
Complementary Strategies to Unlock Biosynthesis Gene Clusters Encoding Secondary Metabolites in the Filamentous Fungus Podospora anserina

 , , , , , and
, , , , , and 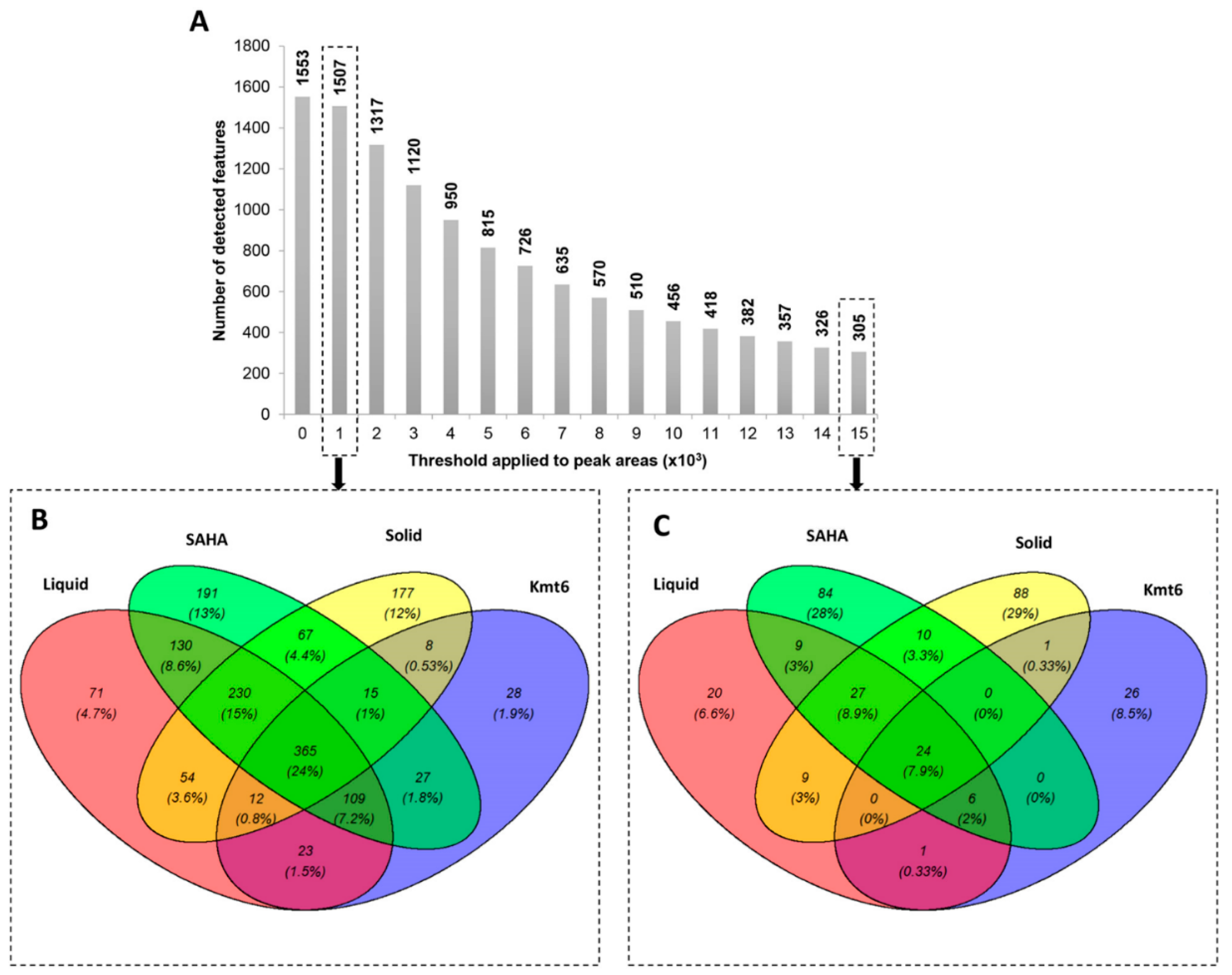{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Growth Conditions
2.2. Fungal Culture Extraction
2.3. LC-MS/MS Analysis
2.4. Data Processing
2.5. Statistical Analysis
2.6. Dereplication
3. Results
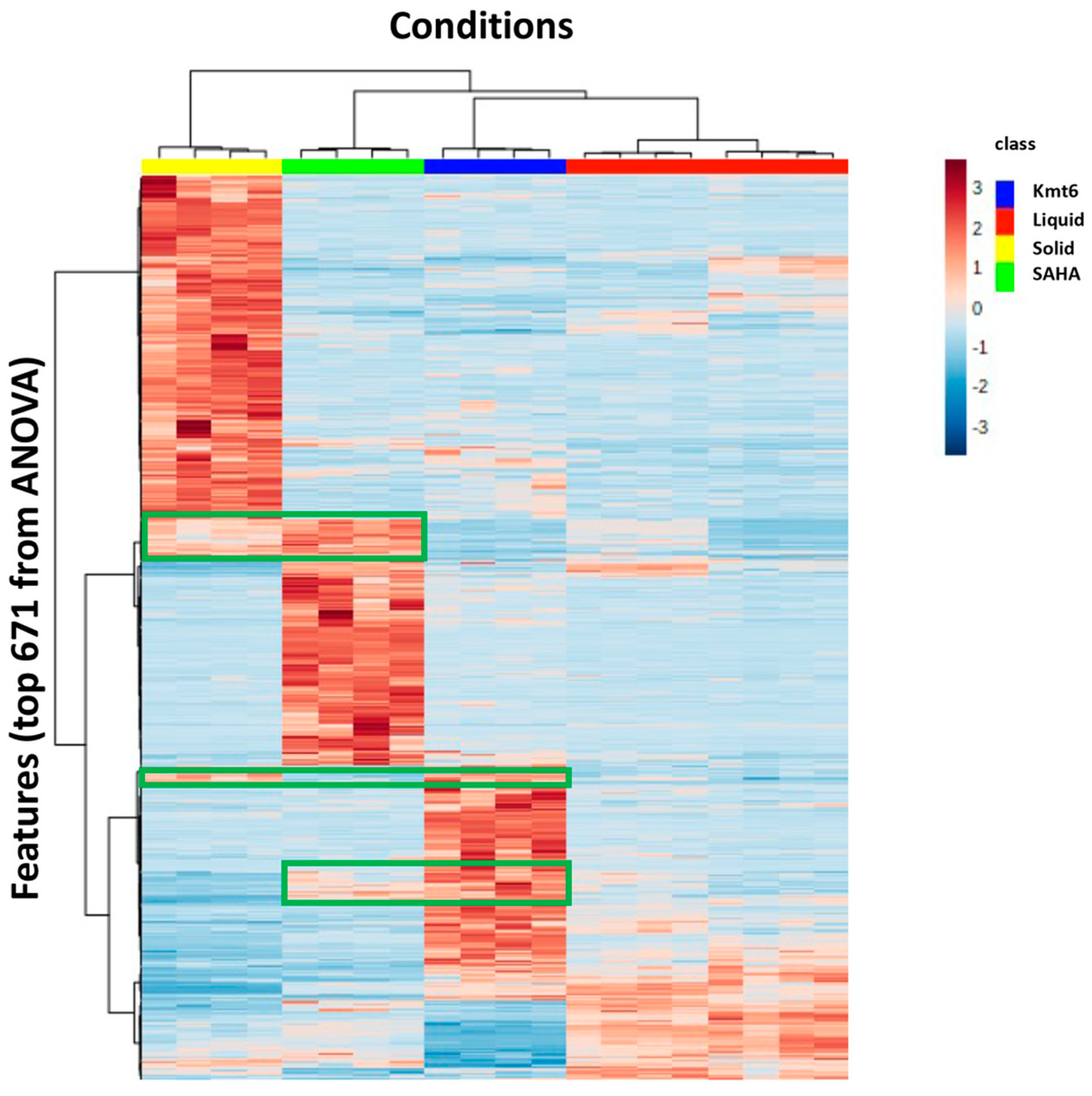
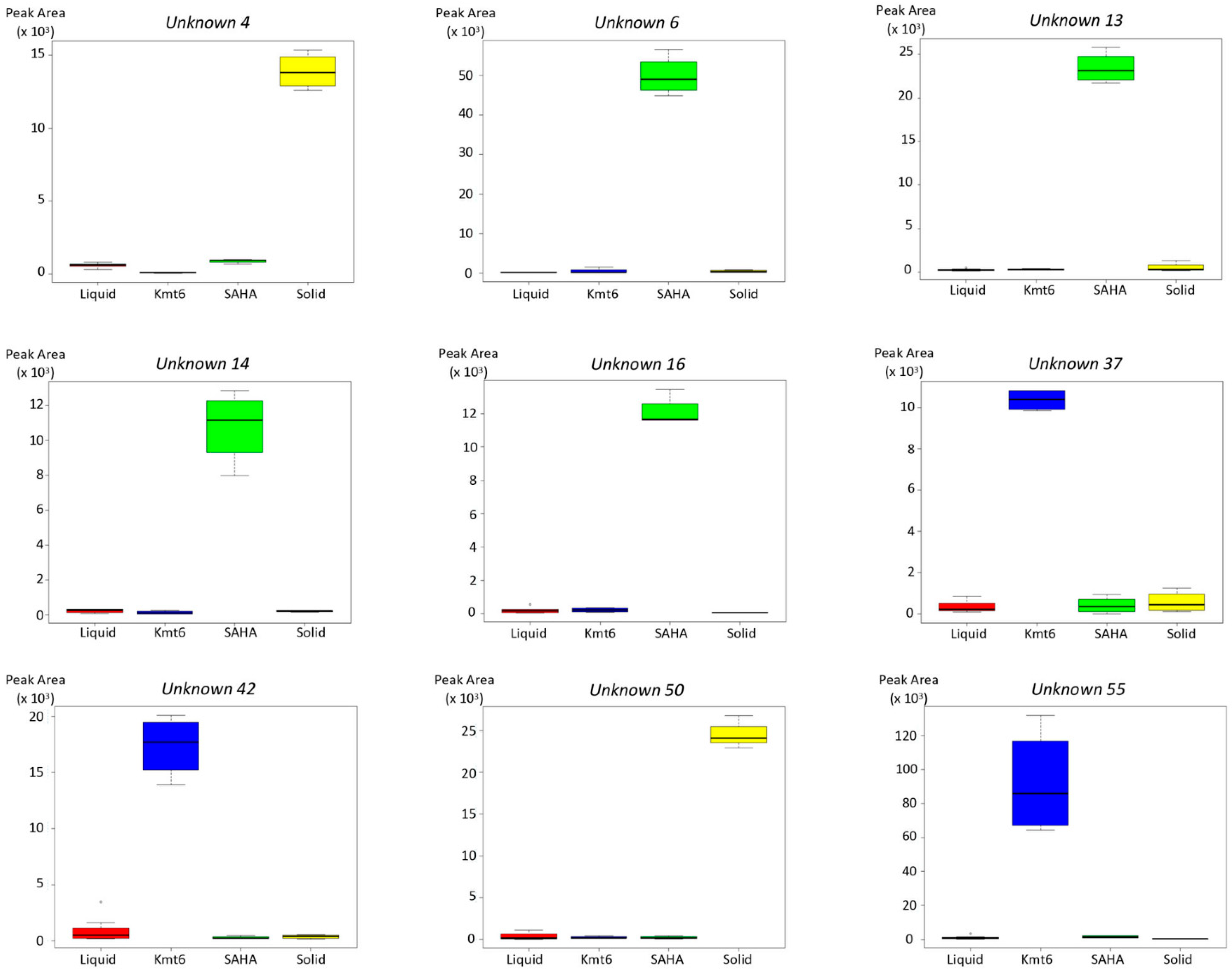
3.1. Metabolomic Study of P. anserina in Different Conditions
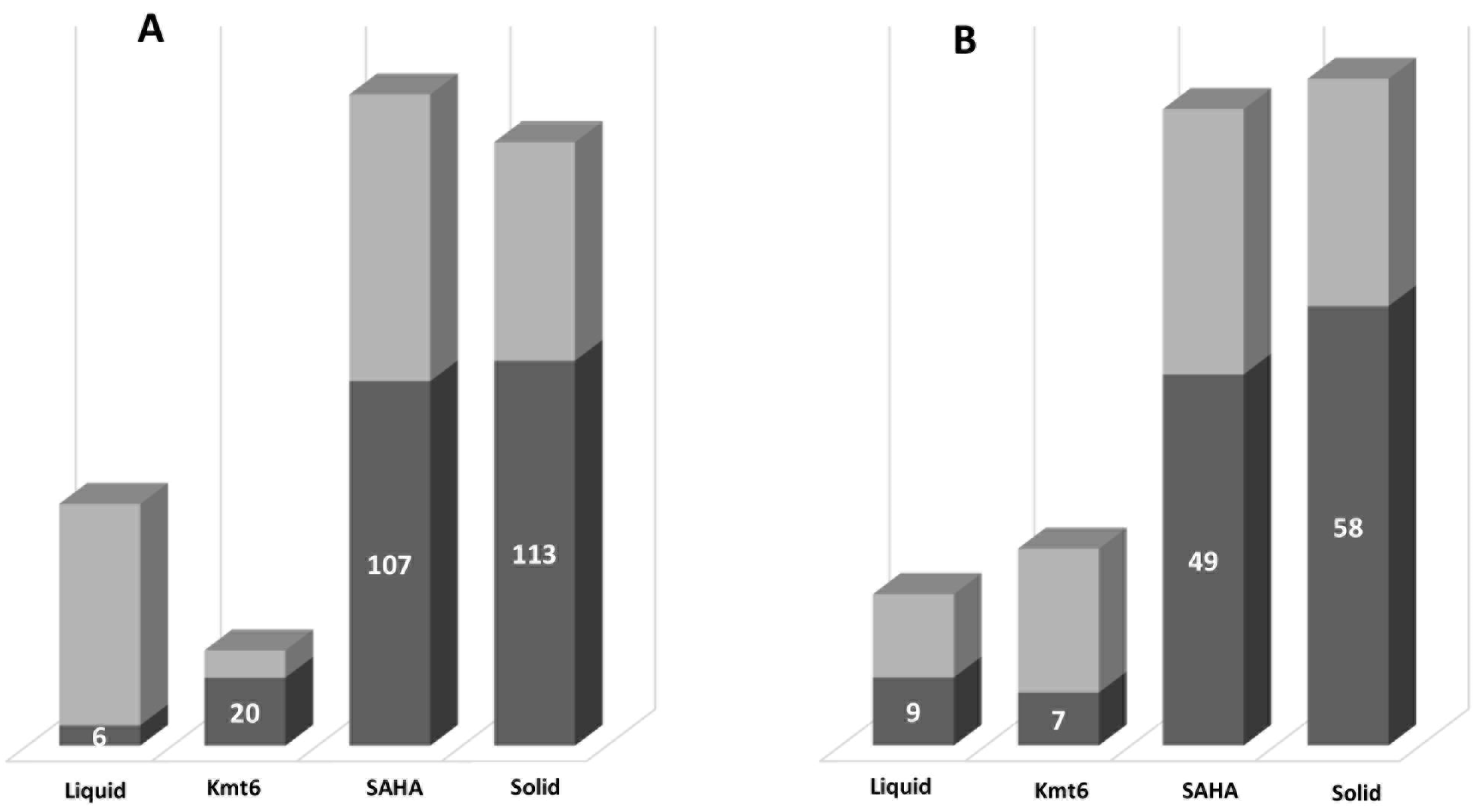
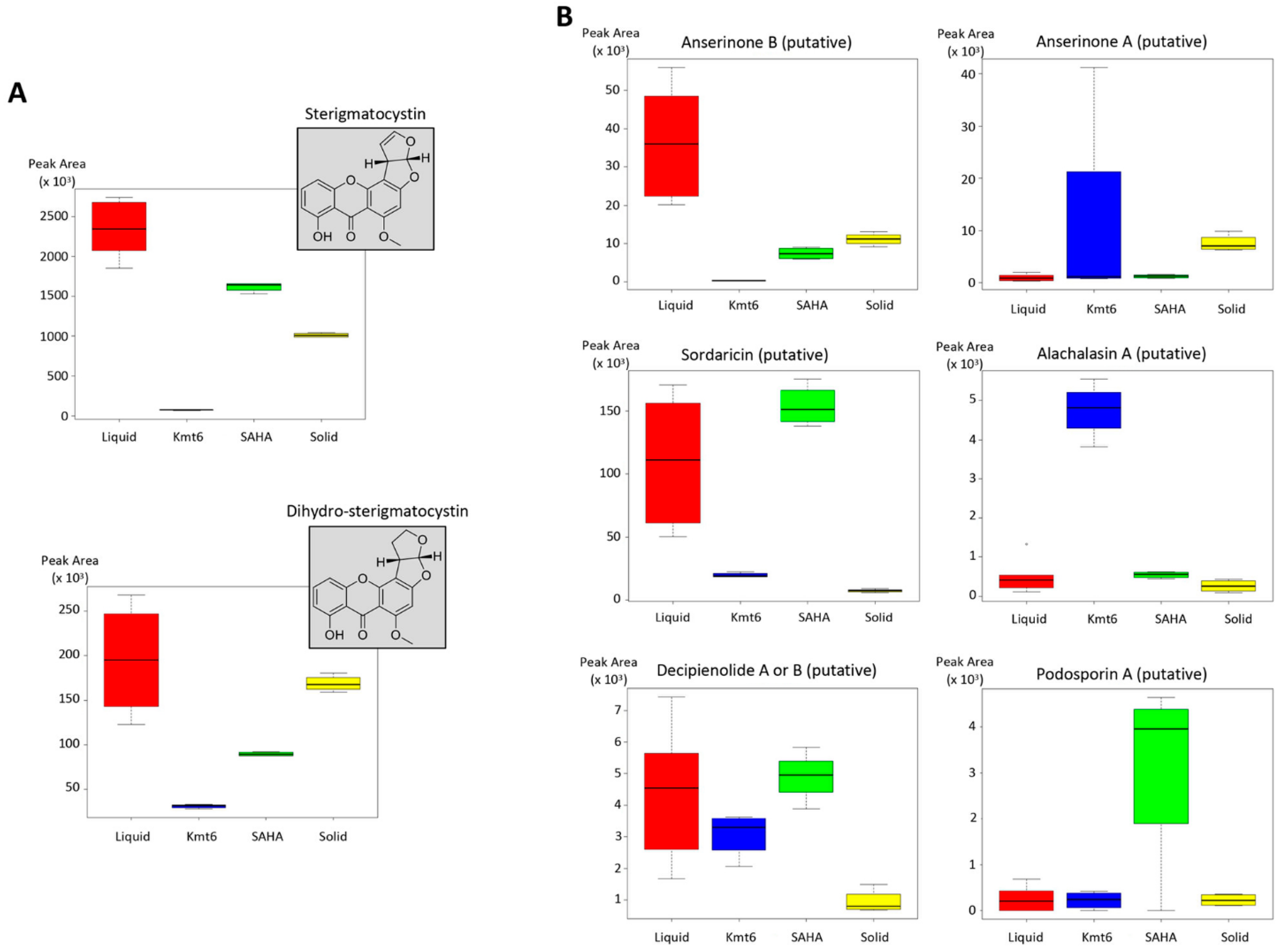
3.2. Identification of Features of Interest
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brakhage, A.A. Regulation of Fungal Secondary Metabolism. Nat. Rev. Microbiol. 2013, 11, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Caesar, L.K.; Kelleher, N.L.; Keller, N.P. In the Fungus Where It Happens: History and Future Propelling Aspergillus nidulans as the Archetype of Natural Products Research. Fungal Genet. Biol. 2020, 144, 103477. [Google Scholar] [CrossRef] [PubMed]
- Keller, N.P. Fungal Secondary Metabolism: Regulation, Function and Drug Discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Kjærbølling, I.; Mortensen, U.H.; Vesth, T.; Andersen, M.R. Strategies to Establish the Link between Biosynthetic Gene Clusters and Secondary Metabolites. Fungal Genet. Biol. 2019, 130, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Wiemann, P.; Keller, N.P. Strategies for Mining Fungal Natural Products. J. Ind. Microbiol. Biotechnol. 2014, 41, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Brakhage, A.A.; Schroeckh, V. Fungal Secondary Metabolites—Strategies to Activate Silent Gene Clusters. Fungal Genet. Biol. 2011, 48, 15–22. [Google Scholar] [CrossRef]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal Secondary Metabolism—From Biochemistry to Genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef]
- Bergmann, S.; Schümann, J.; Scherlach, K.; Lange, C.; Brakhage, A.A.; Hertweck, C. Genomics-Driven Discovery of PKS-NRPS Hybrid Metabolites from Aspergillus nidulans. Nat. Chem. Biol. 2007, 3, 213–217. [Google Scholar] [CrossRef]
- Janevska, S.; Tudzynski, B. Secondary Metabolism in Fusarium fujikuroi: Strategies to Unravel the Function of Biosynthetic Pathways. Appl. Microbiol. Biotechnol. 2018, 102, 615–630. [Google Scholar] [CrossRef]
- Hautbergue, T.; Jamin, E.L.; Debrauwer, L.; Puel, O.; Oswald, I.P. From Genomics to Metabolomics, Moving toward an Integrated Strategy for the Discovery of Fungal Secondary Metabolites. Nat. Prod. Rep. 2018, 35, 147–173. [Google Scholar] [CrossRef]
- Bergmann, S.; Funk, A.N.; Scherlach, K.; Schroeckh, V.; Shelest, E.; Horn, U.; Hertweck, C.; Brakhage, A.A. Activation of a Silent Fungal Polyketide Biosynthesis Pathway through Regulatory Cross Talk with a Cryptic Nonribosomal Peptide Synthetase Gene Cluster. Appl. Environ. Microbiol. 2010, 76, 8143–8149. [Google Scholar] [CrossRef] [Green Version]
