
Description and Genome Characterization of Three Novel Fungal Strains Isolated from Mars 2020 Mission-Associated Spacecraft Assembly Facility Surfaces—Recommendations for Two New Genera and One Species

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Isolation of Fungi
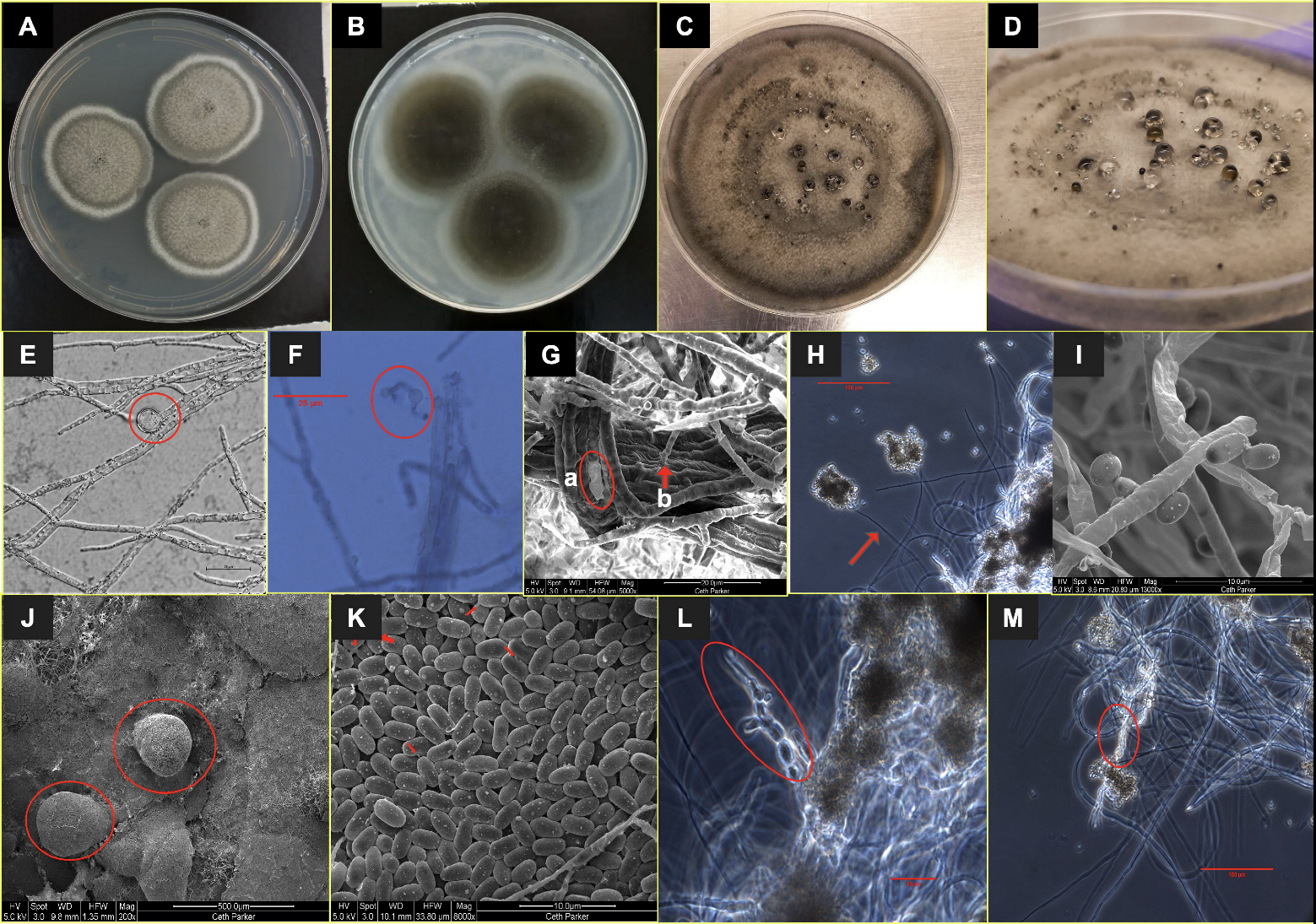
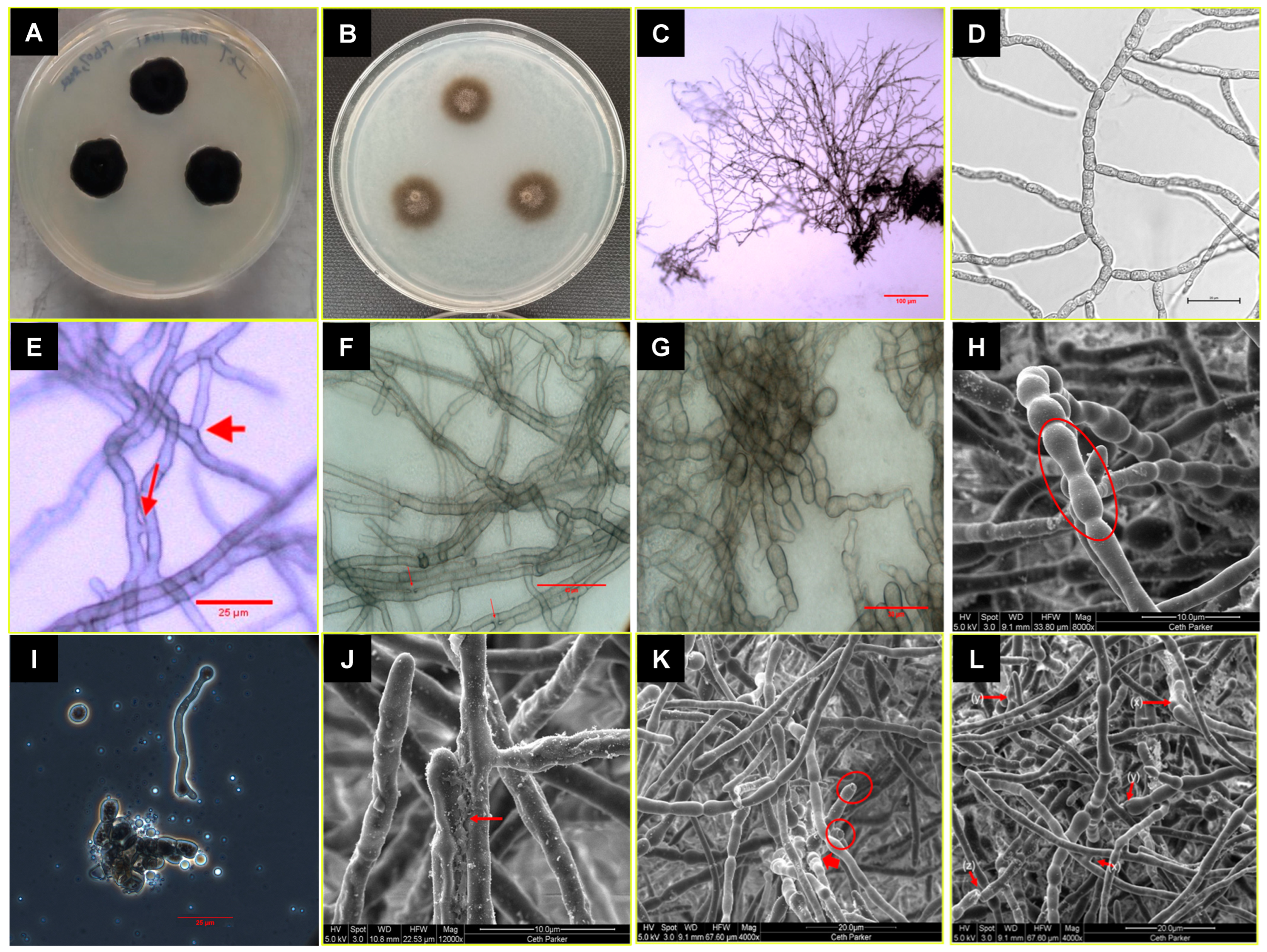
2.2. Morphological Analysis
2.3. Scanning Electron Microscopy
2.4. ITS-Based Fungal Identification
2.5. Whole Genome Sequence Generation
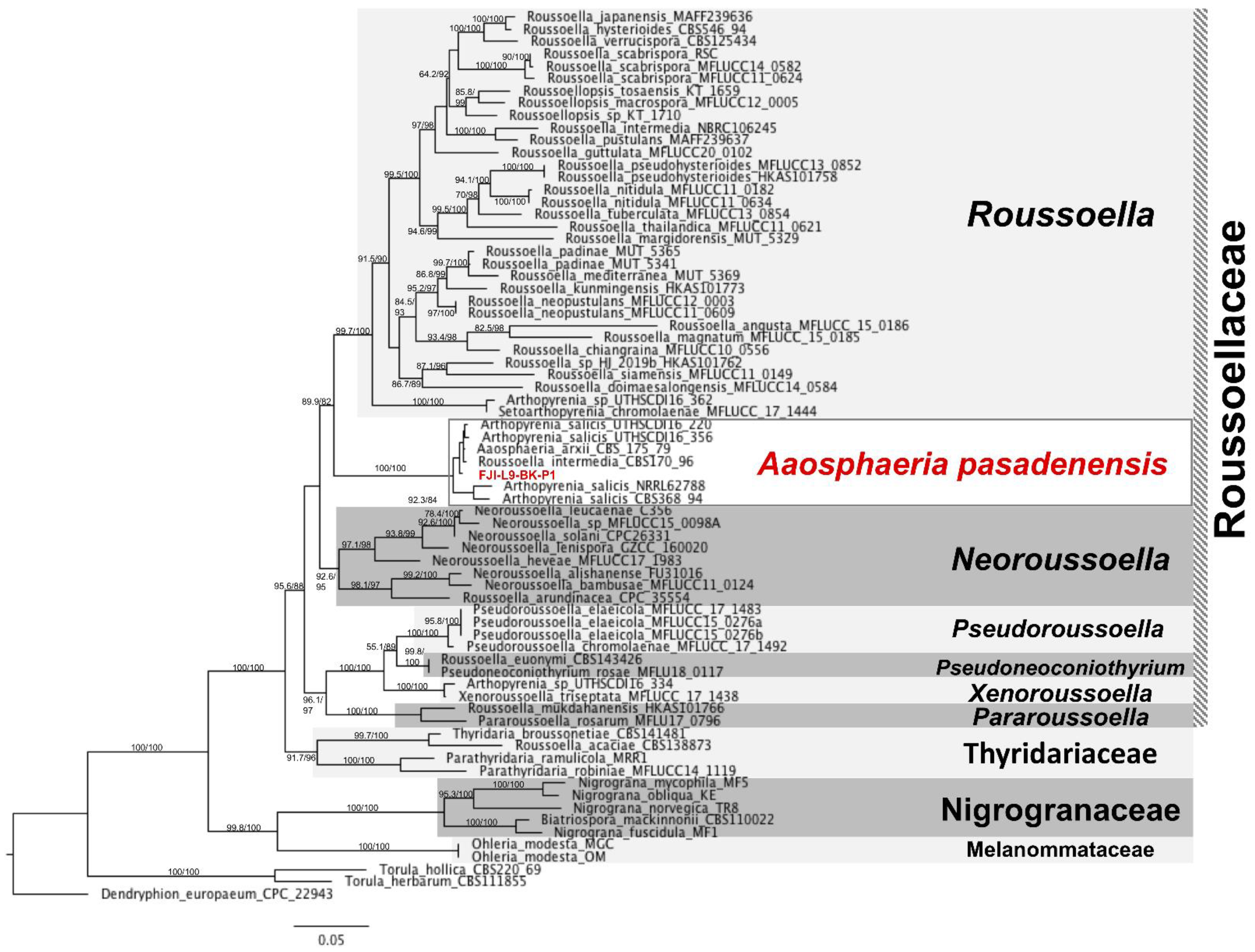
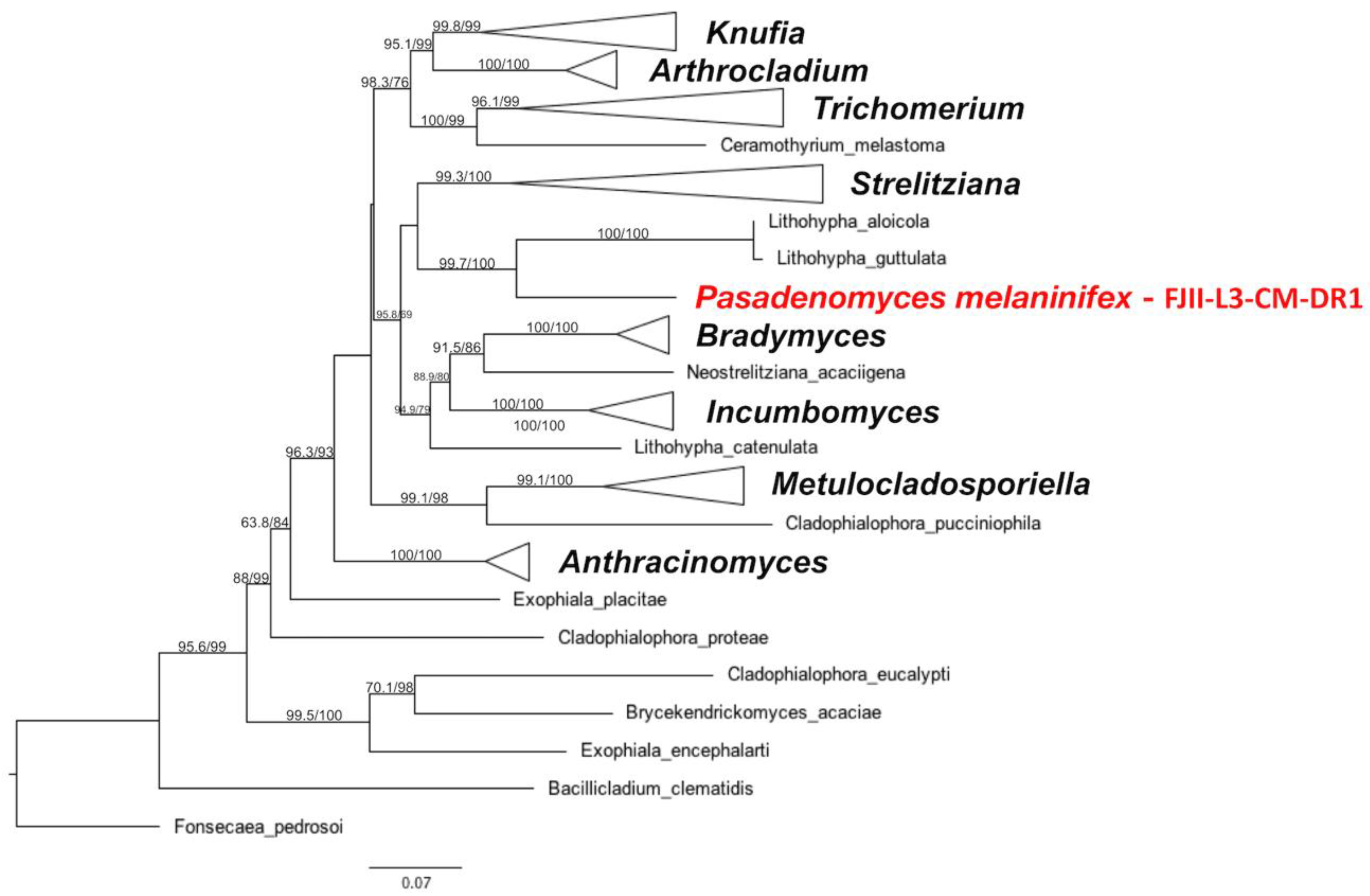
2.6. MLST-Based Phylogenetic Analyses
2.7. Genome Annotation
2.8. Whole Genome-Based Phylogenetic Tree
2.9. UV-C Exposure and Survival Evaluation
3. Results
3.1. Taxonomy and Description of the Strain A. pasadenensis
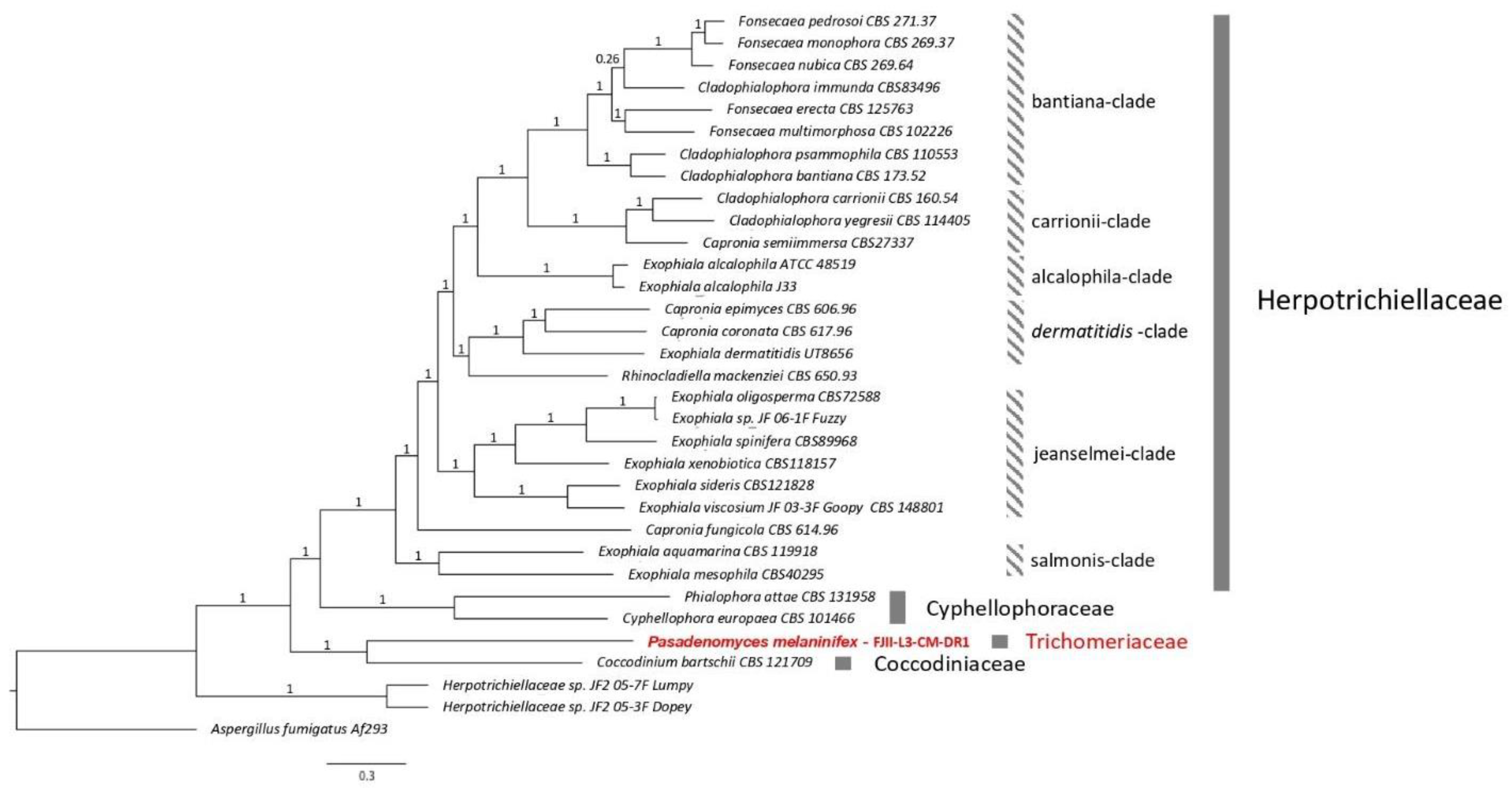
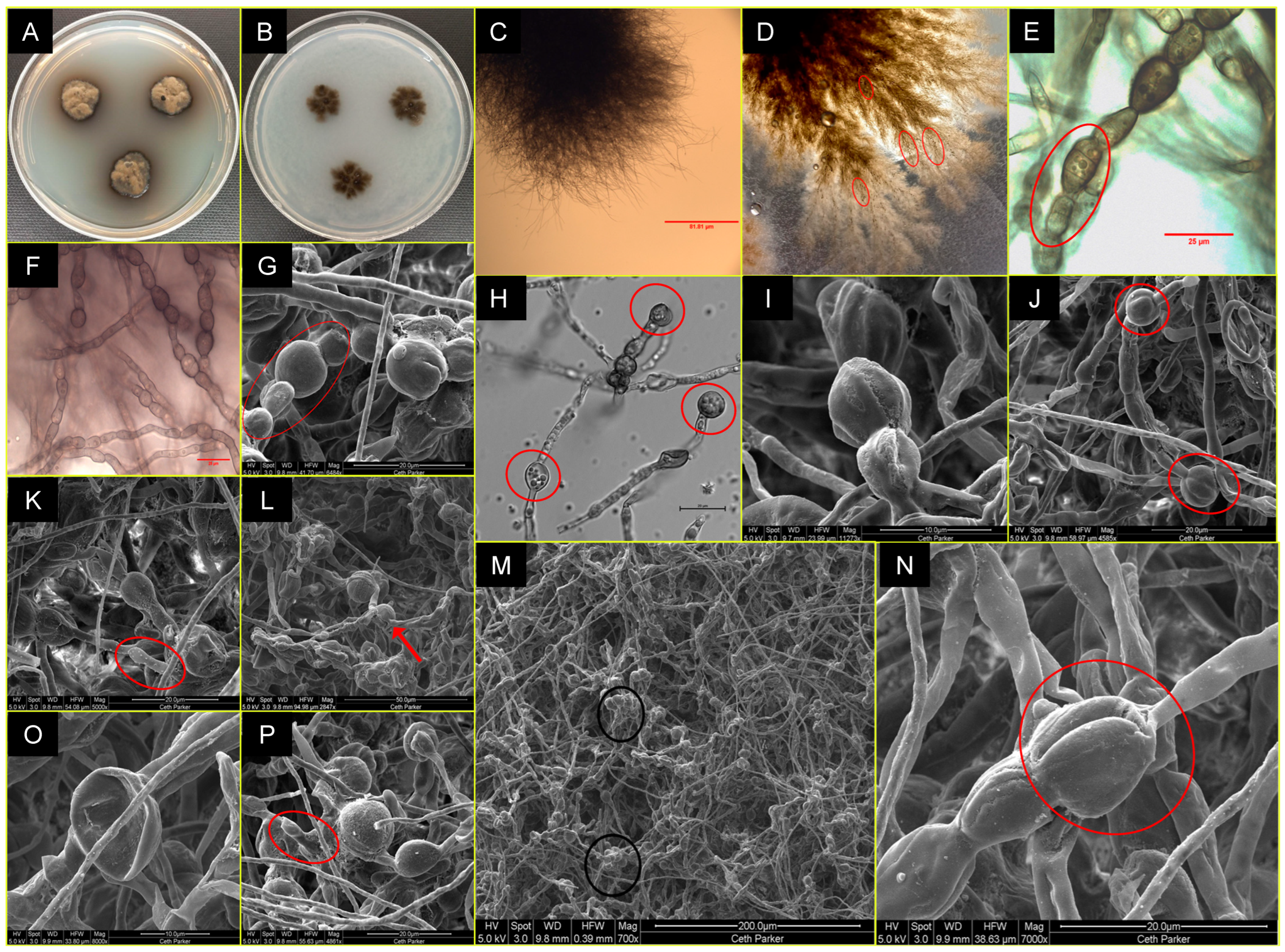
3.2. Taxonomy and Description of the P. melaninifex
3.3. Taxonomy and Characteristics of F. radiotolerans
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- la Duc, M.T.; Nicholson, W.; Kern, R.; Venkateswaran, K. Microbial Characterization of the Mars Odyssey Spacecraft and Its Encapsulation Facility. Environ. Microbiol. 2003, 5, 977–985. [Google Scholar] [CrossRef] [PubMed]
