
Morphology and Phylogeny of Lyophylloid Mushrooms in China with Description of Four New Species
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimen Sampling
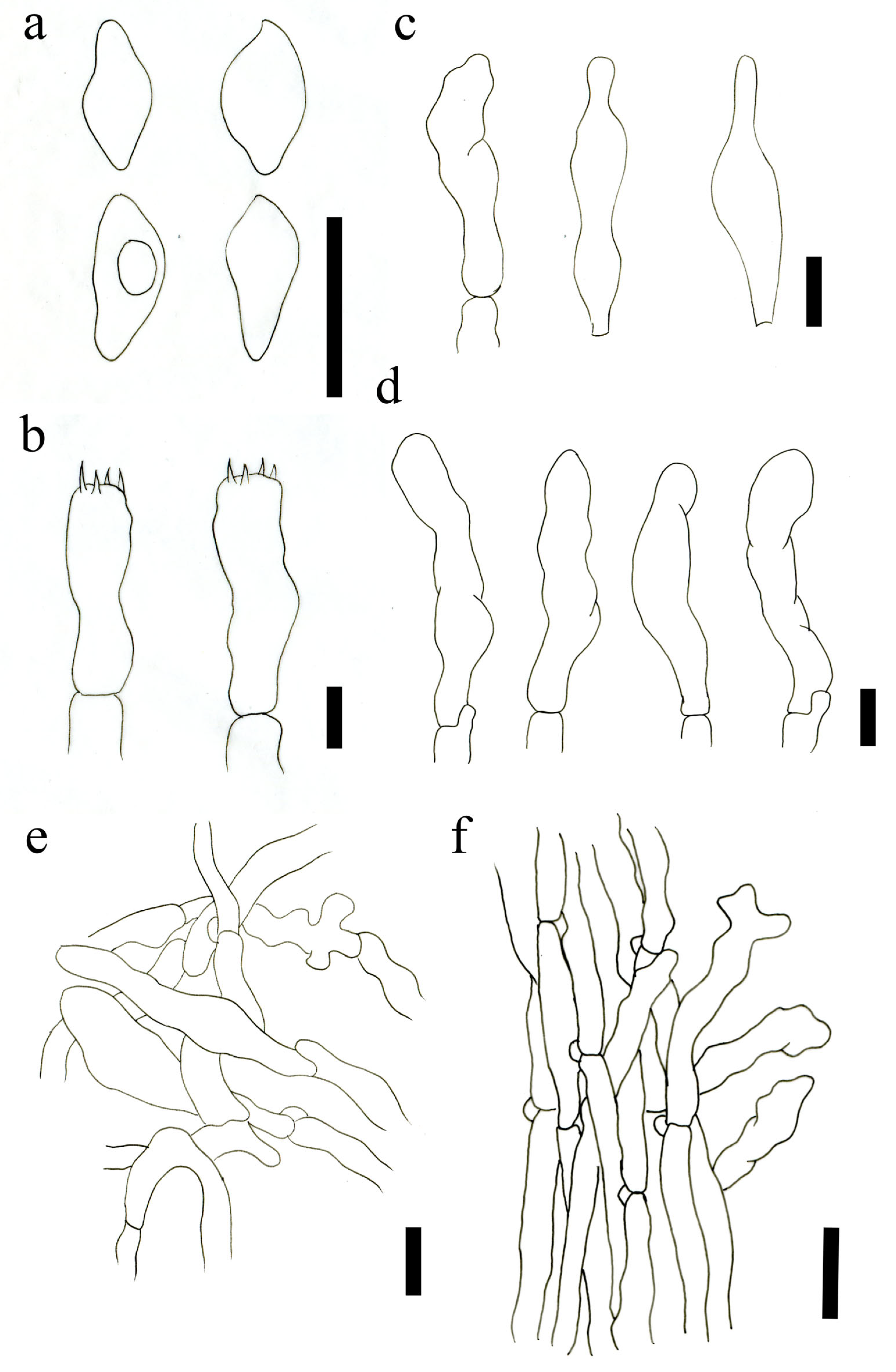
2.2. Morphological Observation
2.3. DNA Extraction, Amplification, and Sequencing
2.4. Phylogenetic Analyses
3. Results
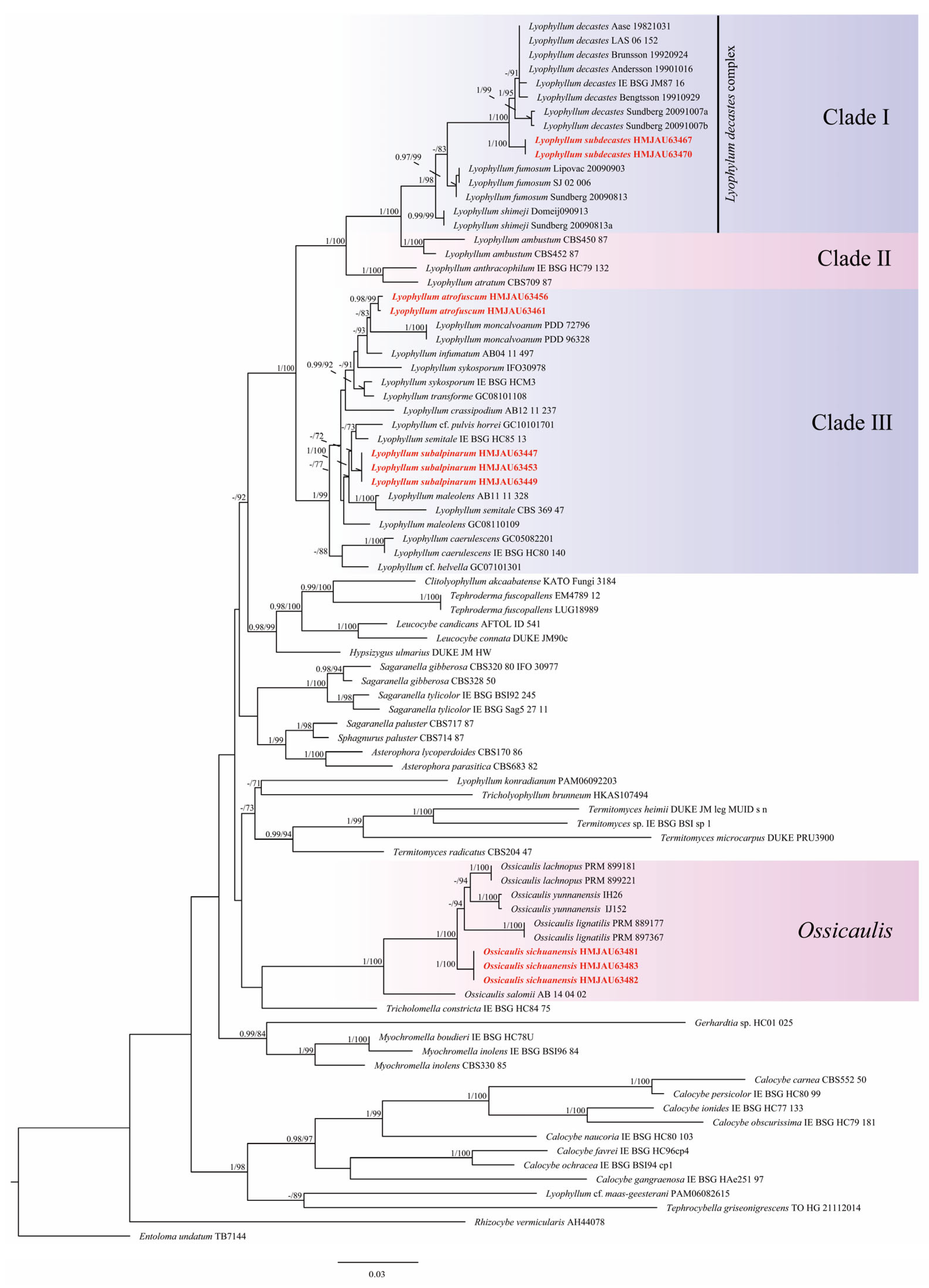
3.1. Molecular Phylogeny
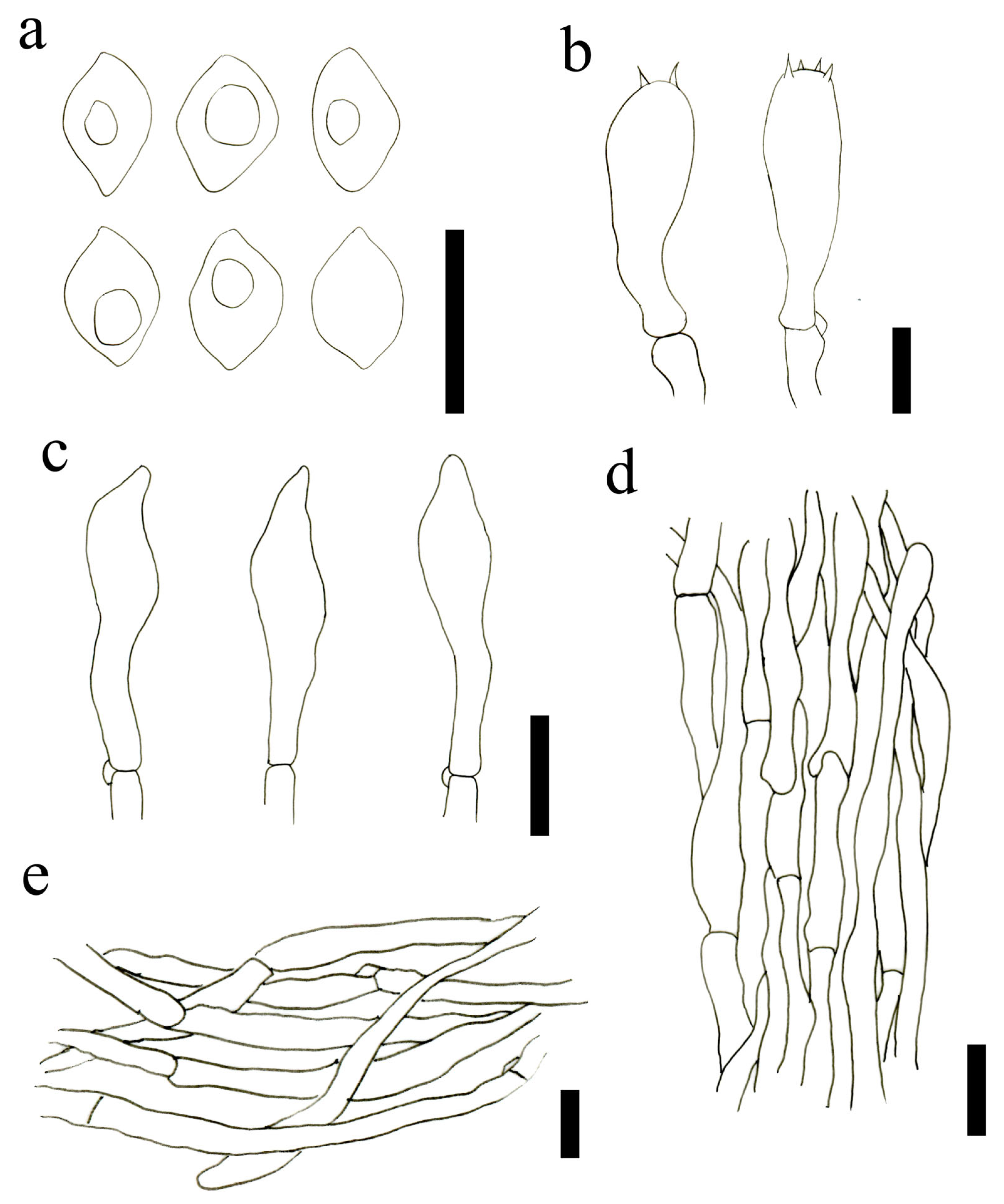
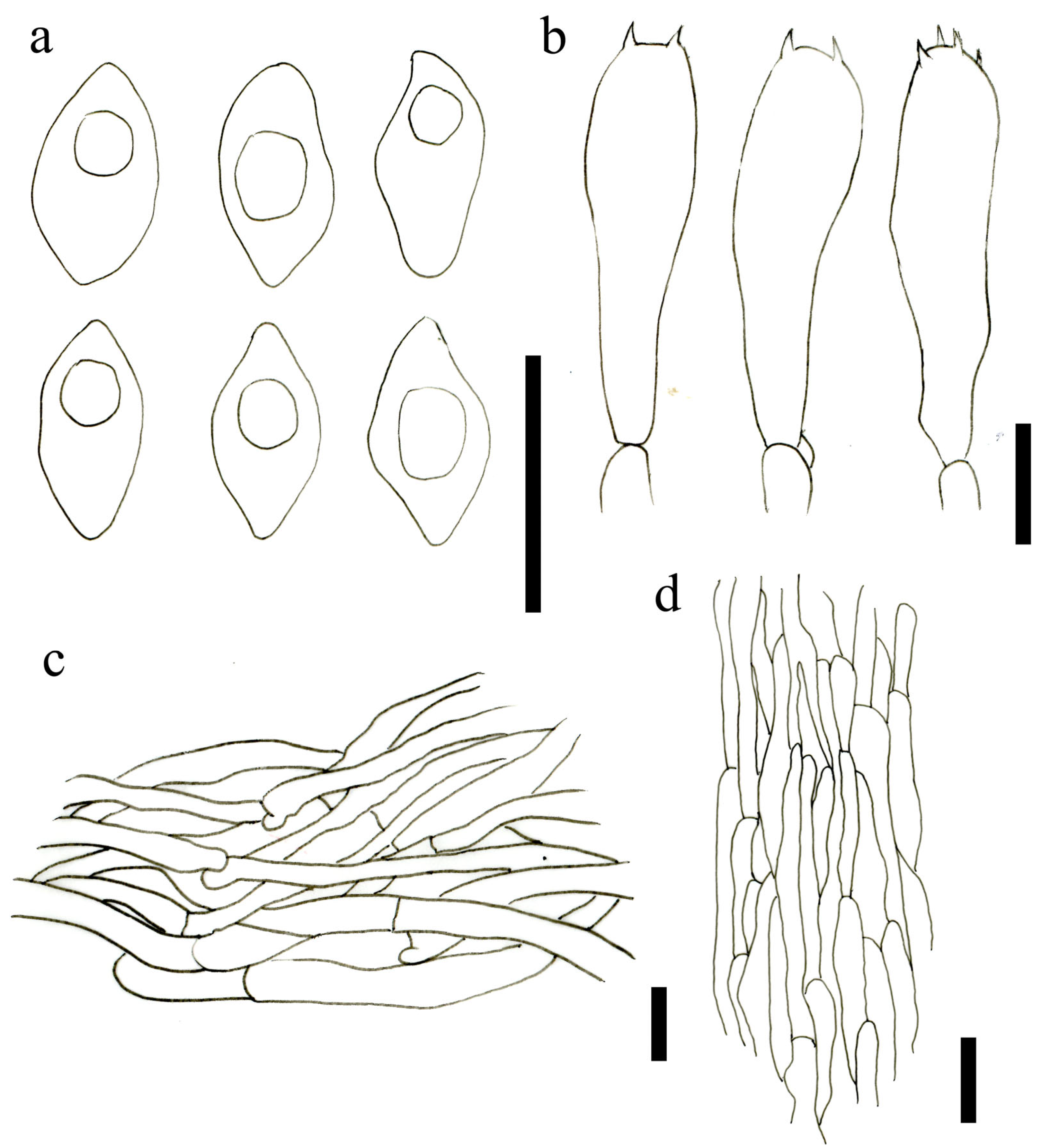
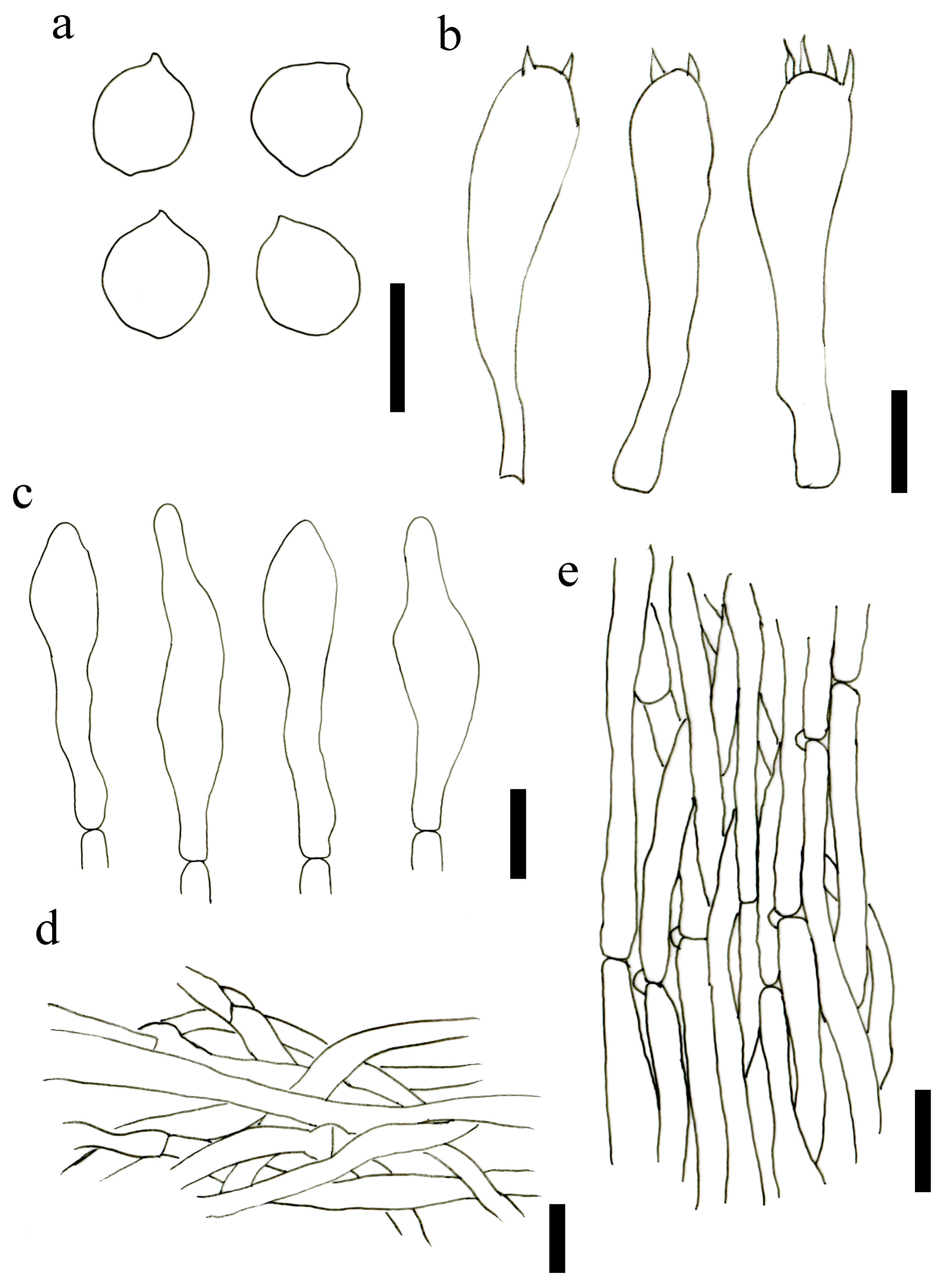
3.2. Taxonomy
4. Discussion
| Key to species of Lyophyllum in China | |
| 1. Lamellae black when bruised | 2 |
| 1’. Lamellae no staining when bruised | 13 |
| 2. Basidiocarps small, pileus usually less than 6 cm, stipe less than 5 cm, and width narrower than 1.2 cm | 3 |
| 2’. Basidiocarps medium to large, pileus usually more than 6 cm, stipe more than 5 cm long, and width exceeding 1.2 cm | 7 |
| 3. Lamellae dark, grayish-black | L. trigonosporum |
| 3’. Lamellae white, gray to light brown | 4 |
| 4. Pileus usually less than 3 cm | 5 |
| 4’. Pileus usually broader than 3 cm | 6 |
| 5. Width of the stipe is narrower than 0.3 cm | L. pusillum |
| 5’. Width of the stipe is usually 0.3–1.2 cm | L. pulvis-horrei |
| 6. Pileipellis hyphae width of more than 4 μm | L. subalpinarum |
| 6’. Pileipellis hyphae width less than or equal to 4 μm | L. semitale |
| 7. Basidiospores usually longer than 14 μm | L. rhombisporum |
| 7’. Basidiospores shorter than 14 µm | 8 |
| 8. Basidiospores globosae | L. immundum |
| 8’. Basidiospores not globosae | 9 |
| 9. Basidiospores triangular | 10 |
| 9’. Basidiospores rhombic or subrhombic | 11 |
| 10. Basidiospores with a hump located in the middle of the abaxial side, not thicker than the main body of the basidiospore and not higher than the length of the Basidiospore | L. sykosporum |
| 10’. Basidiospores with a broad abaxial, thickening and making it widest near the apex | L. transforme |
| 11. Q value less than or equal to 1.5 | L. atrofuscum |
| 11’. Q value higher than 1.5 | 12 |
| 12. Hymenophoral trama are regular, hyphae of the mediostratum are narrow than 15 μm | L. infumatum |
| 12’. Hymenophoral trama are regular, hyphae of the mediostratum exceed 15 μm, up to 20 μm | L. macrosporum |
| 13. Basidiocarps large, up to exceeding 10 cm | 14 |
| 13’. Basidiocarps are small to medium, less than 10 cm | 15 |
| 14. Pileus reddish-brown to chestnut-brown, the width of stipe usually less than 1.5 cm | L. loricatum |
| 14’. Pileus grayish-brown to grayish-yellow, the width of the stipe usually exceeds more than 1.5cm | L. shimeji |
| 15. Basidiospores diam usually less than 5.5 μm | L. subdecastes |
| 15’. Basidiospores diam usually more than 5.5 μm | 16 |