- Yeh, H.-H.; Ahuja, M.; Chiang, Y.-M.; Oakley, C.E.; Moore, S.; Yoon, O.; Hajovsky, H.; Bok, J.-W.; Keller, N.P.; Wang, C.C.C.; et al. Resistance Gene-Guided Genome Mining: Serial Promoter Exchanges in Aspergillus nidulans Reveal the Biosynthetic Pathway for Fellutamide B, a Proteasome Inhibitor. ACS Chem. Biol. 2016, 11, 2275–2284. [Google Scholar] [CrossRef]
- Lin, T.-S.; Chen, B.; Chiang, Y.-M.; Wang, C.C.C. Discovery and Elucidation of the Biosynthesis of Aspernidgulenes: Novel Polyenes from Aspergillus nidulans by Using Serial Promoter Replacement. Chembiochem 2019, 20, 329–334. [Google Scholar] [CrossRef]
- Sung, C.T.; Chang, S.-L.; Entwistle, R.; Ahn, G.; Lin, T.-S.; Petrova, V.; Yeh, H.-H.; Praseuth, M.B.; Chiang, Y.-M.; Oakley, B.R.; et al. Overexpression of a Three-Gene Conidial Pigment Biosynthetic Pathway in Aspergillus nidulans Reveals the First NRPS Known to Acetylate Tryptophan. Fungal Genet. Biol. 2017, 101, 1–6. [Google Scholar] [CrossRef]
- Yeh, H.-H.; Chiang, Y.-M.; Entwistle, R.; Ahuja, M.; Lee, K.-H.; Bruno, K.S.; Wu, T.-K.; Oakley, B.R.; Wang, C.C.C. Molecular Genetic Analysis Reveals That a Nonribosomal Peptide Synthetase-like (NRPS-like) Gene in Aspergillus nidulans Is Responsible for Microperfuranone Biosynthesis. Appl. Microbiol. Biotechnol. 2012, 96, 739–748. [Google Scholar] [CrossRef] [Green Version]
- van Dijk, J.W.A.; Wang, C.C.C. Heterologous Expression of Fungal Secondary Metabolite Pathways in the Aspergillus nidulans Host System. Methods Enzymol. 2016, 575, 127–142. [Google Scholar] [CrossRef]
- He, Q.-P.; Zhao, S.; Wang, J.-X.; Li, C.-X.; Yan, Y.-S.; Wang, L.; Liao, L.-S.; Feng, J.-X. Transcription Factor NsdD Regulates the Expression of Genes Involved in Plant Biomass-Degrading Enzymes, Conidiation, and Pigment Biosynthesis in Penicillium oxalicum. Appl. Environ. Microbiol. 2018, 84, e01039-18. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Y.-M.; Yu, R.-L.; Zhu, P. Advances in Targeting and Heterologous Expression of Genes Involved in the Synthesis of Fungal Secondary Metabolites. RSC Adv. 2019, 9, 35124–35134. [Google Scholar] [CrossRef] [Green Version]
- Harvey, C.J.B.; Tang, M.; Schlecht, U.; Horecka, J.; Fischer, C.R.; Lin, H.-C.; Li, J.; Naughton, B.; Cherry, J.; Miranda, M.; et al. HEx: A Heterologous Expression Platform for the Discovery of Fungal Natural Products. Sci. Adv. 2018, 4, eaar5459. [Google Scholar] [CrossRef] [Green Version]
- Chiang, Y.-M.; Oakley, C.E.; Ahuja, M.; Entwistle, R.; Schultz, A.; Chang, S.-L.; Sung, C.T.; Wang, C.C.C.; Oakley, B.R. An Efficient System for Heterologous Expression of Secondary Metabolite Genes in Aspergillus nidulans. J. Am. Chem. Soc. 2013, 135, 7720–7731. [Google Scholar] [CrossRef]
- Yin, W.-B.; Chooi, Y.H.; Smith, A.R.; Cacho, R.A.; Hu, Y.; White, T.C.; Tang, Y. Discovery of Cryptic Polyketide Metabolites from Dermatophytes Using Heterologous Expression in Aspergillus nidulans. ACS Synth. Biol. 2013, 2, 629–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baral, B.; Akhgari, A.; Metsä-Ketelä, M. Activation of Microbial Secondary Metabolic Pathways: Avenues and Challenges. Synth. Syst. Biotechnol. 