
Strategies for Controlling the Sporulation in Fusarium spp.

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Sporulation in Fusarium
2.1. Sexual Spores
2.2. Asexual Spores
2.3. Genetic Pathway Responsible for Spore Formation
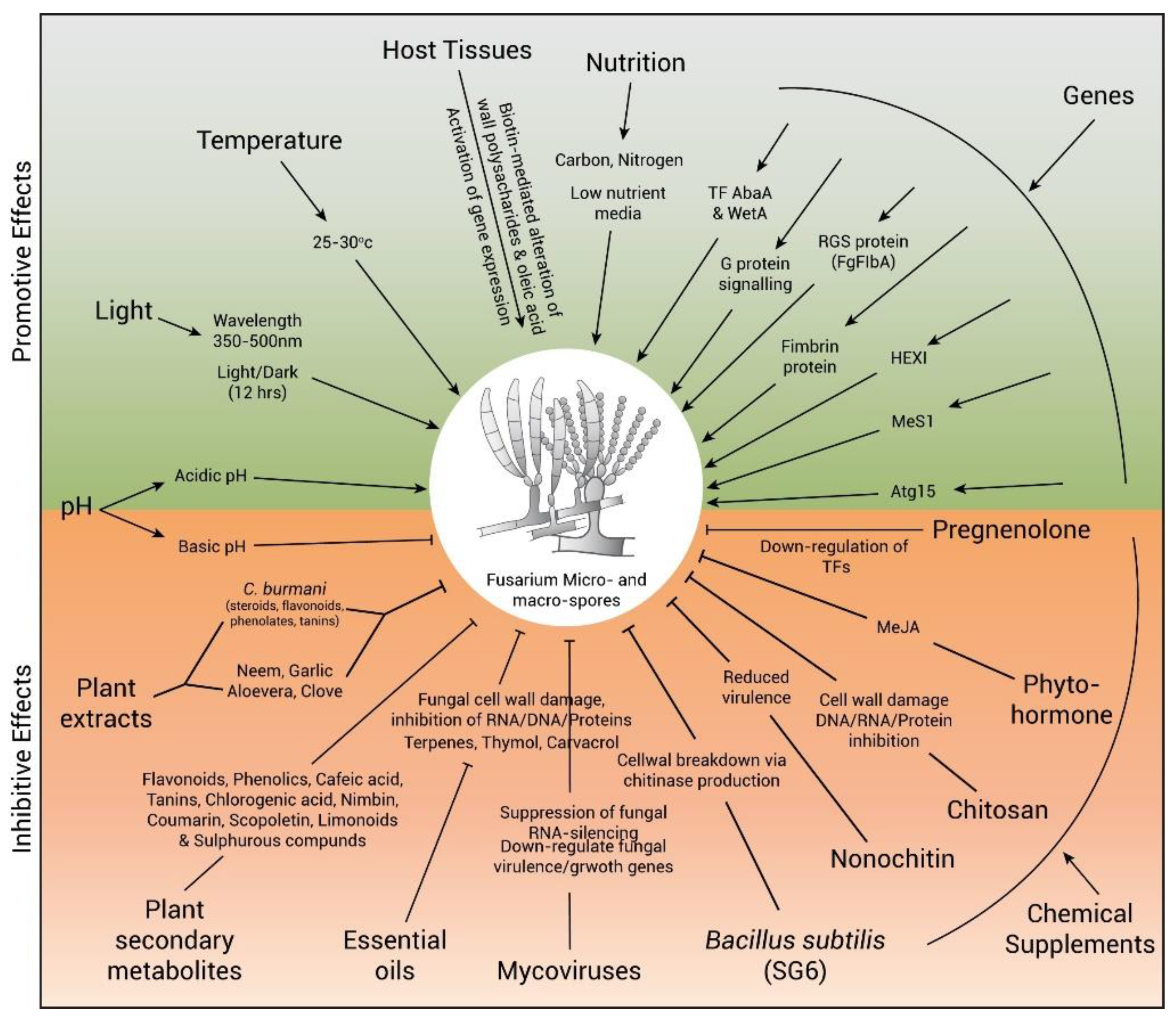
3. Growth Conditions and Environmental Factors Affecting Sporulation
3.1. Temperature
3.2. Light
3.3. PH
3.4. Host Tissue
3.5. Nutrition
4. Artificial Control of Sporulation in Fusarium
4.1. Biological Control Agents
4.1.1. Leaf Extracts
4.1.2. Essential Oils
4.1.3. Mycovirus
4.1.4. Rhizospheric Bacteria
4.2. Chemical Supplements
4.3. Transgenic Approaches to Control of Sporulation in Fusarium
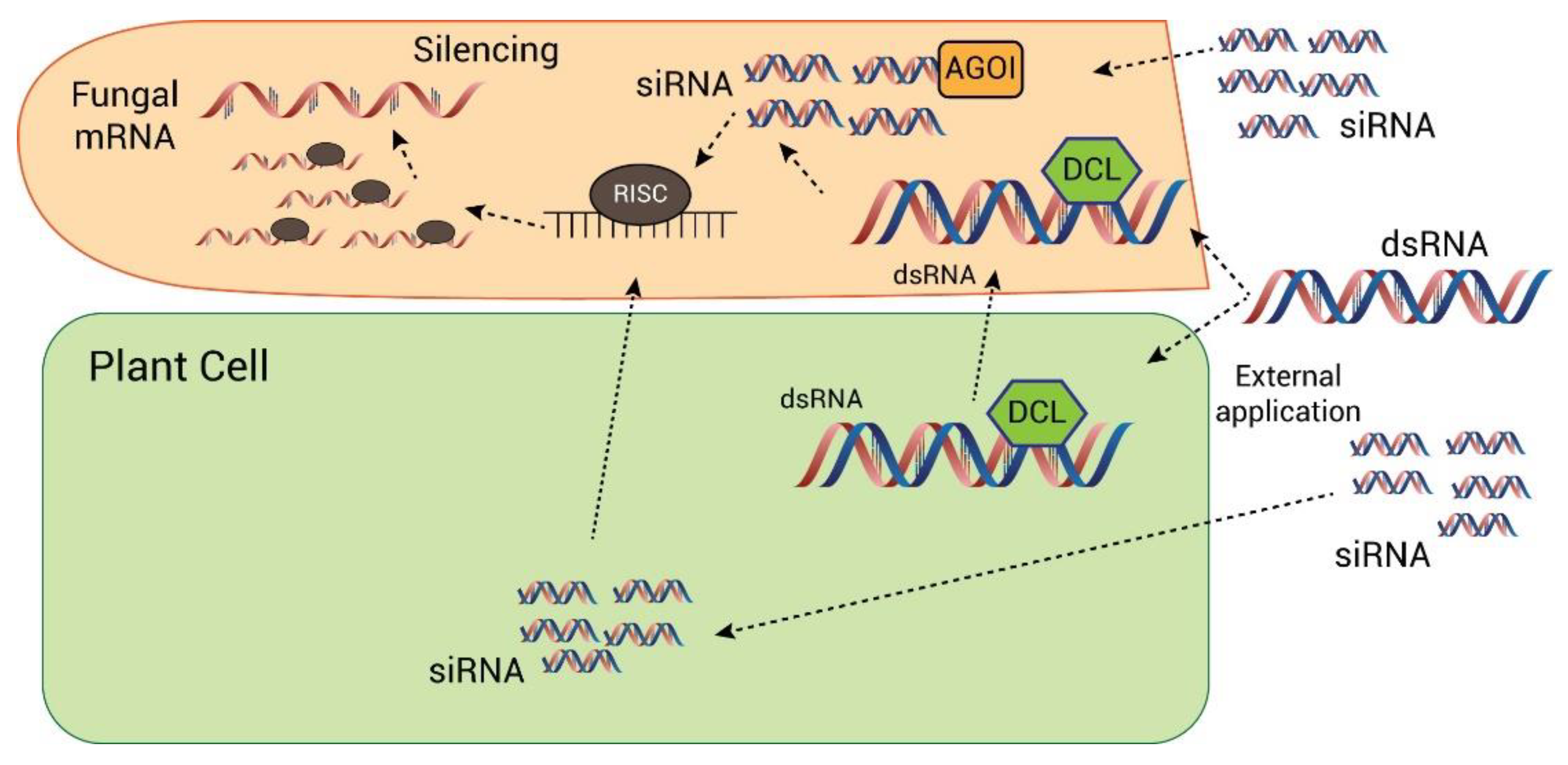
4.3.1. Host-Delivered RNAi or Host-Induced Gene Silencing
4.3.2. Spray-Induced Gene Silencing
5. Future Perspectives in Sporulation Control in Fusarium
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bottalico, A.; Perrone, G. Toxigenic Fusarium Species and Mycotoxins Associated with Head Blight in Small-Grain Cereals in Europe, Mycotoxins in Plant Disease. Eur. J. Plant Pathol. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- McMullen, M.; Bergstrom, G.; De Wolf, E.; Dill-Macky, R.; Hershman, D.; Shaner, G.; Sanford, D.V. A unified effort to fight an enemy of wheat and barley: Fusarium Head Blight. Plant Dis. 2012, 96, 1712–1855. [Google Scholar] [CrossRef] [Green Version]
- Leslie, J.; Summerell, B. Fusarium laboratory workshops-A recent history. Mycotoxin Res. 2006, 22, 73. [Google Scholar] [CrossRef] [PubMed]
- Babadoost, M. Fusarium: Historical and Continued Importance; Books on Demand: Balikesir, Turkey, 2017. [Google Scholar] [CrossRef] [Green Version]
- Early, R. Pathogen control in primary production: Crop foods. Foodborne Pathog. 2009, 2009, 205–279. [Google Scholar]
- Mui-Yun, W. Fusarium oxysporum f. sp. lycopersici (Sacc.): PP728 Soil-Borne Plant Pathogen Class Project; North Carolina State University: Raleigh, NC, USA, 2003. [Google Scholar]
- Nelson, P.E.; Dignani, M.C.; Anaissie, E.J. Taxonomy, biology, and clinical aspects of Fusarium species. Clin. Microbiol. Rev. 1994, 7, 479–504. [Google Scholar] [CrossRef]
- Arif, M.; Pani, D.R.; Zaidi, N.W.; Singh, S.U. PCR-based identification and characterization of Fusarium sp. associated with mango malformation. Biotechnol. Res. Int. 2011, 6, 141649. [Google Scholar]
- Balali, G.; Iranpour, M. Identification and genetic variation of Fusarium species in Isfahan, Iran, using pectic Zymogram technique. Iran J. Sci. Technol. 2006, 30, 91–102. [Google Scholar]
- Wang, H.; Xiao, M.; Kong, F.; Chen, S.; Dou, H.T.; Sorrell, T.; Li, R.Y.; Xu, Y.C. Accurate and practical identification of 20 Fusarium species by seven-locus sequence analysis and reverse line blot hybridization, and an in vitro antifungal susceptibility study. J. Clin. Microbiol. 2011, 49, 1890–1898. [Google Scholar] [CrossRef] [Green Version]
- Bhat, R.; Rai, R.V.; Karim, A.A. Mycotoxins in Food and Feed: Present Status and Future Concerns. Compr. Rev. Food Sci. Food Saf. 2010, 9, 57–81. [Google Scholar] [CrossRef]
- Ma, L.J.; Van Der Does, H.C.; Borkovich, K.A.; Coleman, J.J.; Daboussi, M.J.; Di Pietro, A.; Dufresne, M.; Freitag, M.; Grabherr, M.; Henrissat, B.J. Comparative genomics reveals mobile pathogenicity chromosomes in Fusarium. Nature 2010, 464, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Moss, M.O.; Thrane, U. Fusarium taxonomy with relation to trichothecene formation. Toxicol. Lett. 2004, 153, 23–28. [Google Scholar] [CrossRef]
- Windels, C.E. Economic and social impacts of Fusarium head blight: Changing farms and rural communities in the Northern Great Plains. Phytopathology 2000, 90, 17–21. [Google Scholar] [CrossRef]
- Aoki, T.; O’Donnell, K.; Geiser, D.M. Systematics of key phytopathogenic Fusarium species: Current status and future challenges. J. Gen. Plant Pathol. 2014, 80, 189–201. [Google Scholar] [CrossRef]
- Asam, S.; Habler, K.; Rychlik, M. Fusarium Mycotoxins in Food, Chemical Contaminants and Residues in Food; Elsevier: Amsterdam, The Netherlands, 2017; pp. 295–336. [Google Scholar]
- Bai, G.H.; Desjardins, A.; Plattner, R.D. Deoxynivalenol-nonproducing Fusarium graminearum causes initial infection, but does not cause disease spread in wheat spikes. Mycopathologia 2002, 153, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Desmond, O.J.; Manners, J.M.; Stephens, A.E.; Maclean, D.J.; Schenk, P.M.; Gardiner, D.M.; Munn, A.L.; Kazan, K. The Fusarium mycotoxin deoxynivalenol elicits hydrogen peroxide production, programmed cell death and defence responses in wheat. Mol. Plant Pathol. 2008, 9, 435–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desjardins, A.E.; Proctor, R.; Bai, G.; McCormick, S.; Shaner, G.; Buechley, G.; Hohn, T. Reduced virulence of trichothecene-nonproducing mutants of Gibberella zeae in wheat field tests. Mol. Plant Microbe Interact. 1996, 9, 775–781. [Google Scholar] [CrossRef]
- Ilgen, P.; Maier, F.; Schäfer, W. Trichothecenes and lipases are host-induced and secreted virulence factors of Fusarium graminearum. Cereal Res. Commun. 2008, 36, 421–428. [Google Scholar] [CrossRef]
- Proctor, R.H.; Hohn, T.M.; McCormick, S.P. Reduced virulence of Gibberella zeae caused by disruption of a trichothecene toxin biosynthetic gene. Mol. Plant Microbe Interact. 1995, 8, 593–601. [Google Scholar] [CrossRef] [Green Version]
- Muimba-Kankolongo, A. Food Crop Production by Smallholder Farmers in Southern Africa: Challenges and Opportunities for Improvement; Elsevier: Amsterdam, The Netherlands, 2018; p. 368. [Google Scholar]
- Woloshuk, C.P.; Shim, W.B. Aflatoxins, fumonisins, and trichothecenes: A convergence of knowledge. FEMS Microbiol. Rev. 2013, 37, 94–109. [Google Scholar] [CrossRef] [Green Version]
- Knights, E.; Hobson, K. Chickpea Overview, Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2016; pp. 316–323. [Google Scholar] [CrossRef]
