
Phylogenetic Analyses and Morphological Studies Reveal Four New Species of Phellodon (Bankeraceae, Thelephorales) from China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological Studies
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Phylogenetic Analyses
3. Results
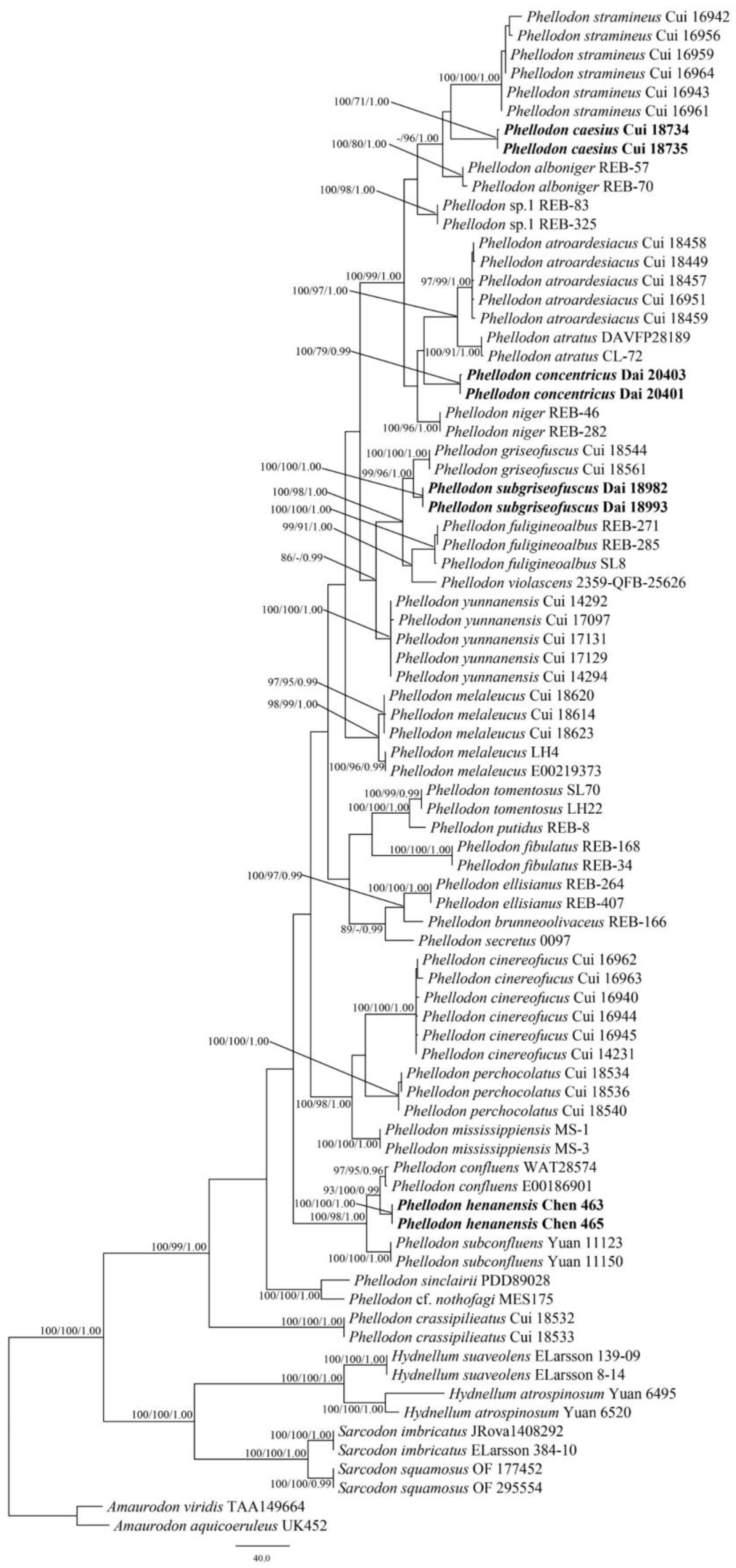
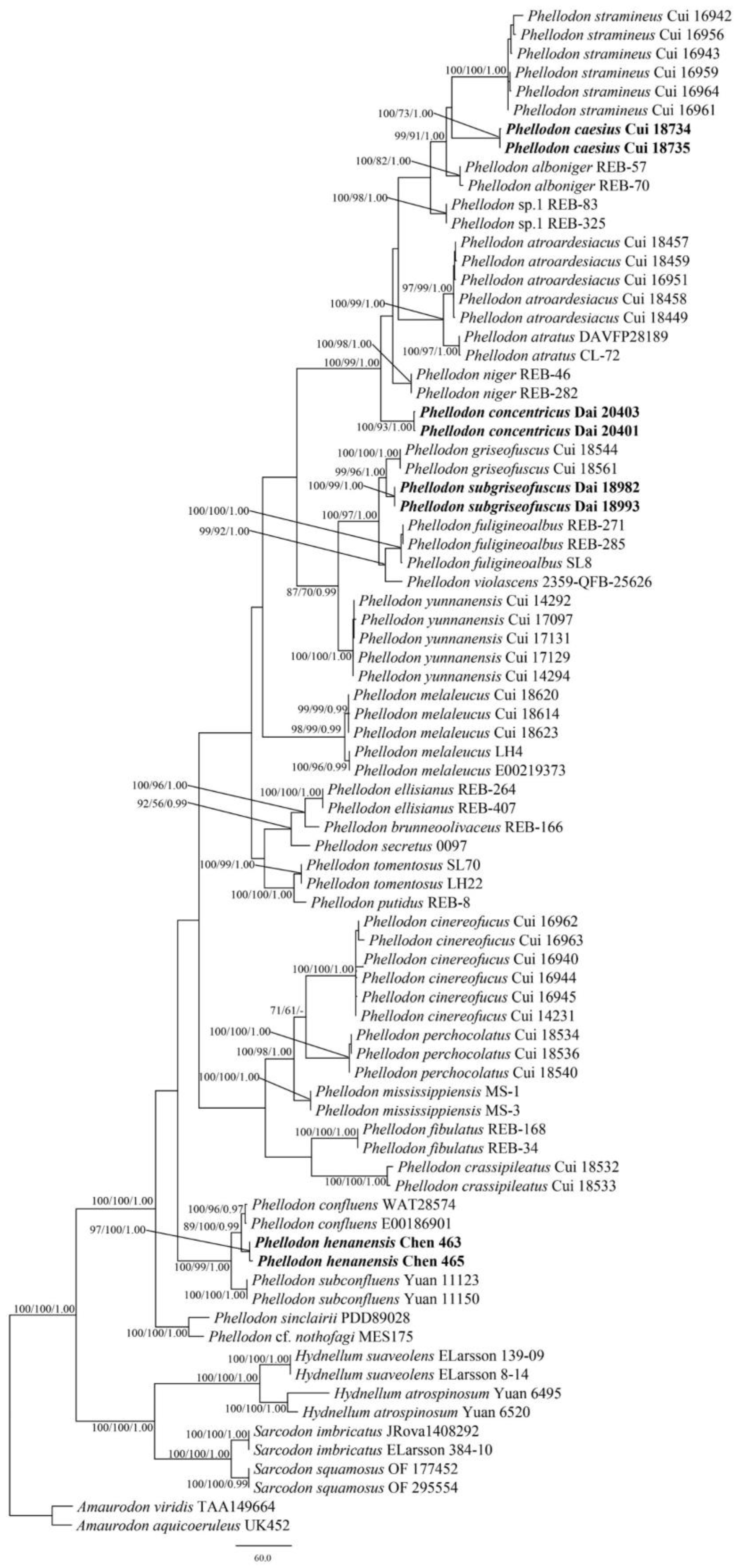
3.1. Phylogenetic Analyses
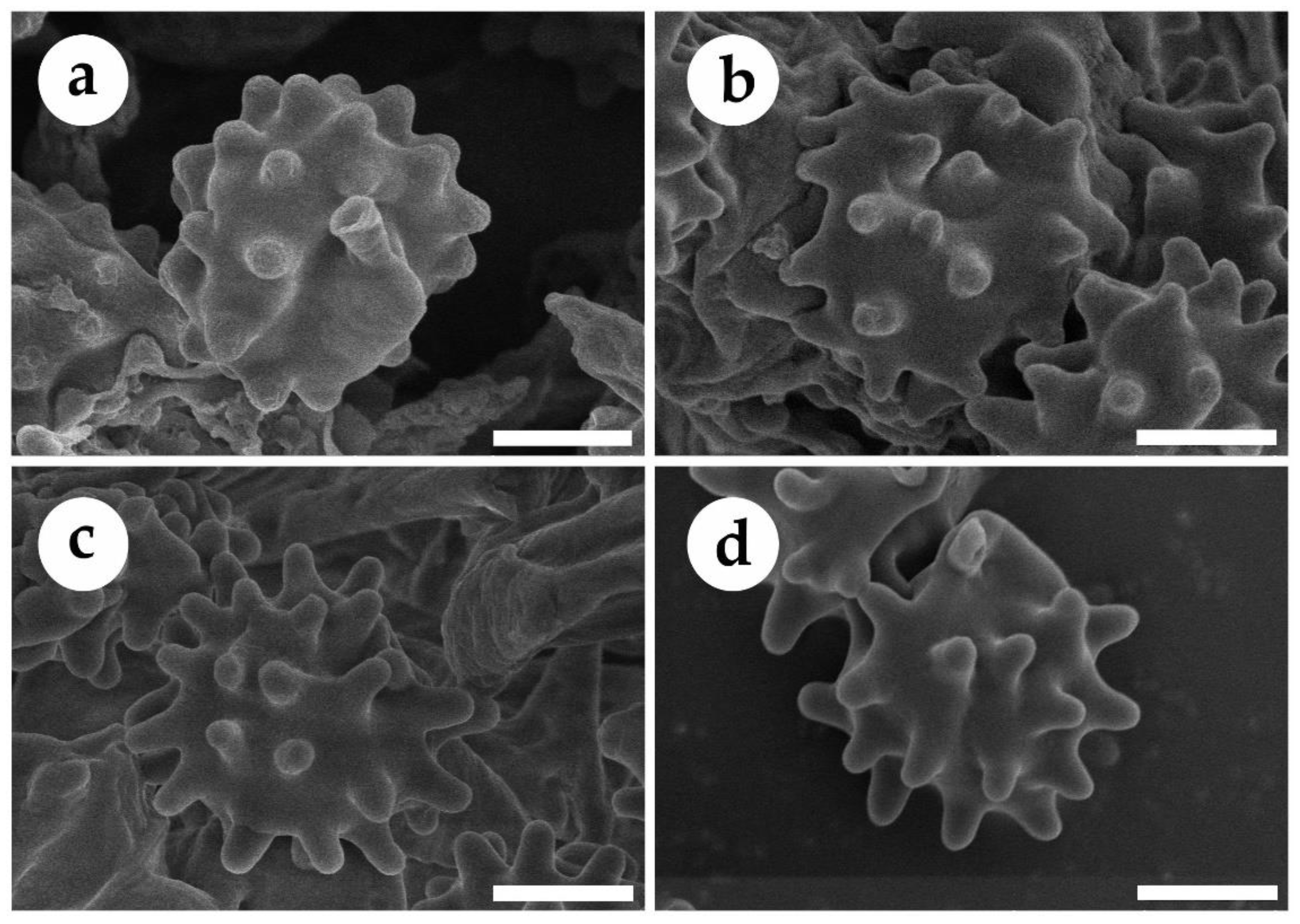
