
Taxonomy and Phylogeny of Peniophora Sensu Lato (Russulales, Basidiomycota)
Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological Studies
2.2. DNA Extraction and Sequencing
2.3. Phylogenetic Analyses
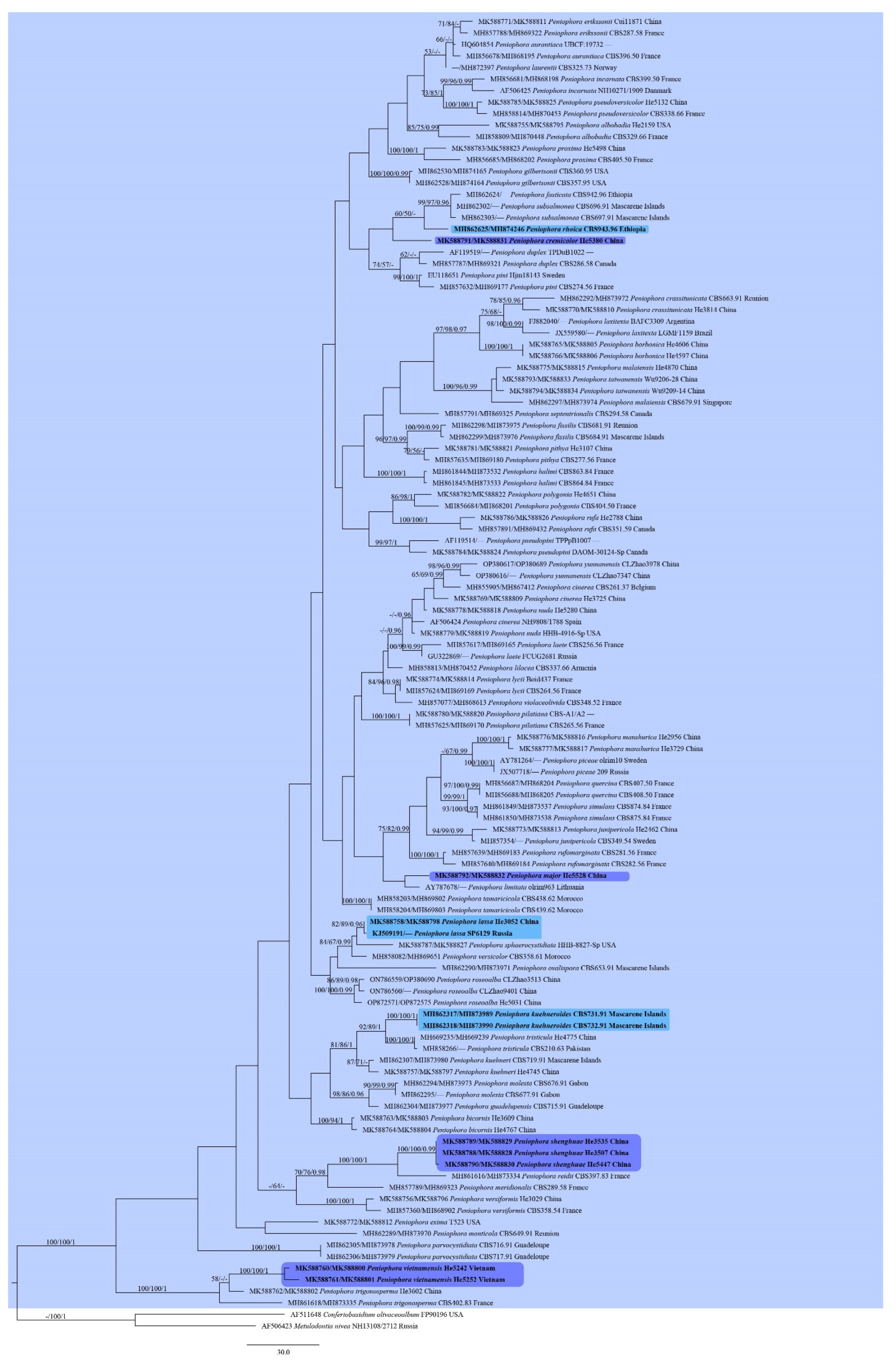
3. Results
3.1. Phylogenetic Analyses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Voucher | Locality | ITS | 28S | Reference |
|---|---|---|---|---|---|
| Amylostereum chailletii | NH8031/1035 | Romania | AF506406 | AF506406 | [6] |
| Asterostroma laxum | EL33-99 | Estonia | AF506410 | AF506410 | [6] |
| A. muscicola | KHL9537 | Puerto Rico | AF506409 | AF506409 | [6] |
| Baltazaria eurasiaticogalactina | CBS666.84 | France | — | AY293211 | [27] |
| B. octopodites | FLOR 56449 | Brazil | MH260025 | MH260047 | [28] |
| Confertobasidium olivaceoalbum | FP90196 | USA | AF511648 | AF511648 | [6] |
| Dichostereum effuscatum | GG930915 | France | AF506390 | AF506390 | [6] |
| D. pallescens | NH7046/673 | Canada | AF506392 | AF506392 | [6] |
| Gloiothele lactescens | EL8-98 | Sweden | AF506453 | AF506453 | [6] |
| G. lamellosa | KHL11031 | Venezuela | AF506454 | AF506454 | [6] |
| Lachnocladium schweinfurthianum | KM 49740 | Cameroon | MH260033 | MH260051 | [28] |
| L. sp. | KHL10556 | Jamaica | AF506461 | AF506461 | [6] |
| Metulodontia nivea | NH13108/2712 | Russia | AF506423 | AF506423 | [6] |
| Peniophora albobadia | CBS329.66 | France | MH858809 | MH870448 | [9] |
| P. albobadia | He2159 | USA | MK588755 | MK588795 | Present study |
| P. aurantiaca | UBCF:19732 | — | HQ604854 | HQ604854 | — |
| P. aurantiaca | CBS396.50 | France | MH856678 | MH868195 | [9] |
| P. bicornis | He3609 | China | MK588763 | MK588803 | Present study |
| P. bicornis | He4767 | China | MK588764 | MK588804 | Present study |
| P. borbonica | He4606 | China | MK588765 | MK588805 | Present study |
| P. borbonica | He4597 | China | MK588766 | MK588806 | Present study |
| P. cinerea | NH9808/1788 | Spain | AF506424 | AF506424 | [6] |
| P. cinerea | CBS261.37 | Belgium | MH855905 | MH867412 | [9] |
| P. cinerea | He3725 | China | MK588769 | MK588809 | Present study |
| P. crassitunicata | CBS663.91 | Reunion | MH862292 | MH873972 | [9] |
| P. crassitunicata | He3814 | China | MK588770 | MK588810 | Present study |
| P. cremicolor | He5380 * | China | MK588791 | MK588831 | Present study |
| P. duplex | TPDuB1022 | — | AF119519 | — | [29] |
| P. duplex | CBS286.58 | Canada | MH857787 | MH869321 | [9] |
| P. erikssonii | CBS287.58 | France | MH857788 | MH869322 | [9] |