- Shanmugam, V.; Chugh, P.; Sharma, P. Cold-tolerant Trichoderma species for the managementof Fusarium wilt of tomato plants. Ann. Microbiol. 2015, 65, 543–551. [Google Scholar] [CrossRef]
- Rampersad, S.N. Pathogenomics and management of Fusarium diseases in plants. Pathogen 2020, 9, 340. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.J.; Rounsley, S.D.; Rodriguez-Carres, M.; Kuo, A.; Wasmann, C.C.; Grimwood, J.; Schmutz, J.; Taga, M.; White, G.J.; Zhou, S.; et al. The genome of Nectria haematococca: Contribution of supernumerary chromosomes to gene xxpansion. PLoS Genet. 2009, 5, e1000618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyde, K.D.; Abd-Elsalam, K.; Cai, L.J.M. Morphology: Still essential in a molecular world. Mycotaxon 2010, 114, 439–451. [Google Scholar] [CrossRef]
- Deacon, J. Fungal Spores, Spore Dormancy, and Spore Dispersal. Fungal Biology, 4th ed.; Blackwell Publishing: Oxford, UK, 2006; pp. 184–212. [Google Scholar]
- Ma, L.J.; Geiser, D.M.; Proctor, R.H.; Rooney, A.P.; O’Donnell, K.; Trail, F.; Gardiner, D.M.; Manners, J.M.; Kazan, K. Fusarium pathogenomics. Annu. Rev. Microbiol. 2013, 67, 399–416. [Google Scholar] [CrossRef] [Green Version]
- Desjardins, A.E.; Plattner, R.D.; Shaner, G.; Brown, D.W.; Buechley, G.; Proctor, R.H.; Turgeon, G. Field release of Gibberella zeae genetically modified to lack ascospores. In Proceedings of the 2006 National Fusarium Head Blight Forum; University of Kentucky: Lexington, KY, USA, 2006; pp. 39–44. [Google Scholar]
- VanEtten, H.D. Identification of additional habitats of Nectria haematococca mating population VI. Phytopathology 1978, 68, 6. [Google Scholar] [CrossRef]
- Desjardins, A.E. Gibberella from A (venaceae) to Z (eae). Annu. Rev. Phytopathol. 2003, 41, 177–198. [Google Scholar] [CrossRef] [PubMed]
- Agrios, G.N. Plant Pathology, 5th ed.; Elsevier Academic Press: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Adams, T.H.; Wieser, J.K.; Yu, J.H.J.M. Asexual sporulation in Aspergillus nidulans. Microbiol. Mol. Biol. Rev. 1998, 62, 35–54. [Google Scholar] [CrossRef] [Green Version]
- Gordon, T.R. Fusarium oxysporum and the Fusarium wilt syndrome. Annu. Rev. Phytopathol. 2017, 55, 23–39. [Google Scholar] [CrossRef]
- Srinivas, C.; Devi, D.; Murthy, K.; Mohan, C.; Lakshmeesha, T.; Singh, B.; Kalagatur, N.; Niranjana, S.; Hashem, A.; Alqarawi, A. Fusarium oxysporum f. sp. lycopersici causal agent of vascular wilt disease of tomato: Biology to diversity—A review. Saudi J. Biol. Sci. 2019, 26, 1315–1324. [Google Scholar] [CrossRef]
- Okungbowa, F.; Shittu, H. Fusarium Wilts: An Overview. Environ. Res. J. 2014, 6, 83–102. [Google Scholar]
- Xu, J.R. MAP kinases in fungal pathogens. Fungal Genet. Biol. 2000, 31, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Roncal, T.; Ugalde, U. Conidiation induction in Penicillium. Res. Microbiol. 2003, 154, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.W.; Zhao, W.; Xu, Y.N.; Zhong, J.J. Isolation and analysis of differentially expressed genes during asexual sporulation in liquid static culture of Ganoderma lucidum by suppression subtractive hybridization. Mol. Biol. Rep. 2012, 39, 3603–3610. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Wang, J.; Qiu, L.; Feng, M.G. A Group III histidine kindase (mhk1) pstream of high-osmolarity glycerol pathway regulates sporulation, multi-stress tolerance and virulence of Metarhizium robertsii, a fungal entomopathogen. Environ. Biol. 2012, 4, 817–829. [Google Scholar]
- Son, H.; Kim, M.G.; Min, K.; Seo, Y.S.; Lim, J.Y.; Choi, G.J.; Kim, J.C.; Chae, S.K.; Lee, Y.W. AbaA regulates conidiogenesis in the ascomycete fungus Fusarium graminearum. PLoS ONE 2013, 8, e72915. [Google Scholar] [CrossRef] [Green Version]
- Son, H.; Kim, M.G.; Min, K.; Lim, J.Y.; Choi, G.J.; Kim, J.C.; Chae, S.K.; Lee, Y. WetA is required for conidiogenesis and conidium maturation in the ascomycete fungus Fusarium graminearum. Eukaryot. Cell 2014, 13, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Park, A.R.; Cho, A.R.; Seo, J.A.; Min, K.; Son, H.; Lee, J.; Choi, G.J.; Kim, J.C.; Lee, Y.W. Functional analyses of regulators of G protein signaling in Gibberella zeae. Fungal Genet. Biol. 2012, 49, 511–520. [Google Scholar] [CrossRef]
- Son, M.; Lee, K.M.; Yu, J.; Kang, M.; Park, J.M.; Kwon, S.J.; Kim, K.H. The HEX1 gene of Fusarium graminearum is required for fungal asexual reproduction and pathogenesis and for efficient viral RNA accumulation of Fusarium graminearum virus 1. J. Virol. 2013, 87, 10356–10367. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.N.; Bormann, J.; Le, G.T.T.; Stärkel, C.; Olsson, S.; Nosanchuk, J.D.; Giese, H.; Schäfer, W. Autophagy-related lipase FgATG15 of Fusarium graminearum is important for lipid turnover and plant infection. Fungal Genet. Biol. 2011, 48, 217–224. [Google Scholar] [CrossRef]
- Rittenour, W.R.; Harris, S.D. Characterization of Fusarium graminearum Mes1 reveals roles in cell-surface organization and virulence. Fungal Genet. Biol. 2008, 45, 933–946. [Google Scholar] [CrossRef]
- Lee, J.; Myong, K.; Kim, J.E.; Kim, H.K.; Yun, S.H.; Lee, Y.W. FgVelB globally regulates sexual reproduction, mycotoxin production and pathogenicity in the cereal pathogen Fusarium graminearum. J. Microbiol. 2012, 158, 1723–1733. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Liu, X.; Yin, Y.; Ma, Z. Involvement of a velvet protein FgVeA in the regulation of asexual development, lipid and secondary metabolisms and virulence in Fusarium graminearum. PLoS ONE 2011, 6, e28291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Yun, Y.; Liu, Y.; Ma, Z. FgVELB is associated with vegetative differentiation, secondary metabolism and virulence in Fusarium graminearum. Fungal Genet. Biol. 2012, 49, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Son, H.; Min, K.; Lee, J.; Choi, G.J.; Kim, J.C.; Lee, Y.W. Mitochondrial carnitine-dependent acetyl coenzyme A transport is required for normal sexual and asexual development of the ascomycete Gibberella zeae. Eukaryot. Cell 2012, 11, 1143–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldwin, T.K.; Urban, M.; Brown, N.; Hammond-Kosack, K.E. A role for topoisomerase I in Fusarium graminearum and F. culmorum pathogenesis and sporulation. Mol. Plant Microbe Interact. 2010, 23, 566–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavinder, B.; Hamam, A.; Lew, R.R.; Trail, F. Mid1, a mechanosensitive calcium ion channel, affects growth, development, and ascospore discharge in the filamentous fungus Gibberella zeae. Eukaryot. Cell 2011, 10, 832–841. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.K.; Lee, T.; Han, K.H.; Yun, S.H.; Lee, Y.W. Functional analysis of the homoserine O-acetyltransferase gene and its identification as a selectable marker in Gibberella zeae. Curr. Genet. 2004, 46, 205–212. [Google Scholar] [CrossRef]
