
Anti-Candida Potential of Sclareol in Inhibiting Growth, Biofilm Formation, and Yeast–Hyphal Transition

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungi Strains and Growth Conditions
2.2. Anti-Fungal Susceptibility Microdilution Assay
2.3. Fungi Cell Growth Test
2.4. Membrane Permeabilization Assay
2.5. Measurement of Mitochondrial Membrane Potential
2.6. ROS Detection
2.7. Biofilm Formation Assay
2.8. Hyphal Transition Assay
2.9. Synergistic Anti-Fungal Activity Assay
2.10. Quantitative Reverse Transcriptase PCR (qRT-PCR) Analysis
2.11. Statistical Analysis
3. Results
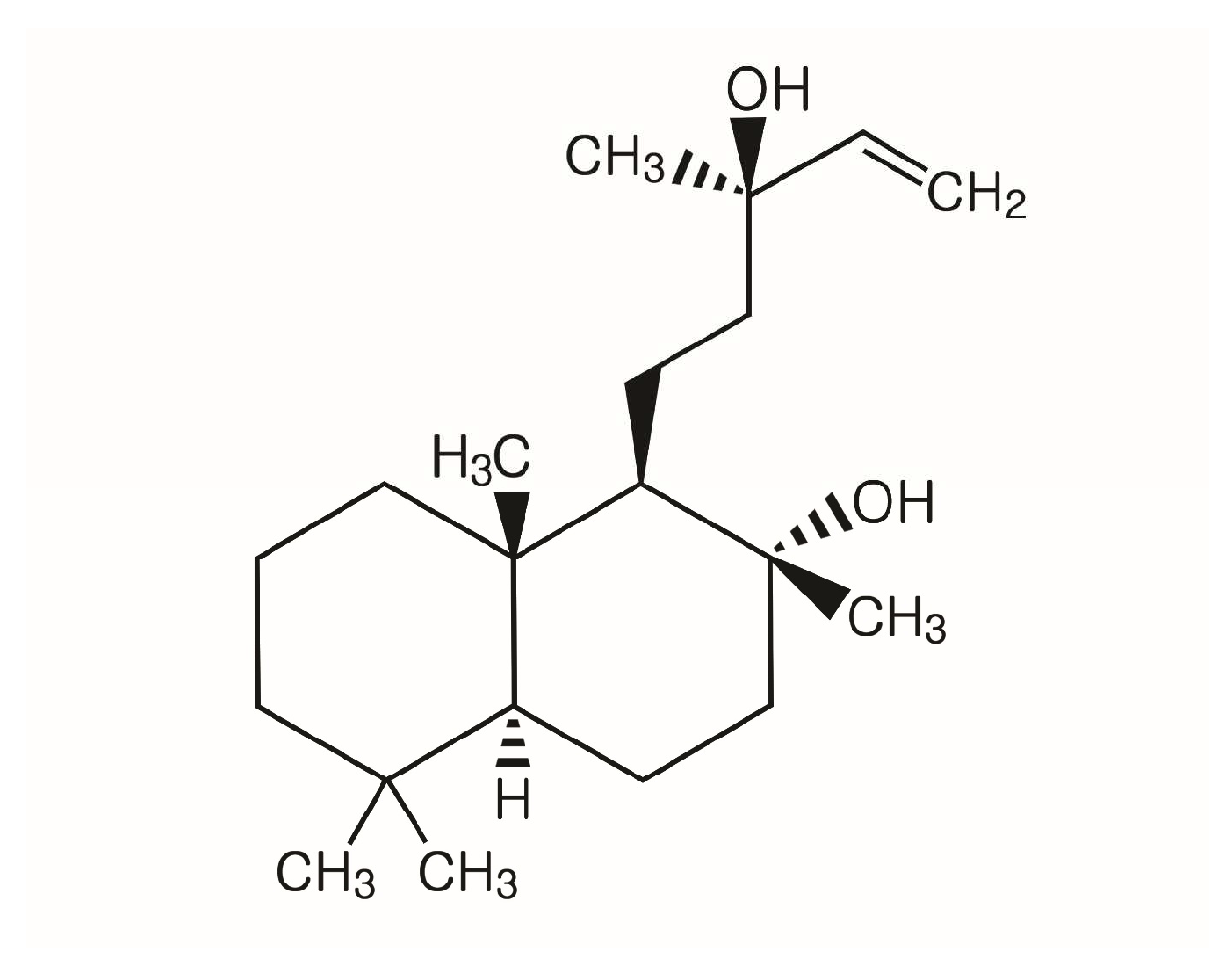
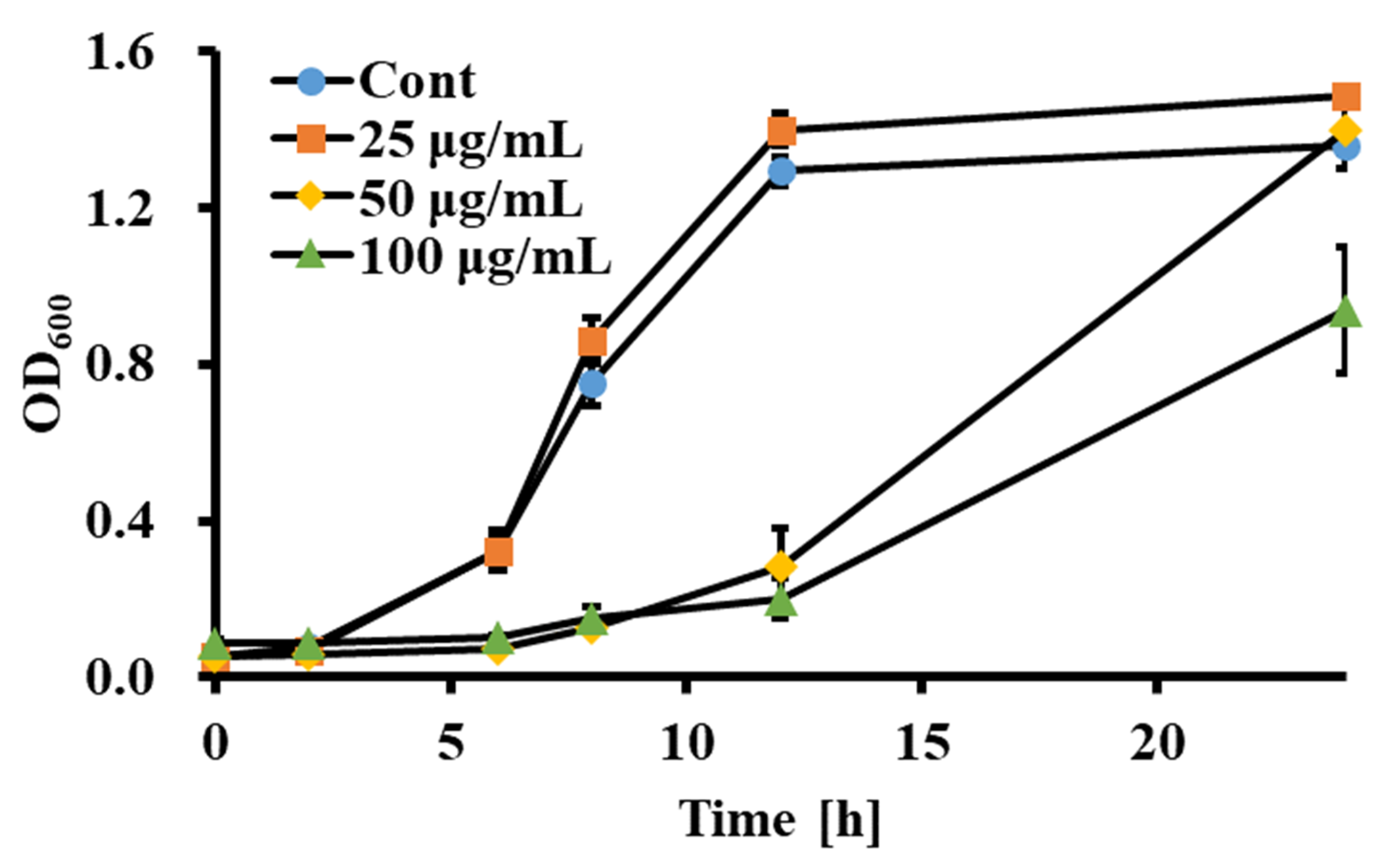
3.1. Sclareol Inhibited the Growth of Candida spp.
3.2. Sclareol Reduced Membrane Permeability of C. albicans
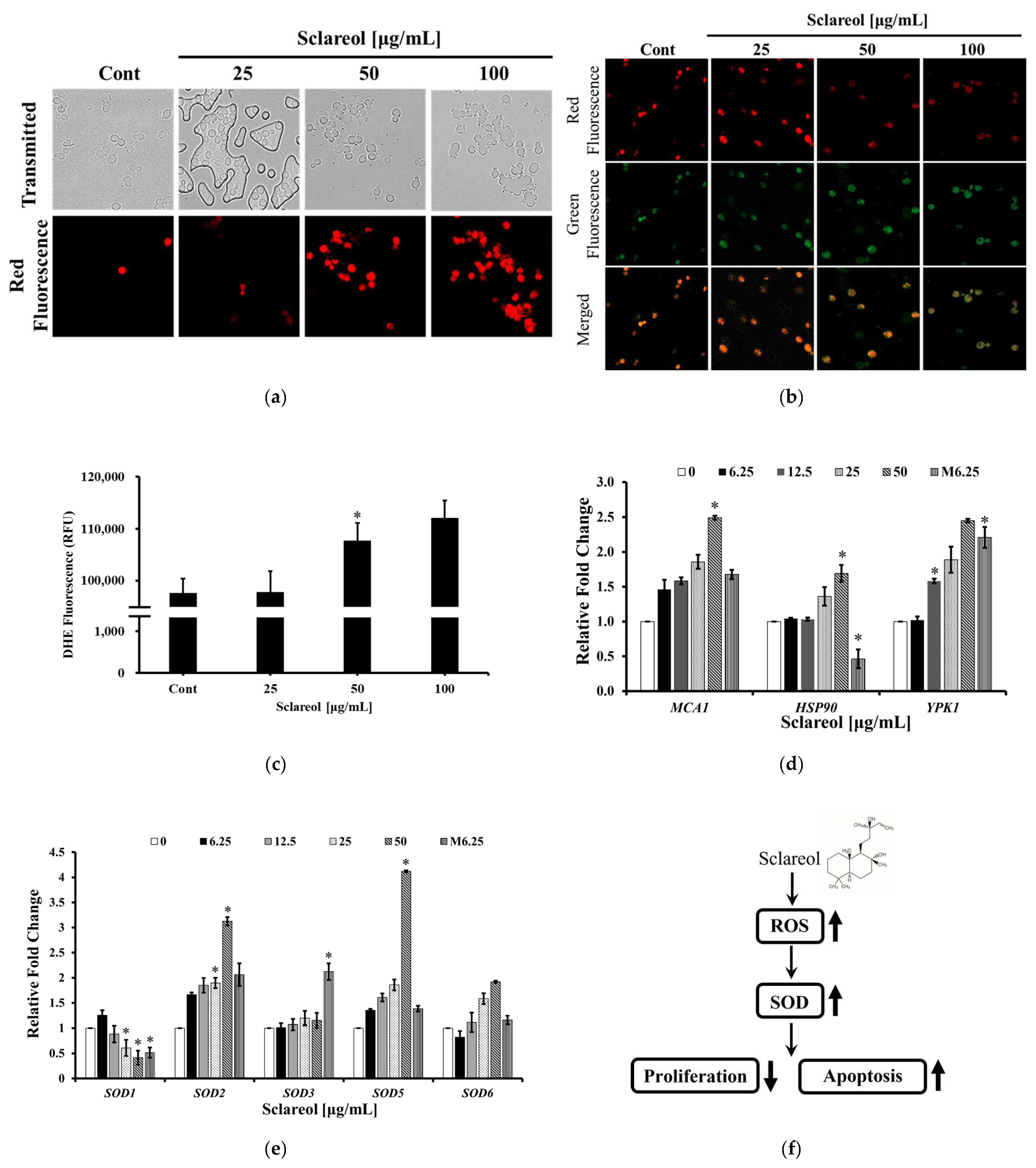
3.3. Sclareol Induced Apoptosis-like Cell Death through Disruption of Mitochondrial Membrane Potential and Increasing Intracellular Concentration of ROS
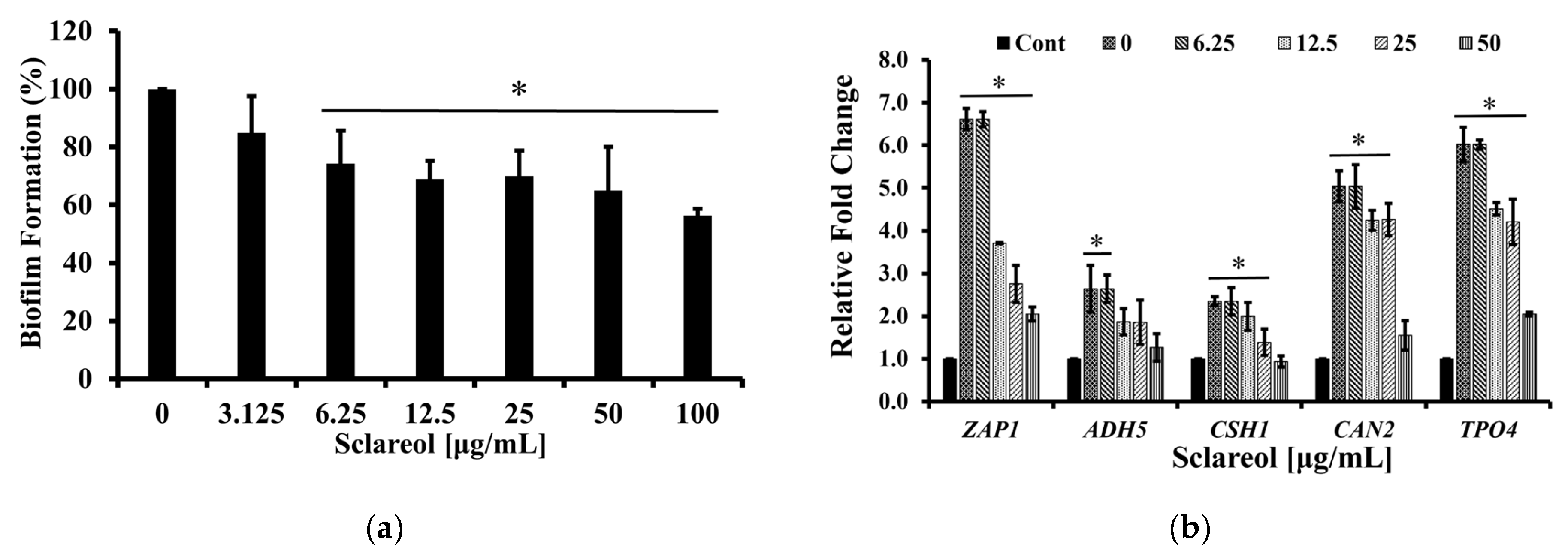
3.4. Sclareol Inhibited the Biofilm Formation of C. albicans
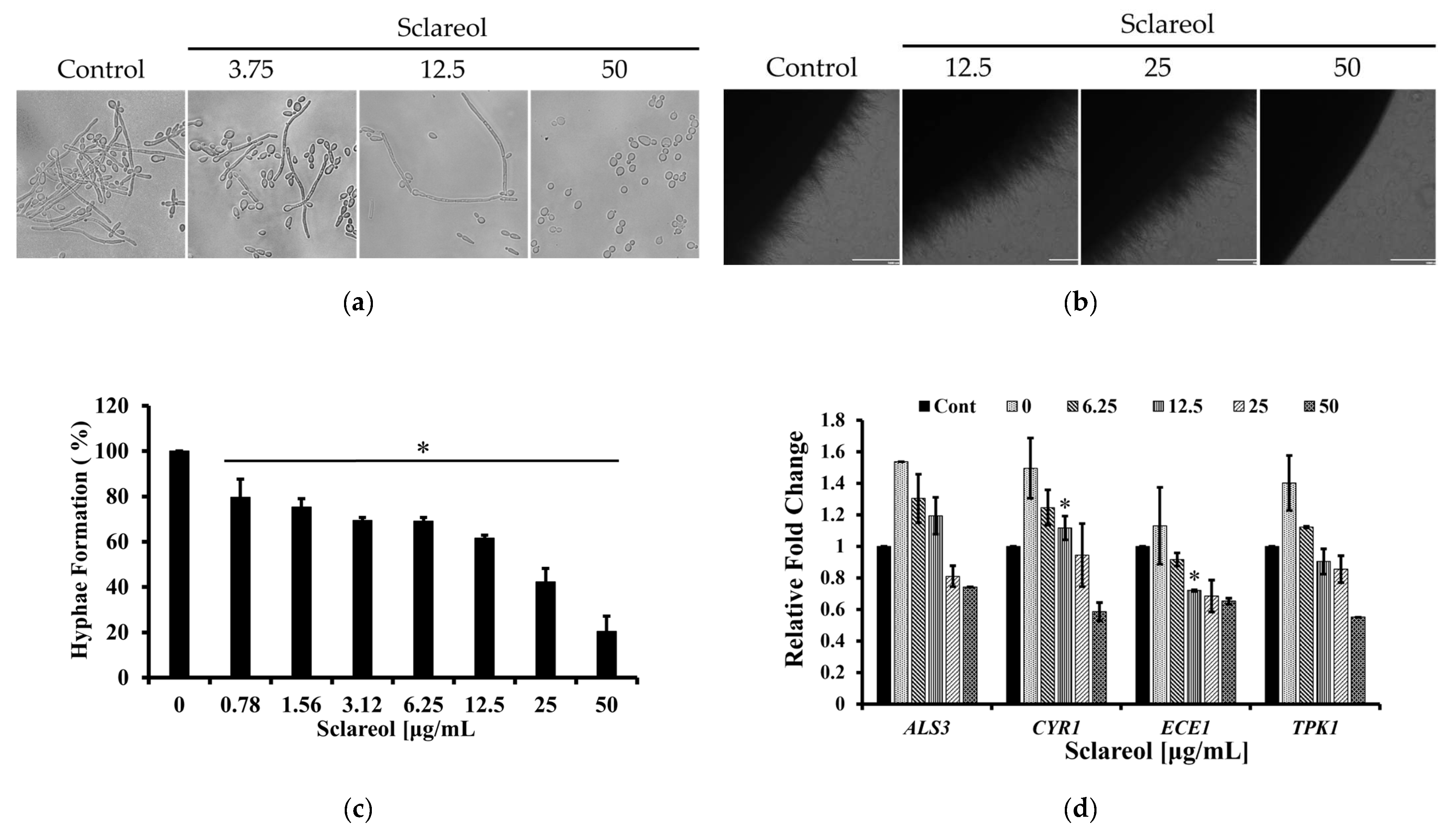
3.5. Sclareol Inhibited the Hyphal Formation of C. albicans
3.6. Sclareol Synergistically Inhibited the Growth of C. albicans with Miconazole
4. Discussion
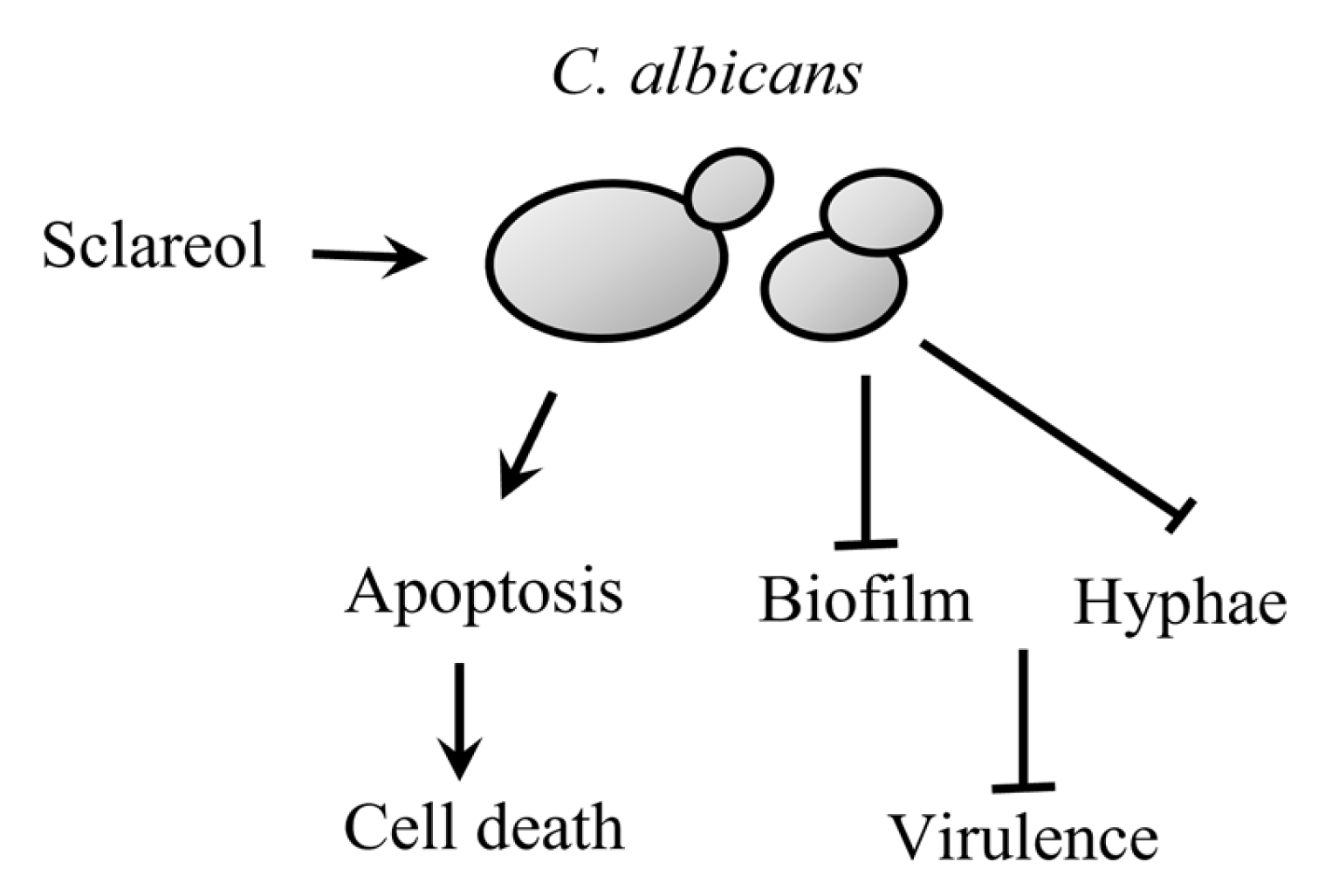
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Phillips, A.J.; Sudbery, I.; Ramsdale, M. Apoptosis induced by environmental stresses and amphotericin B in Candida albicans. Proc. Natl. Acad. Sci. USA 2003, 100, 14327–14332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajjeh, R.A.; Sofair, A.N.; Harrison, L.H.; Lyon, G.M.; Arthington-Skaggs, B.A.; Mirza, S.A.; Phelan, M.; Morgan, J.; Lee-Yang, W.; Ciblak, M.A.; et al. Incidence of bloodstream infections due to Candida species and in vitro susceptibilities of isolates collected from 1998 to 2000 in a population-based active surveillance program. J. Clin. Microbiol. 2004, 42, 1519–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Chakrabarti, A. Invasive candidiasis in the Southeast-Asian region. In Candida albicans: Cellular and Molecular Biology; Springer: Cham, Switzerland, 2017; pp. 25–40. [Google Scholar] [CrossRef]
- Sardi, J.C.O.; Scorzoni, L.; Bernardi, T.; Fusco-Almeida, A.M.; Mendes Giannini, M.J.S. Candida species: Current epidemiology, pathogenicity, biofilm formation, natural antifungal products and new therapeutic options. J. Med. Microbiol. 2013, 62, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Thatchanamoorthy, N.; Rukumani Devi, V.; Chandramathi, S.; Tee Tay, S. Candida auris: A mini review on epidemiology in healthcare facilities in Asia. J. Fungi 2022, 8, 1126. [Google Scholar] [CrossRef] [PubMed]