| P. erikssonii | Cui11871 | China | MK588771 | MK588811 | Present study |
| P. exima | T-523 | USA | MK588772 | MK588812 | Present study |
| P. fasticata | CBS942.96 | Ethiopia | MH862624 | — | [9] |
| P. fissilis | CBS681.91 | Reunion | MH862298 | MH873975 | [9] |
| P. fissilis | CBS684.91 | Mascarene Islands | MH862299 | MH873976 | [9] |
| P. gilbertsonii | CBS357.95 | USA | MH862528 | MH874164 | [9] |
| P. gilbertsonii | CBS360.95 | USA | MH862530 | MH874165 | [9] |
| P. guadelupensis | CBS715.91 | Guadeloupe | MH862304 | MH873977 | [9] |
| P. halimi | CBS863.84 | France | MH861844 | MH873532 | [9] |
| P. halimi | CBS864.84 | France | MH861845 | MH873533 | [9] |
| P. incarnata | NH10271/1909 | Danmark | AF506425 | AF506425 | [6] |
| P. incarnata | CBS399.50 | France | MH856681 | MH868198 | [9] |
| P. junipericola | CBS349.54 | Sweden | MH857354 | — | [9] |
| P. junipericola | He2462 | China | MK588773 | MK588813 | Present study |
| P. kuehneri | CBS719.91 | Mascarene Islands | MH862307 | MH873980 | [9] |
| P. kuehneri | He4745 | China | MK588757 | MK588797 | Present study |
| P. kuehneroides | CBS731.91 | Mascarene Islands | MH862317 | MH873989 | [9] |
| P. kuehneroides | CBS732.91 | Mascarene Islands | MH862318 | MH873990 | [9] |
| P. laete | CBS256.56 | France | MH857617 | MH869165 | [9] |
| P. laete | FCUG 2681 | Russia | GU322869 | — | [30] |
| P. lassa | SP6129 | Russia | KJ509191 | — | [31] |
| P. lassa | He3052 | China | MK588758 | MK588798 | Present study |
| P. lassa | Dai17081A | China | MK588759 | MK588799 | Present study |
| P. laurentii | CBS325.73 | Norway | — | MH872397 | [9] |
| P. laxitexta | BAFC 3309 | Argentina | FJ882040 | — | [32] |
| P. laxitexta | LGMF1159 | Brazil | JX559580 | — | [33] |
| P. lilacea | CBS337.66 | Armenia | MH858813 | MH870452 | [9] |
| P. limitata | olrim963 | Lithuania | AY787678 | — | [34] |
| P. lycii | CBS264.56 | France | MH857624 | MH869169 | [9] |
| P. lycii | Boid-437 | France | MK588774 | MK588814 | Present study |
| P. major | He5528 * | China | MK588792 | MK588832 | Present study |
| P. malaiensis | CBS679.91 | Singapore | MH862297 | MH873974 | [9] |
| P. manshurica | He2956 | China | MK588776 | MK588816 | Present study |
| P. manshurica | He3729 | China | MK588777 | MK588817 | Present study |
| P. meridionalis | CBS289.58 | France | MH857789 | MH869323 | [9] |
| P. molesta | CBS677.91 | Gabon | MH862295 | — | [9] |
| P. molesta | CBS676.91 | Gabon | MH862294 | MH873973 | [9] |
| P. monticola | CBS649.91 | Reunion | MH862289 | MH873970 | [9] |