- la Duc, M.T.; Kern, R.; Venkateswaran, K. Microbial Monitoring of Spacecraft and Associated Environments. Microb. Ecol. 2004, 47, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Blachowicz, A.; Mhatre, S.; Singh, N.K.; Wood, J.M.; Parker, C.W.; Ly, C.; Butler, D.; Mason, C.E.; Venkateswaran, K. The Isolation and Characterization of Rare Mycobiome Associated With Spacecraft Assembly Cleanrooms. Front. Microbiol. 2022, 13, 777133. [Google Scholar] [CrossRef]
- la Duc, M.T.; Dekas, A.; Osman, S.; Moissl, C.; Newcombe, D.; Venkateswaran, K. Isolation and Characterization of Bacteria Capable of Tolerating the Extreme Conditions of Clean Room Environments. Appl. Environ. Microbiol. 2007, 73, 2600–2611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blachowicz, A.; Chiang, A.J.; Elsaesser, A.; Kalkum, M.; Ehrenfreund, P.; Stajich, J.E.; Torok, T.; Wang, C.C.C.; Venkateswaran, K. Proteomic and Metabolomic Characteristics of Extremophilic Fungi Under Simulated Mars Conditions. Front. Microbiol. 2019, 10, 1013. [Google Scholar] [CrossRef] [PubMed]
- Dighton, J.; Tugay, T.; Zhdanova, N. Fungi and Ionizing Radiation from Radionuclides. FEMS Microbiol. Lett. 2008, 281, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Onofri, S.; Barreca, D.; Selbmann, L.; Isola, D.; Rabbow, E.; Horneck, G.; de Vera, J.P.P.; Hatton, J.; Zucconi, L. Resistance of Antarctic Black Fungi and Cryptoendolithic Communities to Simulated Space and Martian Conditions. Stud. Mycol. 