- Gintjee, T.J.; Donnelley, M.A.; Thompson, G.R., III. Aspiring antifungals: Review of current antifungal pipeline developments. J. Fungi 2020, 6, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharon, A.; Finkelstein, A.; Shlezinger, N.; Hatam, I. Fungal apoptosis: Function, genes and gene function. FEMS Microbiol. Rev. 2009, 33, 833–854. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.; Lee, D.G. Cecropin A-induced apoptosis is regulated by ion balance and glutathione antioxidant system in Candida albicans. IUBMB Life 2016, 68, 652–662. [Google Scholar] [CrossRef] [Green Version]
- Madeo, F.; Herker, E.; Wissing, S.; Jungwirth, H.; Eisenberg, T.; Fröhlich, K.U. Apoptosis in yeast. Curr. Opin. Microbiol. 2004, 7, 655–660. [Google Scholar] [CrossRef]
- Wu, X.Z.; Chang, W.Q.; Cheng, A.X.; Sun, L.M.; Lou, H.X. Plagiochin E, an antifungal active macrocyclic bis(bibenzyl), induced apoptosis in Candida albicans through a metacaspase-dependent apoptotic pathway. Biochim. Biophys. Acta 2010, 1800, 439–447. [Google Scholar] [CrossRef]
- Douglas, L.J. Medical importance of biofilms in Candida infections. Rev. Iberoam. Micol. 2002, 19, 139–143. [Google Scholar]
- Donlan, R.M. Biofilm formation: A clinically relevant microbiological process. Clin. Infect. Dis. 2001, 33, 1387–1392. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Zhou, X.; Ren, B.; Cheng, L. The regulation of hyphae growth in Candida albicans. Virulence 2020, 11, 337–348. [Google Scholar] [CrossRef] [Green Version]
- Dimas, K.; Papadaki, M.; Tsimplouli, C.; Hatziantoniou, S.; Alevizopoulos, K.; Pantazis, P.; Demetzos, C. Labd-14-ene-8,13-diol (sclareol) induces cell cycle arrest and apoptosis in human breast cancer cells and enhances the activity of anticancer drugs. Biomed. Pharmacother. 2006, 60, 127–133. [Google Scholar] [CrossRef]
- Wang, L.; He, H.S.; Yu, H.L.; Zeng, Y.; Han, H.; He, N.; Liu, Z.G.; Wang, Z.Y.; Xu, S.J.; Xiong, M. Sclareol, a plant diterpene, exhibits potent antiproliferative effects via the induction of apoptosis and mitochondrial membrane potential loss in osteosarcoma cancer cells. Mol. Med. Rep. 2015, 11, 4273–4278. [Google Scholar] [CrossRef] [Green Version]
- Duan, G.; Hou, S.; Ji, J.; Deng, B. The study of sclareol in inhibiting proliferation of osteosarcoma cells by apoptotic induction and loss of mitochondrial membrane potential. Cancer Biomark. 2018, 22, 29–34. [Google Scholar] [CrossRef]
