
The SUMOylation Pathway Components Are Required for Vegetative Growth, Asexual Development, Cytotoxic Responses, and Programmed Cell Death Events in Fusarium oxysporum f. sp. niveum

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strain and Growth Conditions
2.2. Bioinformatics Analyses
2.3. Generation of Targeted Deletion and Complementation Strains
2.4. Fungal Growth, Conidiation, and Stress Tolerance Assays
2.5. Fluorescence Microscopy
2.6. Transmission Electron Microscopy (TEM)
2.7. RT-qPCR Analyses
2.8. Western Blotting Assays
2.9. Statistical Analysis
3. Results
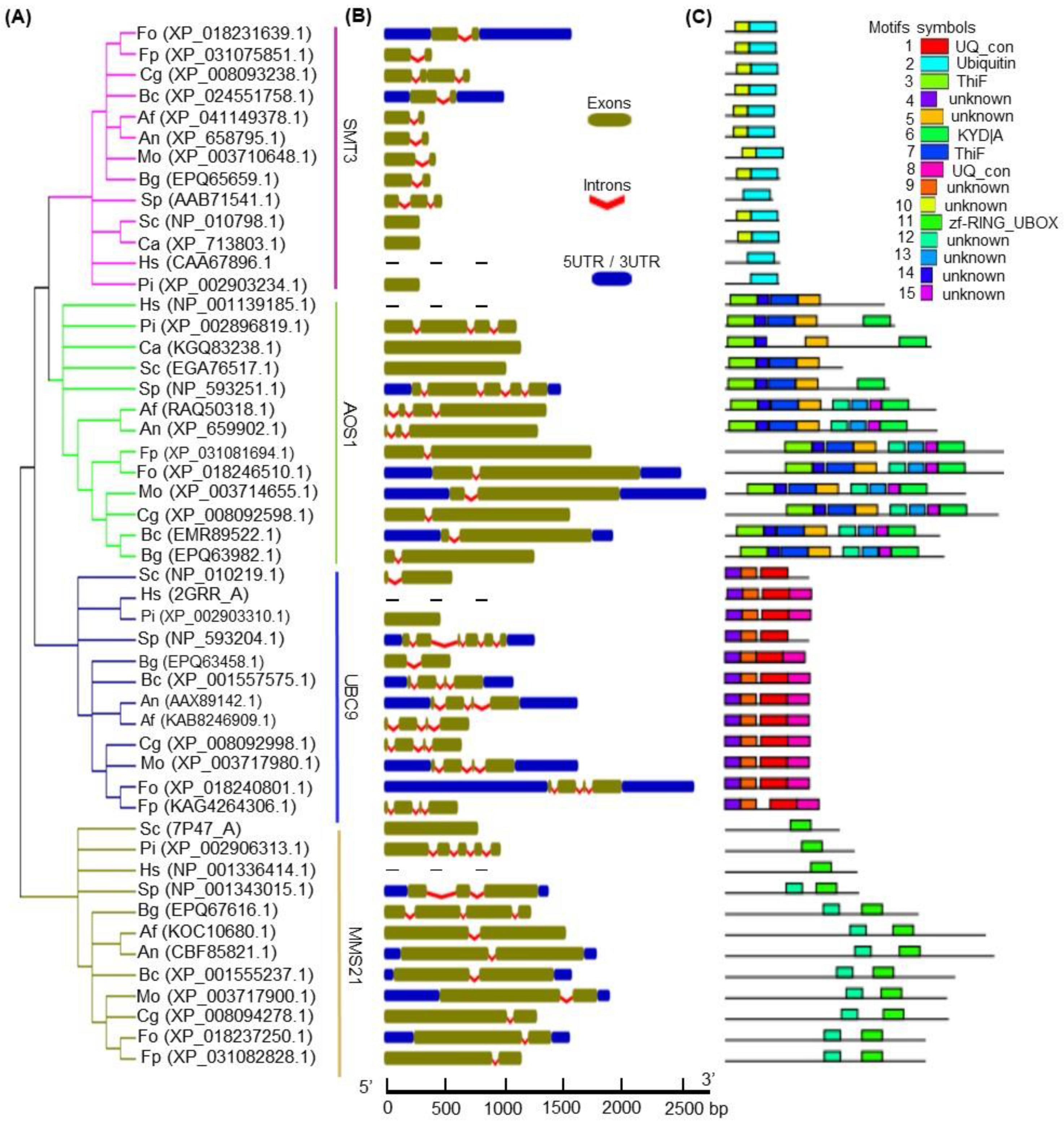
3.1. Identification of the SUMOylation Pathway Components in Fon
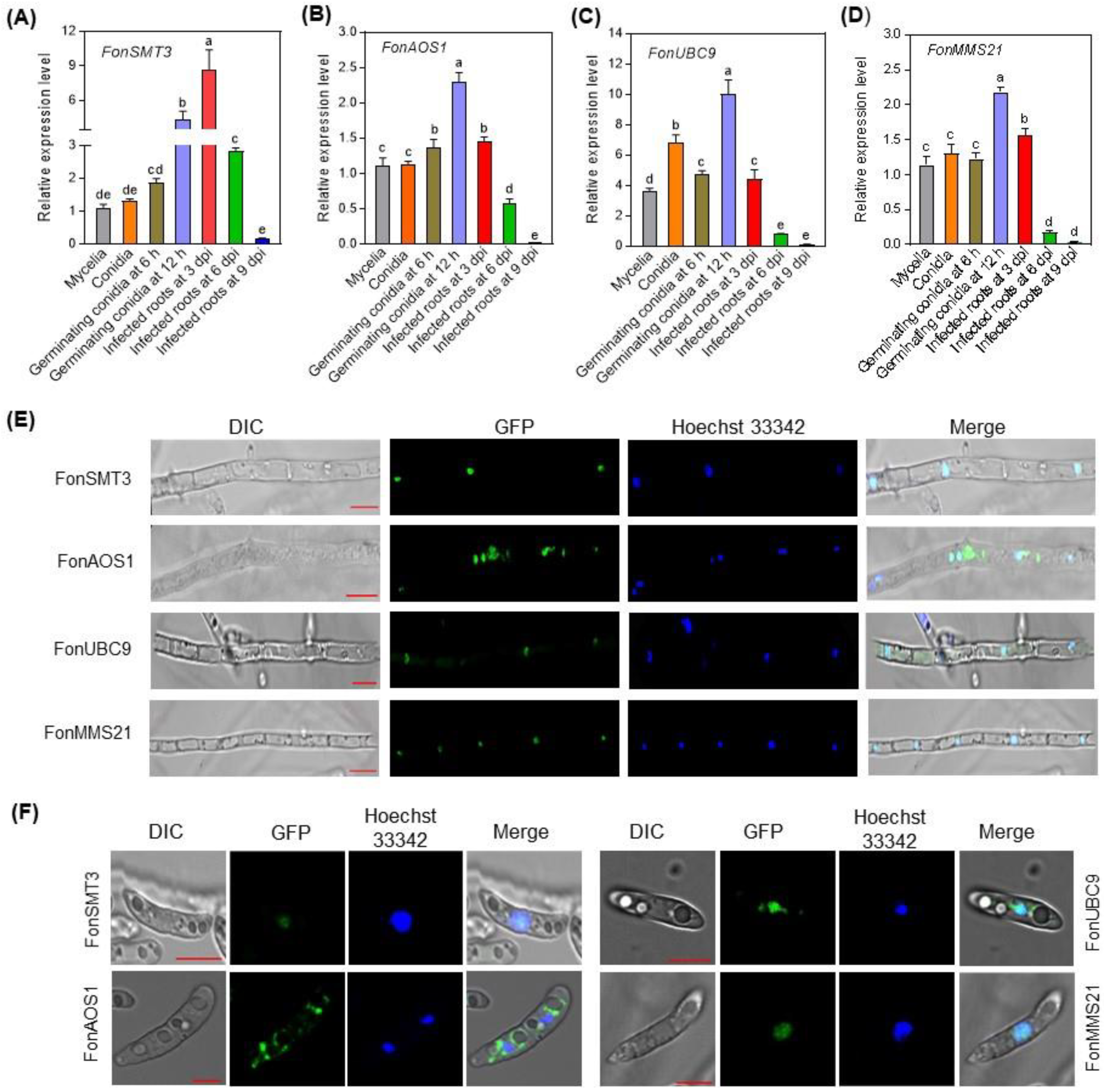
3.2. Expression Profiling of the SUMOylation Pathway Genes in Fon
3.3. Generation and Characterization of the Deletion Mutants and Complementation Strains
3.4. Subcellular Localzation of the SUMOylation Pathway Components in Fon
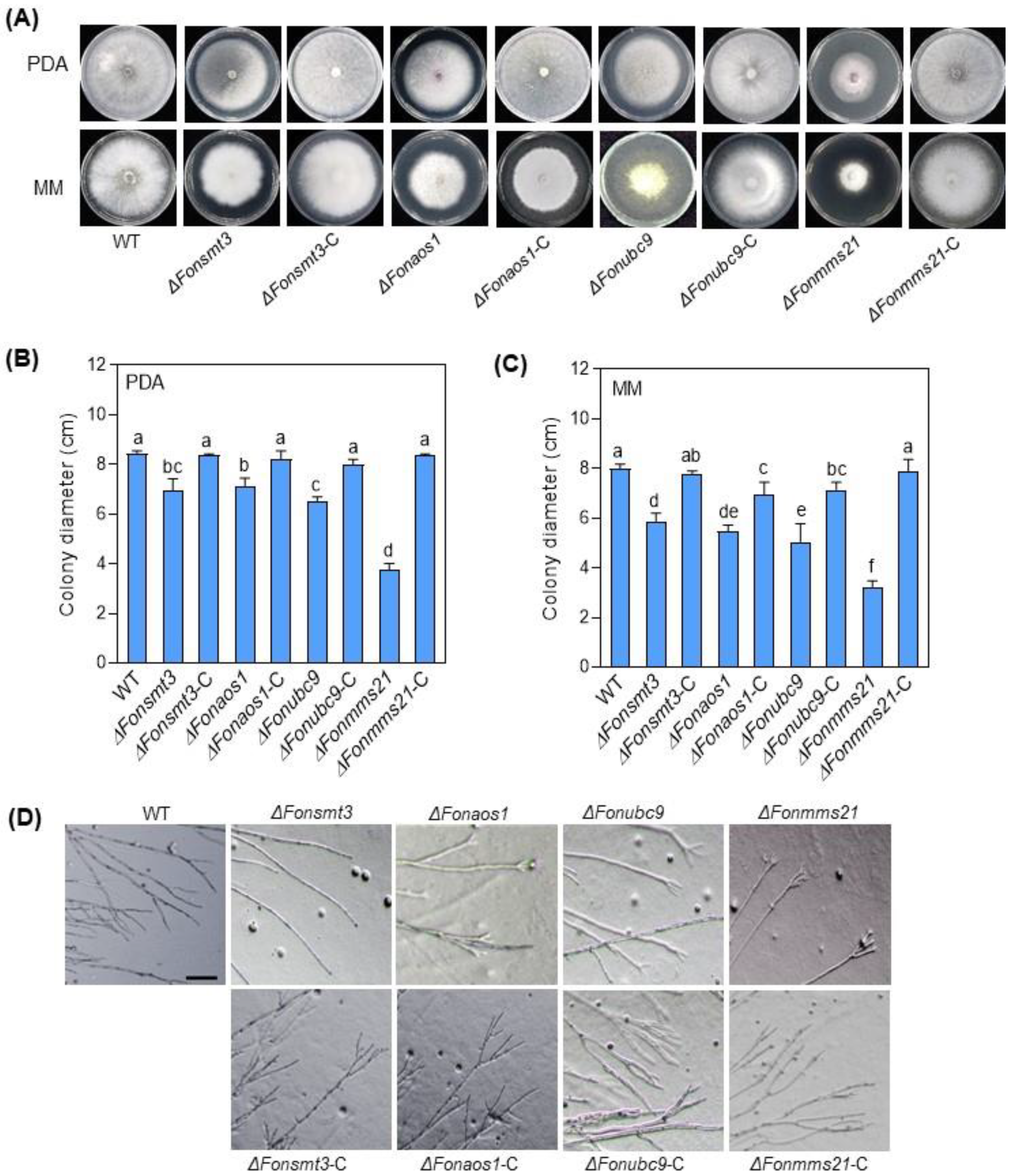
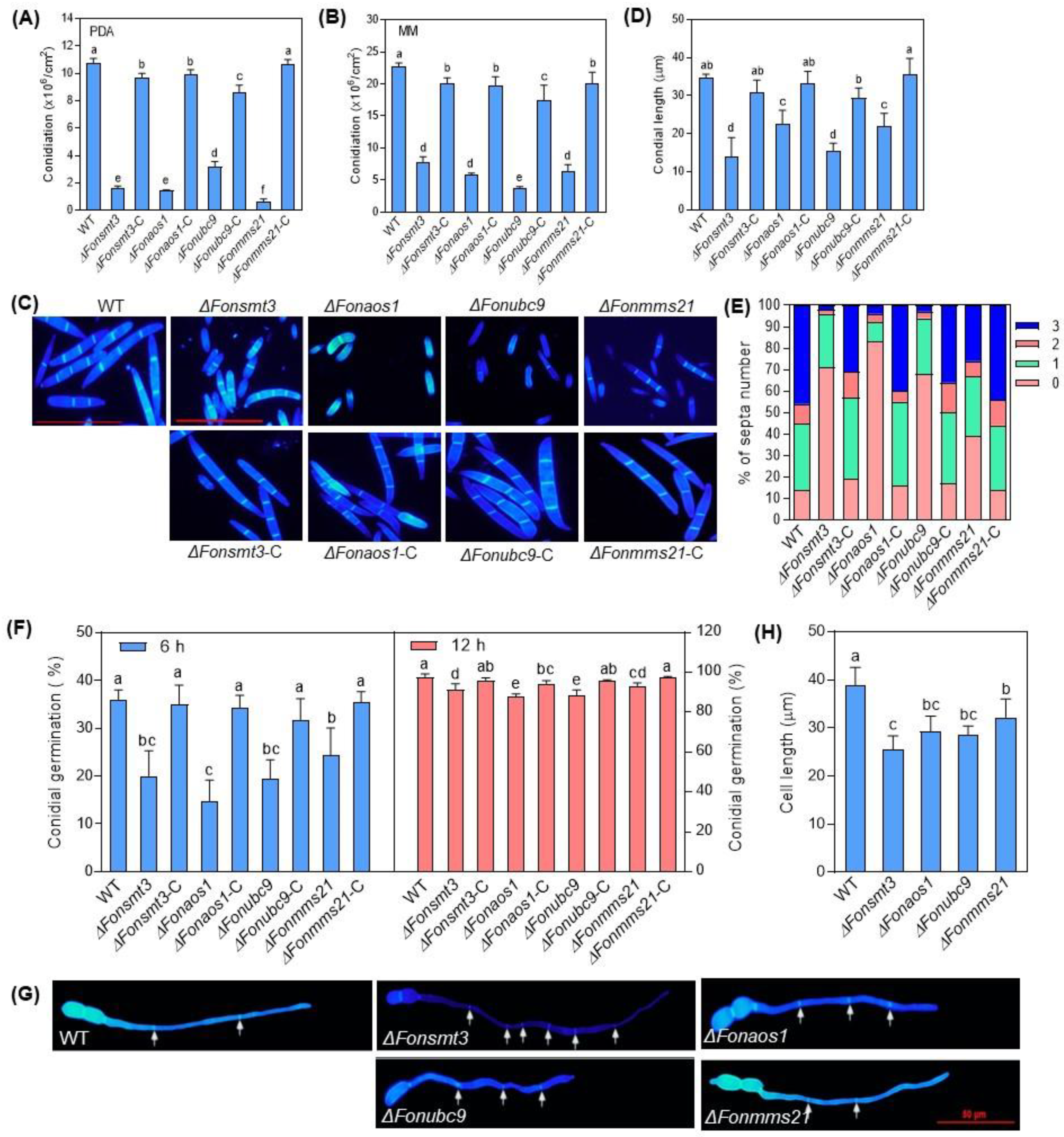
3.5. The SUMOylation Pathway Regulates the Growth and Development in Fon
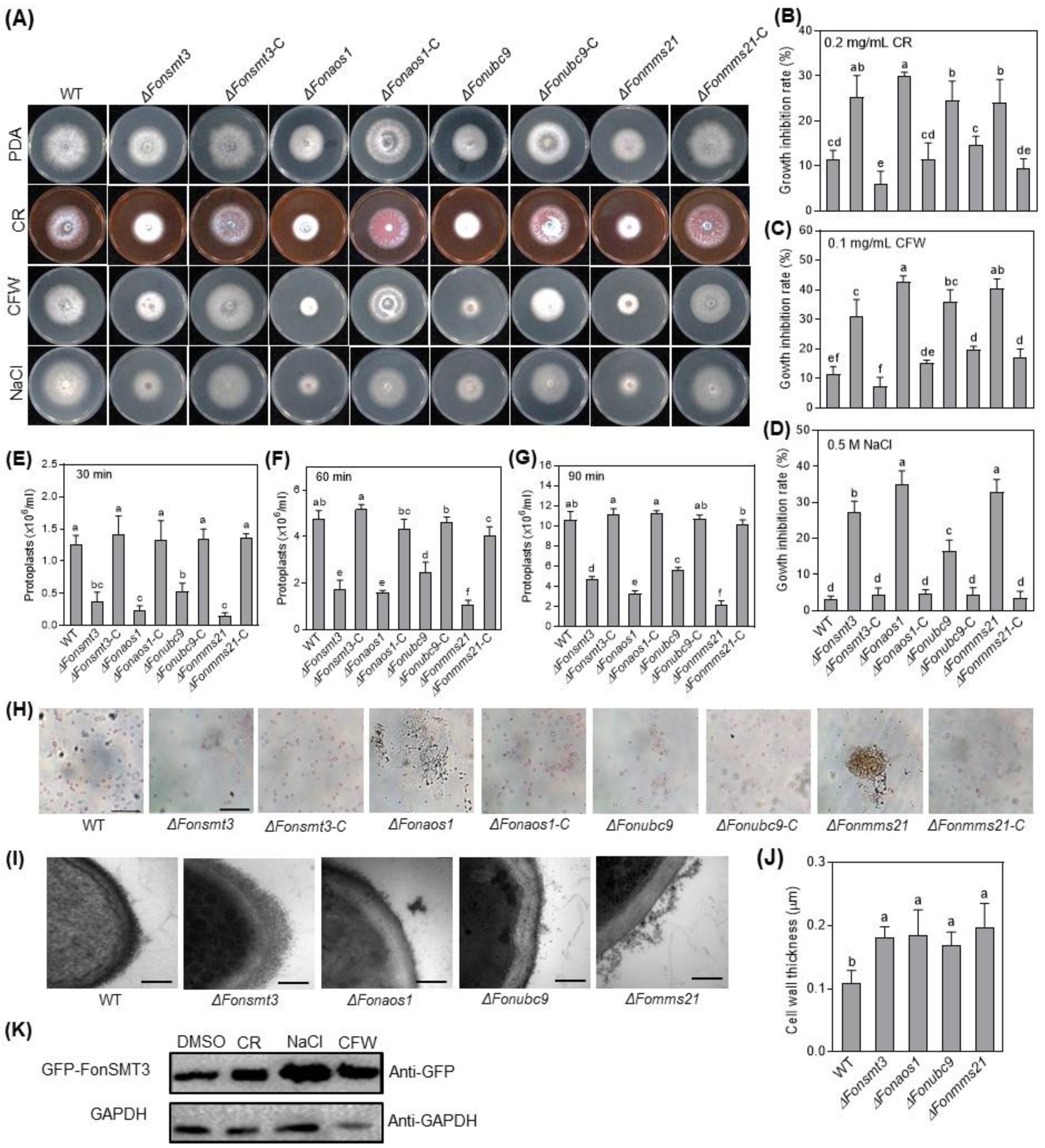
3.6. The SUMOylation Pathway Maintains Cell Wall Integrity in Fon
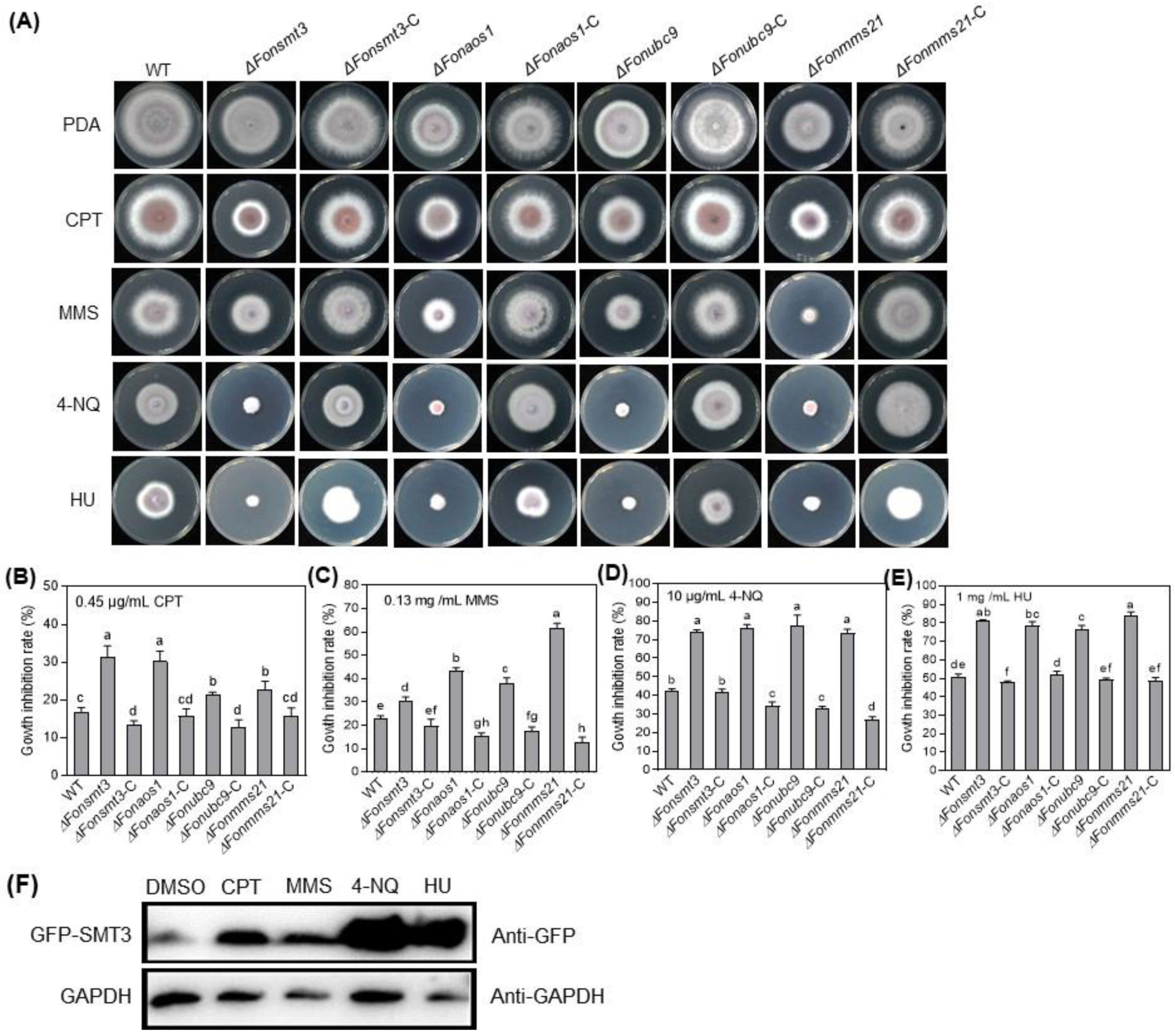
3.7. The SUMOylation Pathway Orchestrates DNA Damage Responses in Fon
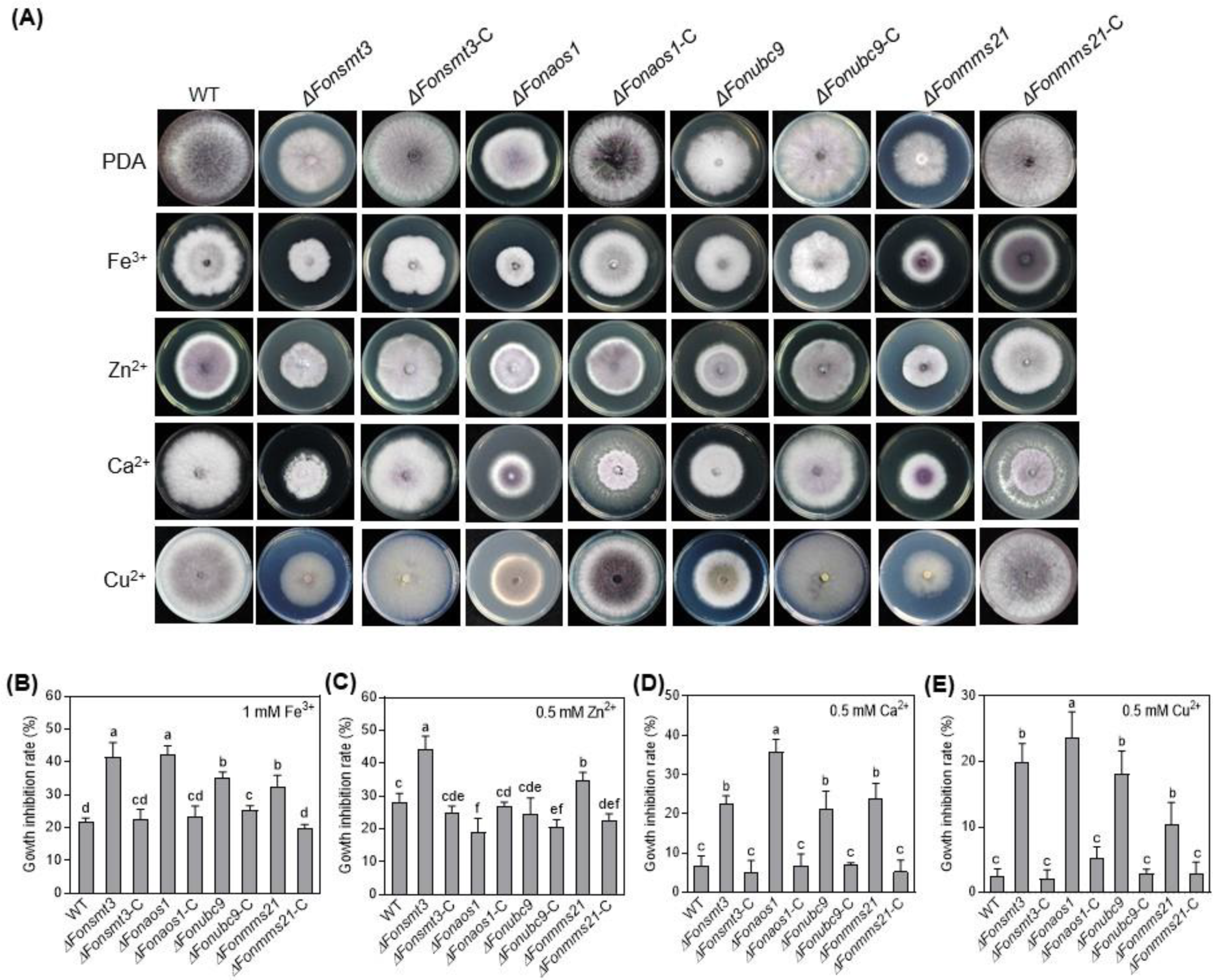
3.8. The SUMOylation Pathway Affects Metal ion Responses in Fon
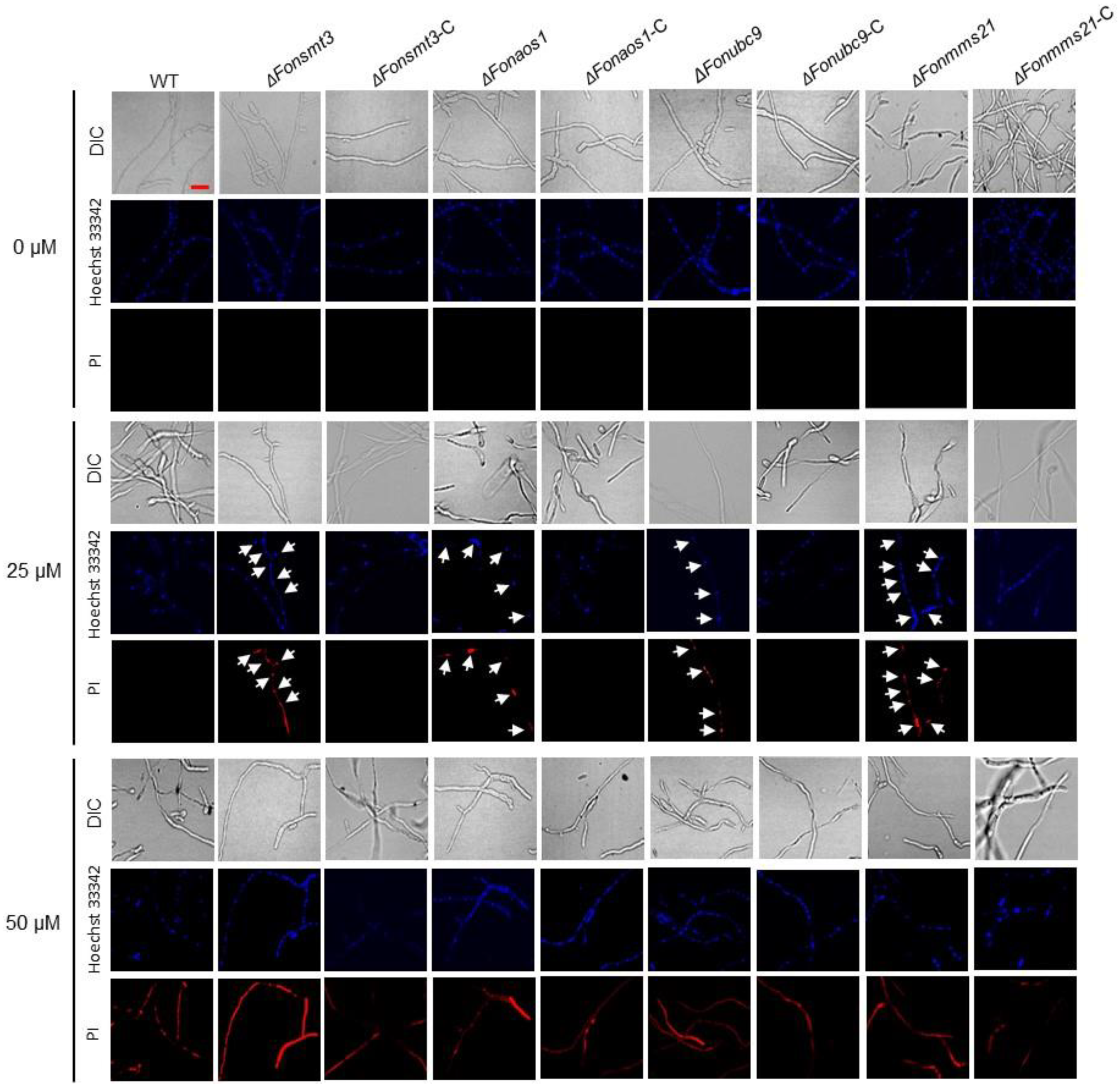
3.9. The SUMOylation Pathway Negativelty Regulates Apoptosis in Fon
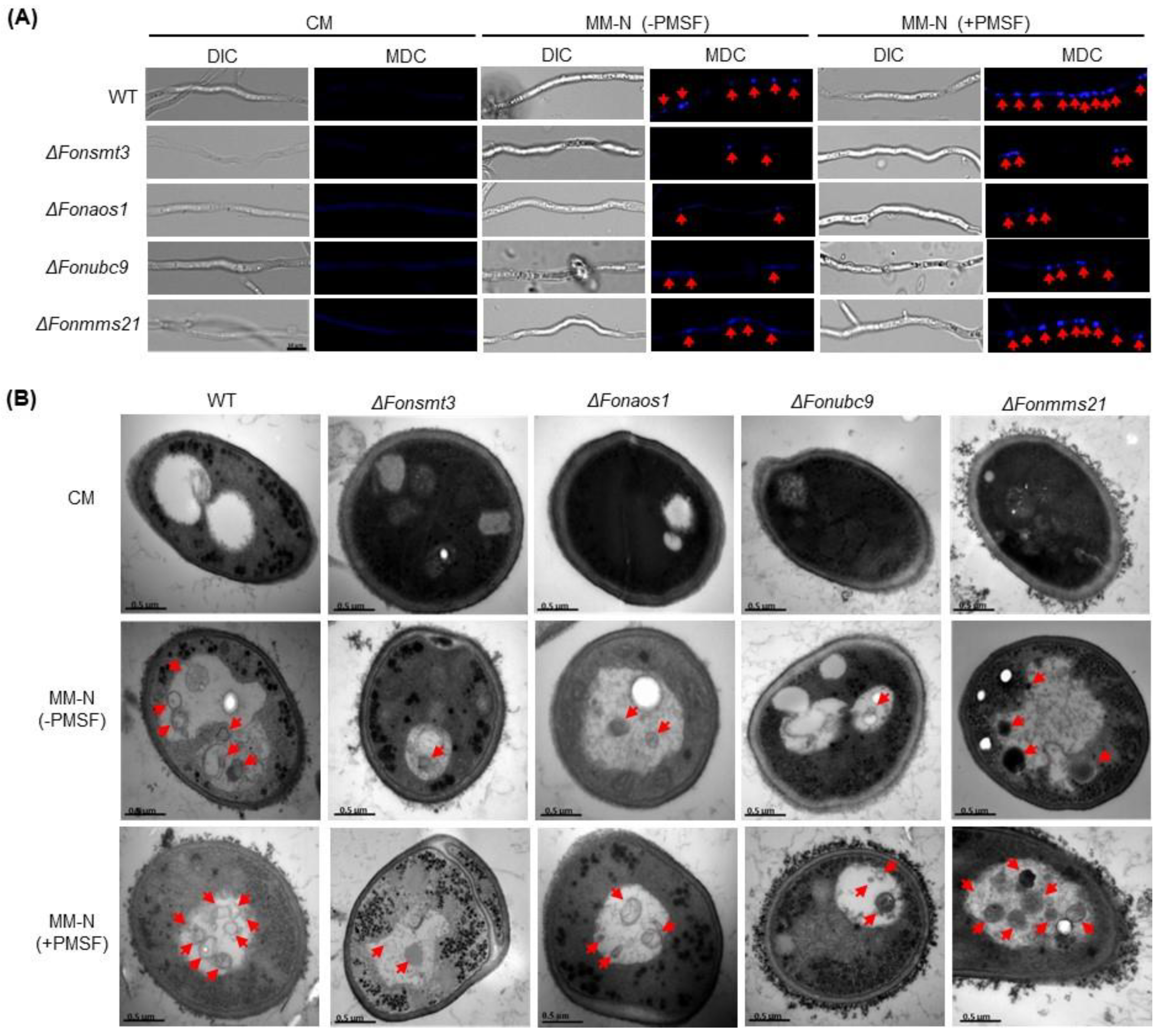
3.10. The SUMOylatin Pathway Mediates Autophagy in Fon
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gupta, D.; Garapati, H.S.; Kakumanu, A.V.S.; Shukla, R.; Mishra, K. SUMOylation in fungi: A potential target for intervention. Comput. Struct. Biotechnol. J. 2020, 18, 3484–3493. [Google Scholar] [CrossRef] [PubMed]
- Melchior, F. SUMO—Nonclassical ubiquitin. Annu. Rev. Cell Dev. Biol. 2000, 16, 591–626. [Google Scholar] [CrossRef] [PubMed]