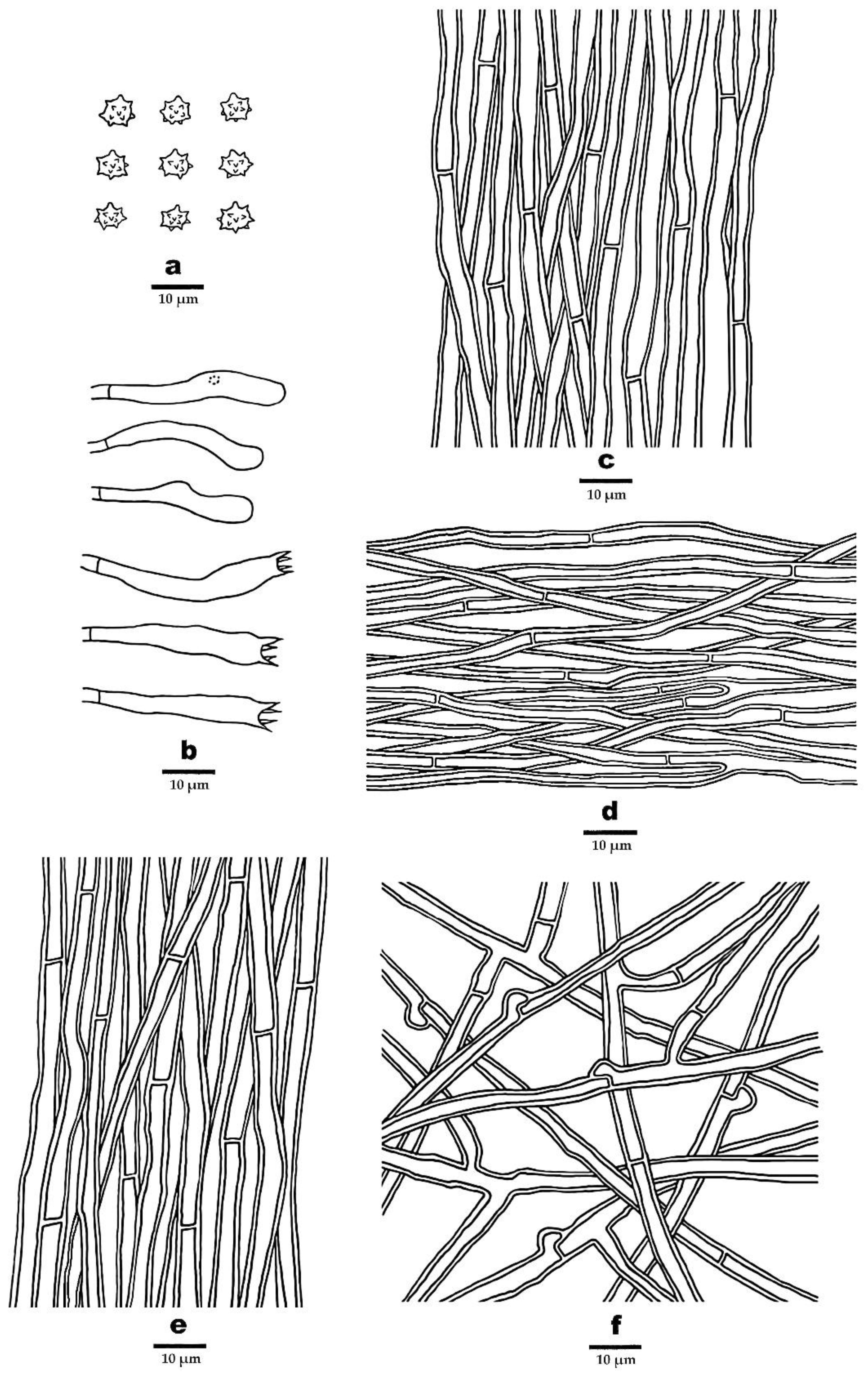
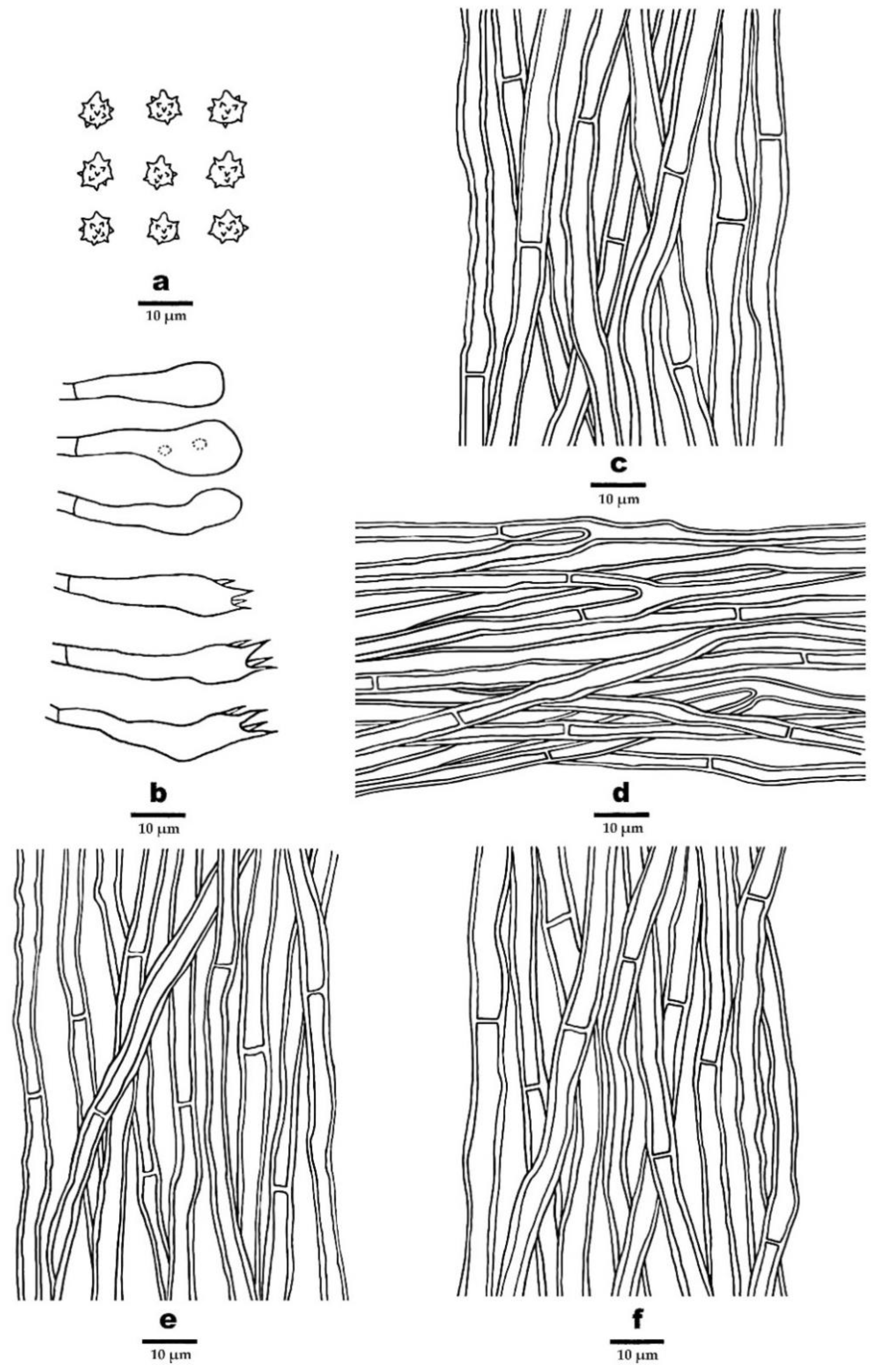
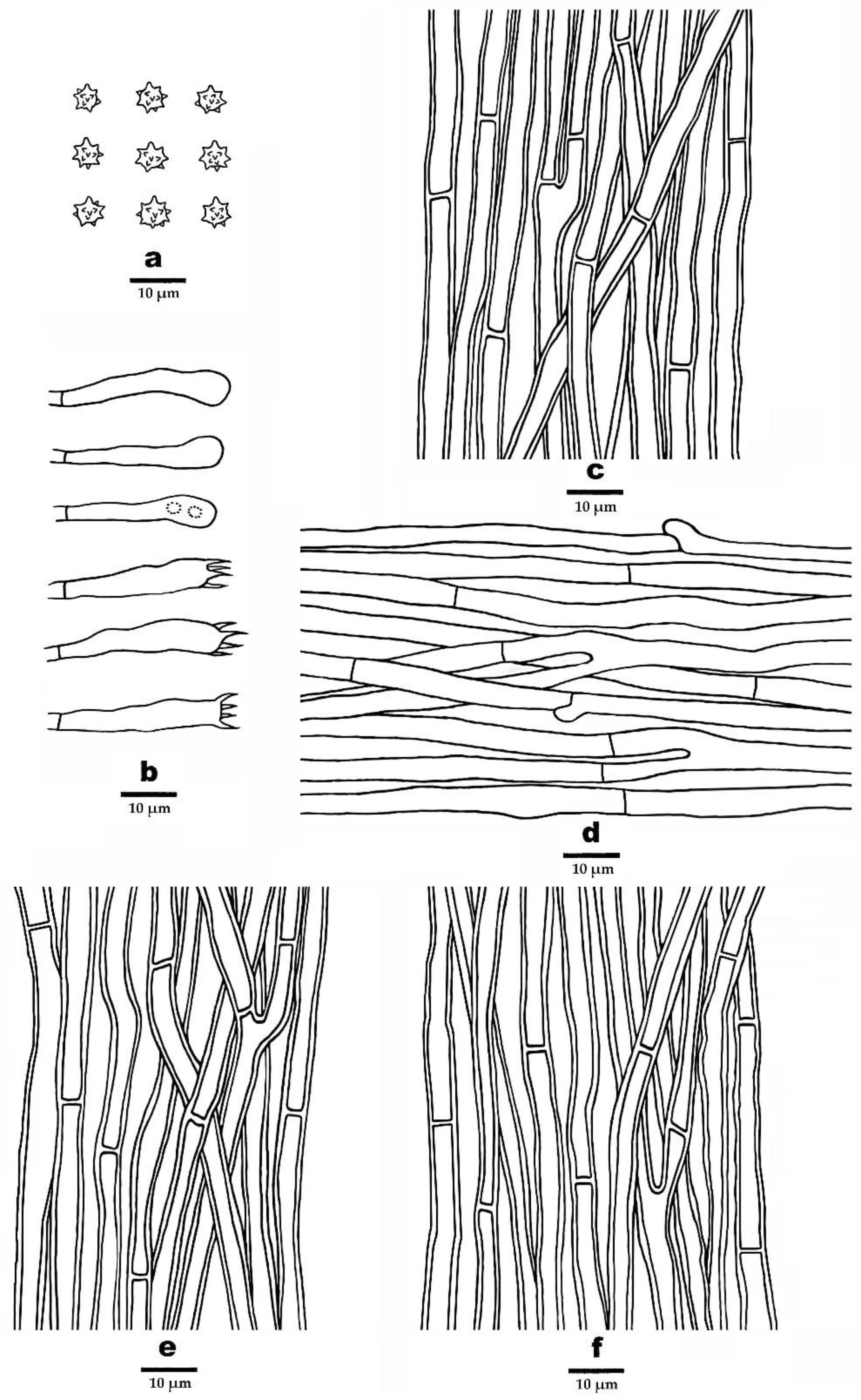
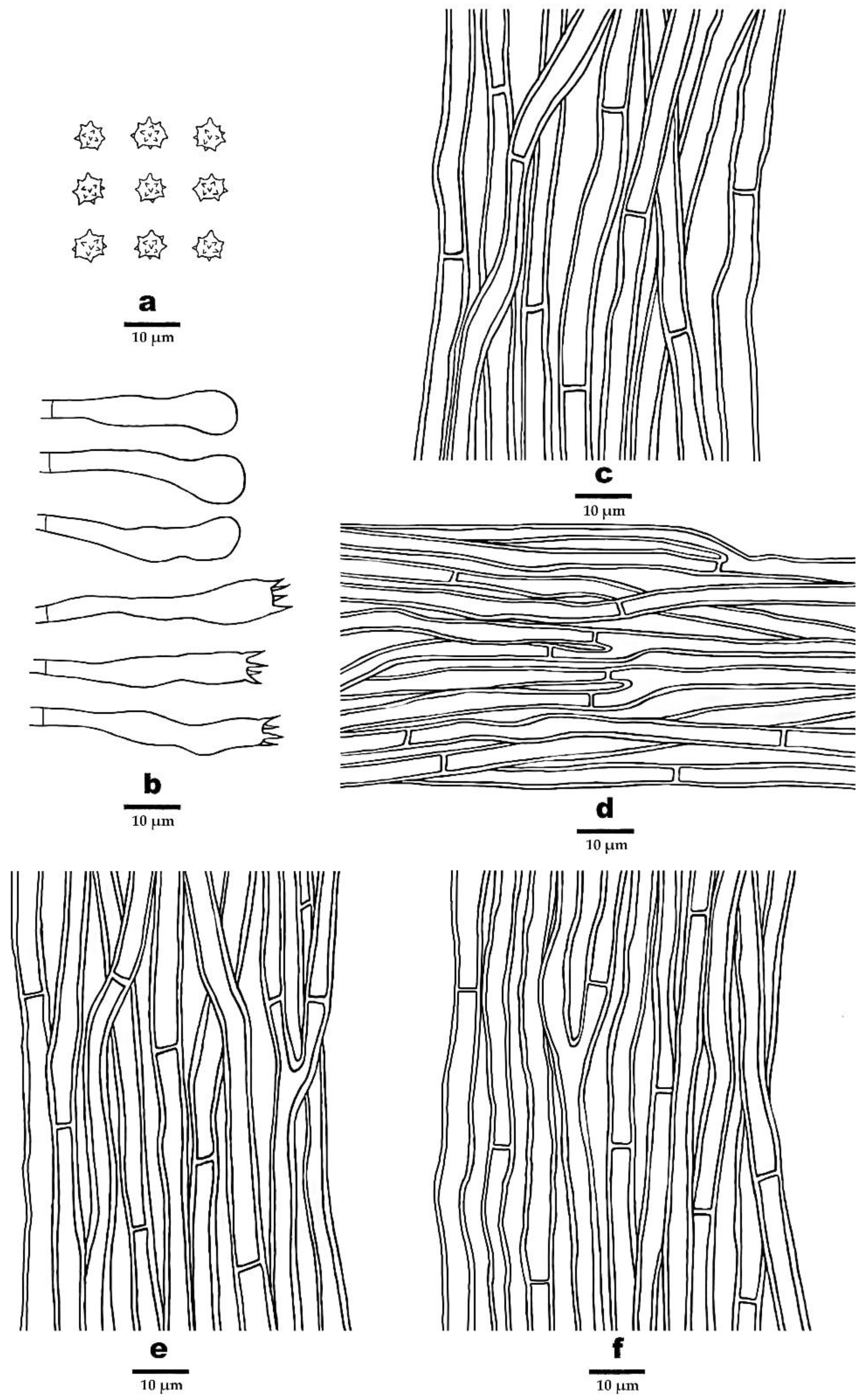
3.2. Taxonomy
- 1.
- Pileal surface straw buff-------------------------------------------------------------------------------P. stramineus
- 1.
- Pileal surface differently colored--------------------------------------------------------------------2
- 2.