| 16. Basidia usually less than 30 μm | L. fumosum |
| 16’. Basidia usually longer than 30 μm | L. decastes |
| Key to worldwide species of Ossicaulis | |
| 1. Basidiospores usually shorter than 4 µm | 2 |
| 1’. Basidiospores usually longer than 4 μm | 3 |
| 2. Pileus white, chalky to orange tinged, without grayish tinge, on living tree trunks in alpine belt | O. yunnanensis |
| 2’. Pileus gray or beige-gray, on dying or decaying woods | O. lachnopus |
| 3. Width of the stipe is narrower than 2.0 cm | O. salomii |
| 3’. Width of stipe is usually more than 2.0 cm | 4 |
| 4. Basidia usually less than 15 μm | O. lignatilis |
| 4’. Basidia usually longer than 15 μm | O. sichuanensis |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hofstetter, V.; Redhead, S.A.; Kauff, F.; Moncalvo, J.M.; Matheny, P.B.; Vilgalys, R. Taxonomic Revision and Examination of Ecological Transitions of the Lyophyllaceae (Basidiomycota, Agaricales) Based on a Multigene Phylogeny. Cryptogam. Mycol. 2014, 35, 399–425. [Google Scholar] [CrossRef]
- Alvarado, P.; Moreau, P.A.; Dima, B.; Vizzini, A.; Consiglio, G. Pseudoclitocybaceae fam. nov.(Agaricales, Tricholomatineae), a new arrangement at family, genus and species level. Fungal Divers. 2018, 90, 109–133. [Google Scholar] [CrossRef]
- Bellanger, J.M.; Moreau, P.A.; Corriol, G.; Bidaud, A.; Chalange, R.; Dudova, Z.; Richard, F. Plunging hands into the mushroom jar: A phylogenetic framework for Lyophyllaceae (Agaricales, Basidiomycota). Genetica 2015, 143, 169–194. [Google Scholar] [CrossRef] [PubMed]
- Lennart, J.J.; Marry, C.A.; Thomas, L.; Ole, S.P.; Victor, R.M.; Thomas, W.K.; Dirk, S.; Duur, K.A.; Timothy, J.B. Four new genera and six new species of lyophylloid agarics (Agaricales, Basidiomycota) from three different continents. Mycol. Prog. 2022, 21, 1–14. [Google Scholar]
- Vizzini, A.; Consiglio, G.; Setti, L.; Ercole, E. Calocybella, a new genus for Rugosomyces pudicus (Agaricales, Lyophyllaceae) and emendation of the genus Gerhardtia. IMA Fungus 2015, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.L.; Wu, G.; Li, Y.C.; Wang, X.H.; Cai, Q. Common Edible and Poisonous Mushrooms of Southwestern China; Science Press: Beijing, China, 2021. [Google Scholar]
- Zhang, F.; Xu, H.; Yuan, Y.; Huang, H.; Wu, X.; Zhang, J.; Fu, J. Lyophyllum decastes fruiting body polysaccharide alleviates acute liver injury by activating the Nrf2 signaling pathway. Food Funct. 2022, 13, 2057–2067. [Google Scholar] [PubMed]
- Wu, F.; Zhou, L.W.; Yang, Z.L.; Bau, T.; Li, T.H.; Dai, Y.C. Resource diversity of Chinese macrofungi: Edible, medicinal and poisonous species. Fungal Divers. 2019, 98, 1–76. [Google Scholar] [CrossRef]