- Gareau, J.R.; Lima, C.D. The SUMO pathway: Emerging mechanisms that shape specificity, conjugation and recognition. Nat. Rev. Mol. Cell Biol. 2010, 11, 861–871. [Google Scholar] [CrossRef] [Green Version]
- Bayer, P.; Arndt, A.; Metzger, S.; Mahajan, R.; Melchior, F.; Jaenicke, R.; Becker, J. Structure determination of the small ubiquitin-related modifier SUMO-1. J. Mol. Biol. 1998, 280, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, K.A.; Henley, J.M. Mechanisms, regulation and consequences of protein SUMOylation. Biochem. J. 2010, 428, 133–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.J.; Kim, W.Y.; Park, H.C.; Lee, S.Y.; Bohnert, H.J.; Yun, D.J. SUMO and SUMOylation in plants. Mol. Cells 2011, 32, 305–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, E.S.; Schwienhorst, I.; Dohmen, R.J.; Blobel, G. The ubiquitin-like protein Smt3p is activated for conjugation to other proteins by an Aos1p/Uba2p heterodimer. EMBO J. 1997, 16, 5509–5519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Li, Z.; Xing, J.; Yang, J.; Wang, Z.; Zhang, H.; Chen, D.; Peng, Y.L.; Chen, X.L. Global analysis of sumoylation function reveals novel insights into development and appressorium-mediated infection of the rice blast fungus. New Phytol. 2018, 219, 1031–1047. [Google Scholar] [CrossRef]
- Lim, Y.J.; Kim, K.T.; Lee, Y.H. SUMOylation is required for fungal development and pathogenicity in the rice blast fungus Magnaporthe oryzae. Mol. Plant Pathol. 2018, 19, 2134–2148. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Nishide, J.; Okazaki, K.; Kato, H.; Niwa, O.; Nakagawa, T.; Matsuda, H.; Kawamukai, M.; Murakami, Y. Characterization of a fission yeast SUMO-1 homologue, pmt3p, required for multiple nuclear events, including the control of telomere length and chromosome segregation. Mol. Cell. Biol. 1999, 19, 8660–8672. [Google Scholar] [CrossRef]
