
Metabolomic Profiling of Different Antrodia cinnamomea Phenotypes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Determination of Total Triterpenoid Contents
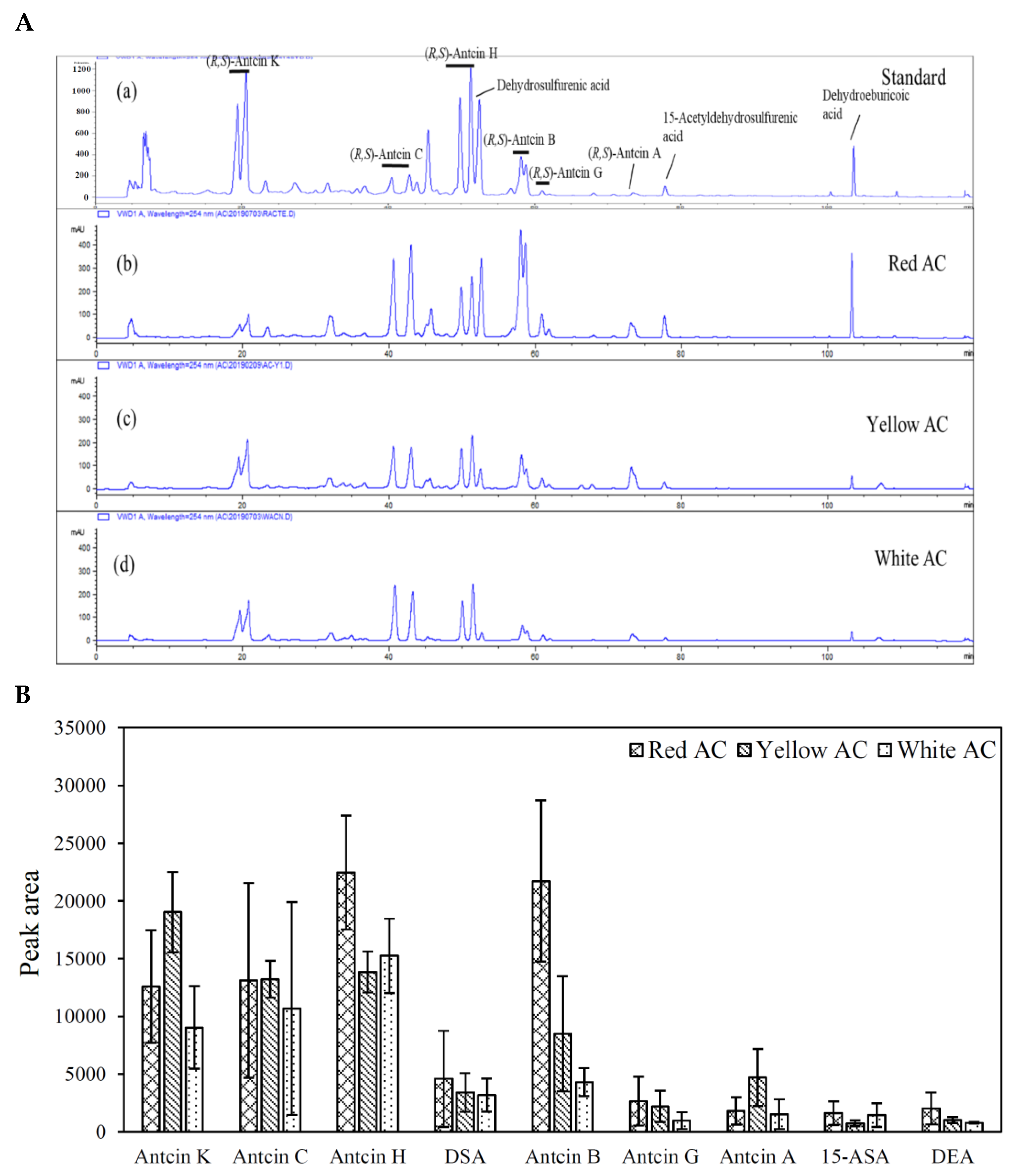
2.3. Analysis of AC Index Compounds
2.4. Metabolomic Analysis
2.5. Statistical Analysis
3. Results and Discussion
3.1. Authentication of A. cinnamomea Phenotypes
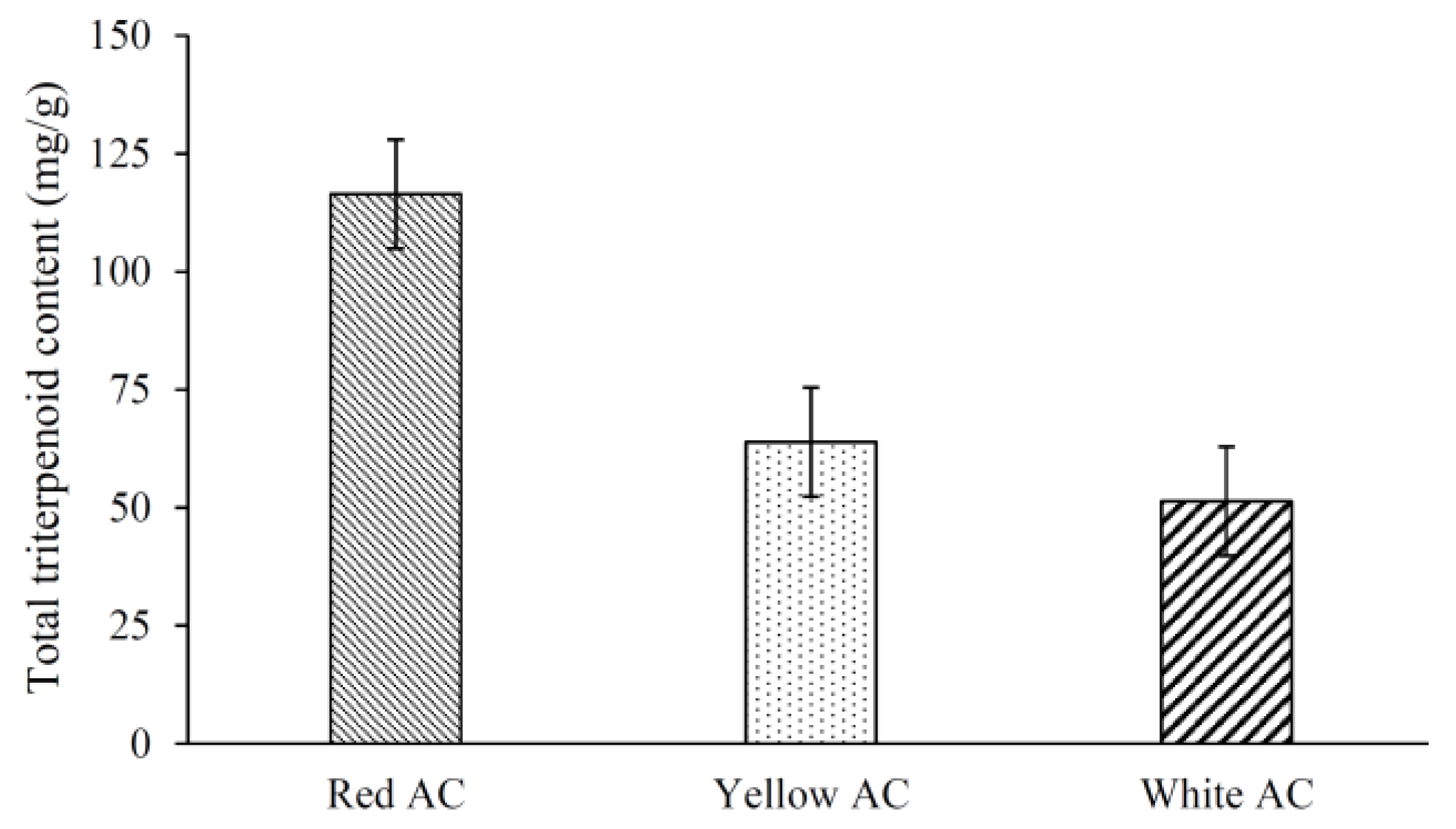
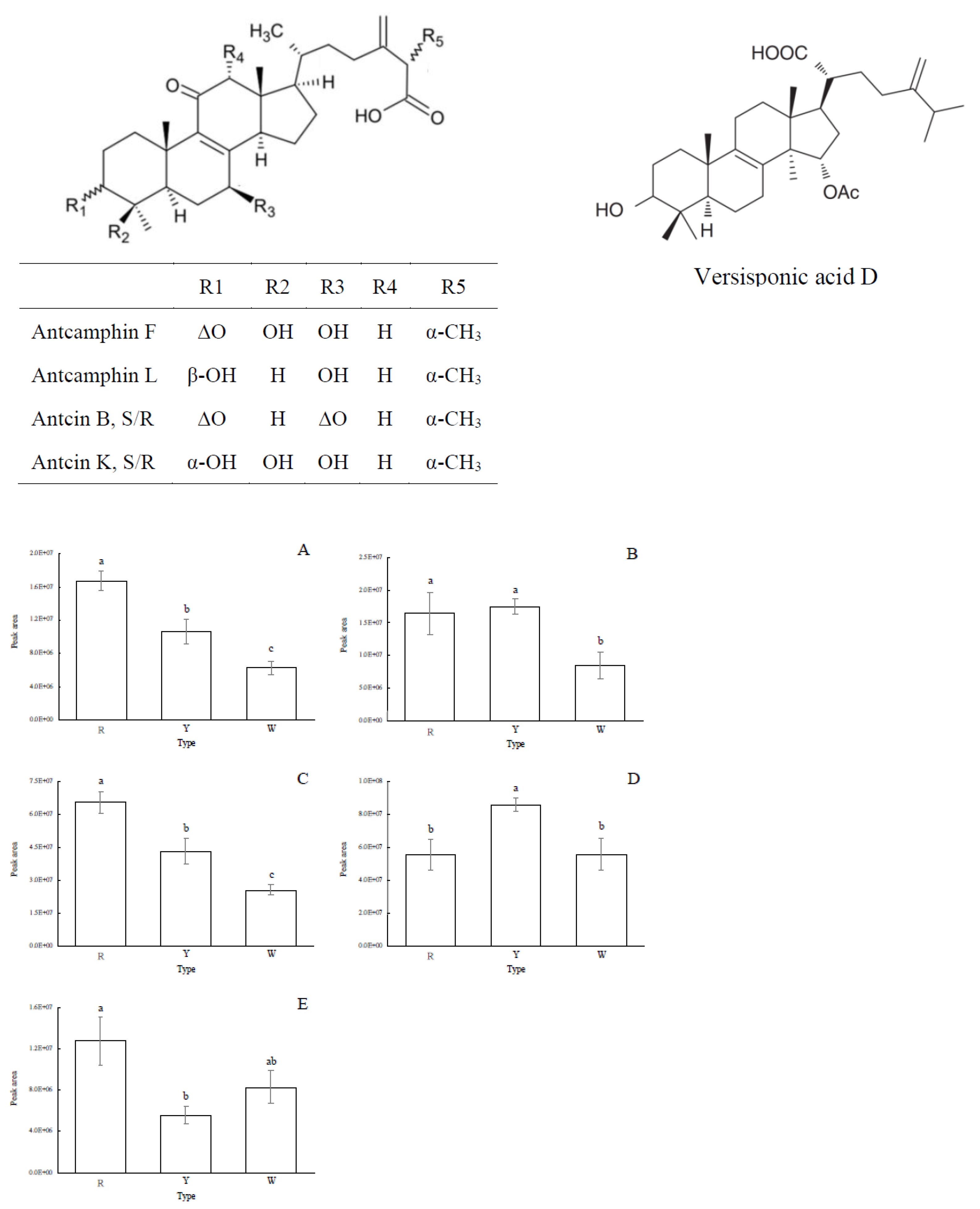
3.2. Triterpenoid Contents and Profiles of Different A. cinnamomea Phenotypes
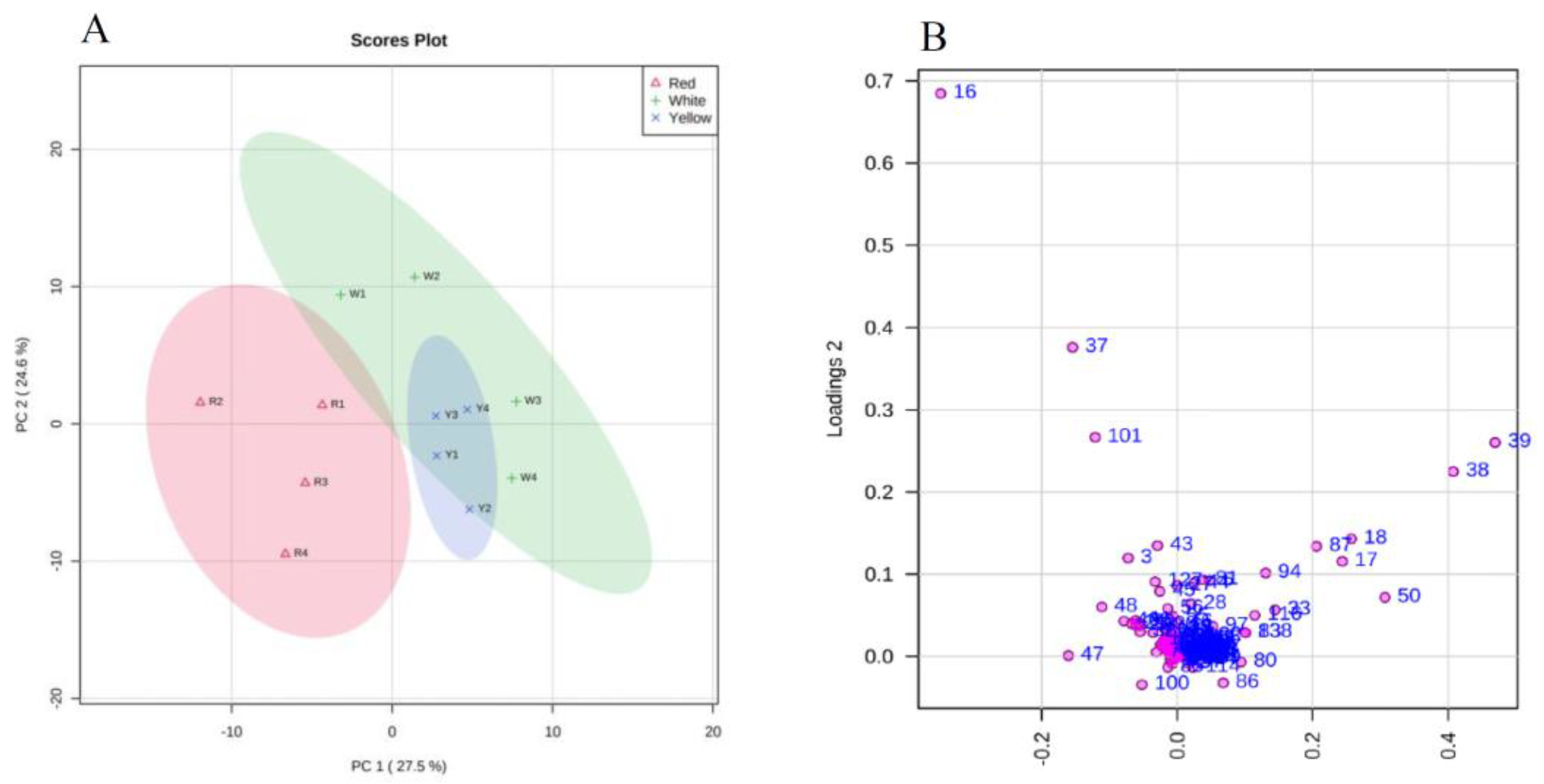
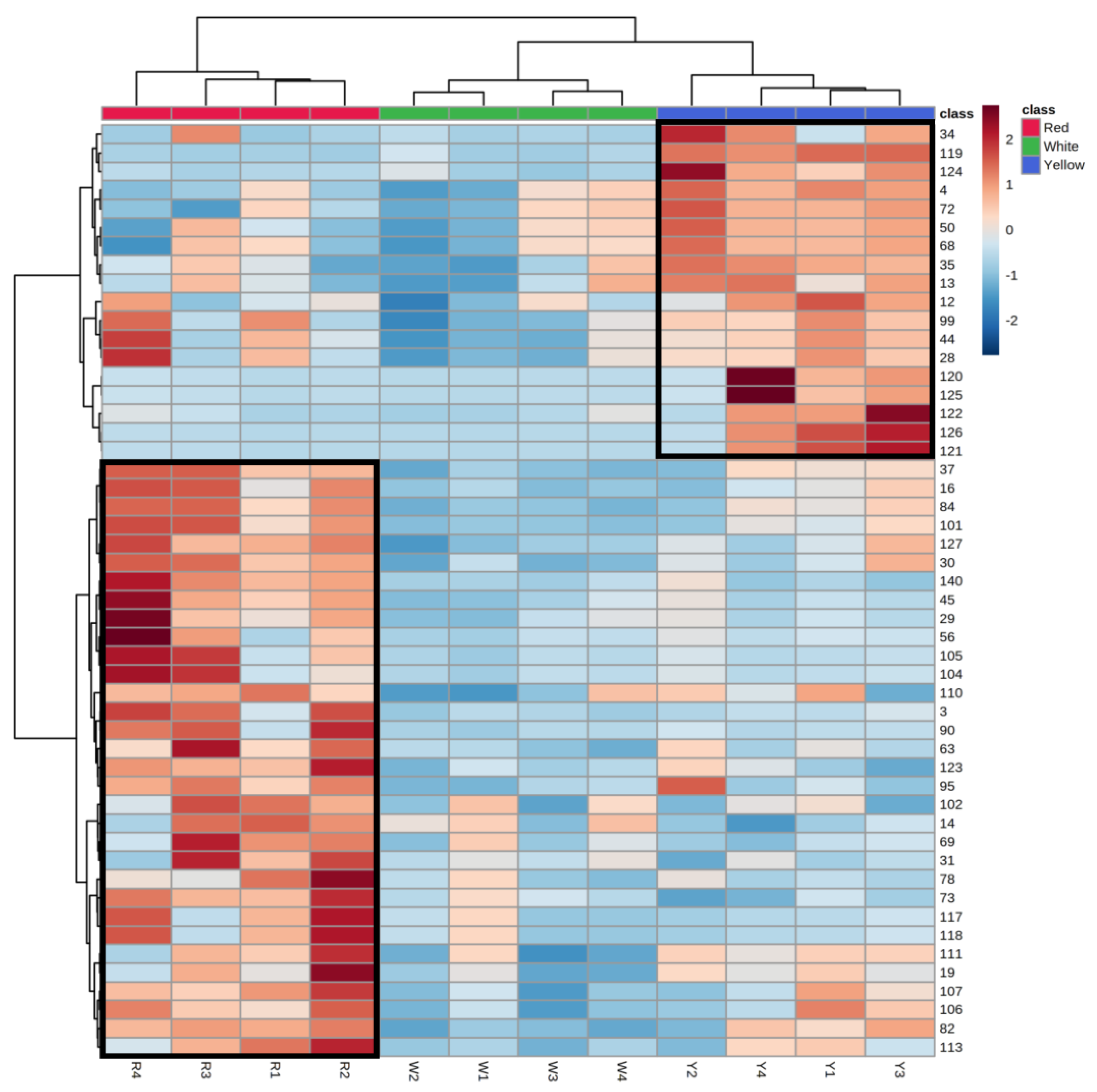
3.3. Untargeted Metabolomic Analysis of Different A. cinnamomea Phenotypes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Geethangili, M.; Tzeng, Y.M. Review of pharmacological effects of Antrodia camphorata and its bioactive compounds. Evid. Based Complement. Alternat. Med. 2011, 2011, 