- Pileal surface blackish-blue to dark grey or bluish-grey to dark bluish-grey-------------3
- 2.
- Pileal surface differently colored--------------------------------------------------------------------4
- 3.
- Clamp connections exist in spines------------------------------------------------------------------P. atroardesiacus
- 3.
- Clamp connections do not exist in spines---------------------------------------------------------P. caesius
- 4.
- Tissues color changed in KOH-----------------------------------------------------------------------5
- 4.
- Tissues color unchanged in KOH-------------------------------------------------------------------P. subconfluens
- 5.
- Pileal surface glabrous---------------------------------------------------------------------------------6
- 5.
- Pileal surface not glabrous----------------------------------------------------------------------------8
- 6.
- Pileal surface reddish-brown to cinnamon brown----------------------------------------------P. cinereofuscus
- 6.
- Pileal surface differently colored--------------------------------------------------------------------7
- 7.
- Pileal surface clay pink to brown-------------------------------------------------------------------P. yunnanensis
- 7.
- Pileal surface fuscous to black-----------------------------------------------------------------------P. subgriseofuscus
- 8.
- Clamp connections exist-------------------------------------------------------------------------------9
- 8.
- Clamp connections absent----------------------------------------------------------------------------P. concentricus
- 9.
- Pileal surface ash grey, light vinaceous grey to light brown---------------------------------P. henanensis
- 9.
- Pileal surface differently colored-------------------------------------------------------------------10
- 10.
- Clamp connections exist in spines----------------------------------------------------------------11
- 10.
- Clamp connections do not exist in spines-------------------------------------------------------P. crassipileatus
- 11.
- Spines brown after mature--------------------------------------------------------------------------P. griseofuscus
- 11.
- Spines white after mature---------------------------------------------------------------------------P. perchocolatus
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karsten, P.A. Enumeratio boletinearum et polyporearum fennicarum, systemate novo dispositarum. Rev. Mycol. 1881, 3, 16–19. [Google Scholar]
- Maas Geesteranus, R.A. Hydnaceous fungi of the eastern old world. Verh. Kon. Ned. Akad. Wetensch. Afd. Natuurk 1971, 60, 176. [Google Scholar]
- Erland, S.; Taylor, A.F.S. Resupinate ectomycorrhizal fungal genera. In Ectomycorrhizal Fungi Key Genera in Profile; Cairney, J.W.G., Chambers, S.M., Eds.; Springer: Berlin/Heidelberg, Germany, 1999; pp. 347–363. [Google Scholar]
- Brundrett, M.C. Mycorrhizal associations and other means of nutrition of vascular plants: Understanding the global diversity of host plants by resolving conflicting information and developing reliable means of diagnosis. Plant Soil 2009, 320, 37–77. [Google Scholar] [CrossRef]
- Van der Heijden, M.; Klironomos, J.; Ursic, M.; Moutoglis, P.; Streitwolf-Engel, R.; Boller, T.; Wiemken, A.; Sanders, I. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 1998, 396, 69–72. [Google Scholar] [CrossRef]
- Stadler, M.; Anke, T.; Dasenbrock, J.; Steglich, W. Phellodonic Acid, a new biologically active hirsutane derivative from Phellodon melaleucus (Thelephoraceae, Basidiomycetes). Z. Nat. C 1993, 48, 545–549. [Google Scholar] [CrossRef]
- Reekie, T.A.; Austin, K.A.; Banwell, M.G.; Willis, A.C. The chemoenzymatic total synthesis of phellodonic acid, a biologically active and highly functionalized hirsutane derivative isolated from the tasmanian fungus Phellodon melaleucus. Aust. J. Chem. 2008, 61, 94–106. [Google Scholar] [CrossRef]
- Fang, S.T.; Zhang, L.; Li, Z.H.; Li, B.; Liu, J.K. Cyathane diterpenoids and nitrogenous terphenyl derivative from the fruiting bodies of basidiomycete Phellodon niger. Chem. Pharm. Bull. 2010, 58, 1176–1179. [Google Scholar] [CrossRef] [Green Version]
- Banker, H.J. A contribution to a revision of the North American Hydnaceae. Mem. Torrey Bot. Club 1906, 12, 99–194. [Google Scholar]
- Donk, M.A. Four new families of Hymenomycetes. Persoonia 1961, 1, 405–407. [Google Scholar]
- Baird, R.E.; Wallace, L.E.; Baker, G.; Scruggs, M. Stipitate hydnoid fungi of the temperate southeastern United States. Fungal Divers. 2013, 62, 41–114. [Google Scholar] [CrossRef]
- Harrison, K.A. New or little known north American stipitate hydnums. Can. J. Bot. 1964, 42, 1205–1233. [Google Scholar] [CrossRef]
- Harrison, K.A. A new species of Phellodon possessing clamp connections. Can. J. Bot. 1972, 50, 1219–1221. [Google Scholar] [CrossRef]
- Hrouda, P. Hydnaceous fungi of the Czech Republic and Slovakia. Czech Mycol. 1999, 51, 99–155. [Google Scholar] [CrossRef]
- Hrouda, P. Bankeraceae in Central Europe. 1. Czech Mycol. 2005, 57, 57–78. [Google Scholar] [CrossRef]
- Hrouda, P. Bankeraceae in Central Europe. 2. Czech Mycol. 2005, 57, 279–297. [Google Scholar] [CrossRef]
- Coker, W.C.; Beers, A.H. The Stipitate Hydnums of the Eastern United States; University of North Carolina Press: Chapel Hill, NC, USA, 1951; p. 211. [Google Scholar]
- Pouzar, Z.D. Příspěvek k poznání našich kloboukatých lošáků. Čes. Mykol. 1956, 10, 65–76. [Google Scholar]
- Maas Geesteranus, R.A. The stipitate hydnums of the Netherlands. Fungus 1958, 28, 48–61. [Google Scholar]
- Maas Geesteranus, R.A. Notes on the hydnums. Persoonia 1960, 1, 341–384. [Google Scholar]
- Maas Geeseranus, R.A. Hyphal structures in the hydnums. Persoonia 1962, 2, 476. [Google Scholar]
- Maas Geesteranus, R.A. The terrestrial hydnums of Europe. Verh. Kon. Ned. Akad. Wetensch. Afd. Natuurk 1975, 65, 127. [Google Scholar]