- Li, Y.; Wang, C.; Liu, W.; Wang, G.; Kang, Z.; Kistler, H.C.; Xu, J.R. The HDF1 histone deacetylase gene is important for conidiation, sexual reproduction, and pathogenesis in Fusarium graminearum. Mol. Plant-Microbe Interact. 2011, 24, 487–496. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Gao, T.; Zsinhang, Y.; Hou, Y.; Wang, J.; Zhou, M. FgFim, a key protein regulating resistance to the fungicide JS 399-19, asexual and sexual development, stress responses and virulence in Fusarium graminearum. Mol. Plant Pathol. 2014, 15, 488–499. [Google Scholar] [CrossRef]
- Steyaert, J.M.; Weld, R.J.; Mendoza-Mendoza, A.; Stewart, A. Reproduction without sex: Conidiation in the filamentous fungus Trichoderma. Microbiology 2010, 156, 2887–2900. [Google Scholar] [CrossRef] [Green Version]
- Katayama, M.; Yanagi, M.; Marumo, S. Isolation of sporogen-PF 1, a blue light-induced sporogenic substance, from Penicillium funiculosum. Agric. Biol. Chem. 1989, 53, 3379–3380. [Google Scholar] [CrossRef]
- Roncal, T.; Cordobés, S.; Sterner, O.; Ugalde, U. Conidiation in Penicillium cyclopium is induced by conidiogenone, an endogenous diterpene. Eukaryot. Cell 2002, 1, 823–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoppacher, N.; Kluger, B.; Zeilinger, S.; Krska, R.; Schuhmacher, R. Identification and profiling of volatile metabolites of the biocontrol fungus Trichoderma atroviride by HS-SPME-GC-MS. J. Microbiol. Methods 2010, 81, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Bosland, P.; Williams, P.; Morrison, R. Influence of soil temperature on the expression of yellows and wilt of crucifers by Fusarium oxysporum. Plant Dis. 1988, 72, 777–780. [Google Scholar] [CrossRef]
- Frans, M.; Aerts, R.; Van Laethem, S.; Ceusters, J. Environmental effects on growth and sporulation of Fusarium spp. causing internal fruit rot in bell pepper. Eur. J. Plant Pathol. 2017, 149, 875–883. [Google Scholar]
- Rossi, V.; Scandolara, A.; Battilani, P. Effect of environmental conditions on spore production by Fusarium verticillioides, the causal agent of maize ear rot. Eur. J. Plant Pathol. 2009, 123, 159–169. [Google Scholar] [CrossRef]
- Marin, S.; Sanchis, V.; Magan, N.J. Water activity, temperature, and pH effects on growth of Fusarium moniliforme and Fusarium proliferatum isolates from maize. Can. J. Microbiol. 1995, 41, 1063–1070. [Google Scholar] [CrossRef]
- Doohan, F.; Brennan, J.; Cooke, B. Influence of Climatic Factors on Fusarium Species Pathogenic to Cereals, Epidemiology of Mycotoxin Producing Fungi; Springer: Dordrecht, Netherlands, 2003; pp. 755–768. [Google Scholar]
- Tonapi, V.A.; Mundada, R.R.; Navi, S.S.; Reddy, R.K.; Thakur, R.P.; Bandyopadhyay, R.; Varanavasiappan, S.; Seetharama, N. Effect of temperature and humidity regimes on grain mold sporulation and seed quality in sorghum (Sorghum bicolor (L.) Moench). Arch. Phytopathol. Plant Prot. 2007, 40, 113–127. [Google Scholar] [CrossRef] [Green Version]
- Daami-Remadi, M.; Jabnoun-Khiareddine, H.; Ayed, F.; El Mahjoub, M. Effect of temperature on aggressivity of Tunisian Fusarium species causing potato (Solanum tuberosum L.) tuber dry rot. J. Agron. 2006, 5, 350–355. [Google Scholar] [CrossRef] [Green Version]
- Desai, A.; Dange, S.; Patel, D.S.; Patel, D.B. Variability in Fusarium oxysporum f. sp. ricini causing wilt of castor. Mycol. Plant Pathol. 2003, 33, 37–41. [Google Scholar]
- Crous, P.W.; Slippers, B.; Wingfield, M.J.; Rheeder, J.; Marasas, W.F.; Philips, A.J.; Alves, A.; Burgess, T.; Barber, P.; Groenewald, J.Z. Phylogenetic lineages in the Botryosphaeriaceae. Stud. Mycol. 2006, 55, 235–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Idnurm, A.; Rodríguez-Romero, J.; Corrochano, L.M.; Sanz, C.; Iturriaga, E.A.; Eslava, A.P.; Heitman, J. The Phycomyces madA gene encodes a blue-light photoreceptor for phototropism and other light responses. Proc. Natl. Acad. Sci. USA 2006, 103, 4546–4551. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Li, F.; Xie, H.; Liu, X. A novel method for promoting conidial production by a nematophagous fungus, Pochonia chlamydosporia AS6.8. World J. Microbiol. Biotechnol. 2009, 25, 1989–1994. [Google Scholar] [CrossRef]
- Starostzik, C.; Marwan, W. A photoreceptor with characteristics of phytochrome triggers sporulation in the true slime mould Physarum polycephalum. FEBS Lett. 1995, 370, 146–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahlberg, K.R.; Etten, J. Physiology and biochemistry of fungal sporulation. Ann. Rev. Phytopathol. 1982, 20, 281–301. [Google Scholar] [CrossRef]
- Rakoczy, L. Influence of monochromatic light on the fructification of Physarum nudum. Acta Soc. Bot. Pol. 1963, 11, 559–562. [Google Scholar]
- Rakoczy, L. Action spectrum in sporulation of slime-mold Physarum nudum Macbr. Acta Soc. Bot. Pol. 1965, 34, 97–112. [Google Scholar] [CrossRef]
- Tisch, D.; Schmoll, M. Light regulation of metabolic pathways in fungi. Appl. Microbiol. Biotechnol. 2010, 85, 1259–1277. [Google Scholar] [CrossRef] [Green Version]
- Leach, C.M. Sporulation of diverse species of fungi under near-ultraviolet radiation. Can. J. Bot. 1962, 40, 151–161. [Google Scholar] [CrossRef]