212641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuang, Y.; Li, B.; Wang, Z.; Qiao, X.; Ye, M. Terpenoids from the medicinal mushroom Antrodia camphorata: Chemistry and medicinal potential. Nat. Prod. Rep. 2021, 38, 83–102. [Google Scholar] [CrossRef] [PubMed]
- Qiao, X.; Wang, Q.; Ji, S.; Huang, Y.; Liu, K.D.; Zhang, Z.X.; Bo, T.; Tzeng, Y.M.; Guo, D.A.; Ye, M. Metabolites identification and multi-component pharmacokinetics of ergostane and lanostane triterpenoids in the anticancer mushroom Antrodia cinnamomea. J. Pharm. Biomed. Anal. 2015, 111, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.Y.; Chen, C.Y.; Chien, S.C.; Hsiao, W.W.; Chu, F.H.; Li, W.H.; Lin, C.C.; Shaw, J.F.; Wang, S.Y. Metabolite profiles for Antrodia cinnamomea fruiting bodies harvested at different culture ages and from different wood substrates. J. Agric. Food Chem. 2011, 59, 7626–7635. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.J.; Deng, J.S.; Huang, S.S.; Lee, C.Y.; Hou, W.C.; Wang, S.Y.; Sung, P.J.; Kuo, Y.H. Hepatoprotective effects of eburicoic acid and dehydroeburicoic acid from Antrodia camphorata in a mouse model of acute hepatic injury. Food Chem. 2013, 141, 3020–3027. [Google Scholar] [CrossRef]
- Chen, Y.C.; Liu, Y.L.; Li, F.Y.; Chang, C.I.; Wang, S.Y.; Lee, K.Y.; Li, S.L.; Chen, Y.P.; Jinn, T.R.; Tzen, J.T. Antcin A, a steroid-like compound from Antrodia camphorata, exerts anti-inflammatory effect via mimicking glucocorticoids. Acta Pharmacol. Sin. 2011, 32, 904–911. [Google Scholar] [CrossRef]
- Hsieh, Y.C.; Rao, Y.K.; Whang-Peng, J.; Huang, C.Y.; Shyue, S.K.; Hsu, S.L.; Tzeng, Y.M. Antcin B and its ester derivative from Antrodia camphorata induce apoptosis in hepatocellular carcinoma cells involves enhancing oxidative stress coincident with activation of intrinsic and extrinsic apoptotic pathway. J. Agric. Food Chem. 2011, 59, 10943–10954. [Google Scholar] [CrossRef]