- Zheng, Z.Y.; Han, J.J.; Mi, P.C.; Chen, B.S.; Bao, L.; Xi, Y.L.; Wang, Z.D.; Xie, L.P.; Guo, C.B.; Wei, S.L. Chemical constituents from fruiting bodies of Lyophyllum decastes. Mycosystema 2020, 39, 1774–1782. [Google Scholar]
- Li, J.H.; Yang, B.; Ren, J.Y.; Jin, W.S.; Sun, S.J.; Hu, K.H. Effects of hot air drying temperature on drying characteristics and volatile flavor compounds for Lyophyllum decastes. Mycosystema 2021, 40, 3304–3319. [Google Scholar]
- Xu, J.; Yu, X.; Suwannarach, N.; Jiang, Y.; Zhao, W.; Li, Y. Additions to Lyophyllaceae s.l. from China. J. Fungi 2021, 7, 1101. [Google Scholar] [CrossRef]
- Joshi, M.; Bhargava, P.; Bhatt, M.; Kadri, S.; Shri, M.; Joshi, C.G. Lyophyllaceae. In Mushrooms of Gujarat; Springer: Singapore, 2021; pp. 31–40. [Google Scholar]
- Vesterholt, J. Funga Nordica, Agaricoid, Boletoid, Cyphelloid and Gasteroid Genera; Nordsvamp: Copenhagen, Denmark, 2012; p. 9811021. [Google Scholar]
- He, M.Q.; Zhao, R.L.; Hyde, K.D.; Begerow, D.; Kemler, M.; Yurkov, A.; McKenzie, E.H.; Raspé, O.; Kakishima, M.; Sánchez-Ramírez, S. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar]
- Kühner, R. Utilisation du carmin acétique dans la classification des agarics leucosporés. Publ. De La Société Linnéenne De Lyon 1938, 7, 204–211. [Google Scholar] [CrossRef]
- Singer, R. The Agaricales in Modern Taxonomy, 4th ed.; Koeltz Scientific Books: Koenigstein, Germany, 1986; p. 981. [Google Scholar]
- Sundberg, H. Phylogeny of Lyophyllum Section Difformia. Does hon-shimeji (L. shimeji) Occur in Sweden? Master’s Thesis, Department of Plant and Environmental Sciences, University of Gothenburg, Gothenburg, Sweden, 2010; pp. 1–19. [Google Scholar]
- Sesli, E.; Vizzini, A.; Contu, M. Lyophyllum turcicum (Agaricomycetes: Lyophyllaceae), a new species from Turkey. Turk. J. Bot. 2015, 39, 512–519. [Google Scholar] [CrossRef]
- Redhead, S.A.; Hofstetter, V.; Clémençon, H.; Moncalvo, J.M.; Vilgalys, R.J. (1742) Proposal to conserve the name Lyophyllum with a conserved type (Basidiomycota). Taxon 2006, 55, 1034–1036. [Google Scholar] [CrossRef]
- Redhead, S.A. A reappraisal of agaric genera associated with brown rots of woods. Trans. Mycol. Soc. Jpn. 1985, 26, 349–389. [Google Scholar]
- Holec, J.; Kolařík, M. Ossicaulis lachnopus (Agaricales, Lyophyllaceae), a species similar to O. lignatilis, is verified by morphological and molecular methods. Mycol. Prog. 2013, 12, 589–597. [Google Scholar] [CrossRef]
- Ye, L.; Karunarathna, S.C.; Li, H.; Xu, J.; Hyde, K.D.; Mortimer, P.E. A survey of Termitomyces (Lyophyllaceae, Agaricales), including a new species, from a subtropical forest in Xishuangbanna, China. Mycobiology 2019, 47, 391–400. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Li, T.H.; Wang, C.Q.; Deng, W.Q.; Song, B. Gerhardtia sinensis (Agaricales, Lyophyllaceae), a new species and a newly recorded genus for China. Phytotaxa 2017, 332, 172–180. [Google Scholar] [CrossRef]