- Das, J.; Busse, H. Light-driven diurnal zonation in the filamentous fungus Fusarium solani. Int. J. Dev. Biol. 1990, 34, 319–322. [Google Scholar]
- Leach, C.; Tulloch, M. Induction of sporulation of fungi isolated from Dactylis glomerata seed by exposure to near-ultraviolet radiation. Ann. Appl. Biol. 1972, 72, 155–159. [Google Scholar] [CrossRef]
- Avalos, J.; Estrada, A.F. Regulation by light in Fusarium. Fungal Genet. Biol. 2010, 47, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Corrochano, L.M. Fungal photoreceptors: Sensory molecules for fungal development and behaviour. Photochem. Photobiol. Sci. 2007, 6, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Romero, J.; Hedtke, M.; Kastner, C.; Müller, S.; Fischer, R. Fungi, hidden in soil or up in the air: Light makes a difference. Annu. Rev. Microbiol. 2010, 64, 585–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruger-Herreros, C.; Rodríguez-Romero, J.; Fernández-Barranco, R.; Olmedo, M.; Fischer, R.; Corrochano, L.M.; Canovas, D. Regulation of conidiation by light in Aspergillus nidulans. Genetics 2011, 188, 809–822. [Google Scholar] [CrossRef] [Green Version]
- Froehlich, A.C.; Liu, Y.; Loros, J.J.; Dunlap, J.C. White Collar-1, a circadian blue light photoreceptor, binding to the frequency promoter. Science 2002, 297, 815–819. [Google Scholar] [CrossRef]
- He, Q.; Cheng, P.; Yang, Y.; Wang, L.; Gardner, K.H.; Liu, Y. White collar-1, a DNA binding transcription factor and a light sensor. Science 2002, 297, 840–843. [Google Scholar] [CrossRef]
- Schafmeier, T.; Diernfellner, A.C. Light input and processing in the circadian clock of Neurospora. FEBS Lett. 2011, 585, 1467–1473. [Google Scholar] [CrossRef] [Green Version]
- Idnurm, A.; Verma, S.; Corrochano, L.M. A glimpse into the basis of vision in the kingdom Mycota. Fungal Genet. Biol. 2010, 47, 881–892. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Arreguín, A.; Pérez-Martínez, A.S.; Herrera-Estrella, A. Proteomic analysis of Trichoderma atroviride reveals independent roles for transcription factors BLR-1 and BLR-2 in light and darkness. Eukaryot. Cell 2012, 11, 30–41. [Google Scholar] [CrossRef] [Green Version]
- Estrada, A.F.; Avalos, J. The White-Collar protein WcoA of Fusarium fujikuroi is not essential for photocarotenogenesis, but is involved in the regulation of secondary metabolism and conidiation. Fungal Genet. Biol. 2008, 45, 705–718. [Google Scholar] [CrossRef] [PubMed]
- Souramma, V.; Singh, J. Effect of temperature and pH on growth and sporulation of wilt causing pathogen in linseed. J. Oilseeds Res. 2004, 21, 206–207. [Google Scholar]
- Groenewald, S. Biology, Pathogenicity and Diversity of Fusarium oxysporum Fsp Cubense; University of Pretoria: Pretoria, South Africa, 2005; p. 316. [Google Scholar]
- Gupta, V.; Misra, A.; Gaur, R. Growth characteristics of Fusarium spp. causing wilt disease in Psidium guajava L. in India. J. Plant Prot. Res. 2010, 50, 453–462. [Google Scholar] [CrossRef]
- Ahamad, S.; Agarwal, D.; Narain, U.; Chauhan, S. Effect of temperature, pH, light and incubation periods on growth, sporulation, biomass and gibberellic acid production. Ann. Plant Prot. Sci. 2002, 10, 343–348. [Google Scholar]
- Sharma, R.; Singh, B.; Thakur, M.; Thapak, S.K. Effect of media, temperature, pH and Light on the growth and sporulation of Fusarium oxysporum f. sp. lini (Bolley) Snyder and Hensan. Ann. Plant Prot. Sci. 2005, 13, 172–174. [Google Scholar]
- Kishore, R.; Pandey, M.; Dubey, M.K.; Kumar, Y. Effect of Temperature and pH on Growth and Sporulation of Fusarium Oxysporum f. sp. Lini (Bolley) Snyder and Hensan Causing Linseed Wilt. Progress. Agric. 2009, 9, 147–149. [Google Scholar]
- Photita, W.; Lumyong, S.; Lumyong, P.; Hyde, K.D. Endophytic fungi of wild banana (Musa acuminata) at doi Suthep Pui National Park, Thailand. Mycol. Res. 2001, 105, 1508–1513. [Google Scholar] [CrossRef]
- Okane, I.; Nakagiri, A.; Ito, T. Identity of Guignardia sp. inhabiting ericaceous plants. Can. J. Bot. 2001, 79, 101–109. [Google Scholar]
- Fisher, N.L.; Burgess, L.; Toussoun, T.; Nelson, P.E. Carnation leaves as a substrate and for preserving cultures of Fusarium species. Phytopathology 1982, 72, 151–153. [Google Scholar] [CrossRef]
- Liu, A.R.; Chen, S.C.; Wu, S.Y.; Xu, T.; Guo, L.D.; Jeewon, R.; Wei, J.G. Cultural studies coupled with DNA based sequence analyses and its implication on pigmentation as a phylogenetic marker in Pestalotiopsis taxonomy. Mol. Phylogenet. Evol. 2010, 57, 528–535. [Google Scholar] [CrossRef]
- Wei, J.; Xu, T.; Guo, L.D. Morphological stability and taxonomical significance of the genus Pestalotiopsis. J. Laiyang Agric. Coll. 2006, 23, 280–284. [Google Scholar]
- Hassan, Y.I.; Bullerman, L.B. Wheat bran as an alternative substrate for macroconidia formation by some Fusarium species. J. Microbiol. Methods 2009, 77, 134–136. [Google Scholar] [CrossRef] [PubMed]
- Leaver, F.; Leal, J.; Brewer, C. Nutritional studies on Piricularia oryzae. J. Bacteriol. 1947, 54, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Misawa, T. Nutritional factors for phytopathogenic fungi on culture media. Jpn. J. Phytopathol. 1965, 31, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Qi, Y.; Cai, L. Induction of sporulation in plant pathogenic fungi. Fungal Biol. 2012, 3, 195–200. [Google Scholar]
- Timberlake, W.E. Developmental gene regulation in Aspergillus nidulans. Dev. Biol. 1980, 78, 497–510. [Google Scholar] [CrossRef]
- Yamaguchi, H. Mycelial development and chemical alteration of Candida albicans from biotin insufficiency. Sabouraudia 1974, 12, 320–328. [Google Scholar] [CrossRef]
- Yoshida, S.; Shirata, A. Biotin induces sporulation of mulberry anthracnose fungus, Colletotrichum dematium. J. Gen. Plant Pathol. 2000, 66, 117–122. [Google Scholar] [CrossRef]
- Timnick, M.B.; Lilly, V.G.; Barnett, H. The effect of nutrition on the sporulation of Melanconium fuligineum in culture. Mycologia 1951, 43, 625–634. [Google Scholar] [CrossRef]
