ando, Y.; Nakazawa, T.; Oka, K.; Nakahori, K.; Kamada, T. Cc.snf5, a gene encoding a putative component of the SWI/SNF chromatin remodeling complex, is essential for sexual development in the agaricomycete Coprinopsis cinerea. Fungal Genet. Biol. 2013, 50, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Muraguchi, H.; Abe, K.; Nakagawa, M.; Nakamura, K.; Yanagi, S.O. Identification and characterisation of structural maintenance of chromosome 1 (smc1) mutants of Coprinopsis cinerea. Mol. Genet. Genom. 2008, 280, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Tsusué, Y.M. Experimental control of fruit-body formation in Coprinus macrorhizus. Dev. Growth Differ. 1969, 11, 164–178. [Google Scholar] [CrossRef] [PubMed]
- Stamets, P. Growing Gourmet and Medicinal Mushrooms, 3rd ed.; Ten Speed Press: Berkeley, CA, USA, 2000. [Google Scholar]
- Palmer, G.E.; Horton, J.S. Mushrooms by magic: Making connections between signal transduction and fruiting body development in the basidiomycete fungus Schizophyllum commune. FEMS Microbiol. Lett. 2006, 262, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Alspaugh, J.A.; Pukkila-Worley, R.; Harashima, T.; Cavallo, L.M.; Funnell, D.; Cox, G.M.; Perfect, J.R.; Kronstad, J.W.; Heitman, J. Adenylyl cyclase functions downstream of the Gα protein Gpa1 and controls mating and pathogenicity of Cryptococcus neoformans. Eukaryot. Cell 2022, 1, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, K.; Kimura, T.; Suzuki, M.; Yamaki, K.J.; Oita, S. Identification and overexpression of genes encoding cAMP-dependent protein kinase catalytic subunits in homobasidiomycete Schizophyllum commune. Biosci. Biotechnol. Biochem. 2005, 69, 2333–2342. [Google Scholar] [CrossRef]
- Linden, H.; Ballario, P.; Macino, G. Blue light regulation in Neurospora crassa. Fungal Genet. Biol. 1997, 22, 141–150. [Google Scholar] [CrossRef]
- Linden, H. A white collar protein senses blue light. Science 2002, 297, 777–779. [Google Scholar] [CrossRef]
- Dunlap, J.C. Proteins in the Neurospora Circadian Clockworks. J. Biol. Chem. 2006, 281, 28489–28493. [Google Scholar] [CrossRef]
- Selenica, M.L.; Jensen, H.S.; Larsen, A.K.; Pedersen, M.L.; Helboe, L.; Leist, M.; Lotharius, J. Efficacy of small-molecule glycogen synthase kinase-3 inhibitors in the postnatal rat model of tau hyperphosphorylation. Br. J. Pharmacol. 2007, 152, 959–979. [Google Scholar] [CrossRef]
- Terashima, K.; Yuki, K.; Muraguchi, H.; Akiyama, M.; Kamada, T. The Dst1 gene involved in mushroom photomorphogenesis of Coprinus cinereus encodes a putative photoreceptor for blue light. Genetics 2005, 171, 101–108. [Google Scholar] [CrossRef]
- Kuratani, M.; Tanaka, K.; Terashima, K.; Muraguchi, H.; Nakazawa, T.; Nakahori, K.; Kamada, T. The Dst2 gene essential for photomorphogenesis of Coprinopsis cinerea encodes a protein with a putative FAD-binding-4 domain. Fungal Genet. Biol. 2010, 47, 152–158. [Google Scholar] [CrossRef]
- Nakazawa, T.; Ando, Y.; Kitaaki, K.; Nakahori, K.; Kamada, T. Efficient gene targeting in ΔCc. ku70 or ΔCc. lig4 mutants of the agaricomycete Coprinopsis cinerea. Fungal Genet. Biol. 2011, 48, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Guo, M.; Yang, H.; Guo, S.; Dong, C. The blue-light receptor CmWC-1 mediates fruit body development and secondary metabolism in Cordyceps militaris. Appl. Microbiol. Biotechnol. 2016, 100, 743–755. [Google Scholar] [CrossRef] [PubMed]
- Esquivel-Naranjo, E.U.; Herrera-Estrella, A. Enhanced responsiveness and sensitivity to blue light by blr-2 overexpression in Trichoderma atroviride. Microbiology 2007, 153, 3909–3922. [Google Scholar] [CrossRef] [PubMed]
- Pei, G.L.; Jun, G.U.O.; Wand, Q.H.; Kang, Z.S. Comparative analysis of protein kinases and associated domains between ascomycota and basidiomycota. J. Integr. Agric. 2019, 18, 96–107. [Google Scholar] [CrossRef]