- Cai, Q.; LvLi, Y.; Kost, G.; Yang, Z.L. Tricholyophyllum brunneum gen. et. sp. nov. with bacilliform basidiospores in the family Lyophyllaceae. Mycosystema 2020, 39, 1728–1740. [Google Scholar]
- Feng, Y.; Tang, X.; Yu, J.F.; Yang, Z.F.; Ma, M.; Guo, X. Recent Research Progress of Lyophyllum. Vegetables 2019, 8, 64–69. [Google Scholar]
- Li, X.; Zhang, S.Y.; Li, Y. Biological Characteristics of Lyophyllum fumosum. Edible Fungi China 2010, 29, 20–22. [Google Scholar]
- Zhang, Y.; Mo, M.; Yang, L.; Mi, F.; Cao, Y.; Liu, C.L.; Tang, X.Z.; Wang, P.F.; Xu, J.P. Exploring the species diversity of edible mushrooms in Yunnan, Southwestern China, by DNA barcoding. J. Fungi 2021, 7, 310. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Q.; Zhou, D.Q.; Zhang, X.L. Taxonomic Study on the Genus Lyophyllum in Sustainable Environment Engineering. Appl. Mech. Mater. 2013, 340, 975–978. [Google Scholar] [CrossRef]
- Yang, S.D.; Huang, H.Y.; Zhao, J.; Zeng, N.K.; Tang, L.P. Ossicaulis yunnanensis sp. nov.(Lyophyllaceae, Agaricales) from southwestern China. Mycoscience 2018, 59, 33–37. [Google Scholar] [CrossRef]
- Li, Y.; Li, T.H.; Yang, Z.L.; Bau, T.; Dai, Y.C. Atlas of Chinese Macrofungal Resources; Central Chinese Farmer: Zhengzhou, China, 2015. [Google Scholar]
- Komerup, A.; Wanscher, J. Methuen Handbook of Colour, 3rd ed.; Pavey, D., Ed.; Eyre Methuen: London, UK, 1978; pp. 1–252. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Bolchacova, E.; Voigt, K.; Crous, P.W. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [Green Version]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. Bioedit: A user–friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Hofstetter, V.; Clémençon, H.; Vilgalys, R.; Moncalvo, J.M. Phylogenetic analyses of the Lyophylleae (Agaricales, Basidiomycota) based on nuclear and mitochondrial rDNA sequences. Mycol. Res. 2002, 106, 1043–1059. [Google Scholar] [CrossRef]
- Sesli, E.; Vizzini, A.; Ercole, E.; Contu, M. Clitolyophyllum akcaabatense gen. nov., sp. nov.(Agaricales, Tricholomatineae); a new fan-shaped clitocyboid agaric from Turkey. Botany 2016, 94, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Larsson, E.; Sundberg, H. Lyophyllum shimeji, a species associated with lichen pine forest in northern Fennoscandia. Mycoscience 2011, 52, 289–295. [Google Scholar]
- Cooper, J. New species and combinations of some New Zealand agarics belonging to Clitopilus, Lyophyllum, Gerhardtia, Clitocybe, Hydnangium, Mycena, Rhodocollybia and Gerronema. Mycosphere 2014, 5, 263–288. [Google Scholar] [CrossRef]