2018, 3, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Bok, J.W.; Keller, N.P. LaeA, a Regulator of Secondary Metabolism in Aspergillus spp. Eukaryot. Cell 2004, 3, 527–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butchko, R.A.E.; Brown, D.W.; Busman, M.; Tudzynski, B.; Wiemann, P. Lae1 Regulates Expression of Multiple Secondary Metabolite Gene Clusters in Fusarium verticillioides. Fungal Genet. Biol. 2012, 49, 602–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chettri, P.; Bradshaw, R.E. LaeA Negatively Regulates Dothistromin Production in the Pine Needle Pathogen Dothistroma septosporum. Fungal Genet. Biol. 2016, 97, 24–32. [Google Scholar] [CrossRef]
- Grau, M.F.; Entwistle, R.; Oakley, C.E.; Wang, C.C.C.; Oakley, B.R. Overexpression of an LaeA-like Methyltransferase Upregulates Secondary Metabolite Production in Aspergillus nidulans. ACS Chem. Biol. 2019, 14, 1643–1651. [Google Scholar] [CrossRef]
- Yu, J.; Han, H.; Zhang, X.; Ma, C.; Sun, C.; Che, Q.; Gu, Q.; Zhu, T.; Zhang, G.; Li, D. Discovery of Two New Sorbicillinoids by Overexpression of the Global Regulator LaeA in a Marine-Derived Fungus Penicillium dipodomyis YJ-11. Mar. Drugs 2019, 17, 446. [Google Scholar] [CrossRef] [Green Version]
- Oakley, C.E.; Ahuja, M.; Sun, W.-W.; Entwistle, R.; Akashi, T.; Yaegashi, J.; Guo, C.-J.; Cerqueira, G.C.; Russo Wortman, J.; Wang, C.C.C.; et al. Discovery of McrA, a Master Regulator of Aspergillus Secondary Metabolism. Mol. Microbiol. 2017, 103, 347–365. [Google Scholar] [CrossRef] [Green Version]
- Pfannenstiel, B.T.; Keller, N.P. On Top of Biosynthetic Gene Clusters: How Epigenetic Machinery Influences Secondary Metabolism in Fungi. Biotechnol. Adv. 2019, 37, 107345. [Google Scholar] [CrossRef]
- Shwab, E.K.; Bok, J.W.; Tribus, M.; Galehr, J.; Graessle, S.; Keller, N.P. Histone Deacetylase Activity Regulates Chemical Diversity in Aspergillus. Eukaryot. Cell 2007, 6, 1656–1664. [Google Scholar] [CrossRef]
- Gacek-Matthews, A.; Noble, L.M.; Gruber, C.; Berger, H.; Sulyok, M.; Marcos, A.T.; Strauss, J.; Andrianopoulos, A. KdmA, a Histone H3 Demethylase with Bipartite Function, Differentially Regulates Primary and Secondary Metabolism in Aspergillus nidulans. Mol. Microbiol. 2015, 96, 839–860. [Google Scholar] [CrossRef] [Green Version]
- Studt, L.; Rösler, S.M.; Burkhardt, I.; Arndt, B.; Freitag, M.; Humpf, H.-U.; Dickschat, J.S.; Tudzynski, B. Knock-down of the Methyltransferase Kmt6 Relieves H3K27me3 and Results in Induction of Cryptic and Otherwise Silent Secondary Metabolite Gene Clusters in Fusarium fujikuroi. Environ. Microbiol. 2016, 18, 4037–4054. [Google Scholar] [CrossRef] [Green Version]
- Fan, A.; Mi, W.; Liu, Z.; Zeng, G.; Zhang, P.; Hu, Y.; Fang, W.; Yin, W.-B. Deletion of a Histone Acetyltransferase Leads to the Pleiotropic Activation of Natural Products in Metarhizium Robertsii. Org. Lett. 2017, 19, 1686–1689. [Google Scholar] [CrossRef]
- Fisch, K.M.; Gillaspy, A.F.; Gipson, M.; Henrikson, J.C.; Hoover, A.R.; Jackson, L.; Najar, F.Z.; Wägele, H.; Cichewicz, R.H. Chemical Induction of Silent Biosynthetic Pathway Transcription in Aspergillus niger. J. Ind. Microbiol. Biotechnol. 2009, 36, 1199–1213. [Google Scholar] [CrossRef]