- Bhatia, S.P.; McGinty, D.; Letizia, C.S.; Api, A.M. Fragrance material review on sclareol. Food Chem. Toxicol. 2008, 46, S270–S274. [Google Scholar] [CrossRef]
- Tsai, S.W.; Hsieh, M.C.; Li, S.; Lin, S.C.; Wang, S.P.; Lehman, C.W.; Lien, C.Z.; Lin, C.C. Therapeutic potential of sclareol in experimental models of rheumatoid arthritis. Int. J. Mol. Sci. 2018, 19, 1351. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Kim, K.Y. Antibacterial effect of sophoraflavanone G by destroying the cell wall of Enterococcus faecium. J. Appl. Pharm. Sci. 2020, 10, 59–64. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Diekema, D. Progress in antifungal susceptibility testing of Candida spp. by use of Clinical and Laboratory Standards Institute broth microdilution methods, 2010 to 2012. J. Clin. Microbiol. 2012, 50, 2846–2856. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Ha Quang Bao, T.; Shin, Y.K.; Kim, K.Y. Antifungal activity of magnoflorine against Candida strains. World J. Microbiol. Biotechnol. 2018, 34, 167. [Google Scholar] [CrossRef]
- Lee, J.; Kim, J.G.; Lee, H.; Lee, T.H.; Kim, K.Y.; Kim, H. Antifungal activity of 1,4-Dialkoxynaphthalen-2-Acyl Imidazolium salts by inducing apoptosis of pathogenic Candida spp. Pharmaceutics 2021, 13, 312. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Kim, J.G.; Kim, K.Y. Trichosanthes kirilowii extract promotes wound healing through the phosphorylation of ERK1/2 in Keratinocytes. Biomimetics 2022, 7, 154. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.H.; Choi, H.; Kim, A.R.; Yun, J.W.; Yu, R.; Woo, E.R.; Lee, D.G. Hibicuslide C-induced cell death in Candida albicans involves apoptosis mechanism. J. Appl. Microbiol. 2014, 117, 1400–1411. [Google Scholar] [CrossRef] [PubMed]
- Sivandzade, F.; Bhalerao, A.; Cucullo, L. Analysis of the mitochondrial membrane potential using the cationic JC-1 dye as a sensitive fluorescent probe. Bio-Protocol 2019, 9, e3128. [Google Scholar] [CrossRef] [PubMed]
- Katherine, M.C.; Cristina, M.A.; Estibaliz, M.; José Manuel, A.U.; Guillermo, Q.; Elena, E. In vitro activities of carvacrol, cinnamaldehyde and thymol against Candida biofilms. Biomed. Pharmacother. 2021, 143, 112218. [Google Scholar] [CrossRef]
- Kim, D.; Kim, K.Y. Adenophora triphylla var. japonica inhibits Candida biofilm formation, increases susceptibility to antifungal agents and reduces infection. Int. J. Mol. Sci. 2021, 22, 12523. [Google Scholar] [CrossRef]
- Hawser, S.P.; Douglas, L.J. Biofilm formation by Candida species on the surface of catheter materials in vitro. Infect. Immun. 1994, 62, 915–921. [Google Scholar] [CrossRef] [Green Version]