- Parfitt, D.; Ainsworth, A.M.; Simpson, D.; Rogers, H.J.; Boddy, L. Molecular and morphological discrimination of stipitate hydnoids in the genera Hydnellum and Phellodon. Mycol. Res. 2007, 3, 761–777. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, A.M.; Parfitt, D.; Rogers, H.J.; Boddy, L. Cryptic taxa within European species of Hydnellum and Phellodon revealed by combined molecular and morphological analysis. Fungal Ecol. 2010, 3, 65–80. [Google Scholar] [CrossRef]
- Baird, R.E.; Wallace, L.E.; Baker, G. Stipitate hydnums of the southern United States 1: Phellodon mississippiensis sp. nov. Mycotaxon 2013, 123, 183–191. [Google Scholar] [CrossRef]
- Li, V. Phylogenetic Study of Family Bankeraceae in Korea. Master’s Thesis, Seoul National University, Seoul, Korea, 2017. [Google Scholar]
- Song, C.G.; Ji, X.; Liu, S.; He, X.L.; Cui, B.K. Taxonomy and molecular phylogeny of Phellodon (Thelephorales) with descriptions of four new species from Southwest China. Forests 2021, 12, 932. [Google Scholar] [CrossRef]
- Song, C.G.; Chen, Y.Y.; Liu, S.; Xu, T.M.; He, X.L.; Wang, D.; Cui, B.K. A phylogenetic and taxonomic study on Phellodon (Bankeraceae, Thelephorales) from China. J. Fungi 2022, 8, 429. [Google Scholar] [CrossRef]
- Mu, Y.H.; Wu, F.; Yuan, H.S. Hydnaceous fungi of China 7. Morphological and molecular characterization of Phellodon subconfluens sp. nov. from temperate, deciduous forests. Phytotaxa 2019, 414, 280–288. [Google Scholar] [CrossRef]
- Han, M.L.; Chen, Y.Y.; Shen, L.L.; Song, J.; Vlasák, J.; Dai, Y.C.; Cui, B.K. Taxonomy and phylogeny of the brown-rot fungi: Fomitopsis and its related genera. Fungal Divers. 2016, 80, 343–373. [Google Scholar] [CrossRef] [Green Version]
- Cui, B.K.; Li, H.J.; Ji, X.; Zhou, J.L.; Song, J.; Si, J.; Yang, Z.L.; Dai, Y.C. Species diversity, taxonomy and phylogeny of Polyporaceae (Basidiomycota) in China. Fungal Divers. 2019, 97, 137–392. [Google Scholar] [CrossRef]
- Mu, Y.H.; Yu, J.R.; Cao, T.; Wang, X.H.; Yuan, H.S. Multi-Gene phylogeny and taxonomy of Hydnellum (Bankeraceae, Basidiomycota) from China. J. Fungi 2021, 7, 818. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.L.; Wang, M.; Zhou, J.L.; Xing, J.H.; Cui, B.K.; Dai, Y.C. Taxonomy and phylogeny of Postia. Multi-gene phylogeny and taxonomy of the brown-rot fungi: Postia (Polyporales, Basidiomycota) and related genera. Persoonia 2019, 42, 101–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.F.; Costa-Rezende, D.H.; Xing, J.H.; Zhou, J.L.; Zhang, B.; Gibertoni, T.B.; Gates, G.; Glen, M.; Dai, Y.C.; Cui, B.K. Multi-gene phylogeny and taxonomy of Amauroderma s. lat. (Ganodermataceae). Persoonia 2020, 44, 206–239. [Google Scholar] [CrossRef] [PubMed]
- Cao, T.; Hu, Y.P.; Yu, J.R.; Wei, T.Z.; Yuan, H.S. A phylogenetic overview of the Hydnaceae (Cantharellales, Basidiomycota) with new taxa from China. Stud. Mycol. 2021, 99, 100–121. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.S.; Kang, R.; Zeng, N.K.; Yu, W.J.; Chang, Z.; Xu, F.; Deng, W.Q.; Qi, L.L.; Zhou, Y.L.; Fan, Y.G. Two new Inosperma (Inocybaceae) species with unexpected muscarine contents from tropical China. MycoKeys 2021, 85, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.S.; Yu, W.J.; Zeng, N.K.; Zhang, Y.Z.; Wu, X.P.; Li, H.J.; Xu, F.; Fan, Y.G. A new muscarine-containing Inosperma (Inocybaceae, Agaricales) species discovered from one poisoning incident occurring in tropical China. Front. Microbiol. 2022, 13, 923435. [Google Scholar] [CrossRef]
- Liu, S.; Han, M.L.; Xu, T.M.; Wang, Y.; Wu, D.M.; Cui, B.K. Taxonomy and phylogeny of the Fomitopsis pinicola complex with descriptions of six new species from east Asia. Front. Microbiol. 2021, 12, 644979. [Google Scholar] [CrossRef]
- Liu, S.; Shen, L.L.; Wang, Y.; Xu, T.M.; Gates, G.; Cui, B.K. Species diversity and molecular phylogeny of Cyanosporus (Polyporales, Basidiomycota). Front. Microbiol. 2021, 12, 631166. [Google Scholar] [CrossRef]
- Liu, S.; Song, C.G.; Xu, T.M.; Ji, X.; Wu, D.M.; Cui, B.K. Species diversity, molecular phylogeny, and ecological habits of Fomitopsis (Polyporales, Basidiomycota). Front. Microbiol. 2022, 13, 859411. [Google Scholar] [CrossRef]
- Liu, S.; Chen, Y.Y.; Sun, Y.F.; He, X.L.; Song, C.G.; Si, J.; Liu, D.M.; Gates, G.; Cui, B.K. Systematic classification and phylogenetic relationships of the brown-rot fungi within the Polyporales. Fungal Divers. 2022. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, C.Q.; Buyck, B.; Deng, W.Q.; Li, T.H. Multigene phylogeny and morphology reveal unexpectedly high number of new species of Cantharellus subgenus Parvocantharellus (Hydnaceae, Cantharellales) in China. J. Fungi 2021, 7, 919. [Google Scholar] [CrossRef]
- Ji, X.; Zhou, J.L.; Song, C.G.; Xu, T.M.; Wu, D.M.; Cui, B.K. Taxonomy, phylogeny and divergence times of Polyporus (Basidiomycota) and related genera. Mycosphere 2022, 13, 1–52. [Google Scholar] [CrossRef]
- Wang, Y.; Tuo, Y.L.; Wu, D.M.; Gao, N.; Zhang, Z.H.; Rao, G.; Wang, X.M.; Wang, J.; Dai, D.; Li, Y.; et al. Exploring the relationships between four new species of boletoid fungi from Northern China and their related species. J. Fungi 2022, 8, 218. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.F.; Xing, J.H.; He, X.L.; Wu, D.M.; Song, C.G.; Liu, S.; Vlasák, J.; Gates, G.; Gibertoni, T.B.; Cui, B.K. Species diversity, systematic revision and molecular phylogeny of Ganodermataceae (Polyporales, Basidiomycota) with an emphasis on Chinese collections. Stud. Mycol. 2022, 101, 287–415. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. Bioedit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.2. 2017. Available online: http://mesquiteproject.org (accessed on 19 December 2022).
- Farris, J.S.; Källersjö, M.; Kluge, A.G.; Kluge, A.G.; Bult, C. Testing significance of incongruence. Cladistics 1994, 10, 315–319. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods), Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002.