- Leach, M.D.; Stead, D.A.; Argo, E.; Brown, A.J.P. Identification of sumoylation targets, combined with inactivation of SMT3, reveals the impact of sumoylation upon growth, morphology, and stress resistance in the pathogen Candida albicans. Mol. Biol. Cell 2011, 22, 687–702. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.H.; Todd, R.B.; Oakley, B.R.; Oakley, C.E.; Hynes, M.J.; Davis, M.A. Sumoylation in Aspergillus nidulans: SumO inactivation, overexpression and live-cell imaging. Fungal Genet. Biol. 2008, 45, 728–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, X.; Yu, S.; Qiu, M.; Wang, X.; Wang, Y.; Bai, Y.; Zhang, F.; Wang, S. Aspergillus flavus SUMO contributes to fungal virulence and toxin attributes. J. Agric. Food Chem. 2016, 64, 6772–6782. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Meng, X.; Chen, J.; Fang, Q.; Wang, J. Smt3, a homologue of yeast SUMO, contributes to asexual development, environmental adaptation, and host infection of a filamentous entomopathogen. Fungal Biol. 2020, 124, 924–931. [Google Scholar] [CrossRef] [PubMed]
- Geiss-Friedlander, R.; Melchior, F. Concepts in sumoylation: A decade on. Nat. Rev. Mol. Cell Biol. 2007, 8, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Sahu, M.S.; Patra, S.; Kumar, K.; Kaur, R. SUMOylation in human pathogenic fungi: Role in physiology and virulence. J. Fungi 2020, 6, 32. [Google Scholar] [CrossRef] [Green Version]
- Hickey, C.M.; Wilson, N.R.; Hochstrasser, M. Function and regulation of SUMO proteases. Nat. Rev. Mol. Cell Biol. 2012, 13, 755–766. [Google Scholar] [CrossRef] [Green Version]
- Desterro, J.M.P.; Rodriguez, M.S.; Kemp, G.D.; Hay, R.T. Identification of the enzyme required for activation of the small ubiquitin-like protein SUMO-1. J. Biol. Chem. 1999, 274, 10618–10624. [Google Scholar] [CrossRef] [Green Version]
- Tatham, M.H.; Jaffray, E.; Vaughan, O.A.; Desterro, J.M.P.; Botting, C.H.; Naismith, J.H.; Hay, R.T. Polymeric chains of SUMO-2 and SUMO-3 are conjugated to protein substrates by SAE1/SAE2 and Ubc9. J. Biol. Chem. 2001, 276, 35368–35374. [Google Scholar] [CrossRef] [Green Version]
- Sriramachandran, A.M.; Dohmen, R.J. SUMO-targeted ubiquitin ligases. Biochim. Biophys. Acta 2014, 1843, 75–85. [Google Scholar] [CrossRef]