- Crous, P.W.; Carnegie, A.J.; Wingfield, M.J.; Sharma, R.; Mughini, G.; Noordeloos, M.E.; Santini, A.; Shouche, Y.S.; Bezerra, J.D.; Dima, B. Fungal Planet description sheets: 868–950. Pers. Mol. Phylogeny Evol. Fungi 2019, 42, 291. [Google Scholar] [CrossRef]
- Alvarado, P.; Moreno, G.; Vizzini, A.; Consiglio, G.; Manjon, J.L.; Setti, L. Atractosporocybe, Leucocybe and Rhizocybe: Three new clitocyboid genera in the Tricholomatoid clade (Agaricales) with notes on Clitocybe and Lepista. Mycologia 2015, 107, 123–136. [Google Scholar] [CrossRef] [Green Version]
- Musumeci, E.; Contu, M. Tephroderma (Agaricomycetidae, Tricholomatoid clade), un nuovo genere di Basidiomiceti lamellati dalla Francia. Bolletino AMER 2014, 91, 20–30. [Google Scholar]
- Dähncke, R.; Contu, M.; Vizzini, A. Some rare or critical taxa of the genus Lyophyllum s.l. Basidiomycota, Agaricomycetes from La Palma Canary Islands, Spain. Osterr. Z. Für Pilzkd. 2009, 18, 129–139. [Google Scholar]
- Dähncke, R.M.; Contu, M.; Vizzini, A. Two new species of Lyophyllum s.l. (Basidiomycota, Agaricomycetes) from La Palma (Canary Islands, Spain). Mycotaxon 2011, 111, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Clémençon, H.; Smith, A. New species of Lyophyllum (Agaricales) from North America and a key to the known staining species. Mycotaxon 1983, 18, 379–437. [Google Scholar]
- Wang, X.Q.; Zhou, D.Q.; Zhao, Y.C.; Zhang, X.L.; Li, L.; Li, S.H. Lyophyllum rhombisporum sp. nov. from China. Mycotaxon 2013, 123, 473–477. [Google Scholar] [CrossRef]
- Breitenbach, J. Fungi of Switzerland. Volume 3, Boletes and Agarics 1; Verlag Mycologia: Luzern, Switzerland, 1991. [Google Scholar]
- Hesler, L.R. Lyophyllum Notebook 1; University of Tennessee: Knoxville, TN, USA, 2013. [Google Scholar]
- Kim, Y.S.; Park, Y.H.; Shin, K.C. Notes on Korean Agaricales (II). Korean J. Mycol. 1978, 6, 37–42. [Google Scholar]
- Arana-Gabriel, Y.; Burrola-Aguilar, C.; Garibay-Orijel, R.; Matías-Ferrer, N.; Franco-Maass, S.; Mata, G. Genetic characterization, evaluation of growth and production of biomass of strains from wild edible mushrooms of Lyophyllum of Central Mexico. Braz. J. Microbiol. 2018, 49, 632–640. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Q.; Jin, X.; Chen, Z.; Xiong, C.; Li, P.; Zhao, J.; Huang, W. Characterization and comparison of the mitochondrial genomes from two Lyophyllum fungal species and insights into phylogeny of Agaricomycetes. Int. J. Biol. Macromol. 2019, 121, 364–372. [Google Scholar] [CrossRef]
- Bocian, M. Nowe stanowisko kępkowca jasnobrązowego Lyophyllum decastes (Fr.) Singer na Pomorzu Zachodnim. Przegląd Przyr. 2017, 28, 32–38. [Google Scholar]