- Engelkes, C.; Nuclo, R.; Fravel, D. Effect of carbon, nitrogen, and C: N ratio on growth, sporulation, and biocontrol efficacy of Talaromyces flavus. J. Phytopathol. 1997, 87, 500–505. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Sun, M.H.; Liu, X.Z.; Che, Y.S. Effects of carbon concentration and carbon to nitrogen ratio on the growth and sporulation of several biocontrol fungi. Mycol. Res. 2007, 111, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Wulandari, N.; To-Anun, C.; Hyde, K.; Duong, L.; De Gruyter, J.; Meffert, J.; Groenewald, J.; Crous, P. Phyllosticta citriasiana sp. nov., the cause of Citrus tan spot of Citrus maxima in Asia. Fungal Divers. 2009, 34, 23–39. [Google Scholar]
- Braun, U.; Crous, P.W.; Groenewald, J.Z.; Scheuer, C. Pseudovirgaria, a fungicolous hyphomycete genus. IMA Fungus. 2011, 2, 65–69. [Google Scholar] [CrossRef] [Green Version]
- Masangkay, R.F.; Paulitz, T.C.; Hallett, S.G.; Watson, A.K. Characterization of sporulation of Alternaria alternata f. sp. sphenocleae. Biocontrol Sci. Technol. 2000, 10, 385–397. [Google Scholar] [CrossRef]
- Booth, C. Chapter II Fungal Culture Media, Methods in Microbiology; Academic Press Inc.: London, UK, 1971; pp. 49–94. [Google Scholar]
- Guo, L.D. A method to promote sporulation in palm endophytic fungi. Fungal Divers. 1998, 1, 109–113. [Google Scholar]
- Li, W.C.; Zhou, J.; Guo, S.Y.; Guo, L.D. Endophytic fungi associated with lichens in Baihua mountain of Beijing, China. Fungal Divers. 2007, 25, 69–80. [Google Scholar]
- Kishore, C.; Kulkarni, S. Evaluation of Plant Extracts and Biocontrol Agents Against Fusarium Oxysporum f. sp. Gerberae Causing Wilt of Gerbera. J. Plant Dis. Sci. 2008, 3, 108–110. [Google Scholar]
- Mezzomo, R.; Rolim, J.M.; Poletto, T.; De Oliveira, M.B.; Lazarotto, M.; Muñiz, M.F. Mycelial growth and sporulation of Fusarium spp. Pathogenic to Ilex paraguariensis in different culture media and under exposure to different light levels. Sci. Agrar. 2018, 19, 14–19. [Google Scholar] [CrossRef]
- Arie, T. Fusarium diseases of cultivated plants, control, diagnosis, and molecular and genetic studies. J. Pestic. Sci. 2019, 44, 275–281. [Google Scholar] [CrossRef] [Green Version]
- Djunaedy, A. Biopesticides as control of plant pests (opt) which are environmentally friendly. Embryo 2009, 6, 88–95. [Google Scholar]
- Astiti, N.P.A.; Suprapta, D.N. Antifungal activity of teak (Tectona grandis Lf) leaf extract against Arthrinium phaeospermum (corda) MB Ellis, the cause of wood decay on Albizia falcataria (L). ISSAAS 2012, 18, 62–69. [Google Scholar]
- Suprapta, D.; Sudana, M.; Alit-Susanta, W.; Sudiarta, P. Plant extracts to control cocoa black pod disease caused by Phytophthora palmivora. J. Int. Soc. Southeast Asian Agric. Sci. 2008, 13, 1–30. [Google Scholar]
- Suprapta, D.; Sudarma, M.; Arya, N.; Ohsawa, K. Plant Extracts to Control Wilt Desease in Banana Seedlings. J. Int. Soc. Southeast Asian Agric. Sci. 2005, 11, 84–90. [Google Scholar]
- Suprapta, D.N.; Khalimi, K. Anti-fungal activities of selected tropical plants from Bali Island. J. Phytopharm. 2012, 2, 265–270. [Google Scholar]
- Suprapta, D.N.; Khalimi, K. Efficacy of plant extract formulations to suppress stem rot disease on vanilla seedlings. J. Int. Soc. Southeast Asian Agric. Sci. 2009, 15, 34–41. [Google Scholar]
- Begum, S.; Devi, R.T.; Singh, N.I. Evaluation of fungicides, biocontrol agents and botanicals for management of damping-off in cabbage seedlings caused by Fursarium moniliforme sheld. J. Appl. Nat. Sci. 2015, 7, 106–110. [Google Scholar] [CrossRef] [Green Version]
- Pfleger, F.; Harman, G. Fungal antisporulant activity of a complex lipid fraction extracted from pea seeds. Can. J. Bot. 1975, 53, 1625–1629. [Google Scholar] [CrossRef]
- Drakopoulos, D.; Luz, C.; Torrijos, R.; Meca, G.; Weber, P.; Bänziger, I.; Voegele, R.T.; Six, J.; Vogelgsang, S. Use of botanicals to suppress different stages of the life cycle of Fusarium graminearum. Phytopathology 2019, 109, 2116–2123. [Google Scholar] [CrossRef] [Green Version]
- Sinaga, S. Principle of Plant Diseases; Jakarta: Penebar, Swadaya, 2006. [Google Scholar]
- Selim, E.M.; Ammar, M.; Amer, G.; Awad, H. Effect of some plant extracts, plant oils and Trichoderma spp. on tomato Fusarium wilt disease. Menoufia J. Plant Prot. 2020, 5, 155–167. [Google Scholar] [CrossRef]
- Danish, P.; Ali, Q.; Hafeez, M.; Malik, A. Antifungal and antibacterial activity of aloe vera plant extract. Biol. Clin. Sci. Res. J. 2020, 2020, 4. [Google Scholar] [CrossRef]
- Castellanos, L.M.; Olivas, N.A.; Ayala-Soto, J.; De La O Contreras, C.M.; Ortega, M.Z.; Salas, F.S.; Hernández-Ochoa, L. In vitro and in vivo antifungal activity of clove (Eugenia caryophyllata) and pepper (Piper nigrum L.) essential oils and functional extracts against Fusarium oxysporum and Aspergillus niger in tomato (Solanum lycopersicum L.). Int. J. Microbiol. 2020, 8, 1702037. [Google Scholar] [CrossRef] [PubMed]
- Suprapta, D.; Swari, I.G.; Arya, N.; Ohsawa, K. Pometia pinnata leaves extract to control light blight disease in potato. J. Int. Soc. Southeast Asian Agric. Sci. 2002, 8, 31–36. [Google Scholar]
- Darmadi, A.A.K.; Suprapta, D.N.; Temaja, I.; Swantara, I.M.D.; Sudirman, J.; Indonesia, D.B. Leaf extract of Cinnamomum burmanni blume effectively suppress the growth of Fusarium oxysporum f. sp. lycopersici the cause of Fusarium wilt disease on tomato. J. Biol. Agric. Healthc. 2015, 5, 131–137. [Google Scholar]
- Shivpuri, A.; Sharma, O.; Jhamaria, S. Fungitoxic properties of plant extracts against pathogenic fungi. J. Mycol. Plant Pathol. 1997, 27, 29–31. [Google Scholar]
- Singh, J.K.; Kumar, M.; Kumar, S.; Kumar, A.; Mehta, N. Inhibitory effect of botanicals on growth and sporulation of Fusarium oxysporum inciting wilt of Chilli (Capsicum annuum L.). J. Pharmacogn. Phytochem. 2017, 6, 2199–2204. [Google Scholar]