- Lomelí, H.; Vázquez, M. Emerging roles of the SUMO pathway in development. Cell. Mol. Life Sci. 2011, 68, 4045–4064. [Google Scholar] [CrossRef] [PubMed]
- Dohmen, R.J. SUMO protein modification. Biochim. Biophys. Acta 2004, 1695, 113–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, E.S.; Blobel, G. Cell cycle-regulated attachment of the ubiquitin-related protein SUMO to the yeast septins. J. Cell Biol. 1999, 147, 981–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, A.; Tebbji, F.; Mallick, J.; Regan, H.; Dumeaux, V.; Omran, R.P.; Whiteway, M. Mms21: A putative SUMO E3 ligase in Candida albicans that negatively regulates invasiveness and filamentation, and is required for the genotoxic and cellular stress response. Genetics 2019, 211, 579–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harting, R.; Bayram, Ö.; Laubinger, K.; Valerius, O.; Braus, G.H. Interplay of the fungal sumoylation network for control of multicellular development. Mol. Microbiol. 2013, 90, 1125–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Yang, N.; Zheng, Y.; Yue, J.; Bhadauria, V.; Peng, Y.-L.; Chen, Q. Ubiquitination in the rice blast fungus Magnaporthe oryzae: From development and pathogenicity to stress responses. Phytopathol. Res. 2022, 4, 1. [Google Scholar] [CrossRef]
- Jian, Y.; Chen, X.; Sun, K.; Liu, Z.; Cheng, D.; Cao, J.; Liu, J.; Cheng, X.; Wu, L.; Zhang, F.; et al. SUMOylation regulates pre-mRNA splicing to overcome DNA damage in fungi. New Phytol. 2022. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Y.; Cao, Z.; Huang, L.; Liu, S.; Shen, Z.; Wang, Y.; Wang, H.; Zhang, H.; Li, D.; Song, F. CCR4-not complex subunit Not2 plays critical roles in vegetative growth, conidiation and virulence in watermelon Fusarium wilt pathogen Fusarium oxysporum f. sp. niveum. Front. Microbiol. 2016, 7, 1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Xiong, X.; Wang, H.; Wang, J.; Bi, Y.; Yan, Y.; Cao, Z.; Li, D.; Song, F. Ero1-Pdi1 module-catalysed dimerization of a nucleotide sugar transporter, FonNst2, regulates virulence of Fusarium oxysporum on watermelon. Environ. Microbiol. 2022, 24, 1200–1220. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, Z.; Huang, M.; Yan, L.; Ma, Z.; Yin, Y. The FgSsb-FgZuo-FgSsz complex regulates multiple stress responses and mycotoxin production via folding the soluble SNARE Vam7 and β2-tubulin in Fusarium graminearum. Environ. Microbiol. 2017, 19, 5040–5059. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Duan, Z.; Chen, P.; Shang, Y.; Wang, C. The Bax inhibitor MrBI-1 regulates heat tolerance, apoptotic-like cell death, and virulence in Metarhizium robertsii. Sci. Rep. 2015, 5, 10625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, M.; Hamid, M.I.; Wang, N.; Bin, L.; Xiang, M.; Liu, X. The transcription factor SKN7 regulates conidiation, thermotolerance, apoptotic-like cell death and parasitism in the nematode endoparasitic fungus Hirsutella minnesotensis. Sci. Rep. 2016, 6, 30047. [Google Scholar] [CrossRef] [Green Version]
- Corral-Ramos, C.; Roca, M.G.; Di Pietro, A.; Roncero, M.I.G.; Ruiz-Roldán, C. Autophagy contributes to regulation of nuclear dynamics during vegetative growth and hyphal fusion in Fusarium oxysporum. Autophagy 2015, 11, 131–144. [Google Scholar] [CrossRef] [Green Version]
- Khalid, A.R.; Zhang, S.; Luo, X.; Mehmood, K.; Rahim, J.; Shaheen, H.; Dong, P.; Qiu, D.; Ren, M. Role of autophagy-related gene atg22 in developmental process and virulence of Fusarium oxysporum. Genes 2019, 10, 365. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.A.; Lou, Y.; Hafeez, R.; Li, X.; Hossain, A.; Xie, T.; Lin, L.; Li, B.; Yin, Y.; Yan, J.; et al. Functional analysis and genome mining reveal high potential of biocontrol and plant growth promotion in nodule-inhabiting bacteria within Paenibacillus polymyxa complex. Front. Microbiol. 2021, 11, 618601. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Gill, G. SUMO and ubiquitin in the nucleus: Different functions, similar mechanisms? Genes Dev. 2004, 18, 2046–2059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truong, K.; Lee, T.D.; Li, B.; Chen, Y. Sumoylation of SAE2 C terminus regulates SAE nuclear localization. J. Biol. Chem. 2012, 287, 