| P. nuda | He5280 | China | MK588778 | MK588818 | Present study |
| P. nuda | HHB-4916-Sp | USA | MK588779 | MK588819 | Present study |
| P. ovalispora | CBS653.91 | Mascarene Islands | MH862290 | MH873971 | [9] |
| P. parvocystidiata | CBS716.91 | Guadeloupe | MH862305 | MH873978 | [9] |
| P. piceae | olrim10 | Sweden | AY781264 | — | [35] |
| P. piceae | 209 | Russia | JX507718 | — | [36] |
| P. pilatiana | CBS265.56 | France | MH857625 | MH869170 | [9] |
| P. pilatiana | CBS-A1/A2 | — | MK588780 | MK588820 | Present study |
| P. pini | Hjm 18143 | Sweden | EU118651 | EU118651 | [7] |
| P. pini | CBS274.56 | France | MH857632 | MH869177 | [9] |
| P. pithya | CBS277.56 | France | MH857635 | MH869180 | [9] |
| P. pithya | He3107 | China | MK588781 | MK588821 | Present study |
| P. polygonia | CBS404.50 | France | MH856684 | MH868201 | [9] |
| P. polygonia | He4651 | China | MK588782 | MK588822 | Present study |
| P. proxima | CBS405.50 | France | MH856685 | MH868202 | [9] |
| P. proxima | He5498 | China | MK588783 | MK588823 | Present study |
| P. pseudopini | TPPpB1007 | — | AF119514 | — | [29] |
| P. pseudopini | DAOM-30124-Sp | Canada | MK588784 | MK588824 | Present study |
| P. pseudoversicolor | CBS338.66 | France | MH858814 | MH870453 | [9] |
| P. pseudoversicolor | He5132 | China | MK588785 | MK588825 | Present study |
| P. quercina | CBS407.50 | France | MH856687 | MH868204 | [9] |
| P. quercina | CBS408.50 | France | MH856688 | MH868205 | [9] |
| P. reidii | CBS397.83 | France | MH861616 | MH873334 | [9] |
| P. rhoica | CBS943.96 | Ethiopia | MH862625 | MH874246 | [9] |
| P. roseoalba | CLZhao3513 | China | ON786559 | OP380690 | [16] |
| P. roseoalba | CLZhao9401 | China | ON786560 | — | [16] |
| P. roseoalba | He5031 | China | OP872571 | OP872575 | Present study |
| P. rufa | CBS351.59 | Canada | MH857891 | MH869432 | [9] |
| P. rufa | He2788 | China | MK588786 | MK588826 | Present study |
| P. rufomarginata | CBS281.56 | France | MH857639 | MH869183 | [9] |
| P. rufomarginata | CBS282.56 | France | MH857640 | MH869184 | [9] |
| P. septentrionalis | CBS294.58 | Canada | MH857791 | MH869325 | [9] |
| P. shenghuae | He3507 * | China | MK588788 | MK588828 | Present study |
| P. shenghuae | He3535 | China | MK588789 | MK588829 | Present study |
| P. shenghuae | He5447 | China | MK588790 | MK588830 | Present study |
| P. simulans | CBS874.84 | France | MH861849 | MH873537 | [9] |
| P. simulans | CBS875.84 | France | MH861850 | MH873538 | [9] |
| P. sphaerocystidiata | HHB-8827-Sp | USA | MK588787 | MK588827 | Present study |