- Ohta, A. Some cultural characteristics of mycelia of a mycorrhizal fungus, Lyophyllum shimeji. Mycoscience 1994, 35, 83–87. [Google Scholar] [CrossRef]
- Gillet, C.C. Les Hyménomycètes: Ou, Description de Tous Les Champignons (Fungi) qui Croissent en France, Avec l’indication de Leurs Propriétés Utiles ou Vénéneuses; C. Thomas: Alençon, France, 1874; Volume 1. [Google Scholar]
- Moncalvo, J.M.; Clémençon, H. A comparative study of fruit body morphology and culture characters in the Lyophyllum decastes complex (Agaricales, Basidiomycetes) from Japan and Europe. Trans. Mycol. Soc. Jpn. 1992, 33, 3–11. [Google Scholar]
- Moncalvo, J.M.; Clémençon, H. Enzymic studies as an aid to the taxonomy of the Lyophyllum decastes complex. Mycol. Res. 1994, 98, 375–383. [Google Scholar] [CrossRef]
- Moncalvo, J.M.; Rehner, S.A.; Vilgalys, R. Systematics of Lyophyllum section Difformia based on evidence from culture studies and ribosomal DNA sequences. Mycologia 1993, 85, 788–794. [Google Scholar] [CrossRef]
- Deng, W.Y.; Li, G.W.; Ge, Z.W.; Zhang, P. Two new species of Clavariadelphus from alpine temperate forests of southwestern China. Mycosystema 2020, 39, 1684–1693. [Google Scholar]
- Wu, F.; Zhao, Q.; Yang, Z.L.; Ye, S.Y.; Rivoire, B.; Dai, Y.C. Exidia yadongensis, a new edible species from East Asia. Mycosystema 2020, 39, 1203–1214. [Google Scholar]
- Wu, F.; Wang, X.H.; Qin, W.Q.; Hao, Y.J.; Guo., T.; Zhang, P.; Chen, Z.H. Five new species and one new record of Lactarius (Russulaceae, Russulales) from tropical and subtropical regions of China. Mycosystema 2022, 41, 1234–1253. [Google Scholar]
- Dai, Y.C.; Yang, Z.L.; Cui, B.K.; Wu, G.; Yuan, H.S. Diversity and systematics of the important macrofungi in Chinese forests. Mycosystema 2021, 40, 770–805. [Google Scholar]
- Guan, Q.X.; Zhao, C.L. Two new corticioid species, Hyphoderma sinense and H. floccosum (Hyphodermataceae, Polyporales), from southern China. Mycosystema 2021, 40, 447–461. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Voucher ID | GenBank Accession No. | References | |
|---|---|---|---|---|
| ITS | nLSU | |||
| Asterophora lycoperdoides | CBS170.86 | AF357037 | AF223190 | [1] |
| A. parasitica | CBS683.82 | AF357038 | AF223191 | [1] |
| Calocybe carnea | CBS552.50 | AF357028 | AF223178 | [1] |
| C. favrei | IE-BSG-HC96cp4 | EF421102 | AF223184 | [1] |
| C. gangraenosa | IE-BSG-HAe251.97 | AF357032 | AF223202 | [1] |
| C. ionides | IE-BSG-HC77/133 | AF357029 | AF223179 | [1] |
| C. naucoria | IE-BSG-HC80/103 | AF357030 | AF223180 | [1] |
| C. obscurissima | IE-BSG-HC79/181 | AF357031 | AF223181 | [1] |
| C. ochracea | IE-BSG-BSI94.cp1 | AF357033 | AF223185 | [41] |