42611–42619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillies, J.; Hickey, C.M.; Su, D.; Wu, Z.; Peng, J.; Hochstrasser, M. SUMO pathway modulation of regulatory protein binding at the ribosomal DNA locus in Saccharomyces cerevisiae. Genetics 2016, 202, 1377–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enserink, J.M. Sumo and the cellular stress response. Cell Div. 2015, 10, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, Z.; Zhu, J.; Deng, Y.; Gao, S.; Liang, S. SUMOylation modification-mediated cell death. Open Biol. 2021, 11, 210050. [Google Scholar] [CrossRef]
- Huang, H.; du, G.; Chen, H.; Liang, X.; Li, C.; Zhu, N.; Xue, L.; Ma, J.; Jiao, R. Drosophila Smt3 negatively regulates JNK signaling through sequestering Hipk in the nucleus. Development 2011, 138, 2477–2485. [Google Scholar] [CrossRef] [Green Version]
- Fang, S.; Yuan, J.; Shi, Q.; Xu, T.; Fu, Y.; Wu, Z.; Guo, W. Downregulation of UBC9 promotes apoptosis of activated human LX-2 hepatic stellate cells by suppressing the canonical NF-κB signaling pathway. PLoS ONE 2017, 12, e0174374. [Google Scholar] [CrossRef] [Green Version]
- Potts, P.R.; Yu, H. Human MMS21/NSE2 is a SUMO ligase required for DNA repair. Mol. Cell. Biol. 2005, 25, 7021–7032. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.J.; Yun, S.M.; Jo, C.; Lee, D.H.; Choi, K.J.; Song, J.C.; Park, S.I.; Kim, Y.J.; Koh, Y.H. SUMO1 promotes Aβ production via the modulation of autophagy. Autophagy 2015, 11, 100–112. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Guo, C.; Lao, Y.; Yang, J.; Chen, F.; Zhao, Y.; Yang, Y.; Yang, J.; Yi, J. A fine-tuning mechanism underlying self-control for autophagy: DeSUMOylation of BECN1 by SENP3. Autophagy 2020, 16, 975–990. [Google Scholar] [CrossRef]
- Miura, K.; Rus, A.; Sharkhuu, A.; Yokoi, S.; Karthikeyan, A.S.; Raghothama, K.G.; Baek, D.; Koo, Y.D.; Jin, J.B.; Bressan, R.A.; et al. The Arabidopsis SUMO E3 ligase SIZ1 controls phosphate deficiency responses. Proc. Natl. Acad. Sci. USA 2005, 102, 17760–17765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazur, M.J.; Kwaaitaal, M.; Mateos, M.A.; Maio, F. The SUMO conjugation complex self-assembles into nuclear bodies independent of SIZ1 and COP1. Plant Physiol. 