- Hassanein, N.; Zeid, M.A.; Youssef, K.; Mahmoud, D. Efficacy of leaf extracts of neem (Azadirachta indica) and chinaberry (Melia azedrach) against early blight and wilt diseases of tomato. Aust. J. Basic Appl. Sci. 2008, 2, 763–772. [Google Scholar]
- Enespa, D.S.; Dwivedi, S. Effectiveness of some antagonistic fungi and botanicals against Fusarium solani and Fusarium oxysporum f. sp. lycopersici infecting brinjal and tomato plants. Asian. J. Plant Pathol. 2014, 8, 18–25. [Google Scholar]
- El-Ghany, A.; Roushdy, M.; Mohamed, A. Efficacy of certain plant extracts as safe fungicides against phytopathogenic and mycotoxigenic fungi. J. Agric. Biol. Sci. 2015, 1, 71–75. [Google Scholar]
- Rai, V.R.; Lokesh, S.; Khan, A. Occurrence and Management of some Seedborne Fungal Pathogens of Maize and Sorghum in vitro. Seed Res. New Delhi 2002, 30, 112–117. [Google Scholar]
- Ramaiah, A.K.; Garampalli, R.K.H. In vitro antifungal activity of some plant extracts against Fusarium oxysporum f. sp. lycopersici. Asian J. Plant Sci. Res. 2015, 5, 22–27. [Google Scholar]
- Yelmame, M.; Mehta, B.; Deshmukh, A.; Patil, V. Evaluation of some organic extracts in in vitro to control Fusarium solani causing chilli wilt. Int. J. Pharma Bio. Sci. 2010, 1, B551 ref 8. [Google Scholar]
- Arora, R.; Singh, S.; Sharma, R. Neem Leaves: Indian Herbal Medicine. In Botanical Medicine in Clinical Practice; CABI: Wallingford, UK, 2008; pp. 85–98. [Google Scholar]
- Atawodi, S.E.; Atawodi, J.C. Azadirachta indica (neem): A plant of multiple biological and pharmacological activities. Phytochem. Rev. 2009, 8, 601–620. [Google Scholar] [CrossRef]
- Brahmachari, G. Neem—An omnipotent plant: A retrospection. Chembiochem 2004, 5, 408–421. [Google Scholar] [CrossRef]
- Girish, K.; Shankara, B.S. Neem–a green treasure. Electron. J. Biol. 2008, 4, 102–111. [Google Scholar]
- Sarkar, K.; Bose, A.; Laskar, S.; Choudhuri, S.K.; Dey, S.; Roychowdhury, P.K.; Baral, R. Antibody response against neem leaf preparation recognizes carcinoembryonic antigen. Int. Immunopharmacol. 2007, 7, 306–312. [Google Scholar] [CrossRef]
- Tiwari, R.; Verma, A.K.; Chakraborty, S.; Dhama, K.; Singh, S.V. Neem (Azadirachta indica) and its potential for safeguarding health of animals and humans: A review. J. Biol. Sci. 2014, 14, 110. [Google Scholar] [CrossRef] [Green Version]
- Singh, U.; Maurya, S.; Singh, A.; Gohain, L. Foliar spray of aqueous extract of neem (Azadirachta indica) cake to control balsam (Impatiens balsaminia) powdery mildew. Arch. Phytopathol. Plant Prot. 2010, 43, 1056–1063. [Google Scholar] [CrossRef]
- Mangi, A.H.; Jiskani, A.M.; Khaskhell, M.I.; Jiskani, M.M.; Poussio, G.B.; Qambrani, R.A.; Mahar, M.A. Evaluation of Neem Products Against Damping of Disease of Tomato. Pak. J. Phytopathol. 2021, 33, 37–45. [Google Scholar] [CrossRef]
- Vani, M.S.; Kumar, S.; Gulya, R. In vitro evaluation of fungicides and plant extracts against Fusarium oxysporum causing wilt of mungbean. J. Pharm. Innov. 2019, 8, 297–302. [Google Scholar]
- Chen, Y.; Zeng, H.; Tian, J.; Ban, X.; Ma, B.; Wang, Y. Antifungal mechanism of essential oil from Anethum graveolens seeds against Candida albicans. J. Med. Microbiol. 2013, 62, 1175–1183. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; Coppola, R.; De Feo, V. Essential oils and antifungal activity. Pharm. J. 2017, 10, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gogoi, P.; Baruah, P.; Nath, S.C. Antifungal Activity of the Essential Oil of Litsea cubeba. Pers. J. Essent. Oil Res. 1997, 9, 213–215. [Google Scholar] [CrossRef]
- Xia, Z.; Yang, J.; Li, P. Study on antifungal mechanism of Litsea cubeba oil in Candida albicans. Bull. Hunan Med. Univ. 1995, 20, 107–108. [Google Scholar]
- Ark, P.A.; Thompson, J.P. Control of certain diseases of plants with antibiotics from garlic (Allium sativum L.). Plant Dis. Rep. 1959, 43, 276–282. [Google Scholar]
- Council, N.R. Regulating Pesticides in Food: The Delaney Paradox; National Academies Press: Cambridge, MA, USA, 1987. [Google Scholar]
- Perello, A.E.; Noll, U.; Slusarenko, A.J. In vitro efficacy of garlic extracts to control fungal pathogens of wheat. J. Med. Plant Res. 2013, 7, 1809–1817. [Google Scholar]
- Wilson, C.; Solar, J.; El Ghaouth, A.; Wisniewski, M. Rapid evaluation of plant extracts and essential oils for antifungal activity against Botrytis cinerea. Plant Dis. 1997, 81, 204–210. [Google Scholar] [CrossRef] [Green Version]
- Cavallito, C.J.; Bailey, J.H. Allicin, the antibacterial principle of Allium sativum. I. Isolation, physical properties and antibacterial action. J. Am. Chem. Soc. 1944, 66, 1950–1951. [Google Scholar] [CrossRef]
- Muhsin, T.M.; Al-Zubaidy, S.R.; Ali, E.T. Effect of garlic bulb extract on the growth and enzymatic activities of rhizosphere and rhizoplane fungi. Mycopathologia 2001, 152, 143–146. [Google Scholar] [CrossRef]
- Achimón, F.; Brito, V.D.; Pizzolitto, R.P.; Sanchez, A.R.; Gómez, E.A.; Zygadlo, J.A. Chemical composition and antifungal properties of commercial essential oils against the maize phytopathogenic fungus Fusarium verticillioides. Rev. Argent. Microbiol. 2021, 53, 292–303. [Google Scholar] [CrossRef]
- Ghabrial, S.A.; Caston, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. J. Virol. 2015, 479–480, 356–368. [Google Scholar] [CrossRef] [Green Version]
- Ghabrial, S.A.; Suzuki, N. Viruses of plant pathogenic fungi. Annu. Rev. Phytopathol. 2009, 47, 353–384. [Google Scholar] [CrossRef]
- Pearson, M.N.; Beever, R.E.; Boine, B.; Arthur, K. Mycoviruses of filamentous fungi and their relevance to plant pathology. Mol. Plant Pathol. 2009, 10, 115–128. [Google Scholar] [CrossRef]
- Xie, J.; Jiang, D. New insights into mycoviruses and exploration for the biological control of crop fungal diseases. Annu. Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Cheng, J.; Fu, Y.; Chen, T.; Jiang, D.; Ghabrial, S.A.; Xie, J. Virus-mediated suppression of host non-self recognition facilitates horizontal transmission of heterologous viruses. PLoS Pathog. 2017, 3, e1006234. [Google Scholar] [CrossRef] [Green Version]