- Huo, Y.; Win, S.; Than, T.A.; Yin, S.; Ye, M.; Hu, H.; Kaplowitz, N. Antcin H protects against acute liver injury through disruption of the interaction of c-Jun-N-terminal kinase with mitochondria. Antioxid. Redox Signal. 2017, 26, 207–220. [Google Scholar] [CrossRef]
- Kuo, Y.H.; Lin, C.H.; Shih, C.C. Antidiabetic and antihyperlipidemic properties of a triterpenoid compound, dehydroeburicoic acid, from Antrodia camphorata in vitro and in streptozotocin-induced mice. J. Agric. Food Chem. 2015, 63, 10140–10151. [Google Scholar] [CrossRef]
- Lin, C.H.; Kuo, Y.H.; Shih, C.C. Eburicoic acid, a triterpenoid compound from Antrodia camphorata, displays antidiabetic and antihyperlipidemic effects in palmitate treated C2C12 myotubes and in high-fat diet-fed mice. Int. J. Mol. Sci. 2017, 18, 2314. [Google Scholar] [CrossRef]
- Yeh, C.T.; Huang, W.C.; Rao, Y.K.; Ye, M.; Lee, W.H.; Wang, L.S.; Tzeng, D.T.; Wu, C.H.; Shieh, Y.S.; Huang, C.Y.; et al. A sesquiterpene lactone antrocin from Antrodia camphorata negatively modulates JAK2/STAT3 signaling via microRNA let-7c and induces apoptosis in lung cancer cells. Carcinogenesis 2013, 34, 2918–2928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tikunov, Y. A novel approach for nontargeted data analysis for metabolomics. Large-scale profiling of tomato fruit volatiles. Plant Physiol. 2005, 139, 1125–1137. [Google Scholar] [CrossRef] [Green Version]
- Arbona, V.; Iglesias, D.J.; Talón, M.; Gómez-Cadenas, A. Plant phenotype demarcation using nontargeted LC-MS and GC-MS metabolite profiling. J. Agric. Food Chem. 2009, 57, 7338–7347. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; Mohsen, M.; Heinke, R.; Wessjohann, L.A. Metabolomic fingerprints of 21 date palm fruit varieties from Egypt using UPLC/PDA/ESI–qTOF-MS and GC-MS analyzed by chemometrics. Food Res. Int. 2014, 64, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.W.; Xu, S.Y.; Sun, D.M.; Chen, W. Dehydration of concentrated Ganoderma lucidum extraction by combined microwave vacuum and conventional vacuum drying. Dry. Technol. 2006, 24, 595–599. [Google Scholar] [CrossRef]
- Dührkop, K.; Fleischauer, M.; Ludwig, M.; Aksenov, A.A.; Melnik, A.V.; Meusel, M.; Dorrestein, P.C.; Rousu, J.; Böcker, S. SIRIUS 4: A rapid tool for turning tandem mass spectra into metabolite structure information. Nat. Methods 2019, 16, 299–302. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.L.; Lee, Y.R.; Tsao, N.W.; Wang, S.Y.; Shaw, J.F.; Chu, F.H. Characterization of the 2,3-oxidosqualene cyclase gene from Antrodia cinnamomea and enhancement of cytotoxic triterpenoid compound production. J. Nat. Prod. 2015, 78, 1556–1562. [Google Scholar] [CrossRef]
- Chinese National Standards (CNS Number 16152); CNS. Fruiting Body of Niu-Chang-Chih (Ku). The Ministry of Economic Affairs of Taiwan: Taipei City, Taiwan, 2021; p. 15.