- Bravo-Chaucanés, C.P.; Vargas-Casanova, Y.; Chitiva-Chitiva, L.C.; Ceballos-Garzon, A.; Modesti-Costa, G.; Parra-Giraldo, C.M. Evaluation of anti-Candida potential of Piper nigrum extract in inhibiting growth, yeast-hyphal transition, virulent enzymes, and biofilm formation. J. Fungi 2022, 8, 784. [Google Scholar] [CrossRef]
- Guilherme, M.C.; Walicyranison, P.S. Superoxide dismutases and glutaredoxins have a distinct role in the response of Candida albicans to oxidative stress generated by the chemical compounds menadione and diamide. Mem. Inst. Oswaldo Cruz 2012, 107, 998–1005. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Kohler, J.; Fink, G.R. Suppression of hyphal formation in Candida albicans by mutation of a STE12 homolog. Science 1994, 266, 1723–1726. [Google Scholar] [CrossRef]
- Haghdoost, N.S.; Salehi, T.Z.; Khosravi, A.; Sharifzadeh, A. Antifungal activity and influence of propolis against germ tube formation as a critical virulence attribute by clinical isolates of Candida albicans. J. Mycol. Med. 2016, 26, 298–305. [Google Scholar] [CrossRef]
- Dongliang, Y.; Yanling, H.; Zixin, Y.; Qianru, G.; Yuqian, Z.; Fong, Y.C.; Guisheng, Z.; Lixing, W.; Lianhui, W.; Yue, W. Candida albicans ubiquitin and heat shock factor-type transcriptional factors are involved in 2-Dodecenoic acid-mediated inhibition of hyphal growth. Microorganisms 2020, 8, 75. [Google Scholar] [CrossRef] [Green Version]
- Yamada-Okabe, T.; Mio, T.; Ono, N.; Kashima, Y.; Matsui, M.; Arisawa, M.; Yamada-Okabe, H. Roles of three histidine kinase genes in hyphal development and virulence of the pathogenic fungus Candida albicans. J. Bacteriol. 1999, 181, 7243–7247. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Kim, K.Y. Pectolinarin inhibits the bacterial biofilm formation and thereby reduces bacterial pathogenicity. Antibiotics 2022, 11, 598. [Google Scholar] [CrossRef]
- Konaté, K.; Mavoungou, J.F.; Lepengué, A.N.; Aworet-Samseny, R.R.; Hilou, A.; Souza, A.; Dicko, M.H.; M’Batchi, B. Antibacterial activity against β-lactamase producing methicillin and ampicillin-resistants Staphylococcus aureus: Fractional inhibitory concentration index (FICI) determination. Ann. Clin. Microbiol. Antimicrob. 2012, 11, 18. [Google Scholar] [CrossRef] [Green Version]