2008, 61, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Guarro, J.; Gené, J.; Stchigel, A.M. Developments in Fungal Taxonomy. Clin Microbiol Rev. 1999, 12, 454–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, L.G.; Kovács, G.M.; Krizsán, K. Complex Multicellularity in Fungi: Evolutionary Convergence, Single Origin, or Both? Biol. Rev. Camb. Philos. Soc. 2018, 93, 1778–1794. [Google Scholar] [CrossRef]
- Lücking, R.; Aime, M.C.; Robbertse, B.; Miller, A.N.; Ariyawansa, H.A.; Aoki, T.; Cardinali, G.; Crous, P.W.; Druzhinina, I.S.; Geiser, D.M.; et al. Unambiguous Identification of Fungi: Where Do We Stand and How Accurate and Precise Is Fungal DNA Barcoding? IMA Fungus 2020 11:1 2020, 11, 14. [Google Scholar] [CrossRef]
- Wertheimer, N.B.; Stone, N.; Berman, J. Ploidy Dynamics and Evolvability in Fungi. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150461. [Google Scholar] [CrossRef] [Green Version]
- Marcet-Houben, M.; Gabaldón, T. Evolutionary and Functional Patterns of Shared Gene Neighbourhood in Fungi. Nat. Microbiol. 2019, 4, 2383–2392. [Google Scholar] [CrossRef] [PubMed]
- Houbraken, J.; Visagie, C.M.; Frisvad, J.C. Recommendations To Prevent Taxonomic Misidentification of Genome-Sequenced Fungal Strains. Microbiol. Resour. Announc. 2021, 10, e01074-20. [Google Scholar] [CrossRef]