- Darissa, O.; Adam, G.; Schäfer, W. A dsRNA mycovirus causes hypovirulence of Fusarium graminearum to wheat and maize. Eur. J. Plant Pathol. 2012, 134, 181–189. [Google Scholar] [CrossRef]
- Lee, K.M.; Cho, W.K.; Yu, J.; Son, M.; Choi, H.; Min, K.; Lee, Y.W.; Kim, K.H. A comparison of transcriptional patterns and mycological phenotypes following infection of Fusarium graminearum by four mycoviruses. PLoS ONE 2014, 9, e100989. [Google Scholar] [CrossRef]
- Lemus-Minor, C.G.; Cañizares, M.C.; García-Pedrajas, M.D.; Pérez-Artés, E.J. Fusarium oxysporum f. sp. dianthi virus 1 accumulation is correlated with changes in virulence and other phenotypic traits of its fungal host. Phytopathology 2018, 108, 957–963. [Google Scholar] [CrossRef] [Green Version]
- Aydi Ben Abdallah, R.; Jabnoun-Khiareddine, H.; Nefzi, A.; Mokni-Tlili, S.; Daami-Remadi, M. Biocontrol of Fusarium wilt and growth promotion of tomato plants using endophytic bacteria isolated from Solanum elaeagnifolium stems. J. Phytopathol. 2016, 164, 811–824. [Google Scholar] [CrossRef]
- Wang, S.; Liang, Y.; Shen, T.; Yang, H.; Shen, B. Biological characteristics of Streptomyces albospinus CT205 and its biocontrol potential against cucumber Fusarium wilt. Biocontrol Sci. Technol. 2016, 26, 951–963. [Google Scholar] [CrossRef]
- Zhao, Y.; Selvaraj, J.N.; Xing, F.; Zhou, L.; Wang, Y.; Song, H.; Tan, X.; Sun, L.; Sangare, L.; Folly, Y.M.E. Antagonistic action of Bacillus subtilis strain SG6 on Fusarium graminearum. PLoS ONE 2014, 9, e92486. [Google Scholar] [CrossRef]
- Shi, L.; Du, N.; Shu, S.; Sun, J.; Li, S.; Guo, S. Paenibacillus polymyxa NSY50 suppresses Fusarium wilt in cucumbers by regulating the rhizospheric microbial community. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- García-Rincón, J.; Vega-Pérez, J.; Guerra-Sanchez, M.G.; Hernandez-Lauzardo, A.N.; Peña-Díaz, A.; Valle, M.G.V.-D. Effect of chitosan on growth and plasma membrane properties of Rhizopus stolonifer (Ehrenb.: Fr.) Vuill. Biochem. Physiol. 2010, 97, 275–278. [Google Scholar] [CrossRef]
- Liu, H.; Du, Y.; Wang, X.; Sun, L.J. Chitosan kills bacteria through cell membrane damage. Int. J. Food Microbiol. 2004, 95, 147–155. [Google Scholar] [CrossRef]
- Kong, M.; Chen, X.G.; Xing, K.; Park, H.J. Antimicrobial properties of chitosan and mode of action: A state of the art review. Int. J. Food Microbiol. 2010, 144, 51–63. [Google Scholar] [CrossRef]
- Roller, S.; Covill, N. The antifungal properties of chitosan in laboratory media and apple juice. Int. J. Food Microbiol. 1999, 47, 67–77. [Google Scholar] [CrossRef]
- Guo, Z.; Chen, R.; Xing, R.; Liu, S.; Yu, H.; Wang, P.; Li, C.; Li, P. Novel derivatives of chitosan and their antifungal activities in vitro. Carbohydr. Res. 2006, 341, 351–354. [Google Scholar] [CrossRef]
- Tikhonov, V.; Stepnova, E.; Babak, V.; Yamskov, I.; Palma-Guerrero, J.; Jasson, H.; lopez-Lorca, l.V.; Salinas, J.; Gerasimenko, D.V.; Avdienko, I.D.; et al. Bactericidal and antifungal activities of a low molecular weight chitosan and its n-/2 (3)-(Dodec-2-Enyl) Succinoyl/-derivatives. Carbohydr. Polym. 2006, 64, 66–72. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, X.; Han, X.; Du, Y. Antifungal activity of oligochitosan against Phytophthora capsici and other plant pathogenic fungi in vitro. Pestic. Biochem. Physiol. 2007, 87, 220–228. [Google Scholar] [CrossRef]
- Anirban, B. Fungicidal potential of chitosan against phytopathogenic Fusarium solani. J. Exp. Biol. Agric. Sci. 2013, 1, 258–263. [Google Scholar]
- Ramteke, P.K. Effect of resistance inducers on in vitro inhibition of mycelial growth and sporulation of Fusarium solani causing root rot of fenugreek. Plant Pathol. Quar. 2019, 9, 198–209. [Google Scholar] [CrossRef]
- Liang, R.; Li, X.; Yuan, W.; Jin, S.; Hou, S.; Wang, M.; Wang, H. Antifungal activity of nanochitin whisker against crown rot diseases of wheat. J. Agric. Food Chem. 2018, 66, 9907–9913. [Google Scholar] [CrossRef]
- Davis, A.J.; Grant, B.R. The effect of phosphonate on the sporulation of Fusarium oxysporum f.sp. cubense. Australas. Plant Pathol. 1996, 25, 31–35. [Google Scholar] [CrossRef]
- Lin, H.; Travisano, M.; Kazlauskas, R.J. The Fungus Trichoderma Regulates Submerged Conidiation Using the Steroid Pregnenolone. ACS Chem. Biol. 2016, 11, 2568–2575. [Google Scholar] [CrossRef]
- Schreiber, K.J.; Nasmith, C.G.; Allard, G.; Singh, J.; Subramaniam, R.; Desveaux, D. Found in translation: High-throughput chemical screening in Arabidopsis thaliana identifies small molecules that reduce Fusarium head blight disease in wheat. Mol. Plant-Microbe Interact. 2011, 24, 640–648. [Google Scholar] [CrossRef] [Green Version]
- Faurie, B.; Cluzet, S.; Corio-Costet, M.F.; Mérillon, J.M. Methyl jasmonate/ethephon cotreatment synergistically induces stilbene production in" Vitis vinifera" cell suspensions but fails to trigger resistance to Erysiphe necator. J. Int. Sci. Vigne. Vin. 2009, 43, 99–110. [Google Scholar] [CrossRef]
- Konan, Y.K.F.; Kouassi, K.M.; Kouakou, K.L.; Koffi, E.; Kouassi, K.N.; Sekou, D.; Kone, M.; Kouakou, T.H. Effect of methyl jasmonate on phytoalexins biosynthesis and induced disease resistance to Fusarium oxysporum f. sp. Vasinfectum in cotton (Gossypium hirsutum L.). Int. J. Agron. 2014, 11, 806439. [Google Scholar]
- Wasternack, C. Jasmonates: An update on biosynthesis, signal transduction and action in plant stress response, growth and development. Ann. Bot. 2007, 100, 681–697. [Google Scholar] [CrossRef] [Green Version]
- Larronde, F.; Gaudillère, J.P.; Krisa, S.; Decendit, A.; Deffieux, G.; Mérillon, J.M. Airborne methyl jasmonate induces stilbene accumulation in leaves and berries of grapevine plants. Am. J. Enol. Vitic. 2003, 54, 63–66. [Google Scholar] [CrossRef]