- Lane, S.; Zhou, S.; Pan, T.; Dai, Q.; Liu, H. The basic helix-loop-helix transcription factor Cph2 regulates hyphal development in Candida albicans partly via TEC1. Mol. Cell. Biol. 2001, 21, 6418–6428. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, A.T.; Kim, K.Y. Inhibition of proinflammatory cytokines in Cutibacterium acnes-induced inflammation in HaCaT cells by using Buddleja davidii aqueous extract. Int. J. Inflam. 2020, 2020, 8063289. [Google Scholar] [CrossRef] [Green Version]
- Carmona-Gutierrez, D.; Eisenberg, T.; Büttner, S.; Meisinger, C.; Kroemer, G.; Madeo, F. Apoptosis in yeast: Triggers, pathways, subroutines. Cell Death Differ. 2010, 17, n763–n773. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Luo, L.; Guo, J.; Liu, H.; Wang, B.; Deng, B.; Long, C.A.; Cheng, Y. Farnesol induces apoptosis and oxidative stress in the fungal pathogen Penicillium expansum. Mycologia 2010, 102, 311–318. [Google Scholar] [CrossRef]
- Da, X.; Nishiyama, Y.; Tie, D.; Hein, K.Z.; Yamamoto, O.; Morita, E. Antifungal activity and mechanism of action of Ougon (Scutellaria root extract) components against pathogenic fungi. Sci. Rep. 2019, 9, 1683. [Google Scholar] [CrossRef]
- Liu, X.; Ma, Z.; Zhang, J.; Yang, L. Antifungal compounds against Candida infections from traditional chinese medicine. BioMed Res. Int. 2017, 2017, 4614183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rockenfeller, P.; Madeo, F. Apoptotic death of ageing yeast. Exp. Gerontol. 2008, 43, 876–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.; Reiners, J.J.; Santiago, A.M.; Kessel, D. Monitoring singlet oxygen and hydroxyl radical formation with fluorescent probes during photodynamic therapy. Photochem. Photobiol. 2009, 85, 1177–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benaroudj, N.; Lee, D.H.; Goldberg, A.L. Trehalose accumulation during cellular stress protects cells and cellular proteins from damage by oxygen radicals. J. Biol. Chem. 2001, 276, 24261–24267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza, L.; Sepulveda, C.; Melo, R.; Cotoras, M. Characterization of the antifungal activity against Botrytis cinerea of sclareol and 13-epi-sclareol, two labdane-type diterpenoids. J. Chil. Chem. 2015, 60, 3024–3028. [Google Scholar] [CrossRef] [Green Version]
- Jia, C.; Zhang, J.; Yu, L.; Wang, C.; Yang, Y.; Rong, X.; Xu, K.; Chu, M. Antifungal activity of Coumarin against Candida albicans is related to apoptosis. Front. Cell. Infect. Microbiol. 2019, 8, 445. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.; Silva, R.D.; Saraiva, L.; Johansson, B.; Sousa, M.J.; Côrte-Real, M. Mitochondria-dependent apoptosis in yeast. Biochim. Biophys. Acta 2008, 1783, 1286–1302. [Google Scholar] [CrossRef] [Green Version]