- Naranjo-Ortiz, M.A.; Gabaldón, T. Fungal Evolution: Cellular, Genomic and Metabolic Complexity. Biol. Rev. Camb. Philos. Soc. 2020, 95, 1198–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keepers, K.G.; Pogoda, C.S.; White, K.H.; Anderson Stewart, C.R.; Hoffman, J.R.; Ruiz, A.M.; McCain, C.M.; Lendemer, J.C.; Kane, N.C.; Tripp, E.A. Whole Genome Shotgun Sequencing Detects Greater Lichen Fungal Diversity Than Amplicon-Based Methods in Environmental Samples. Front. Ecol. Evol. 2019, 7, 484. [Google Scholar] [CrossRef] [Green Version]
- Chow, N.A.; Muñoz, J.F.; Gade, L.; Berkow, E.L.; Li, X.; Welsh, R.M.; Forsberg, K.; Lockhart, S.R.; Adam, R.; Alanio, A.; et al. Tracing the Evolutionary History and Global Expansion of Candida Auris Using Population Genomic Analyses. mBio 2020, 11, e03364-19. [Google Scholar] [CrossRef]
- Welsh, R.M.; Misas, E.; Forsberg, K.; Lyman, M.; Chow, N.A. Candida Auris Whole-Genome Sequence Benchmark Dataset for Phylogenomic Pipelines. J. Fungi 2021, 7, 214. [Google Scholar] [CrossRef]
- Parker, C.W.; Teixeira, M.D.M.; Singh, N.K.; Raja, H.A.; Cank, K.B.; Spigolon, G.; Oberlies, N.H.; Barker, B.M.; Stajich, J.E.; Mason, C.E.; et al. Genomic Characterization of Parengyodontium Torokii Sp. Nov., a Biofilm-Forming Fungus Isolated from Mars 2020 Assembly Facility. J. Fungi 2022, 8, 66. [Google Scholar] [CrossRef]
- Checinska Sielaff, A.; Urbaniak, C.; Mohan, G.B.M.; Stepanov, V.G.; Tran, Q.; Wood, J.M.; Minich, J.; McDonald, D.; Mayer, T.; Knight, R.; et al. Characterization of the Total and Viable Bacterial and Fungal Communities Associated with the International Space Station