| C. persicolor | IE-BSG-HC80/99 | AF357026 | AF223176 | [1] |
| Clitolyophyllum akcaabatense | KATO Fungi 3184 | KT934393 | KT934394 | [42] |
| Entoloma undatum | TB7144 | EF421108 | AF261315 | [1] |
| Gerhardtia sp. | HC01/025 | EF421103 | EF421091 | [1] |
| Hypsizygus ulmarius | DUKE-JM/HW | EF421105 | AF042584 | [1] |
| Leucocybe candicans | AFTOL-ID 541 | DQ202268 | AY645055 | [1] |
| L. connata | DUKE-JM90c | EF421104 | AF042590 | [1] |
| Lyophyllum ambustum | CBS452.87 | AF357057 | AF223216 | [1] |
| L. ambustum | CBS450.87 | AF357058 | AF223214 | [1] |
| L. anthracophilum | IE-BSG-HC79/132 | AF357055 | AF223212 | [1] |
| L. atratum | CBS709.87 | AF357053 | AF223210 | [1] |
| L. atrofuscum | HMJAU63461 | OP605493 | OP605513 | this study |
| L. atrofuscum | HMJAU63456 | OP605494 | OP605514 | this study |
| L. caerulescens | GC05082201 | KP192628 | NA | [3] |
| L. caerulescens | IE-BSG-HC80/140 | AF357052 | AF223209 | [1] |
| L. cf. helvella | GC07101301 | KP192625 | NA | [3] |
| L. cf. maas-geesterani | PAM06082615 | KP192553 | NA | [3] |
| L. cf. pulvis-horrei | GC10101701 | KP192665 | NA | [3] |
| L. crassipodium | AB12-11-237 | KP192608 | NA | [3] |
| L. decastes | IE-BSG-JM87/16 | AF357059 | AF042583 | [1] |
| L. decastes | Brunsson 19920924 | HM572544 | NA | [43] |
| L. decastes | Andersson 19901016 | HM572546 | NA | [43] |
| L. decastes | Bengtsson 19910929 | HM572545 | NA | [43] |
| L. decastes | LAS 06-152 | HM572547 | NA | [43] |
| L. decastes | Aase 19821031 | HM572543 | NA | [43] |
| L. decastes | Sundberg 20091007a | HM572548 | NA | [43] |
| L. decastes | Sundberg 20091007b | HM572549 | NA | [43] |
| L. fumosum | SJ 02-006 | HM572539 | NA | [43] |
| L. fumosum | Lipovac 20090903 | HM572538 | NA | [43] |
| L. fumosum | Sundberg 20090813 | HM572537 | NA | [43] |
| L. infumatum | AB04-11-497 | KP192584 | NA | [3] |
| L. konradianum | PAM06092203 | KP192569 | NA | [3] |
| L. maleolens | AB11-11-328 | KP192607 | NA | [3] |
| L. maleolens | GC08110109 | KP192624 | NA | [3] |
| L. moncalvoanum | PDD 72796 | KJ461890 | KJ461891 | [44] |
| L. moncalvoanum | PDD 96328 | KJ461904 | KJ461905 | [44] |
| L. semitale | CBS 369.47 | AF357048 | AF223207 | [41] |
| L. semitale | IE-BSG-HC85/13 | AF357049 | AF042581 | [1] |
| L. shimeji | Domeij090913 | HM572525 | NA | [43] |
| L. shimeji | Sundberg 20090813a | HM572524 | NA | [43] |
| L. subalpinarum | HMJAU63449 | OP605490 | OP605510 | this study |
| L. subalpinarum | HMJAU63453 | OP605491 | OP605511 | this study |
| L. subalpinarum | HMJAU63447 | OP605492 | OP605512 | this study |
| L. subdecastes | HMJAU63470 | OP605488 | OP605508 | this study |
| L. subdecastes | HMJAU63467 | OP605489 | OP605509 | this study |