| P. subsalmonea | CBS696.91 | Mascarene Islands | MH862302 | — | [9] |
| P. subsalmonea | CBS697.91 | Mascarene Islands | MH862303 | — | [9] |
| P. taiwanensis | He4870 | China | MK588775 | MK588815 | Present study |
| P. taiwanensis | Wu 9206-28 | China | MK588793 | MK588833 | Present study |
| P. taiwanensis | Wu 9209-14 | China | MK588794 | MK588834 | Present study |
| P. tamaricicola | CBS438.62 | Morocco | MH858203 | MH869802 | [9] |
| P. tamaricicola | CBS439.62 | Morocco | MH858204 | MH869803 | [9] |
| P. trigonosperma | CBS402.83 | France | MH861618 | MH873335 | [9] |
| P. trigonosperma | He3602 | China | MK588762 | MK588802 | Present study |
| P. tristicula | CBS210.63 | Pakistan | MH858266 | — | [9] |
| P. tristicula | He4775 | China | MH669235 | MH669239 | [37] |
| P. versicolor | CBS358.61 | Morocco | MH858082 | MH869651 | [9] |
| P. versiformis | CBS358.54 | France | MH857360 | MH868902 | [9] |
| P. versiformis | He3029 | China | MK588756 | MK588796 | Present study |
| P. vietnamensis | He5242 | Vietnam | MK588760 | MK588800 | Present study |
| P. vietnamensis | He5252 * | Vietnam | MK588761 | MK588801 | Present study |
| P. violaceolivida | CBS348.52 | France | MH857077 | MH868613 | [9] |
| P. yunnanensi | CLZhao3978 | China | OP380617 | OP380689 | [16] |
| P. yunnanensi | CLZhao7347 | China | OP380616 | — | [16] |
| Scytinostroma jacksonii | NH6626/635 | Canada | AF506467 | AF506467 | [6] |
| S. portentosum | EL11-99 | Sweden | AF506470 | AF506470 | [6] |
| S. renisporum | CBS770.86 | Indonesia | MH862050 | MH873737 | [9] |
| Vararia investiens | TAA164122 | Norway | AF506484 | AF506484 | [6] |
| V. ochroleuca | JS24400 | Norway | AF506485 | AF506485 | [6] |
| Vesiculomyces citrinus | EL53-97 | Sweden | AF506486 | AF506486 | [6] |
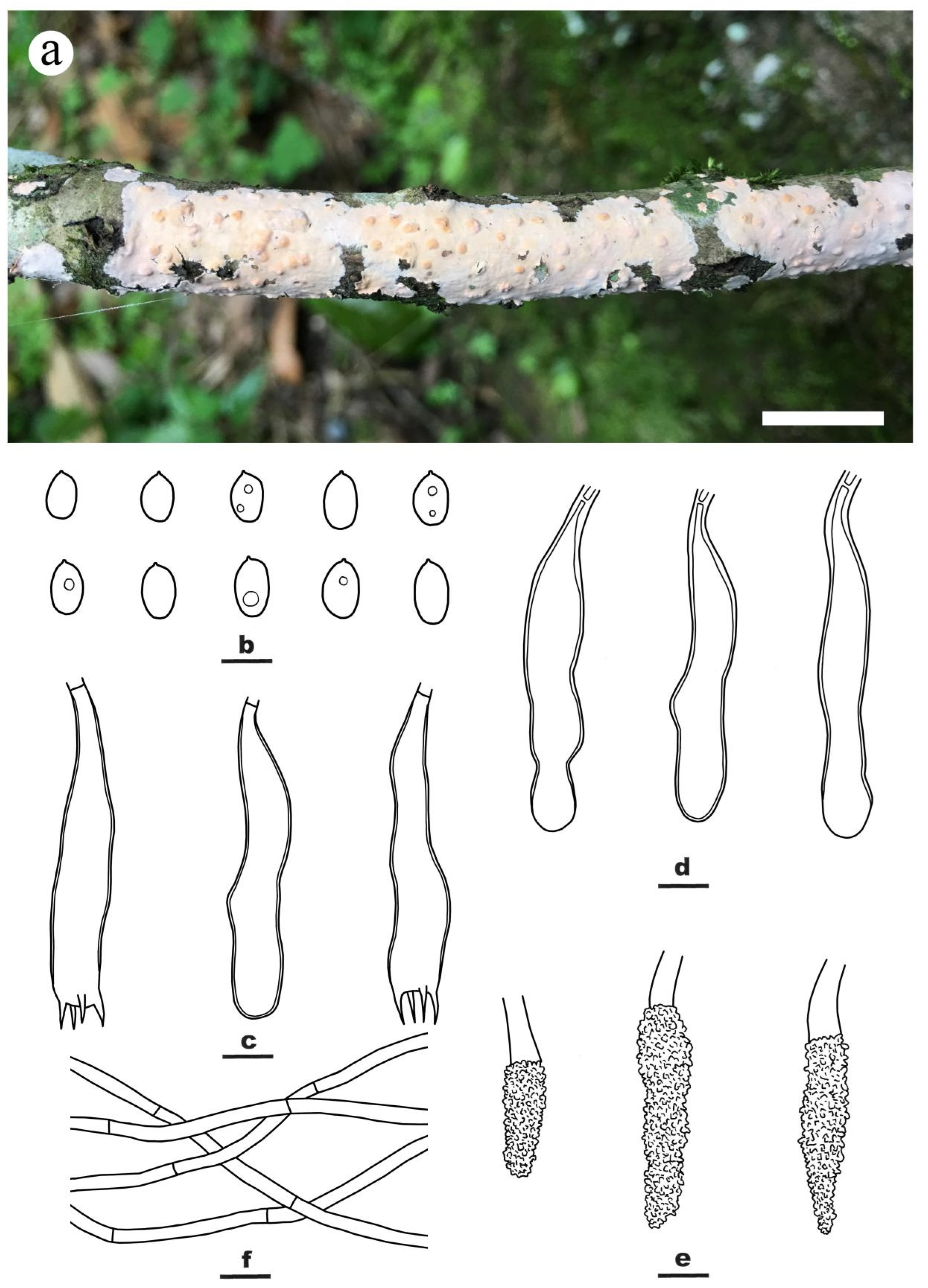
3.2. Taxonomy
| A key to Peniophora species in China | |
| 1. Dendrohyphidia well-developed, brown | P. versiformis |
| 1. Dendrohyphidia absent, or colorless when present | 2 |
| 2. Lamprocystidia brown over the entire length | 3 |
| 2. Lamprocystidia colorless or only brown at the basal part | 6 |
| 3. Skeletocystidia present | 4 |
| 3. Skeletocystidia absent | 5 |
| 4. Basidiospores 10–12.5 × 3.7–4.2 µm | P. tristicula |
| 4. Basidiospores 5.2–7.5 × 2–3.5 µm | P. kuehneri |
| 5. Basidiospores triangular, on palms | P. trignosperma |
| 5. Basidiospores cylindrical to narrowly ellipsoid or ovoid, on angiosperms | P. lassa |
| 6. Dendrohyphidia present, often colorless and may be difficult to see | 7 |
| 6. Dendrohyphidia absent | 8 |