Surfaces. Microbiome 2019, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Riddell, R.W. Permanent Stained Mycological Preparations Obtained by Slide Culture. Mycologia 2018, 42, 265–270. [Google Scholar] [CrossRef]
- Lai, X.; Cao, L.; Tan, H.; Fang, S.; Huang, Y.; Zhou, S. Fungal Communities from Methane Hydrate-Bearing Deep-Sea Marine Sediments in South China Sea. ISME J. 2007, 1, 756–762. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.L.; Bruns, T.D. Community Structure of Ectomycorrhizal Fungi in a Pinus Muricata Forest: Minimal Overlap between the Mature Forest and Resistant Propagule Communities. Mol. Ecol. 1999, 8, 1837–1850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blachowicz, A.; Mayer, T.; Bashir, M.; Pieber, T.R.; de León, P.; Venkateswaran, K. Human Presence Impacts Fungal Diversity of Inflated Lunar/Mars Analog Habitat. Microbiome 2017, 5, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, R.K.; Jain, M. NGS QC Toolkit: A Toolkit for Quality Control of next Generation Sequencing Data. PLoS ONE 2012, 7, e30619. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stajich, J.; Palmer, J. Stajichlab/AAFTF: V0.2.3 Release With. 2019. Available online: https://doi.org/10.5281/ZENODO.3437300 (accessed on 20 June 2022).
- BBMap: A Fast, Accurate, Splice-Aware Aligner (Conference)|OSTI.GOV. Available online: https://www.osti.gov/biblio/1241166 (accessed on 2 September 2022).