| L. sykosporum | IE-BSG-HCM3 | AF357051 | AF357073 | [41] |
| L. sykosporum | IFO30978 | AF357050 | AF223208 | [1] |
| L. transforme | GC08101108 | KP192653 | NA | [3] |
| Myochromella boudieri | IE-BSG-HC78U | AF357046 | AF223206 | [41] |
| M. inolens | IE-BSG-BSI96/84 | AF357047 | AF223204 | [1] |
| M. inolens | CBS330.85 | AF357045 | AF223201 | [1] |
| Ossicaulis lachnopus | PRM 899221 | HE649956 | NA | [21] |
| O. lachnopus | PRM 899181 | HE649955 | NA | [21] |
| O. lignatilis | PRM 897367 | HE649952 | NA | [21] |
| O. lignatilis | PRM 889177 | HE649953 | NA | [21] |
| O. salomii | AB 14-04-02 | MK650044 | MK650043 | [45] |
| O. sichuanensis | HMJAU63481 | OP605495 | OP605515 | this study |
| O. sichuanensis | HMJAU63482 | OP605496 | OP605516 | this study |
| O. sichuanensis | HMJAU63483 | OP605497 | OP605517 | this study |
| O. yunnanensis | IJ152 | KY411962 | KY411960 | [29] |
| O. yunnanensis | IH26 | KY411961 | KY411959 | [29] |
| Rhizocybe vermicularis | AH44078 | KJ681032 | KJ681039 | [46] |
| Sagaranella gibberosa | CBS328.50 | AF357041 | AF223197 | [1] |
| S. gibberosa | CBS320.80/IFO 30977 | AF357042 | AF223198 | [41] |
| S. paluster | CBS717.87 | AF357044 | AF223200 | [1] |
| S. tylicolor | IE-BSG-BSI92/245 | AF357040 | AF223195 | [1] |
| S. tylicolor | IE-BSG-Sag5-27/11 | AF357039 | AF223194 | [41] |
| S. paluster | CBS714.87 | AF357043 | AF223199 | [41] |
| Tephrocybella griseonigrescens | TO HG 21112014 | KR105775 | KR476785 | [47] |
| Tephroderma fuscopallens | LUG18989 | KJ701327 | KJ701333 | [47] |
| T. fuscopallens | EM4789-12 | KJ701326 | KJ701332 | [47] |
| Termitomyces heimii | DUKE-JMleg.MUIDs.n. | AF357022 | AF042586 | [41] |
| T. microcarpus | DUKE-PRU3900 | AF357023 | AF042578 | [1] |
| T. radicatus | CBS204.47 | AF357025 | AF223203 | [1] |
| T. sp. | IE-BSG-BSI sp.1 | AF357024 | AF223174 | [1] |
| Tricholomella constricta | IE-BSG-HC84/75 | AF357036 | AF223188 | [1] |
| Tricholyophyllum brunneum | HKAS107494 | MT705717 | MT734655 | [24] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, S.-W.; Lu, B.-Y.; Wang, Y.; Dou, W.-J.; Wang, Q.; Li, Y. Morphology and Phylogeny of Lyophylloid Mushrooms in China with Description of Four New Species. J. Fungi 2023, 9, 77. https://doi.org/10.3390/jof9010077
Wei S-W, Lu B-Y, Wang Y, Dou W-J, Wang Q, Li Y. Morphology and Phylogeny of Lyophylloid Mushrooms in China with Description of Four New Species. Journal of Fungi. 2023; 9(1):77. https://doi.org/10.3390/jof9010077
Chicago/Turabian StyleWei, Shu-Wei, Bo-Yu Lu, Yang Wang, Wen-Jun Dou, Qi Wang, and Yu Li. 2023. "Morphology and Phylogeny of Lyophylloid Mushrooms in China with Description of Four New Species" Journal of Fungi 9, no. 1: 77. https://doi.org/10.3390/jof9010077