- Wang, Q.; Qiu, B.; Li, S.; Zhang, Y.; Cui, X.; Ge, F.; Liu, D. A methyl jasmonate induced defensin like protein from Panax notoginseng confers resistance against Fusarium solani in transgenic tobacco. Biol. Plant 2019, 63, 797–807. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Monaim, M.F.; Abdel-Gaid, M.A.W.; Armanious, A.H. Effect of chemical inducers on root rot and wilt diseases, yield and quality of tomato. Int. J. Agric. Sci. 2012, 2, 211–220. [Google Scholar]
- Wu, H.S.; Raza, W.; Liu, D.Y.; Wu, C.L.; Mao, Z.S.; Xu, Y.C.; Shen, Q.-R.J. Allelopathic impact of artificially applied coumarin on Fusarium oxysporum f. sp. niveum. World J. Microbiol. Biotechnol. 2008, 24, 1297–1304. [Google Scholar] [CrossRef]
- Ali, I.; Husnain, T.; Riazuddin, S. RNA interference: The story of gene silencing in plants and humans. Biotechnol. Adv. 2008, 26, 202–209. [Google Scholar]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Jinek, M.; Doudna, J.A. A three-dimensional view of the molecular machinery of RNA interference. Nature 2009, 457, 405–412. [Google Scholar] [CrossRef]
- Watson, J.M.; Fusaro, A.F.; Wang, M.; Waterhouse, P.M. RNA silencing platforms in plants. FEMS. Lett. 2005, 579, 5982–5987. [Google Scholar] [CrossRef]
- Napoli, C.; Lemieux, C.; Jorgensen, R. Introduction of a chimeric chalcone synthase gene into petunia results in reversible co-suppression of homologous genes in trans. Plant Cell 1990, 2, 279–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fairbairn, D.J.; Cavallaro, A.S.; Bernard, M.; Mahalinga-Iyer, J.; Graham, M.W.; Botella, J.R. Host-delivered RNAi: An effective strategy to silence genes in plant parasitic nematodes. Planta 2007, 226, 1525–1533. [Google Scholar] [CrossRef]
- Hu, Z.; Parekh, U.; Maruta, N.; Trusov, Y.; Botella, J.R. Down-regulation of Fusarium oxysporum endogenous genes by host-delivered RNA interference enhances disease resistance. Front. Chem. 2015, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Tinoco, M.L.P.; Dias, B.; Dall’Astta, R.C.; Pamphile, J.A.; Aragão, F. In vivo trans-specific gene silencing in fungal cells by in planta expression of a double-stranded RNA. BioMed Central. 2010, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Koch, A.; Kumar, N.; Weber, L.; Keller, H.; Imani, J.; Kogel, K.H. Host-induced gene silencing of cytochrome P450 lanosterol C14α-demethylase–encoding genes confers strong resistance to Fusarium species. Proc. Natl. Acad. Sci. USA 2013, 110, 19324–19329. [Google Scholar] [CrossRef] [Green Version]
- Takano, Y.; Kikuchi, T.; Kubo, Y.; Hamer, J.E.; Mise, K.; Furusawa, I. The Colletotrichum lagenarium MAP kinase gene CMK1 regulates diverse aspects of fungal pathogenesis. Mol. Plant-Microbe Interact 2000, 13, 374–383. [Google Scholar] [CrossRef] [Green Version]
- Pareek, M.; Rajam, M.V. RNAi-mediated silencing of MAP kinase signalling genes (Fmk1, Hog1, and Pbs2) in Fusarium oxysporum reduces pathogenesis on tomato plants. Fungal Biol. 2017, 121, 775–784. [Google Scholar] [CrossRef]
- Zheng, D.; Zhang, S.; Zhou, X.; Wang, C.; Xiang, P.; Zheng, Q.; Xu, J.R. The FgHOG1 pathway regulates hyphal growth, stress responses, and plant infection in Fusarium graminearum. PLoS ONE 2012, 7, e49495. [Google Scholar] [CrossRef]
- Shanmugam, V.; Sharma, V.; Bharti, P.; Jyoti, P.; Yadav, S.K.; Aggarwal, R.; Jain, S. RNAi induced silencing of pathogenicity genes of Fusarium spp. for vascular wilt management in tomato. Ann. Microbiol. 2017, 67, 359–369. [Google Scholar] [CrossRef]
- Fernandes, J.; Angelo, P.; Cruz, J.; Santos, J.; Sousa, N.R.; Silva, G. Post-transcriptional silencing of the SGE1 gene induced by a dsRNA hairpin in Fusarium oxysporum f. sp cubense, the causal agent of Panama disease. Genet. Mol. Res. 2016, 15, 1–142. [Google Scholar] [CrossRef]
- Singh, N.; Mukherjee, S.K.; Rajam, M.V. Silencing of the ornithine decarboxylase gene of Fusarium oxysporum f. sp. lycopersici by host-induced RNAi confers resistance to Fusarium wilt in tomato. Plant Mol. Biol. Rep 2020, 38, 419–429. [Google Scholar] [CrossRef]
- Yin, G.; Sun, Z.; Liu, N.; Zhang, L.; Song, Y.; Zhu, C.; Wen, F. Production of double-stranded RNA for interference with TMV infection utilizing a bacterial prokaryotic expression system. Appl. Microbiol. Biotechnol. 2009, 84, 323–333. [Google Scholar] [CrossRef]
- Dalakouras, A.; Wassenegger, M.; McMillan, J.N.; Cardoza, V.; Maegele, I.; Dadami, E.; Runne, M.; Krczal, G.; Wassenegger, M. Induction of silencing in plants by high-pressure spraying of in vitro-synthesized small RNAs. Front. Plant Sci. 2016, 7, 1327. [Google Scholar] [CrossRef] [Green Version]
- Gebremichael, D.E.; Haile, Z.M.; Negrini, F.; Sabbadini, S.; Capriotti, L.; Mezzetti, B.; Baraldi, E. RNA interference strategies for future management of plant pathogenic fungi: Prospects and challenges. Plants 2021, 10, 650. [Google Scholar] [CrossRef]
- Koch, A.; Biedenkopf, D.; Furch, A.; Weber, L.; Rossbach, O.; Abdellatef, E.; Linicus, L.; Johannsmeier, J.; Jelonek, L.; Goesmann, A.; et al. An RNAi-based control of Fusarium graminearum infections through spraying of long dsRNAs involves a plant passage and is controlled by the fungal silencing machinery. PLoS Pathog. 2016, 12, e1005901. [Google Scholar] [CrossRef]
- Song, X.S.; Gu, K.X.; Duan, X.X.; Xiao, X.M.; Hou, Y.P.; Duan, Y.B.; Wang, J.X.; Yu, N.; Zhou, M.G. Secondary amplification of siRNA machinery limits the application of spray-induced gene silencing. Mole. Plant Pathol. 2018, 19, 2543–2560. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ajmal, M.; Hussain, A.; Ali, A.; Chen, H.; Lin, H. Strategies for Controlling the Sporulation in Fusarium spp. J. Fungi 2023, 9, 10. https://doi.org/10.3390/jof9010010
Ajmal M, Hussain A, Ali A, Chen H, Lin H. Strategies for Controlling the Sporulation in Fusarium spp. Journal of Fungi. 2023; 9(1):10. https://doi.org/10.3390/jof9010010
Chicago/Turabian StyleAjmal, Maria, Adil Hussain, Asad Ali, Hongge Chen, and Hui Lin. 2023. "Strategies for Controlling the Sporulation in Fusarium spp." Journal of Fungi 9, no. 1: 10. https://doi.org/10.3390/jof9010010