- Dai, B.; Wang, Y.; Li, D.; Xu, Y.; Liang, R.; Zhao, L.; Cao, Y.; Jia, J.; Jiang, Y. Hsp90 is involved in apoptosis of Candida albicans by regulating the calcineurin-caspase apoptotic pathway. PLoS ONE 2012, 7, e45109. [Google Scholar] [CrossRef] [Green Version]
- Swamy, K.S.; Sirsi, M.; Ramananda, G.R. Studies on the mechanism of action of miconazole: Effect of miconazole on respiration and cell permeability of Candida albicans. Antimicrob. Agents Chemother. 1974, 5, 420–425. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, D.; Kondo, K.; Uehara, N.; Otokozawa, S.; Tsuji, N.; Yagihashi, A.; Watanabe, N. Endogenous reactive oxygen species is an important mediator of miconazole antifungal effect. Antimicrob. Agents Chemother. 2002, 46, 3113–3117. [Google Scholar] [CrossRef]
- Cheng, J.; Park, T.S.; Chio, L.C.; Fischl, A.S.; Ye, X.S. Induction of apoptosis by sphingoid long-chain bases in Aspergillus nidulans. Mol. Cell. Biol. 2003, 23, 163–177. [Google Scholar] [CrossRef] [Green Version]
- Niles, B.J.; Mogri, H.; Hill, A.; Vlahakis, A.; Powers, T. Plasma membrane recruitment and activation of the AGC kinase Ypk1 is mediated by target of rapamycin complex 2 (TORC2) and its effector proteins Slm1 and Slm2. Proc. Natl. Acad. Sci. USA 2012, 109, 1536–1541. [Google Scholar] [CrossRef] [Green Version]
- Salazar, V.A.; Arranz-Trullén, J.; Prats-Ejarque, G.; Torrent, M.; Andreu, D.; Pulido, D.; Boix, E. Insight into the antifungal mechanism of action of human RNase N-terminus derived peptides. Int. J. Mol. Sci. 2019, 20, 4558. [Google Scholar] [CrossRef] [Green Version]
- Barber, S.C.; Mead, R.J.; Shaw, P.J. Oxidative stress in ALS: A mechanism of neurodegeneration and a therapeutic target. Biochim. Biophys. Acta 2006, 1762, 1051–1067. [Google Scholar] [CrossRef] [Green Version]
- Morici, P.; Fais, R.; Rizzato, C.; Tavanti, A.; Lupetti, A. Inhibition of Candida albicans biofilm formation by the synthetic Lactoferricin derived peptide hLF1-11. PLoS ONE 2016, 11, e0167470. [Google Scholar] [CrossRef] [Green Version]
- Nobile, C.J.; Nett, J.E.; Hernday, A.D.; Homann, O.R.; Deneault, J.S.; Nantel, A.; Andres, D.R.; Johnson, A.D.; Mitchell, A.P. Biofilm matrix regulation by Candida albicans Zap1. PLoS Biol. 2009, 7, e1000133. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, P.K.; Mohamed, S.; Chandra, J.; Kuhn, D.; Liu, S.; Antar, O.S.; Munyon, R.; Mitchell, A.P.; Andes, D.; Chance, M.R.; et al. Alcohol dehydrogenase restricts the ability of the pathogen Candida albicans to form a biofilm on catheter surfaces through an ethanol-based mechanism. Infect. Immun. 2006, 74, 3804–3816. [Google Scholar] [CrossRef] [Green Version]