2019, 179, 168–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jmii, S.; Cappadocia, L. Plant SUMO E3 ligases: Function, structural organization, and connection with DNA. Front. Plant Sci. 2021, 12, 652170. [Google Scholar] [CrossRef] [PubMed]
- Ishida, T.; Fujiwara, S.; Miura, K.; Stacey, N.; Yoshimura, M.; Schneider, K.; Adachi, S.; Minamisawa, K.; Umeda, M.; Sugimoto, K. SUMO E3 ligase HIGH PLOIDY2 regulates endocycle onset and meristem maintenance in Arabidopsis. Plant Cell 2009, 21, 2284–2297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, S.E.; Matuschewski, K.; Liakopoulos, D.; Scheffner, M.; Jentsch, S. The ubiquitin-like proteins SMT3 and SUMO-1 are conjugated by the UBC9 E2 enzyme. Proc. Natl. Acad. Sci. USA 1998, 95, 560–564. [Google Scholar] [CrossRef] [Green Version]
- Nie, M.; Vashisht, A.A.; Wohlschlegel, J.A.; Boddy, M.N. High confidence fission yeast SUMO conjugates identified by tandem denaturing affinity purification. Sci. Rep. 2015, 5, 14389. [Google Scholar] [CrossRef] [Green Version]
- Geoghegan, I.; Steinberg, G.; Gurr, S. The role of the fungal cell wall in the infection of plants. Trends Microbiol. 2017, 25, 957–967. [Google Scholar] [CrossRef]
- Dichtl, K.; Samantaray, S.; Wagener, J. Cell wall integrity signalling in human pathogenic fungi. Cell. Microbiol. 2016, 18, 1228–1238. [Google Scholar] [CrossRef]
- Gujjula, R.; Veeraiah, S.; Kumar, K.; Thakur, S.S.; Mishra, K.; Kaur, R. Identification of components of the SUMOylation machinery in Candida glabrata: ROLE OF THE DESUMOYLATION PEPTIDASE CgUlp2 IN VIRULENCE. J. Biol. Chem. 2016, 291, 19573–19589. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Augustine, R.C.; Suzuki, M.; Feng, J.; Char, S.N.; Yang, B.; McCarty, D.R.; Vierstra, R.D. The SUMO ligase MMS21 profoundly influences maize development through its impact on genome activity and stability. PLoS Genet. 2021, 17, e1009830. [Google Scholar] [CrossRef]