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. methods. 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence intervals on phylogenetics: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analysis with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [Green Version]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest v2. Program. Distributed by the Author; Evolutionary Biology Center, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Specimen No. | Locality | GenBank Accession No. | ||||
|---|---|---|---|---|---|---|---|
| ITS | nrLSU | nuSSU | RPB1 | RPB2 | |||
| Amaurodon aquicoeruleus | UK 452 | Australia | AM490944 | AM490944 | - | - | - |
| A. viridis | TAA 149664 | Russia | AM490942 | AM490942 | - | - | - |
| Hydnellum atrospinosum | Yuan 6520 | China | MW579912 | - | MW579912 | - | - |
| H. atrospinosum | Yuan 6495 | China | MW579938 | MW579885 | MW579911 | - | - |
| H. suaveolens | ELarsson 139-09 | Norway | MK602734 | MK602734 | - | - | - |
| H. suaveolens | ELarsson 8-14 | Sweden | MK602735 | MK602735 | - | - | - |
| P. alboniger | REB-70 | USA | KC571749 | - | - | - | - |
| P. alboniger | REB-57 | USA | JN135206 | - | - | - | - |
| P. atratus | CL-72 | Canada | MK281471 | - | - | - | - |
| P. atratus | DAVFP 28189 | Canada | HQ650766 | - | - | - | - |
| P. atroardesiacus | Cui 18449 | China | MZ221189 | MZ225598 | MZ225636 | - | - |
| P. atroardesiacus | Cui 18457 | China | MZ225577 | MZ225599 | MZ225637 | - | - |
| P. atroardesiacus | Cui 18458 | China | MZ225633 | MZ225600 | MZ225638 | - | - |
| P. atroardesiacus | Cui 18459 | China | MZ225634 | MZ225601 | MZ225639 | - | - |
| P. atroardesiacus | Cui 16951 | China | MZ225632 | MZ225597 | MZ225635 | MZ343209 | MZ343197 |
| P. brunneoolivaceus | REB-166 | USA | KC571752 | - | - | - | - |
| P. caesius | Cui 18734 | China | OP751005 | OP751407 | OP751414 | OP755302 | OP755305 |
| P. caesius | Cui 18735 | China | - | OP751408 | OP751415 | OP755303 | - |
| P. cinereofuscus | Cui 14231 | China | MZ225579 | - | - | - | - |
| P. cinereofuscus | Cui 16940 | Australia | MZ225580 | MZ225602 | MZ225640 | MZ343210 | MZ343198 |
| P. cinereofuscus | Cui 16944 | China | MZ225581 | MZ225603 | MZ225641 | MZ343211 | MZ343199 |
| P. cinereofuscus | Cui 16945 | China | MZ225582 | MZ225604 | MZ225642 | - | - |
| P. cinereofuscus | Cui 16962 | China | MZ225583 | MZ225605 | MZ225643 | MZ352084 | MZ343200 |
| P. cinereofuscus | Cui 16963 | China | MZ225584 | MZ225606 | MZ225644 | MZ352085 | MZ343201 |
| P. concentricus | Dai 20401 | China | - | OP751406 | OP751413 | OP755301 | - |
| P. concentricus | Dai 20403 | China | OP751004 | OP751405 | OP751412 | - | - |
| P. confluens | WAT 28574 | UK | EU622361 | - | - | - | - |
| P. confluens | E00 186901 | UK | EU622362 | - | - | - | - |
| P. crassipilieatus | Cui 18532 | China | OL449267 | OL439037 | OL439027 | - | - |
| P. crassipilieatus | Cui 18533 | China | OL449268 | OL439038 | OL439028 | - | - |
| P. ellisianus | REB-264 | USA | KC571757 | - | - | - | - |
| P. ellisianus | REB-407 | USA | KC571759 | - | - | - | - |
| P. fibulatus | REB-168 | USA | JN135205 | - | - | - | - |
| P. fibulatus | REB-34 | USA | KC571761 | - | - | - | - |
| P. fuligineoalbus | REB-271 | USA | KC571760 | - | - | - | - |
| P. fuligineoalbus | REB-285 | USA | JN135196 | - | - | - | - |
| P. fuligineoalbus | SL8 | - | EU622316 | - | - | - | - |
| P. griseofuscus | Cui 18544 | China | OL449265 | OL439035 | OL439025 | OL456229 | OL449087 |
| P. griseofuscus | Cui 18561 | China | OL449266 | OL439036 | OL439026 | - | - |
| P. henanensis | Chen 463 | China | OP751002 | - | OP751410 | - | - |
| P. henanensis | Chen 465 | China | OP751003 | OP751404 | OP751411 | - | - |
| P. melaleucus | LH4 | UK | EU622368 | - | - | - | - |
| P. melaleucus | E00219373 | UK | EU622369 | - | - | - | - |
| P. melaleucus | Cui 18614 | China | OL449262 | OL439032 | OL439022 | - | |
| P. melaleucus | Cui 18620 | China | OL449263 | OL439033 | OL439023 | - | - |
| P. melaleucus | Cui 18623 | China | OL449264 | OL439034 | OL439024 | - | - |
| P. mississippiensis | MS-1 | USA | JN247563 | - | - | - | - |
| P. mississippiensis | MS-3 | USA | JN247564 | - | - | - | - |
| P. niger | REB-46 | USA | JN135202 | - | - | - | - |
| P. niger | REB-282 | USA | KC571766 | - | - | - | - |
| P. cf. nothofagi | MES-175 | Chile | MH930224 | - | - | - | - |
| P. perchocolatus | Cui 18534 | China | OL449259 | OL439029 | OL439020 | - | |
| P. perchocolatus | Cui 18536 | China | OL449260 | OL439030 | - | - | - |
| P. perchocolatus | Cui 18540 | China | OL449261 | OL439031 | OL439021 | - | - |
| P. putidus | REB-8 | USA | JN135200 | - | - | - | - |
| P. secretus | 0097 | Russia | MG597404 | - | - | - | - |
| P. sinclairii | PDD 89028 | New Zealand | GU222291 | - | - | - | - |
| P. stramineus | Cui 16942 | China | MZ225585 | MZ225607 | MZ225645 | MZ352086 | - |
| P. stramineus | Cui 16943 | China | MZ225586 | MZ225608 | MZ225646 | MZ352087 | MZ343202 |
| P. stramineus | Cui 16956 | China | MZ225587 | MZ225609 | MZ225647 | MZ352088 | MZ343203 |
| P. stramineus | Cui 16959 | China | MZ225588 | MZ225610 | MZ225648 | MZ352089 | MZ343204 |
| P. stramineus | Cui 16961 | China | MZ225589 | MZ225611 | MZ225649 | MZ352090 | MZ343205 |
| P. stramineus | Cui 16964 | China | MZ225590 | MZ225612 | MZ225650 | MZ352091 | - |
| P. subconfluens | Yuan 11123 | China | MK677464 | - | - | - | - |
| P. subconfluens | Yuan 11150 | China | MK677465 | - | - | - | - |