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A Better Web Interface. Nucleic Acids Res. 2008, 36, W5. [Google Scholar] [CrossRef]
- Li, H. Minimap2: Pairwise Alignment for Nucleotide Sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An Integrated Tool for Comprehensive Microbial Variant Detection and Genome Assembly Improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [Green Version]
- Manchanda, N.; Portwood, J.L.; Woodhouse, M.R.; Seetharam, A.S.; Lawrence-Dill, C.J.; Andorf, C.M.; Hufford, M.B. GenomeQC: A Quality Assessment Tool for Genome Assemblies and Gene Structure Annotations. BMC Genomics 2020, 21, 193. [Google Scholar] [CrossRef] [PubMed]
- Kõljalg, U.; Nilsson, H.R.; Schigel, D.; Tedersoo, L.; Larsson, K.H.; May, T.W.; Taylor, A.F.S.; Jeppesen, T.S.; Frøslev, T.G.; Lindahl, B.D.; et al. The Taxon Hypothesis Paradigm—On the Unambiguous Detection and Communication of Taxa. Microorganisms 2020, 8, 1910. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; White, J.B.; Miadlikowska, J.; Arnold, A.E.; Miller, M.A.; Magain, N.; U’Ren, J.M.; Lutzoni, F. T-BAS Version 2.1: Tree-Based Alignment Selector Toolkit for Evolutionary Placement of DNA Sequences and Viewing Alignments and Specimen Metadata on Curated and Custom Trees. Microbiol. Resour. Announc. 2019, 8, e00328-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirarab, S.; Nguyen, N.; Warnow, T. PASTA: Ultra-Large Multiple Sequence Alignment. In Lecture Notes in Computer Science (including Subseries Lecture Notes in Artificial Intelligence and Lecture Notes in Bioinformatics); Springer: Cham, Switzherland, 2014; pp. 177–191. [Google Scholar] [CrossRef]
- Steenwyk, J.L.; Buida, T.J.; Li, Y.; Shen, X.X.; Rokas, A. ClipKIT: A Multiple Sequence Alignment Trimming Software for Accurate Phylogenomic Inference. PLoS Biol. 2020, 18, e3001007. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R.; Teeling, E. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anisimova, M.; Gil, M.; Dufayard, J.F.; Dessimoz, C.; Gascuel, O. Survey of Branch Support Methods Demonstrates Accuracy, Power, and Robustness of Fast Likelihood-Based Approximation Schemes. Syst. Biol. 2011, 60, 685. [Google Scholar] [CrossRef] [Green Version]
- Palmer, J.M.; Stajich, J. Funannotate v1.8.1: Eukaryotic Genome Annotation. 2020. Available online: https://doi.org/10.5281/ZENODO.4054262 (accessed on 15 June 2022).
- Frith, M.C. A New Repeat-Masking Method Enables Specific Detection of Homologous Sequences. Nucleic Acids Res. 2011, 39, e23. [Google Scholar] [CrossRef] [Green Version]
- Seppey, M.; Manni, M.; Zdobnov, E.M. BUSCO: Assessing Genome Assembly and Annotation Completeness. Methods Mol. Biol. 2019, 1962, 227–245. [Google Scholar] [CrossRef]
- Delcher, A.L.; Harmon, D.; Kasif, S.; White, O.; Salzberg, S.L. Improved Microbial Gene Identification with GLIMMER. Nucleic Acids Res. 1999, 27, 4636–4641. [Google Scholar] [CrossRef]
- Korf, I. Gene Finding in Novel Genomes. BMC Bioinform. 2004, 5, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, B.J.; Salzberg, S.L.; Zhu, W.; Pertea, M.; Allen, J.E.; Orvis, J.; White, O.; Robin, C.R.; Wortman, J.R. Automated Eukaryotic Gene Structure Annotation Using EVidenceModeler and the Program to Assemble Spliced Alignments. Genome Biol. 2008, 9, R7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, P.P.; Lowe, T.M. TRNAscan-SE: Searching for TRNA Genes in Genomic Sequences. Methods Mol. Biol. 2019, 1962, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Blum, M.; Chang, H.Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro Protein Families and Domains Database: 20 Years On. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef]
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernández-Plaza, A.; Forslund, S.K.; Cook, H.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J.; et al. EggNOG 5.0: A Hierarchical, Functionally and Phylogenetically Annotated Orthology Resource Based on 5090 Organisms and 2502 Viruses. Nucleic Acids Res. 2019, 47, D309–D314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The Protein Families Database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Drula, E.; Garron, M.L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The Carbohydrate-Active Enzyme Database: Functions and Literature. Nucleic Acids Res. 2022, 50, D571–D577. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J.; Thomas, P.D.; Huang, X.; Bateman, A.; Finn, R.D. The MEROPS Database of Proteolytic Enzymes, Their Substrates and Inhibitors in 2017 and a Comparison with Peptidases in the PANTHER Database. Nucleic Acids Res. 2018, 46, D624–D632. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Park, J.; Kim, D.; Jung, K.; Kang, S.; Lee, Y.H. Fungal Secretome Database: Integrated Platform for Annotation of Fungal Secretomes. BMC Genomics 2010, 11, 105. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. AntiSMASH 6.0: Improving Cluster Detection and Comparison Capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- GitHub. Stajichlab/PHYling_unified: Unified PHYling Pipeline for Species Tree Building from Annotated Genomes. Available online: Https://Github.Com/Stajichlab/AAFTF; Https://Github.Com/Nextgenusfs/Funannotate; https://github.com/stajichlab/PHYling_unified (accessed on 18 September 2022). (for Assembly and Annotation Steps).
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F.; et al. MycoCosm Portal: Gearing up for 1000 Fungal Genomes. Nucleic Acids Res. 2014, 42, D699–D704. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing Large Minimum Evolution Trees with Profiles Instead of a Distance Matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef] [PubMed]
- Mirarab, S.; Reaz, R.; Bayzid, M.S.; Zimmermann, T.; Swenson, M.S.; Warnow, T. ASTRAL: Genome-Scale Coalescent-Based Species Tree Estimation. Bioinformatics 2014, 30, i541–i548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blachowicz, A.; Raffa, N.; Bok, J.W.; Choera, T.; Knox, B.; Lim, F.Y.; Huttenlocher, A.; Wang, C.C.C.; Venkateswaran, K.; Keller, N.P. Contributions of Spore Secondary Metabolites to UV-C Protection and Virulence Vary in Different Aspergillus Fumigatus Strains. mBio 2020, 11, e03415-19. [Google Scholar] [CrossRef] [Green Version]
- Osman, S.; Peeters, Z.; la Duc, M.T.; Mancinelli, R.; Ehrenfreund, P.; Venkateswaran, K. Effect of Shadowing on Survival of Bacteria under Conditions Simulating the Martian Atmosphere and UV Radiation. Appl. Environ. Microbiol. 2008, 74, 959–970. [Google Scholar] [CrossRef] [Green Version]
- Brancini, G.T.P.; Rangel, D.E.N.; Braga, G.Ú.L. Exposure of Metarhizium Acridum Mycelium to Light Induces Tolerance to UV-B Radiation. FEMS Microbiol. Lett. 2016, 363, fnw036. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.H.; Lin, C.H.; Chung, K.R. A Nonribosomal Peptide Synthetase Mediates Siderophore Production and Virulence in the Citrus Fungal Pathogen Alternaria Alternata. Mol. Plant Pathol. 2013, 14, 497. [Google Scholar] [CrossRef]
- Godio, R.P.; Martín, J.F. Modified Oxidosqualene Cyclases in the Formation of Bioactive Secondary Metabolites: Biosynthesis of the Antitumor Clavaric Acid. Fungal. Genet. Biol. 2009, 46, 232–242. [Google Scholar] [CrossRef]
- Pacelli, C.; Bryan, R.A.; Onofri, S.; Selbmann, L.; Shuryak, I.; Dadachova, E. Melanin Is Effective in Protecting Fast and Slow Growing Fungi from Various Types of Ionizing Radiation. Environ. Microbiol. 2017, 19, 1612–1624. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Becerra, A.G.; Schnittler, M.; Shchepin, O.N.; Novozhilov, Y.K.; Silva-Filho, A.G.S.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Thiyagaraja, V.; Lücking, R.; Ertz, D.; Coppins, B.J.; Wanasinghe, D.N.; Karunarathna, S.C.; Suwannarach, N.; To-Anun, C.; Cheewangkoon, R.; Hyde, K.D. Sequencing of the Type Species of Arthopyrenia Places Arthopyreniaceae as a Synonym of Trypetheliaceae. Mycosphere 2020. [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal Diversity Notes 367–490: Taxonomic and Phylogenetic Contributions to Fungal Taxa. Fungal Divers 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Dothideomycetes. Available online: https://www.dothideomycetes.org/pleosporales/dacampiaceae/aaosphaeria/aaosphaeria-arxii.html (accessed on 2 September 2022).