- Nagai, S.; Dubrana, K.; Tsai-Pflugfelder, M.; Davidson, M.B.; Roberts, T.M.; Brown, G.W.; Varela, E.; Hediger, F.; Gasser, S.M.; Krogan, N.J. Functional targeting of DNA damage to a nuclear pore-associated SUMO-dependent ubiquitin ligase. Science 2008, 322, 597–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Cook, D.E. The contribution of DNA repair pathways to genome editing and evolution in filamentous pathogens. FEMS Microbiol. Rev. 2022, 46, fuac035. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Fang, H.; Chiou, S.; Yi, S.; Huang, C.; Chiang, S.; Chang, H.; Lin, T.; Chiang, I.; Chow, K. Sumoylation of eukaryotic elongation factor 2 is vital for protein stability and anti-apoptotic activity in lung adenocarcinoma cells. Cancer Sci. 2011, 102, 1582–1589. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; He, Y.; Wang, X.; Liang, Z.; He, G.; Zhang, P.; Zhu, H.; Xu, N.; Liang, S. Protein SUMOylation modification and its associations with disease. Open Biol. 2017, 7, 170167. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azizullah; Noman, M.; Gao, Y.; Wang, H.; Xiong, X.; Wang, J.; Li, D.; Song, F. The SUMOylation Pathway Components Are Required for Vegetative Growth, Asexual Development, Cytotoxic Responses, and Programmed Cell Death Events in Fusarium oxysporum f. sp. niveum. J. Fungi 2023, 9, 94. https://doi.org/10.3390/jof9010094
Azizullah, Noman M, Gao Y, Wang H, Xiong X, Wang J, Li D, Song F. The SUMOylation Pathway Components Are Required for Vegetative Growth, Asexual Development, Cytotoxic Responses, and Programmed Cell Death Events in Fusarium oxysporum f. sp. niveum. Journal of Fungi. 2023; 9(1):94. https://doi.org/10.3390/jof9010094
Chicago/Turabian StyleAzizullah, Muhammad Noman, Yizhou Gao, Hui Wang, Xiaohui Xiong, Jiajing Wang, Dayong Li, and Fengming Song. 2023. "The SUMOylation Pathway Components Are Required for Vegetative Growth, Asexual Development, Cytotoxic Responses, and Programmed Cell Death Events in Fusarium oxysporum f. sp. niveum" Journal of Fungi 9, no. 1: 94. https://doi.org/10.3390/jof9010094