- Chung, C.H.; Yeh, S.C.; Tseng, H.C.; Siu, M.L.; Lee, K.T. Chemical quality evaluation of Antrodia cinnamomea fruiting bodies using phytomics similarity index analysis. J. Food Drug Anal. 2016, 24, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Liao, S.C.; Wen, F.C.; Chien, W.C.; Liao, Y.H. Anticancer effects of different extracts of various Antrodia cinnamomea types on human liver HepG2 cells and analyses of compositions and bioactivities. Int. J. Adv. Sci. Eng. 2017, 5, 41–51. [Google Scholar]
- Liu, S.C.; Wu, T.Y.; Hsu, T.H.; Lai, M.N.; Wu, Y.C.; Ng, L.T. Chemical composition and chronic toxicity of disc-cultured Antrodia cinnamomea fruiting bodies. Toxics 2022, 10, 587. [Google Scholar] [CrossRef]
- Zhang, B.B.; Guan, Y.Y.; Hu, P.F.; Chen, L.; Xu, G.R.; Liu, L.; Cheung, P.C.K. Production of bioactive metabolites by submerged fermentation of the medicinal mushroom Antrodia cinnamomea: Recent advances and future development. Crit. Rev. Biotechnol. 2019, 39, 541–554. [Google Scholar] [CrossRef]
- Zeng, W.W.; Chen, T.C.; Liu, C.H.; Wang, S.Y.; Shaw, J.F.; Chen, Y.T. Identification and isolation of an intermediate metabolite with dual antioxidant and anti-proliferative activity present in the fungus Antrodia cinnamomea cultured on an alternative medium with Cinnamomum kanehirai leaf extract. Plants 2021, 10, 737. [Google Scholar] [CrossRef] [PubMed]
- Chiu, H.H.; Lee, F.P.; Wang, J.K.; Chou, C.C. Identification and phylogenetic analysis of Antrodia camphorata and related species based on the polymorphic D2 region of LSU rDNA. Fooyin J. Health Sci. 2009, 1, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Li, H.X.; Wang, J.J.; Lu, C.L.; Gao, Y.J.; Gao, L.; Yang, Z.Q. Review of bioactivity, isolation, and identification of active compounds from Antrodia cinnamomea. Bioengineering 2022, 9, 494. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Lou, H.X. Strategies to diversify natural products for drug discovery. Med. Res. Rev. 2018, 38, 1255–1294. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.L.; Ho, Y.P.; Chou, J.C. Phenologic variation of major triterpenoids in regular and white Antrodia cinnamomea. Bot. Stud. 2016, 57, 33. [Google Scholar] [CrossRef] [Green Version]
- Ma, T.W.; Lai, Y.; Chen, L.T.; Yang, F.C. The cultivation strategy of enhancing triterpenoid production in submerged cultures of Antrodia cinnamomea by adding monoterpenes. J. Taiwan Inst. Chem. Eng. 2016, 58, 210–218. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, Y.; Yuan, X.L.; Luo, Y.N.; Luo, M.N.; Zheng, Y. Effects of culture mechanism of Cinnamomum kanehirae and C. camphora on the expression of genes related to terpene biosynthesis in Antrodia cinnamomea. Mycobiology 2022, 50, 121–131. [Google Scholar] [CrossRef]
- Ma, T.W.; Lai, Y.; Yang, F.C. Enhanced production of triterpenoid in submerged cultures of Antrodia cinnamomea with the addition of citrus peel extract. Bioprocess Biosyst. Eng. 2014, 37, 2251–2261. [Google Scholar] [CrossRef]
- Shu, C.H.; Wu, C.J.; Hsiao, W.J. Enhancement of triterpenoids production of Antrodia cinnamomea by co-culture with Saccharomyces cerevisiae. J. Bioprocess Biotech. 2015, 5, 253. [Google Scholar]