- Nishida, E.; Gotoh, Y. The MAP kinase cascade is essential for diverse signal transduction pathways. Trends Biochem. Sci. 1993, 18, 128–131. [Google Scholar] [CrossRef] [PubMed]
- Giles, J.J.; Bannigan, J.G. Teratogenic and developmental effects of lithium. Curr. Pharm. Des. 2006, 12, 1531–1541. [Google Scholar] [CrossRef]
- Wang, X.; Liu, X.T.; Dunn, R.; Ohl, D.A.; Smith, G.D. Glycogen synthase kinase-3 regulates mouse oocyte homologue segregation. Mol. Reprod. 2003, 64, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Ferruz, N.; Tresadern, G.; Pineda-Lucena, A.; De Fabritiis, G. Multibody cofactor and substrate molecular recognition in the myo-inositol monophosphatase enzyme. Sci. Rep. 2016, 6, 30275. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Huang, W.; Li, X.; Huang, Z.; Liu, X.; Chen, Y.; Shi, T.; Zhang, J. Insights into the role of magnesium triad in myo-inositol monophosphatase: Metal mechanism, substrate binding, and lithium therapy. J. Chem. Inf. Model. 2012, 52, 2398–2409. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Wang, G.; Jiang, C.; Xu, J.R.; Wang, C. Fgk3 glycogen synthase kinase is important for development, pathogenesis, and stress responses in Fusarium graminearum. Sci. Rep. 2015, 5, 8504. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Dagdas, Y.F.; Zhu, X.; Zheng, S.; Chen, L.; Cartwright, Z.; Talbot, N.J.; Wang, Z. The glycogen synthase kinase MoGsk1, regulated by Mps1 MAP kinase, is required for fungal development and pathogenicity in Magnaporthe oryzae. Sci. Rep. 2017, 7, 945. [Google Scholar] [CrossRef] [PubMed]
- Murry, R.; Kniemeyer, O.; Krause, K.; Saiardi, A.; Kothe, E. Crosstalk between Ras and inositol phosphate signaling revealed by lithium action on inositol monophosphatase in Schizophyllum commune. Adv. Biol. Regul. 2019, 72, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.; Cheng, L.; Mudge, A.; Harwood, A.J. A common mechanism of action for three mood-stabilizing drugs. Nature 2022, 417, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Turrà, D.; Segorbe, D.; Di Pietro, A. Protein Kinases in Plant-Pathogenic Fungi: Conserved Regulators of Infection. Annu. Rev. Phytopathol. 2014, 52, 267–288. [Google Scholar] [CrossRef]
- Xiao, Y.; Mitchell, A.P. Shared Roles of Yeast Glycogen Synthase Kinase 3 Family Members in Nitrogen-Responsive Phosphorylation of Meiotic Regulator Ume6p. Mol. Cell. Biol. 2000, 20, 5447–5453. [Google Scholar] [CrossRef]
- Andoh, T.; Hirata, Y.; Kikuchi, A. Yeast Glycogen Synthase Kinase 3 Is Involved in Protein Degradation in Cooperation with Bul1, Bul2, and Rsp5. Mol. Cell. Biol. 2000, 2, 6712–6720. [Google Scholar] [CrossRef] [PubMed]
- Tataroǧlu, Ö.; Lauinger, L.; Sancar, G.; Jakob, K.; Brunner, M.; Diernfellner, A.C.R. Glycogen synthase kinase is a regulator of the circadian clock of Neurospora crassa. J. Biol. Chem. 2012, 287, 36936–36943. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chan, P.-L.; Kwan, H.-S.; Xie, Y.; Wong, K.-H.; Chang, J. Transcriptome Analysis Reveals Mycelial and Fruiting Responses to Lithium Chloride in Coprinopsis cinerea. J. Fungi 2024, 10, 140. https://doi.org/10.3390/jof10020140
Chan P-L, Kwan H-S, Xie Y, Wong K-H, Chang J. Transcriptome Analysis Reveals Mycelial and Fruiting Responses to Lithium Chloride in Coprinopsis cinerea. Journal of Fungi. 2024; 10(2):140. https://doi.org/10.3390/jof10020140
Chicago/Turabian StyleChan, Po-Lam, Hoi-Shan Kwan, Yichun Xie, Ka-Hing Wong, and Jinhui Chang. 2024. "Transcriptome Analysis Reveals Mycelial and Fruiting Responses to Lithium Chloride in Coprinopsis cinerea" Journal of Fungi 10, no. 2: 140. https://doi.org/10.3390/jof10020140