- Yakasai, A.A.; Davison, J.; Wasil, Z.; Halo, L.M.; Butts, C.P.; Lazarus, C.M.; Bailey, A.M.; Simpson, T.J.; Cox, R.J. Nongenetic Reprogramming of a Fungal Highly Reducing Polyketide Synthase. J. Am. Chem. Soc. 2011, 133, 10990–10998. [Google Scholar] [CrossRef]
- Chung, Y.-M.; El-Shazly, M.; Chuang, D.-W.; Hwang, T.-L.; Asai, T.; Oshima, Y.; Ashour, M.L.; Wu, Y.-C.; Chang, F.-R. Suberoylanilide Hydroxamic Acid, a Histone Deacetylase Inhibitor, Induces the Production of Anti-Inflammatory Cyclodepsipeptides from Beauveria felina. J. Nat. Prod. 2013, 76, 1260–1266. [Google Scholar] [CrossRef]
- Asai, T.; Morita, S.; Taniguchi, T.; Monde, K.; Oshima, Y. Epigenetic Stimulation of Polyketide Production in Chaetomium cancroideum by an NAD(+)-Dependent HDAC Inhibitor. Org. Biomol. Chem. 2016, 14, 646–651. [Google Scholar] [CrossRef]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big Effects from Small Changes: Possible Ways to Explore Nature’s Chemical Diversity. Chembiochem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Schroeckh, V.; Scherlach, K.; Nützmann, H.-W.; Shelest, E.; Schmidt-Heck, W.; Schuemann, J.; Martin, K.; Hertweck, C.; Brakhage, A.A. Intimate Bacterial-Fungal Interaction Triggers Biosynthesis of Archetypal Polyketides in Aspergillus nidulans. Proc. Natl. Acad. Sci. USA 2009, 106, 14558–14563. [Google Scholar] [CrossRef] [Green Version]
- Jančič, S.; Frisvad, J.C.; Kocev, D.; Gostinčar, C.; Džeroski, S.; Gunde-Cimerman, N. Production of Secondary Metabolites in Extreme Environments: Food- and Airborne Wallemia spp. Produce Toxic Metabolites at Hypersaline Conditions. PLoS ONE 2016, 11, e0169116. [Google Scholar] [CrossRef]
- Vinale, F.; Nicoletti, R.; Borrelli, F.; Mangoni, A.; Parisi, O.A.; Marra, R.; Lombardi, N.; Lacatena, F.; Grauso, L.; Finizio, S.; et al. Co-Culture of Plant Beneficial Microbes as Source of Bioactive Metabolites. Sci. Rep. 2017, 7, 14330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romsdahl, J.; Blachowicz, A.; Chiang, A.J.; Chiang, Y.-M.; Masonjones, S.; Yaegashi, J.; Countryman, S.; Karouia, F.; Kalkum, M.; Stajich, J.E.; et al. International Space Station Conditions Alter Genomics, Proteomics, and Metabolomics in Aspergillus nidulans. Appl. Microbiol. Biotechnol. 2019, 103, 1363–1377. [Google Scholar] [CrossRef] [PubMed]
- Rutledge, P.J.; Challis, G.L. Discovery of Microbial Natural Products by Activation of Silent Biosynthetic Gene Clusters. Nat. Rev. Microbiol. 2015, 13, 509–523. [Google Scholar] [CrossRef]
- Silar, P. Podospora anserina. 2020. Available online: https://hal.archives-ouvertes.fr/hal-02475488 (accessed on 12 February 2020).
- Bills, G.F.; Gloer, J.B.; An, Z. Coprophilous Fungi: Antibiotic Discovery and Functions in an Underexplored Arena of Microbial Defensive Mutualism. Curr. Opin. Microbiol. 2013, 16, 549–565. [Google Scholar] [CrossRef] [PubMed]
- Sarrocco, S. Dung-Inhabiting Fungi: A Potential Reservoir of Novel Secondary Metabolites for the Control of Plant Pathogens. Pest Manag. Sci. 2016, 72, 643–652. [Google Scholar] [CrossRef]
- Espagne, E.; Lespinet, O.; Malagnac, F.; Da Silva, C.; Jaillon, O.; Porcel, B.M.; Couloux, A.; Aury, J.-M.; Ségurens, B.; Poulain, J.; et al. The Genome Sequence of the Model Ascomycete Fungus Podospora anserina. Genome Biol. 2008, 9, R77. [Google Scholar] [CrossRef]