| 7. Lamprocystidia absent; basidiospores > 7.5 µm long | P. polygonia |
| 7. Lamprocystidia present; basidiospores < 7.5 µm long | P. meridionalis |
| 8. Basidiospores ellipsoid or ovoid | 9 |
| 8. Basidiospores cylindrical or allantoid | 12 |
| 9. Basidiospores ovoid | P. roseoalba |
| 9. Basidiospores ellipsoid | 10 |
| 10. Generative hyphae with clamps | P. proxima |
| 10. Generative hyphae without clamps | 11 |
| 11. Basidiospores 9–13 × 6–7 µm | P. cremicolor |
| 11. Basidiospores 11–20 × 8–14 µm | P. erikssonii |
| 12. Basidiomes red | 13 |
| 12. Basidiomes grey, cream, pale orange, pinkish, yellowish or purplish | 14 |
| 13. Basidiomes pulvinate, only found on Populus | P. rufa |
| 13. Basidiomes effused, found on other hosts | P. pseudoversicolor |
| 14. Generative hyphae without clamps | 15 |
| 14. Generative hyphae with clamps | 19 |
| 15. Basidia with two sterigmata | P. bicornis |
| 15. Basidia with four sterigmata | 16 |
| 16. Basidiospores > 10 µm long | P. major |
| 16. Basidiospores < 10 µm long | 17 |
| 17. Basidiospores < 7 µm long | P. malaiensis |
| 17. Basidiospores > 7 µm long | 18 |
| 18. Basidiomes up to 1.2 mm thick; basidia thick-walled | P. borbonica |
| 18. Basidiomes up to 200 µm thick; basidia thin-walled | P. shenghuae |
| 19. Gloeocystidia indistinct or absent | 20 |
| 19. Gloeocystidia present | 22 |
| 20. Basidiomes pinkish, effused-reflexed, usually on Quercus. | P. manshurica |
| 20. Basidiomes greyish, resupinate, on gymnosperms or angiosperms | 21 |
| 21. Lamprocystidia 40–80 × 6–14 µm; on Juniperus | P. junipericola |
| 21. Lamprocystidia 15–25 × 5–10 µm; on gymnosperms or angiosperms | P. cinerea |
| 22. Gloeocystidia distinctly thick-walled, with walls up to 2–3 µm thick | P. crassitunicata |
| 22. Gloeocystidia thin- to slightly thick-walled, with walls < 1 µm thick | 23 |
| 23. Lamprocystidia up to 20 µm wide; basidiospores up to 7 µm long | P. pithya |
| 23. Lamprocystidia up to 12 µm wide; basidiospores up to 10 µm long | 24 |
| 24. Gloeocystidia 30–80 × 8–20 µm; basidiospores 2.5–3.5 µm wide | P. nuda |
| 24. Gloeocystidia 12.5–58 × 5.5–15.5 µm; basidiospores 3–5.5 µm wide | P. yunnanensis |
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eriksson, J. A taxonomical study with special reference to the Swedish species. Symb. Bot. Ups. 1950, 10, 1–76. [Google Scholar]
- Boidin, J.; Lanquetin, P. Peniophora (sect. Duportella) kuehneri et halimi novae sp.; réflexions sur les genres Peniophora et Duportella. Bull. Mens. Soc. Linn. Lyon 1974, 43, 47–60. [Google Scholar]