- Clarissa, J.N.; Emily, P.F.; Jeniel, E.N.; Trevor, R.S.; Quinn, M.M.; Aaron, D.H.; Brian, B.T.; David, R.A.; Alexander, D.J. A recently evolved transcriptional network controls biofilm development in Candida albicans. Cell 2012, 148, 126–138. [Google Scholar] [CrossRef] [Green Version]
- Igarashi, K.; Kashiwagi, K. Characteristics of cellular polyamine transport in prokaryotes and eukaryotes. Plant Physiol. Biochem. 2010, 48, 506–512. [Google Scholar] [CrossRef]
- Igarashi, K.; Kashiwagi, K. Modulation of cellular function by polyamines. Int. J. Biochem. Cell Biol. 2010, 42, 39–51. [Google Scholar] [CrossRef]
- Fan, Y.; He, H.; Dong, Y.; Pan, H. Hyphae-specific genes HGC1, ALS3, HWP1, and ECE1 and relevant signaling pathways in Candida albicans. Mycopathologia 2013, 176, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, Y.G.; Choi, P.J.; Ham, J.Y.; Park, J.G.; Lee, J.T. Antibiofilm and antivirulence activities of 6-Gingerol and 6-Shogaol against Candida albicans due to hyphal inhibition. Front. Cell. Infect. Microbiol. 2018, 8, 299. [Google Scholar] [CrossRef] [PubMed]
- Robertson, L.S.; Causton, H.C.; Young, R.A.; Fink, G.R. The yeast A-kinases differentially regulate iron uptake and respiratory function. Proc. Natl. Acad. Sci. USA 2000, 97, 5984–5988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, T.; Asahara, M.; Yamamoto, M.; Yamaura, M.; Matsumura, M.; Goto, K.; Rezaei-Matehkolaei, A.; Mirhendi, H.; Makimura, M.; Makimura, K. In vitro susceptibility of dermatomycoses agents to six antifungal drugs and evaluation by fractional inhibitory concentration index of combined effects of amorolfine and itraconazole in dermatophytes. Microbiol. Immunol. 2014, 58, 1–8. [Google Scholar] [CrossRef]
- Odds, F.C. Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 2003, 52, 1. [Google Scholar] [CrossRef]
- Bahn, Y.S.; Sundstrom, P. CAP1, an adenylate cyclase-associated protein gene, regulates bud-hypha transitions, filamentous growth, and cyclic AMP levels and is required for virulence of Candida albicans. J. Bacteriol. 2001, 183, 3211–3223. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Function | References |
|---|---|---|---|
| ACT1 | F: TAGGTTTGGAAGCTGCTGG R: CCTGGGAACATGGTAGTAC | Control | [39] |
| MCA1 | F: TATAATAGACCTTCTGGAC R: TTGGTGGACGAGAATAATG | Apoptosis | In this study |
| HSP90 | F: GGGAATCTAACGCTGGTGGTAA R: TTCGGTTTCTGGAACTTCTTTT | Apoptosis | In this study |
| YPK1 | F: CAACACAACACAGTAGCACC R: GTTGTGGATAAAGGTGGTTCG | Apoptosis | In this study |