- Wang, H.; Gloer, K.B.; Gloer, J.B.; Scott, J.A.; Malloch, D. Anserinones A and B: New Antifungal and Antibacterial Benzoquinones from the Coprophilous Fungus Podospora anserina. J. Nat. Prod. 1997, 60, 629–631. [Google Scholar] [CrossRef]
- Matasyoh, J.C.; Dittrich, B.; Schueffler, A.; Laatsch, H. Larvicidal Activity of Metabolites from the Endophytic Podospora sp. against the Malaria Vector Anopheles gambiae. Parasitol. Res. 2011, 108, 561–566. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Porée, F.-H.; Gaslonde, T.; Lalucque, H.; Chapeland-Leclerc, F.; Ruprich-Robert, G. Functional Characterization of the Sterigmatocystin Secondary Metabolite Gene Cluster in the Filamentous Fungus Podospora anserina: Involvement in Oxidative Stress Response, Sexual Development, Pigmentation and Interspecific Competitions. Environ. Microbiol. 2019, 21, 3011–3026. [Google Scholar] [CrossRef]
- Shen, L.; Gaslonde, T.; Roullier, C.; Wang, H.; Bodin, J.; Porée, F.-H.; Ruprich-Robert, G.; Chapeland-Leclerc, F. Functional Characterization of the GATA-Type Transcription Factor PaNsdD in the Filamentous Fungus Podospora anserina and Its Interplay with the Sterigmatocystin Pathway. Appl. Environ. Microbiol. 2022, 88, aem0237821. [Google Scholar] [CrossRef]
- Rizet, G. Les Phénomènes de Barrage Chez Podospora anserina. I. Analyse Génétique Des Barrages Entre Souches S and s. Rev. Cytol. Biol. Veg. 1952, 13, 51–92. [Google Scholar]
- Carlier, F.; Li, M.; Maroc, L.; Debuchy, R.; Souaid, C.; Noordermeer, D.; Grognet, P.; Malagnac, F. Loss of EZH2-like or SU(VAR)3-9-like Proteins Causes Simultaneous Perturbations in H3K27 and H3K9 Tri-Methylation and Associated Developmental Defects in the Fungus Podospora anserina. Epigenetics Chromatin 2021, 14, 22. [Google Scholar] [CrossRef]
- Comai, L.; Katz, J.E.; Mallick, P. Proteomics: Methods and Protocols; Humana: Louisville, KY, USA, 2017; ISBN 978-1-4939-6745-2. [Google Scholar]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Oresic, M. MZmine 2: Modular Framework for Processing, Visualizing, and Analyzing Mass Spectrometry-Based Molecular Profile Data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [Green Version]
- Allard, P.-M.; Péresse, T.; Bisson, J.; Gindro, K.; Marcourt, L.; Pham, V.C.; Roussi, F.; Litaudon, M.; Wolfender, J.-L. Integration of Molecular Networking and In-Silico MS/MS Fragmentation for Natural Products Dereplication. Anal. Chem. 2016, 88, 3317–3323. [Google Scholar] [CrossRef]
- Triastuti, A. Exploration de La Diversité Chimique Dans Les Endophytes Fongiques: Influence de L’addition Des Modificateurs Épigénétiques et Des Co-Cultures Fongiques Sur Le Métabolome de Botryosphaeria mamane. Ph.D. Thesis, Université Toulouse III–Paul Sabatier, Toulouse, France, 2018. [Google Scholar]
- Kind, T.; Fiehn, O. Seven Golden Rules for Heuristic Filtering of Molecular Formulas Obtained by Accurate Mass Spectrometry. BMC Bioinform. 2007, 8, 105. [Google Scholar] [CrossRef] [Green Version]
- Weber, H.A.; Baenziger, N.C.; Gloer, J.B. Podosporin A: A Novel Antifungal Metabolite from the Coprophilous Fungus Podospora decipiens (Wint.) Niessl. J. Org. Chem. 1988, 53, 4567–4569. [Google Scholar] [CrossRef]