- Boidin, J.; Lanquetin, P.; Gilles, G. Les Peniophoraceae de la zone intertropicale (Basidiomycètes, Aphyllophorales). A. Espèces paléotropicales. Bull. Trimest. Société Mycol. Fr. 1991, 107, 91–156. [Google Scholar]
- Boidin, J.; Mugnier, J.; Canales, R. Taxonomie molecularies des Aphyllophorales. Mycotaxon 1998, 66, 445–491. [Google Scholar]
- Andreasen, M.; Hallenberg, N. A taxonomic survey of the Peniophoraceae. Synop. Fungorum 2009, 26, 56–119. [Google Scholar]
- Larsson, E.; Larsson, K.H. Phylogenetic relationships of russuloid basidiomycetes with emphasis on aphyllophoralean taxa. Mycologia 2003, 95, 1037–1065. [Google Scholar] [CrossRef]
- Larsson, K.H. Re-thinking the classification of corticioid fungi. Mycol. Res. 2007, 111, 1040–1063. [Google Scholar] [CrossRef]
- Miller, S.L.; Larsson, E.; Larsson, K.H.; Verbeken, A.; Nuytinck, J. Perspectives in the new Russulales. Mycologia 2006, 98, 960–970. [Google Scholar] [CrossRef] [PubMed]
- Vu, D.; Groenewald, M.; De Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 91, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.H. New records of the Corticiaceae from mainland China. Mycotaxon 2002, 82, 289–294. [Google Scholar]
- Wu, S.H. A study of Peniophora species with simple-septate hyphae occurring in Taiwan. Mycotaxon 2003, 85, 187–199. [Google Scholar]
- Wu, S.H. Twenty species of corticioid fungi newly recorded from China. Mycotaxon 2008, 104, 79–88. [Google Scholar]
- Wu, S.H. Survey of corticioid fungi in Taiwan, to 2010. Fungal Sci. 2010, 25, 49–60. [Google Scholar]
- Maekawa, N. Japanece Corticaceaea II. Rep. Tottori Mycol. Inst. 1994, 32, 1–123. [Google Scholar]
- Maekawa, N.; Yang, Z.L.; Zang, M. Corticioid Fungi (Basidiomycetes) collected in Sichuan Province, China. Mycotaxon 2002, 83, 81–95. [Google Scholar]
- Zou, L.; Zhang, X.; Deng, Y.; Zhao, C. Four New Wood-Inhabiting Fungal Species of Peniophoraceae (Russulales, Basidiomycota) from the Yunnan-Guizhou Plateau, China. J. Fungi 2022, 8, 1227. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Methuen: London, UK, 1978; p. 493. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. PCR Protocols: A guide to methods and applications. In Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.5.1. 2018. Available online: http://www.mesquiteproject.org (accessed on 20 October 2022).
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods); Version 4.0b10; Sinauer Associates: Sunderland, UK, 2002; Available online: http://paup.csit.fsu.edu/ (accessed on 25 October 2022).