- Li, H.X.; Lu, Z.M.; Geng, Y.; Gong, J.S.; Zhang, X.J.; Shi, J.S.; Xu, Z.H.; Ma, Y.H. Efficient production of bioactive metabolites from Antrodia camphorata ATCC 200183 by asexual reproduction-based repeated batch fermentation. Bioresour. Technol. 2015, 194, 334–343. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HRMS (m/z) | ||||||
|---|---|---|---|---|---|---|
| Identity | Triterpenoid Types | Formula | tR (min) | Calculated | Measured | ∆m/z (ppm) |
| Antcin K, S + R | Ergostanes | C29H44O6 | 4.00 | 487.3065 | 487.3061 | −0.8 |
| Camphoratin A | Ergostanes | C29H44O6 | 4.31 | 487.3065 | 487.3062 | −0.6 |
| 12-OH antcamphin E/F | Ergostanes | C29H42O7 | 4.46 | 501.2858 | 501.2853 | −1.0 |
| 12-OH antcamphin E/F | Ergostanes | C29H42O7 | 4.59 | 501.2858 | 501.2852 | −1.2 |
| Hydrated antcin F (∆14) | Ergostanes | C29H42O6 | 4.87 | 485.2909 | 485.2906 | −0.6 |
| Antcamphin E, S | Ergostanes | C29H42O6 | 5.16 | 485.2909 | 485.2905 | −0.8 |
| Antcamphin K, S | Ergostanes | C29H44O5 | 5.16 | 471.3116 | 471.3113 | −0.6 |
| Antcamphin F, R | Ergostanes | C29H42O6 | 5.23 | 485.2909 | 485.2904 | −1.0 |
| Antcamphin L, R | Ergostanes | C29H44O5 | 5.24 | 471.3116 | 471.3113 | −0.6 |
| Antcin F, S + R | Ergostanes | C29H40O5 | 5.90 | 467.2803 | 467.2801 | −0.4 |
| Antcin C, S | Ergostanes | C29H42O5 | 6.04 | 469.2959 | 469.2957 | −0.4 |
| Antcin C, R | Ergostanes | C29H42O5 | 6.25 | 469.2959 | 469.2956 | −0.6 |
| Antcin H (zhankuic acid C), R | Ergostanes | C29H42O6 | 6.53 | 485.2909 | 485.2906 | −0.6 |
| Antcin H, S | Ergostanes | C29H42O6 | 6.71 | 485.2909 | 485.2905 | −0.8 |
| Dehydrosulfurenic acid | Ergostanes | C31H48O4 | 7.97 | 483.3480 | 483.3475 | −1.0 |
| Antcin I (zhankuic acid B), R | Ergostanes | C29H42O5 | 8.13 | 469.2959 | 469.2957 | −0.4 |
| Antcin I, S | Ergostanes | C29H42O5 | 8.15 | 469.2959 | 469.2957 | −0.4 |
| Sulfurenic acid | Ergostanes | C31H50O4 | 8.16 | 485.3636 | 485.3632 | −0.8 |
| Antcin B (zhankuic acid A), S | Ergostanes | C29H40O5 | 8.41 | 467.2803 | 467.2800 | −0.6 |
| Antcin G (25R/S epimer) | Ergostanes | C29H44O6 | 8.78 | 511.3065 | 511.3061 | −0.8 |
| Antcin A, S + R | Ergostanes | C29H40O4 | 10.02 | 453.3010 | 453.3008 | −0.4 |
| 15α-Acetyldehydrosulfurenic acid | Lanostanes | C33H50O5 | 10.69 | 525.3585 | 525.3582 | −0.6 |
| Versisponic acid D | Lanostanes | C33H52O5 | 11.00 | 527.3742 | 527.3738 | −0.8 |
| 3β-Hydroxylanosta-7,9(11), 24(28)-triene-21-oic acid | Lanostanes | C30H46O3 | 13.12 | 453.3374 | 453.3372 | −0.4 |
| Dehydroeburicoic acid | Lanostanes | C31H48O3 | 13.73 | 467.3531 | 467.3528 | −0.6 |
| Eburicoic acid | Lanostanes | C31H50O3 | 14.05 | 469.3687 | 469.3683 | −0.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, C.-H.; Hsieh, Y.-C.; Chng, J.-Y.; Lai, M.-N.; Ng, L.-T. Metabolomic Profiling of Different Antrodia cinnamomea Phenotypes. J. Fungi 2023, 9, 97. https://doi.org/10.3390/jof9010097
Su C-H, Hsieh Y-C, Chng J-Y, Lai M-N, Ng L-T. Metabolomic Profiling of Different Antrodia cinnamomea Phenotypes. Journal of Fungi. 2023; 9(1):97. https://doi.org/10.3390/jof9010097
Chicago/Turabian StyleSu, Chun-Han, Yun-Cheng Hsieh, Jin-Yi Chng, Ming-Nan Lai, and Lean-Teik Ng. 2023. "Metabolomic Profiling of Different Antrodia cinnamomea Phenotypes" Journal of Fungi 9, no. 1: 97. https://doi.org/10.3390/jof9010097