- Che, Y.; Gloer, J.B.; Koster, B.; Malloch, D. Decipinin A and Decipienolides A and B: New Bioactive Metabolites from the Coprophilous Fungus Podospora decipiens. J. Nat. Prod. 2002, 65, 916–919. [Google Scholar] [CrossRef]
- Zhang, Y.; Tian, R.; Liu, S.; Chen, X.; Liu, X.; Che, Y. Alachalasins A-G, New Cytochalasins from the Fungus Stachybotrys Podospora vesticola [Corrected]. Bioorg. Med. Chem. 2008, 16, 2627–2634. [Google Scholar] [CrossRef]
- Nielsen, K.F.; Larsen, T.O.; Frisvad, J.C. Lightweight Expanded Clay Aggregates (LECA), a New up-Scaleable Matrix for Production of Microfungal Metabolites. J. Antibiot. 2004, 57, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, J.C.; Prigent, S.; Grijseels, S.; Workman, M.; Ji, B.; Nielsen, J. Comparative Transcriptome Analysis Shows Conserved Metabolic Regulation during Production of Secondary Metabolites in Filamentous Fungi. mSystems 2019, 4, e00012-19. [Google Scholar] [CrossRef] [Green Version]
- Guljamow, A.; Kreische, M.; Ishida, K.; Liaimer, A.; Altermark, B.; Bähr, L.; Hertweck, C.; Ehwald, R.; Dittmann, E. High-Density Cultivation of Terrestrial Nostoc Strains Leads to Reprogramming of Secondary Metabolome. Appl. Environ. Microbiol. 2017, 83, e01510-17. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-J.; Andrei, A.-Ş.; Bulzu, P.-A.; Kavagutti, V.S.; Ghai, R.; Mosier, A.C. Expanded Diversity and Metabolic Versatility of Marine Nitrite-Oxidizing Bacteria Revealed by Cultivation- and Genomics-Based Approaches. Appl. Environ. Microbiol. 2020, 86, e01667-20. [Google Scholar] [CrossRef] [PubMed]
- Garrigues, S.; Kun, R.S.; Peng, M.; Gruben, B.S.; Benoit Gelber, I.; Mäkelä, M.; de Vries, R.P. The Cultivation Method Affects the Transcriptomic Response of Aspergillus niger to Growth on Sugar Beet Pulp. Microbiol. Spectr. 2021, 9, e0106421. [Google Scholar] [CrossRef]
- Gong, W.; Zhang, H.; Liu, S.; Zhang, L.; Gao, P.; Chen, G.; Wang, L. Comparative Secretome Analysis of Aspergillus niger, Trichoderma reesei, and Penicillium oxalicum During Solid-State Fermentation. Appl. Biochem. Biotechnol. 2015, 177, 1252–1271. [Google Scholar] [CrossRef] [PubMed]
- Volke-Sepulveda, T.; Salgado-Bautista, D.; Bergmann, C.; Wells, L.; Gutierrez-Sanchez, G.; Favela-Torres, E. Secretomic Insight into Glucose Metabolism of Aspergillus brasiliensis in Solid-State Fermentation. J. Proteome Res. 2016, 15, 3856–3871. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhang, N.; Pearce, R.; Yi, S.; Zhao, X. Comparative Secretomics Analysis Reveals the Major Components of Penicillium oxalicum 16 and Trichoderma reesei RUT-C30. Microorganisms 2021, 9, 2042. [Google Scholar] [CrossRef] [PubMed]
- Hoang, T.P.T.; Roullier, C.; Boumard, M.-C.; Robiou du Pont, T.; Nazih, H.; Gallard, J.-F.; Pouchus, Y.F.; Beniddir, M.A.; Grovel, O. Metabolomics-Driven Discovery of Meroterpenoids from a Mussel-Derived Penicillium ubiquetum. J. Nat. Prod. 2018, 81, 2501–2511. [Google Scholar] [CrossRef]
- Kim, H.Y.; Heo, D.Y.; Park, H.M.; Singh, D.; Lee, C.H. Metabolomic and Transcriptomic Comparison of Solid-State and Submerged Fermentation of Penicillium expansum KACC 40815. PLoS ONE 2016, 11, e0149012. [Google Scholar] [CrossRef] [Green Version]