| P. subgriseofuscus | Dai 18982 | China | OP751000 | - | - | - | - |
| P. subgriseofuscus | Dai 18993 | China | OP751001 | OP751403 | OP751409 | - | OP755301 |
| Phellodon sp.1 | REB-83 | USA | KC571747 | - | - | - | - |
| Phellodon sp.1 | REB-325 | USA | KC571748 | - | - | - | - |
| P. tomentosus | SL70 | UK | EU622381 | - | - | - | - |
| P. tomentosus | LH22 | UK | EU622382 | - | - | - | |
| P. yunnanensis | Cui 14292 | China | MZ225591 | - | - | - | - |
| P. yunnanensis | Cui 14294 | China | MZ225592 | - | - | - | - |
| P. yunnanensis | Cui 17097 | China | MZ225593 | MZ225613 | MZ225651 | - | MZ343206 |
| P. yunnanensis | Cui 17129 | China | MZ225594 | MZ225614 | MZ225652 | - | MZ343207 |
| P. yunnanensis | Cui 17131 | China | MZ225595 | MZ225615 | MZ225653 | - | MZ343208 |
| P. violascens | 2359-QFB-25626 | - | KM406977 | - | - | - | - |
| Sarcodon imbricatus | JRova 1408292 | Sweden | MK602746 | MK602746 | - | - | - |
| S. imbricatus | ELarsson 384-10 | Norway | MK602747 | MK602747 | - | - | - |
| S. squamosus | OF 177452 | Norway | MK602768 | MK602768 | - | - | - |
| S. squamosus | OF 295554 | Norway | MK602769 | MK602769 | - | - | - |
| Species | Distribution in China | Ecological Habits | Alt. | Pileal Surface | Spines Color | Spines Size (mm) | Clamp Connection | Basidios-Pores (µm) | References |
|---|---|---|---|---|---|---|---|---|---|
| P. atroardesiacus | Xizang Autonomous Region | in Pinus densata forest | 2900 m | blackish-blue to dark grey when fresh | dark greyish-blue to ash grey when fresh | up to 5 | occasionally with clamp connections in spines | 4–5 × (3–) 3.5–4.5 | Song et al., 2021 [27] |
| P. caesius | Sichuan Province | on the ground of forest dominated by Quercus aquifolioides | 3320 m | bluish-grey, dark bluish-grey to black-grey when fresh | white, ash grey to dark bluish-grey when fresh | up to 2 | occasionally with clamp connections in the surface layer of stipe | 4–5.6(–6) × (3.8–)4–5.2 | This study |
| P. cinereofuscus | Yunnan Province | on the ground of forest dominated by Pinus and Fagaceae forest, and mixed forest | 1800–2250 m | reddish-brown to cinnamon brown when fresh | greyish-brown to white when fresh | up to 6 | unclamped | 4–5 × (3.5–) 4–4.5 | Song et al., 2021 [27] |
| P. concentricus | Yunnan Province | on the ground of forest dominated by Quercus | 2088 m | dark olive to mouse grey when dry | ash grey when dry | up to 2.5 | unclamped | 5–6.2 × 4.5–5.5(–5.7) | This study |
| P. crassipileatus | Sichuan Province | on the ground of forest dominated by Quercus | 1190 m | pale brown to dark brown when fresh | white when fresh | up to 3 | with clamp connections in pileus and stipe | (3.5–) 4–5 × 4–5 | Song et al., 2022 [28] |
| P. griseofuscus | Sichuan Province | i on the ground of forest dominated by Pinus and Picea | 2400 m | dark brown to black when fresh | white when young and brown with age when fresh | up to 1 | with clamp connections in spines | 4–5 × 3.5–4.5 | Song et al., 2022 [28] |
| P. henanensis | Henan Province | on the ground of mixed forest | 2000 m | ash grey, light vinaceous grey to light brown when fresh | ash grey to light brown when fresh | up to 1 | occasionally with clamp connections in spines | (3.2–)3.8–5 × (3–)3.5–4.5(–4.8) | This study |
| P. perchocolatus | Sichuan Province | on the ground of forest dominated by Quercus | 1190 m | brown to greyish-brown when fresh | white when fresh | up to 3 | with clamp connections in spines | 4–5 (–5.5) × (3.5–) 4–4.5 (–5) | Song et al., 2022 [28] |
| P. stramineus | Yunnan Province | on the ground of forest dominated by Pinus yunnanensis and Fagaceae | 2250 m | straw buff when fresh | dark grey to ash grey when fresh | up to 3 | unclamped | 4–5.5 (–6) × 4–5 (–5.5) | Song et al., 2021 [27] |
| P. subconfluens | Liaojing Province | on the ground of forest dominated by Quercus | 870 m | greyish-buff, brownish-orange to reddish-brown when fresh | cream to greyish-buff when fresh | up to 1 | unclamped | (3.0–) 3.1–4.1 (–4.8) × (2.5–) 2.9–3.5 (–3.8) | Mu et al., 2019 [29] |
| P. subgriseofuscus | Gansu Province | on the ground of forest dominated by Picea crassifolia | 2250–3000 m | dark brown to black when fresh | white to light brown when fresh | up to 2.5 | unclamped | 4–5 × (3–)3.2–4.8 | This study |
| P. yunnanensis | Yunnan Province | on the ground of Pinus and Fagaceae forest or Pinus forest or Pinus armandii and Rhododendron forest | 2300–2600 m | clay pink to brown when fresh | pale brown to white when fresh | up to 5 | occasionally with clamp connections in stipe | 3.5–4.5(–5) × 3–4 (–4.5) | Song et al., 2021 [27] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, C.-G.; Sun, Y.-F.; Liu, S.; Chen, Y.-Y.; Cui, B.-K. Phylogenetic Analyses and Morphological Studies Reveal Four New Species of Phellodon (Bankeraceae, Thelephorales) from China. J. Fungi 2023, 9, 30. https://doi.org/10.3390/jof9010030
Song C-G, Sun Y-F, Liu S, Chen Y-Y, Cui B-K. Phylogenetic Analyses and Morphological Studies Reveal Four New Species of Phellodon (Bankeraceae, Thelephorales) from China. Journal of Fungi. 2023; 9(1):30. https://doi.org/10.3390/jof9010030
Chicago/Turabian StyleSong, Chang-Ge, Yi-Fei Sun, Shun Liu, Yuan-Yuan Chen, and Bao-Kai Cui. 2023. "Phylogenetic Analyses and Morphological Studies Reveal Four New Species of Phellodon (Bankeraceae, Thelephorales) from China" Journal of Fungi 9, no. 1: 30. https://doi.org/10.3390/jof9010030