- Riko, R.; Nakamura, H.; Shindo, K. Studies on Pyranonigrins-Isolation of Pyranonigrin E and Biosynthetic Studies on Pyranonigrin A. J. Antibiot. 2014, 67, 179–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, L.; Tu, X.; He, K.; Wang, C.; Yin, S.; Zhou, Y.; Chen, W. Monascorubrin and rubropunctatin: Preparation and reaction characteristics with amines. Dye. Pigment. 2019, 170, 107629. [Google Scholar] [CrossRef]
- Nakamoto, S. Promotion of chlamydoconidium formation in Candida albicans by corn meal broth incubation. Med. Mycol. 1998, 36, 123–125. [Google Scholar] [CrossRef] [PubMed]
- Chander, A.M.; Singh, N.K.; Simpson, A.C.; Seuylemezian, A.; Mason, C.E.; Venkateswaran, K.; Stajich, J.E. Draft Genome Sequences of Fungi Isolated from Mars 2020 Spacecraft Assembly Facilities. Microbiol. Resour. Announc. 2022, 11, e00464-22. [Google Scholar] [CrossRef]
- Chang, P.K. Authentication of Aspergillus Parasiticus Strains in the Genome Database of the National Center for Biotechnology Information. BMC Res. Notes 2021, 14, 111. [Google Scholar] [CrossRef]
- Selbmann, L.; de Hoog, G.S.; Zucconi, L.; Isola, D.; Ruisi, S.; Gerrits van den Ende, A.H.G.; Ruibal, C.; de Leo, F.; Urzì, C.; Onofri, S. Drought Meets Acid: Three New Genera in a Dothidealean Clade of Extremotolerant Fungi. Stud. Mycol. 2008, 61, 1. [Google Scholar] [CrossRef]
- Mugambi, G.K.; Huhndorf, S.M. Molecular Phylogenetics of Pleosporales: Melanommataceae and Lophiostomataceae Re-Circumscribed (Pleosporomycetidae, Dothideomycetes, Ascomycota). Stud. Mycol. 2009, 64, 103–121. [Google Scholar] [CrossRef]
- Houbraken, J.; Kocsubé, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.A.; et al. Classification of Aspergillus, Penicillium, Talaromyces and Related Genera (Eurotiales): An Overview of Families, Genera, Subgenera, Sections, Series and Species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef]
- Hubka, V.; Barrs, V.; Dudová, Z.; Sklenář, F.; Kubátová, A.; Matsuzawa, T.; Yaguchi, T.; Horie, Y.; Nováková, A.; Frisvad, J.C.; et al. Unravelling Species Boundaries in the Aspergillus Viridinutans Complex (Section Fumigati): Opportunistic Human and Animal Pathogens Capable of Interspecific Hybridization. Persoonia 2018, 41, 142–174. [Google Scholar] [CrossRef] [PubMed]
- Ciufo, S.; Kannan, S.; Sharma, S.; Badretdin, A.; Clark, K.; Turner, S.; Brover, S.; Schoch, C.L.; Kimchi, A.; DiCuccio, M. Using Average Nucleotide Identity to Improve Taxonomic Assignments in Prokaryotic Genomes at the NCBI. Int. J. Syst. Evol. Microbiol. 2018, 68, 2386–2392. [Google Scholar] [CrossRef] [PubMed]
- Boehm, E.W.A.; Mugambi, G.K.; Miller, A.N.; Huhndorf, S.M.; Marincowitz, S.; Spatafora, J.W.; Schoch, C.L. A Molecular Phylogenetic Reappraisal of the Hysteriaceae, Mytilinidiaceae and Gloniaceae (Pleosporomycetidae, Dothideomycetes) with Keys to World Species. Stud. Mycol. 2009, 64, 49–83. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Crous, P.W.; Groenewald, J.Z.; Boehm, E.W.A.; Burgess, T.I.; de Gruyter, J.; de Hoog, G.S.; Dixon, L.J.; Grube, M.; Gueidan, C.; et al. A Class-Wide Phylogenetic Assessment of Dothideomycetes. Stud. Mycol. 2009, 64, 1. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.K.; Phookamsak, R.; Dai, D.Q.; Tanaka, K.; Jones, E.B.G.; Xu, J.C.; Chukeatirote, E.; Hyde, K.D. Roussoellaceae, a New Pleosporalean Family to Accommodate the Genera Neoroussoella Gen. Nov., Roussoella and Roussoellopsis. Phytotaxa 2014, 181, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Pažoutová, S.; Šrůtka, P.; Holuša, J.; Chudíčková, M.; Kubátová, A.; Kolařík, M. Liberomyces Gen. Nov. with Two New Species of Endophytic Coelomycetes from Broadleaf Trees. Mycologia 2017, 104, 198–210. [Google Scholar] [CrossRef]
- Quan, Y.; Ahmed, S.A.; Menezes da Silva, N.; Al-Hatmi, A.M.S.; Mayer, V.E.; Deng, S.; Kang, Y.; Sybren de Hoog, G.; Shi, D. Novel Black Yeast-like Species in Chaetothyriales with Ant-Associated Life Styles. Fungal. Biol. 2021, 125, 276–284. [Google Scholar] [CrossRef]
- Sun, W.; Su, L.; Yang, S.; Sun, J.; Liu, B.; Fu, R.; Wu, B.; Liu, X.; Cai, L.; Guo, L.; et al. Unveiling the Hidden Diversity of Rock-Inhabiting Fungi: Chaetothyriales from China. J. Fungi 2020, 6, 187. [Google Scholar] [CrossRef]