- Stamatakis, A. RAxML Version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Binder, M.; Hibbett, D.S.; Larsson, K.H.; Larsson, E.; Langer, E.; Langer, G. The phylogenetic distribution of resupinate forms across the major clades of mushroom-forming fungi (Homobasidiomycetes). Syst. Biodivers. 2005, 3, 113–157. [Google Scholar] [CrossRef]
- Leal-Dutra, C.A.; Neves, M.A.; Griffith, G.W.; Reck, M.A.; Clasen, L.A.; Dentinger, B.T. Reclassification of Parapterulicium Corner (Pterulaceae, Agaricales), contributions to Lachnocladiaceae and Peniophoraceae (Russulales) and introduction of Baltazaria gen. nov. MycoKeys 2018, 37, 39–56. [Google Scholar] [CrossRef] [PubMed]
- Hsiau, P.T.; Harrington, T.C. Phylogenetics and adaptations of basidiomycetous fungi fed upon by bark beetles (Coleoptera: Scolytidae). Symbiosis 2003, 34, 111–131. [Google Scholar]
- Hallenberg, N.; Yurchenko, E.O.; Ghobad-Nejhad, M. Peniophora pseudonuda is a synonym of P. laeta. Mycotaxon 2010, 112, 153–162. [Google Scholar] [CrossRef]
- Spirin, V.; Kout, J. Duportella lassa sp. nov. from Northeast Asia. Mycotaxon 2015, 130, 483–488. [Google Scholar] [CrossRef]
- Robles, C.A.; Carmarán, C.C.; Lopez, S.E. Screening of xylophagous fungi associated with Platanus acerifolia in urban landscapes: Biodiversity and potential biodeterioration. Landsc. Urban Plan. 2011, 100, 129–135. [Google Scholar] [CrossRef]
- Hokama, Y.; Savi, D.C.; Assad, B.; Aluizio, R.; Gomes-Figueiredo, J.; Adamoski, D.; Glienke, C. Endophytic fungi isolated from Vochysia divergens in the Pantanal, Mato Grosso do Sul: Diversity, phylogeny and biocontrol of Phyllosticta citricarpa. In Endophytic Fungi: Divers. Charact. Biocontrol; Hughes, E., Ed.; Nova Publishers: New York, NY, USA, 2016; pp. 93–124. [Google Scholar]
- Lygis, V.; Vasiliauskas, R.; Larsson, K.H.; Stenlid, J. Wood-inhabiting fungi in stems of Fraxinus excelsior in declining ash stands of northern Lithuania, with particular reference to Armillaria cepistipes. Scand. J. For. Res. 2005, 20, 337–346. [Google Scholar] [CrossRef]
- Vasiliauskas, R.; Larsson, E.; Larsson, K.H.; Stenlid, J. Persistence and long-term impact of Rotstop biological control agent on mycodiversity in Picea abies stumps. Biol. Control. 2005, 32, 295–304. [Google Scholar] [CrossRef]
- Grum-Grzhimaylo, O.A.; Debets, A.J.; Bilanenko, E.N. The diversity of microfungi in peatlands originated from the White Sea. Mycologia 2016, 108, 233–254. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.L.; He, S.H. Taxonomy and phylogeny of Dichostereum (Russulales), with descriptions of three new species from southern China. MycoKeys 2018, 40, 111–126. [Google Scholar] [CrossRef]
- Boidin, J. Les Peniophoraceae des parties tempérées et froides de l’hémisphère nord (Basidiomycotina). Publ. Société Linnéenne Lyon 1994, 63, 317–334. [Google Scholar] [CrossRef]
- Hjortstam, K.; Ryvarden, L. Some new tropical genera and species of corticioid fungi (Basidiomycotina, Aphyllophorales). Synop. Fungorum 2004, 18, 20–32. [Google Scholar]
- Boidin, J.; Lanquetin, P. Sur quelques corticiés (Basidiomycotina) de l’Ethiopie. Cryptogamie Mycologie 1995, 16, 85–99. [Google Scholar]
- Zhao, C.L.; Cui, B.K.; Song, J.; Dai, Y.C. Fragiliporiaceae, a new family of Polyporales (Basidiomycota). Fungal Divers. 2015, 70, 115–126. [Google Scholar] [CrossRef]