- Zhong, J.-J.; Tang, Y.-J. Submerged Cultivation of Medicinal Mushrooms for Production of Valuable Bioactive Metabolites. Adv Biochem. Eng. Biotechnol. 2004, 87, 25–59. [Google Scholar] [CrossRef]
- Camelini, C.M.; Rossi, M.J.; de Sousa, F.T.G.; Giachini, A. Fungal Polysaccharide Production for Dermatological Purposes. In Polysaccharides of Microbial Origin: Biomedical Applications; Oliveira, J., Radhouani, H., Reis, R.L., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–32. ISBN 978-3-030-35734-4. [Google Scholar]
- Hölker, U.; Höfer, M.; Lenz, J. Biotechnological Advantages of Laboratory-Scale Solid-State Fermentation with Fungi. Appl. Microbiol. Biotechnol. 2004, 64, 175–186. [Google Scholar] [CrossRef]
- Droce, A.; Sørensen, J.L.; Giese, H.; Sondergaard, T.E. Glass Bead Cultivation of Fungi: Combining the Best of Liquid and Agar Media. J. Microbiol. Methods 2013, 94, 343–346. [Google Scholar] [CrossRef]
- Bigelis, R.; He, H.; Yang, H.Y.; Chang, L.-P.; Greenstein, M. Production of Fungal Antibiotics Using Polymeric Solid Supports in Solid-State and Liquid Fermentation. J. Ind. Microbiol. Biotechnol. 2006, 33, 815–826. [Google Scholar] [CrossRef]
- Barrios-González, J.; Baños, J.G.; Covarrubias, A.A.; Garay-Arroyo, A. Lovastatin Biosynthetic Genes of Aspergillus terreus Are Expressed Differentially in Solid-State and in Liquid Submerged Fermentation. Appl. Microbiol. Biotechnol. 2008, 79, 179–186. [Google Scholar] [CrossRef]
- Couto, S.R.; Sanromán, M.Á. Application of Solid-State Fermentation to Food Industry—A Review. J. Food Eng. 2006, 76, 291–302. [Google Scholar] [CrossRef]
- Levin, A.M.; de Vries, R.P.; Conesa, A.; de Bekker, C.; Talon, M.; Menke, H.H.; van Peij, N.N.M.E.; Wösten, H.A.B. Spatial Differentiation in the Vegetative Mycelium of Aspergillus niger. Eukaryot. Cell 2007, 6, 2311–2322. [Google Scholar] [CrossRef] [Green Version]
- Coconi Linares, N.; Di Falco, M.; Benoit-Gelber, I.; Gruben, B.S.; Peng, M.; Tsang, A.; Mäkelä, M.R.; de Vries, R.P. The Presence of Trace Components Significantly Broadens the Molecular Response of Aspergillus niger to Guar Gum. New Biotechnol. 2019, 51, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Salgado-Bautista, D.; Volke-Sepúlveda, T.; Figueroa-Martínez, F.; Carrasco-Navarro, U.; Chagolla-López, A.; Favela-Torres, E. Solid-State Fermentation Increases Secretome Complexity in Aspergillus brasiliensis. Fungal Biol. 2020, 124, 723–734. [Google Scholar] [CrossRef]
- Collemare, J.; Seidl, M.F. Chromatin-Dependent Regulation of Secondary Metabolite Biosynthesis in Fungi: Is the Picture Complete? FEMS Microbiol. Rev. 2019, 43, 591–607. [Google Scholar] [CrossRef] [Green Version]
- Bayram, O.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.-J.; Keller, N.P.; Yu, J.-H.; et al. VelB/VeA/LaeA Complex Coordinates Light Signal with Fungal Development and Secondary Metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef]