- Su, H.-Y.; Df, B.; Zl, L.; Jk, L.; Dj, B.; Li, W.L.; Hy, S.; Kd, H. Lignicolous Freshwater Fungi in China III: Three New Species and a New Record of Kirschsteiniothelia from Northwestern Yunnan Province. Mycosphere. 2018, 755–768. [Google Scholar] [CrossRef]
- Jennings, Z.; Kable, K.; Halliday, C.L.; Nankivell, B.J.; Kok, J.; Wong, G.; Chen, S.C.A. Verruconis Gallopava Cardiac and Endovascular Infection with Dissemination after Renal Transplantation: Case Report and Lessons Learned. Med. Mycol. Case Rep. 2016, 15, 5–8. [Google Scholar] [CrossRef]
- Aptroot, A.; Lücking, R. A Revisionary Synopsis of the Trypetheliaceae (Ascomycota: Trypetheliales). Lichenologist 2016, 48, 763–982. [Google Scholar] [CrossRef] [Green Version]
- Aptroot, A.; Sipman, H.J.M.; Barreto, F.M.O.; Nunes, A.D.; da Silva Cáceres, M.E. Ten New Species and 34 New Country Records of Trypetheliaceae. Lichenologist 2019, 51, 27–43. [Google Scholar] [CrossRef]
- da Silva Cáceres, M.E.; Aptroot, A. Lichens from the Brazilian Amazon, with Special Reference to the Genus Astrothelium. Bryologist 2017, 120, 166–182. [Google Scholar] [CrossRef]
- Kocourková, J.; Knudsen, K. Lichenological Notes 2: Lichenothelia Convexa, a Poorly Known Rock-Inhabiting and Lichenicolous Fungus. Mycotaxon 2011, 115, 345–351. [Google Scholar] [CrossRef]
- Muggia, L.; Kocourková, J.; Knudsen, K. Disentangling the Complex of Lichenothelia Species from Rock Communities in the Desert. Mycologia 2015, 107, 1233–1253. [Google Scholar] [CrossRef] [Green Version]
- Selbmann, L.; Isola, D.; Egidi, E.; Zucconi, L.; Gueidan, C.; de Hoog, G.S.; Onofri, S. Mountain Tips as Reservoirs for New Rock-Fungal Entities: Saxomyces Gen. Nov. and Four New Species from the Alps. Fungal Diversity 2013, 65, 167–182. [Google Scholar] [CrossRef] [Green Version]
- Freitas, D.F.; da Rocha, I.M.; Vieira-da-Motta, O.; de Paula Santos, C. The Role of Melanin in the Biology and Ecology of Nematophagous Fungi. J. Chem. Ecol. 2021, 47, 597–613. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Novel Fungal Strains | Aaosphaeria pasadenensis (FJI-L9-BK-P1) | Pasadenomyces melaninifex (FJII-L3-CM-DR1) | Floridaphiala radiotolerans (FKI-L1-BK-DR1) |
|---|---|---|---|
| Number of contigs | 122 | 972 | 109 |
| Length | 38,798,968 | 27,602,062 | 24,887,818 |
| N50 | 943,274 | 498,971 | 518,619 |
| L50 | 14 | 17 | 17 |
| N90 | 314,675 | 48,404 | 169,985 |
| L90 | 40 | 74 | 49 |
| GC content (%) | 49.73 | 48.13 | 52.18 |
| Genes | 12,248 | 10,961 | 8582 |
| mRNA | 12,141 | 10,856 | 8505 |
| tRNA | 107 | 105 | 77 |
| Average gene length | 1605.34 | 1536.55 | 1708.37 |
| Total exons | 33,005 | 20,143 | 22,516 |
| Average exon length | 467.7 | 545.7 | 512.5 |
| Average protein length | 492.79 | 488.12 | 526.9 |
| GO terms | 2295 | 2,107 | 2099 |
| Interproscan | 3140 | 2968 | 2911 |
| Eggnog | 11,235 | 9309 | 7651 |
| Pfam | 8359 | 7070 | 5866 |
| CAZYme | 588 | 297 | 266 |
| MEROPS | 395 | 346 | 257 |
| BUSCO | 3682 | 2928 | 3412 |
| Secretion | 1204 | 604 | 578 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chander, A.M.; Teixeira, M.d.M.; Singh, N.K.; Williams, M.P.; Simpson, A.C.; Damle, N.; Parker, C.W.; Stajich, J.E.; Mason, C.E.; Torok, T.; et al. Description and Genome Characterization of Three Novel Fungal Strains Isolated from Mars 2020 Mission-Associated Spacecraft Assembly Facility Surfaces—Recommendations for Two New Genera and One Species. J. Fungi 2023, 9, 31. https://doi.org/10.3390/jof9010031
Chander AM, Teixeira MdM, Singh NK, Williams MP, Simpson AC, Damle N, Parker CW, Stajich JE, Mason CE, Torok T, et al. Description and Genome Characterization of Three Novel Fungal Strains Isolated from Mars 2020 Mission-Associated Spacecraft Assembly Facility Surfaces—Recommendations for Two New Genera and One Species. Journal of Fungi. 2023; 9(1):31. https://doi.org/10.3390/jof9010031
Chicago/Turabian StyleChander, Atul Munish, Marcus de Melo Teixeira, Nitin K. Singh, Michael P. Williams, Anna C. Simpson, Namita Damle, Ceth W. Parker, Jason E. Stajich, Christopher E. Mason, Tamas Torok, and et al. 2023. "Description and Genome Characterization of Three Novel Fungal Strains Isolated from Mars 2020 Mission-Associated Spacecraft Assembly Facility Surfaces—Recommendations for Two New Genera and One Species" Journal of Fungi 9, no. 1: 31. https://doi.org/10.3390/jof9010031