- Chen, J.J.; Cui, B.K.; Dai, Y.C. Global diversity and molecular systematics of Wrightoporia s.l. (Russulales, Basidiomycota). Persoonia 2016, 37, 21–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, M.L.; Chen, Y.Y.; Shen, L.L.; Song, J.; Vlasák, J.; Dai, Y.C.; Cui, B.K. Taxonomy and phylogeny of the brown-rot fungi: Fomitopsis and its related genera. Fungal Divers. 2016, 80, 343–373. [Google Scholar] [CrossRef] [Green Version]
- Cui, B.K.; Li, H.J.; Ji, X.; Zhou, J.L.; Song, J.; Si, J.; Dai, Y.C. Species diversity, taxonomy and phylogeny of Polyporaceae (Basidiomycota) in China. Fungal Divers. 2019, 97, 137–302. [Google Scholar] [CrossRef]
- Wu, F.; Zhou, L.W.; Vlasák, J.; Dai, Y.C. Global diversity and systematics of Hymenochaetaceae with poroid hymenophore. Fungal Divers. 2022, 113, 1–192. [Google Scholar] [CrossRef]
- Chen, C.C.; Chen, C.Y.; Wu, S.H. Species diversity, taxonomy and multi-gene phylogeny of phlebioid clade (Phanerochaetaceae, Irpicaceae, Meruliaceae) of Polyporales. Fungal Divers. 2021, 111, 337–442. [Google Scholar] [CrossRef]
- He, S.H.; Dai, Y.C. Taxonomy and phylogeny of Hymenochaete and allied genera of Hymenochaetaceae (Basidiomycota) in China. Fungal Divers. 2012, 56, 77–93. [Google Scholar] [CrossRef]
- Li, Y.; Nakasone, K.K.; Chen, C.C.; Zhao, C.L.; Cao, T.; Yuan, H.S.; He, S.H. Taxonomy and phylogeny of Cystostereaceae (Agaricales, Basidiomycota): A new genus, five new species, and three new combinations. J. Fungi 2022, 8, 1229. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.L.; Nakasone, K.K.; Wu, S.H.; He, S.H.; Dai, Y.C. Taxonomy and phylogeny of Lopharia s.s., Dendrodontia, Dentocorticium and Fuscocerrena (Basidiomycota, Polyporales). MycoKeys 2018, 32, 25–48. [Google Scholar] [CrossRef] [Green Version]
- Riebeseh, J.; Yurchenko, E.; Nakasone, K.K.; Langer, E. Phylogenetic and morphological studies in Xylodon (Hymenochaetales, Basidiomycota) with the addition of four new species. MycoKeys 2019, 47, 97–137. [Google Scholar] [CrossRef]
- Yurchenko, E. The Genus Peniophora (Basidiomycota) of Eastern Europe: Morphology, Taxonomy, Ecology, Distribution; National Academy of Sciences of Belarus, VF Kuprevich Insitute of Experimental Botany: Minsk, Belarus, 2010. [Google Scholar]
- Dai, Y.C. A revised checklist of corticioid and hydnoid fungi in China for 2010. Mycoscience 2011, 52, 69–79. [Google Scholar] [CrossRef]
- Wang, H.; He, X.; Zhao, C.L. Scytinostroma yunnanense sp. nov. (Russulales, Basidiomycota) evidenced by morphological characteristics and phylogenetic analyses in China. Phytotaxa 2020, 451, 145–153. [Google Scholar] [CrossRef]
- Liu, S.L.; He, S.H. The genus Vararia (Russulales, Basidiomycota) in China. Two new species and two new Chinese records. Nord. J. Bot. 2016, 34, 553–558. [Google Scholar] [CrossRef]
- Liu, S.L.; Tian, Y.; Nie, T.; Thawthong, A.; Hyde, K.D.; Xu, L.L.; He, S.H. Updates on East Asian Asterostroma (Russulales, Basidiomycota): New species and new records from Thailand and China. Mycol. Prog. 2017, 16, 667–676. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.-L.; Tian, Y.; He, S.-H. Taxonomy and Phylogeny of Peniophora Sensu Lato (Russulales, Basidiomycota). J. Fungi 2023, 9, 93. https://doi.org/10.3390/jof9010093
Xu Y-L, Tian Y, He S-H. Taxonomy and Phylogeny of Peniophora Sensu Lato (Russulales, Basidiomycota). Journal of Fungi. 2023; 9(1):93. https://doi.org/10.3390/jof9010093
Chicago/Turabian StyleXu, Yun-Lin, Yan Tian, and Shuang-Hui He. 2023. "Taxonomy and Phylogeny of Peniophora Sensu Lato (Russulales, Basidiomycota)" Journal of Fungi 9, no. 1: 93. https://doi.org/10.3390/jof9010093