- Studt, L.; Schmidt, F.J.; Jahn, L.; Sieber, C.M.K.; Connolly, L.R.; Niehaus, E.-M.; Freitag, M.; Humpf, H.-U.; Tudzynski, B. Two Histone Deacetylases, FfHda1 and FfHda2, Are Important for Fusarium fujikuroi Secondary Metabolism and Virulence. Appl. Environ. Microbiol. 2013, 79, 7719–7734. [Google Scholar] [CrossRef] [Green Version]
- Kadooka, C.; Nakamura, E.; Mori, K.; Okutsu, K.; Yoshizaki, Y.; Takamine, K.; Goto, M.; Tamaki, H.; Futagami, T. LaeA Controls Citric Acid Production through Regulation of the Citrate Exporter-Encoding CexA Gene in Aspergillus luchuensis Mut. kawachii. Appl. Environ. Microbiol. 2020, 86, e01950-19. [Google Scholar] [CrossRef] [PubMed]
- Saha, P.; Ghosh, S.; Roy-Barman, S. MoLAEA Regulates Secondary Metabolism in Magnaporthe Oryzae. mSphere 2020, 5, e00936-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connolly, L.R.; Smith, K.M.; Freitag, M. The Fusarium graminearum Histone H3 K27 Methyltransferase KMT6 Regulates Development and Expression of Secondary Metabolite Gene Clusters. PLoS Genet. 2013, 9, e1003916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chujo, T.; Scott, B. Histone H3K9 and H3K27 Methylation Regulates Fungal Alkaloid Biosynthesis in a Fungal Endophyte-Plant Symbiosis. Mol. Microbiol. 2014, 92, 413–434. [Google Scholar] [CrossRef]
- Niehaus, E.-M.; Studt, L.; von Bargen, K.W.; Kummer, W.; Humpf, H.-U.; Reuter, G.; Tudzynski, B. Sound of Silence: The Beauvericin Cluster in Fusarium fujikuroi Is Controlled by Cluster-Specific and Global Regulators Mediated by H3K27 Modification. Environ. Microbiol. 2016, 18, 4282–4302. [Google Scholar] [CrossRef]
- Meng, S.; Liu, Z.; Shi, H.; Wu, Z.; Qiu, J.; Wen, H.; Lin, F.; Tao, Z.; Luo, C.; Kou, Y. UvKmt6-Mediated H3K27 Trimethylation Is Required for Development, Pathogenicity, and Stress Response in Ustilaginoidea virens. Virulence 2021, 12, 2972–2988. [Google Scholar] [CrossRef]
- Triastuti, A.; Vansteelandt, M.; Barakat, F.; Trinel, M.; Jargeat, P.; Fabre, N.; Amasifuen Guerra, C.A.; Mejia, K.; Valentin, A.; Haddad, M. How Histone Deacetylase Inhibitors Alter the Secondary Metabolites of Botryosphaeria mamane, an Endophytic Fungus Isolated from Bixa orellana. Chem. Biodivers. 2019, 16, e1800485. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, L.; Roullier, C.; Porée, F.-H.; Gaslonde, T.; Riffault-Valois, L.; Grovel, O.; Ruprich-Robert, G.; Chapeland-Leclerc, F. Complementary Strategies to Unlock Biosynthesis Gene Clusters Encoding Secondary Metabolites in the Filamentous Fungus Podospora anserina. J. Fungi 2023, 9, 9. https://doi.org/10.3390/jof9010009
Shen L, Roullier C, Porée F-H, Gaslonde T, Riffault-Valois L, Grovel O, Ruprich-Robert G, Chapeland-Leclerc F. Complementary Strategies to Unlock Biosynthesis Gene Clusters Encoding Secondary Metabolites in the Filamentous Fungus Podospora anserina. Journal of Fungi. 2023; 9(1):9. https://doi.org/10.3390/jof9010009
Chicago/Turabian StyleShen, Ling, Catherine Roullier, François-Hugues Porée, Thomas Gaslonde, Ludivine Riffault-Valois, Olivier Grovel, Gwenaël Ruprich-Robert, and Florence Chapeland-Leclerc. 2023. "Complementary Strategies to Unlock Biosynthesis Gene Clusters Encoding Secondary Metabolites in the Filamentous Fungus Podospora anserina" Journal of Fungi 9, no. 1: 9. https://doi.org/10.3390/jof9010009