| SOD1 | F: TTAAAGCTGTCGCTGTTGTC R: AATATGGAAACCTCTCAAGGC | Antioxidant | In this study |
| SOD2 | F: AACTTGGCTCCTGTCTC R: TATCACCATTGGCTTTG | Antioxidant | In this study |
| SOD3 | F: TATCACCATTGGCTTTG R: TATCACCATTGGCTTTG | Antioxidant | In this study |
| SOD5 | F: ACGAGGGACACGGCAATGCT R: ACGAGGGACACGGCAATGCT | Antioxidant | In this study |
| SOD6 | F: GACCCCGACCCACCTCAACAA R: GGGTAGCAAGGAGTGCCGGT | Antioxidant | In this study |
| ZAP1 | F: ATCTGTCCAGTGTTGTTTGTA R: AGGTCTCTTTGAAAGTTGTG | Biofilm | [39] |
| ADH5 | F: ACCTGCAAGGGCTCATTCTG R: CGGCTCTCAACTTCTCCATA | Biofilm | [39] |
| CSH1 | F: CGTGAGGACGAGAGAGAAT R: CGAATGGACGACACAAAACA | Biofilm | [39] |
| TPO4 | F: GCTGCTACCAATGTCAGTCC R: ACGGAGCTATCCGAATCGTC | Biofilm | In this study |
| CAN2 | F: GCGGAATGGATATGCATGGG R: CGGATTGCTCTTGGAGAAGC | Biofilm | [39] |
| ALS3 | F: GGTTATCGTCCATTTGTTG R: TTCTGTATCCAGTCCATCT | Hyphae | [39] |
| CYR1 | F: GTTTCCCCCACCACTCA R: TTGCGGTAATGACACAACAG | Hyphae | [39] |
| ECE1 | F: ACAGTTTCCAGGACGCCAT R: ATTGTTGCTCGTGTTGCCA | Hyphae | [39] |
| TPK1 | F: CCAACGATTCCCTACTCCAG R: GCAATATAATCTGGAGTCCCAC | Hyphae | In this study |
| Candida spp. | Sclareol [μg/mL] | ||
|---|---|---|---|
| 24 h | 48 h | 72 h | |
| C. albicans (KCTC7965) | 50 | 100 | 100 |
| C. auris (KCTC17809) | 50 | >100 | >100 |
| C. glabrata (KCTC7219) | >100 | >100 | >100 |
| C. parapsilosis var. parapsilosis (KACC45480) | 50 | >100 | >100 |
| C. parapsilosis (KACC49573) | >100 | >100 | >100 |
| C. tropicalis var. tropicalis (KCTC17762) | >100 | >100 | >100 |
| Candida spp. | Sclareol [μg/mL] | |
|---|---|---|
| MIC | MFC | |
| C. albicans (KCTC7965) | 50 | 100 |
| Candida spp. | MIC(μg/mL) of Miconazole | MIC(μg/mL) of Sclareol | FICI | Synergy | ||
|---|---|---|---|---|---|---|
| Alone | With Sclareol | Alone | With Miconazole | |||
| C. albicans (KCTC7965) | 3.125 | 0.78 | 50 | 3.125 | 0.31 | Synergy |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, C.; Kim, J.-G.; Kim, K.-Y. Anti-Candida Potential of Sclareol in Inhibiting Growth, Biofilm Formation, and Yeast–Hyphal Transition. J. Fungi 2023, 9, 98. https://doi.org/10.3390/jof9010098
Kim C, Kim J-G, Kim K-Y. Anti-Candida Potential of Sclareol in Inhibiting Growth, Biofilm Formation, and Yeast–Hyphal Transition. Journal of Fungi. 2023; 9(1):98. https://doi.org/10.3390/jof9010098
Chicago/Turabian StyleKim, Chaerim, Jae-Goo Kim, and Ki-Young Kim. 2023. "Anti-Candida Potential of Sclareol in Inhibiting Growth, Biofilm Formation, and Yeast–Hyphal Transition" Journal of Fungi 9, no. 1: 98. https://doi.org/10.3390/jof9010098