
Function of the Mitochondrial Transport Protein BcMtp1 in Regulating Vegetative Development, Asexual Reproduction, Stress Response, Fungicide Sensitivity, and Virulence of Botrytis cinerea

Abstract
:1. Introduction
2. Materials and methods
2.1. Strain, Medium, and Incubation Conditions
2.2. Sequence Analysis of BcMtp1
2.3. The Deletion and Complementation of BcMtp1
2.4. Standard Molecular Methods
2.5. qRT-PCR
2.6. Fungicide Sensitivity Assay
2.7. Glycerol Accumulation Assay
2.8. Virulence Assays
2.9. Statistical Analysis
3. Results
3.1. Identification of the BcMtp1 Gene
3.2. Deletion of BcMtp1 and Complementation
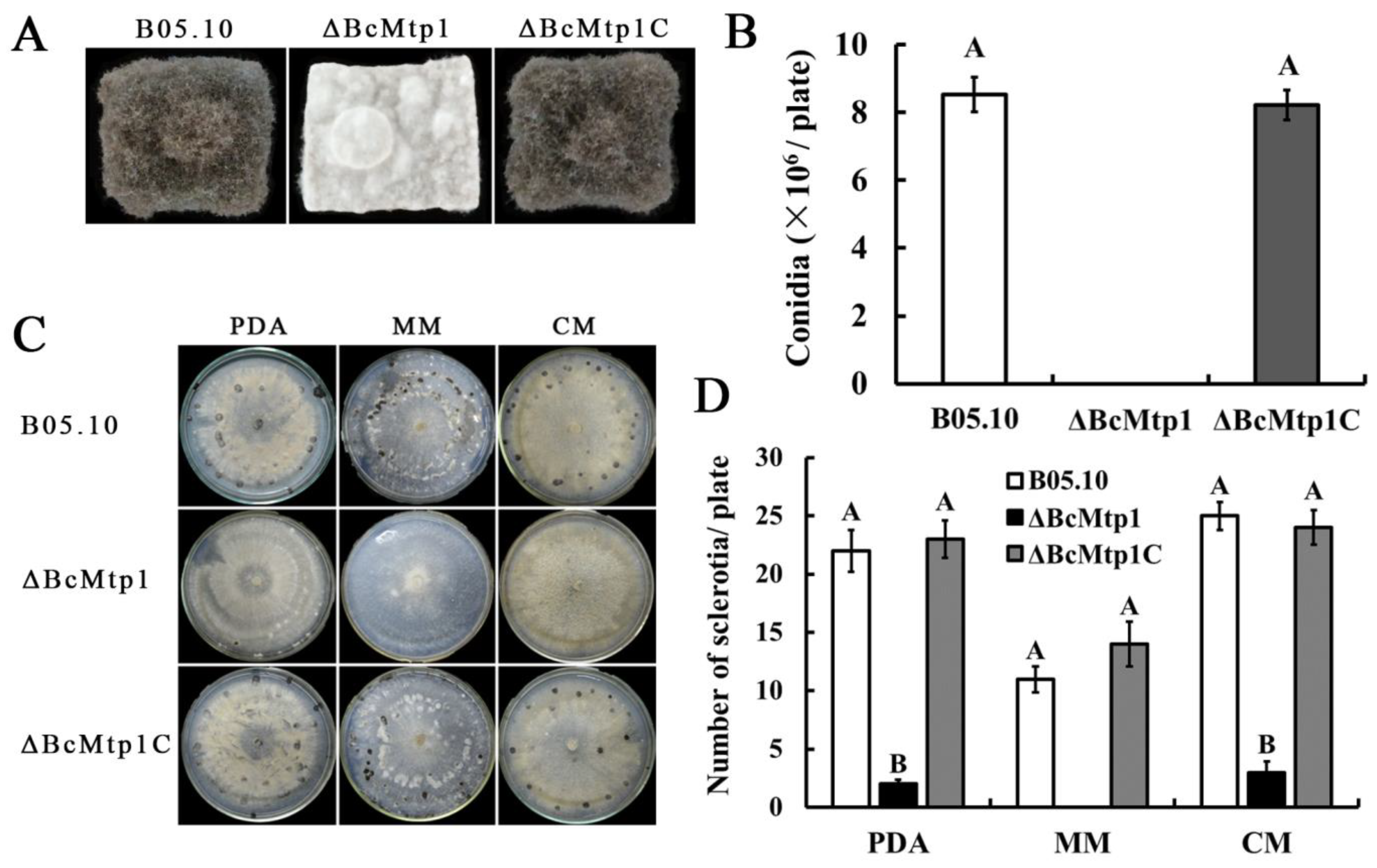
3.3. BcMtp1 Is Involved in Vegetative Growth
3.4. BcMtp1 Is Required for Conidiation and Sclerotia Production
3.5. The Role of BcMtp1 in Stress Response
3.6. BcMtp1 Is Required for Glycerol Accumulation
3.7. BcMtp1 Is Involved in the Transcriptional Regulation of the Genes Related to Environmental Stress
3.8. Involvement of BcMtp1 in the Sensitivity of B. cinerea to Fungicides
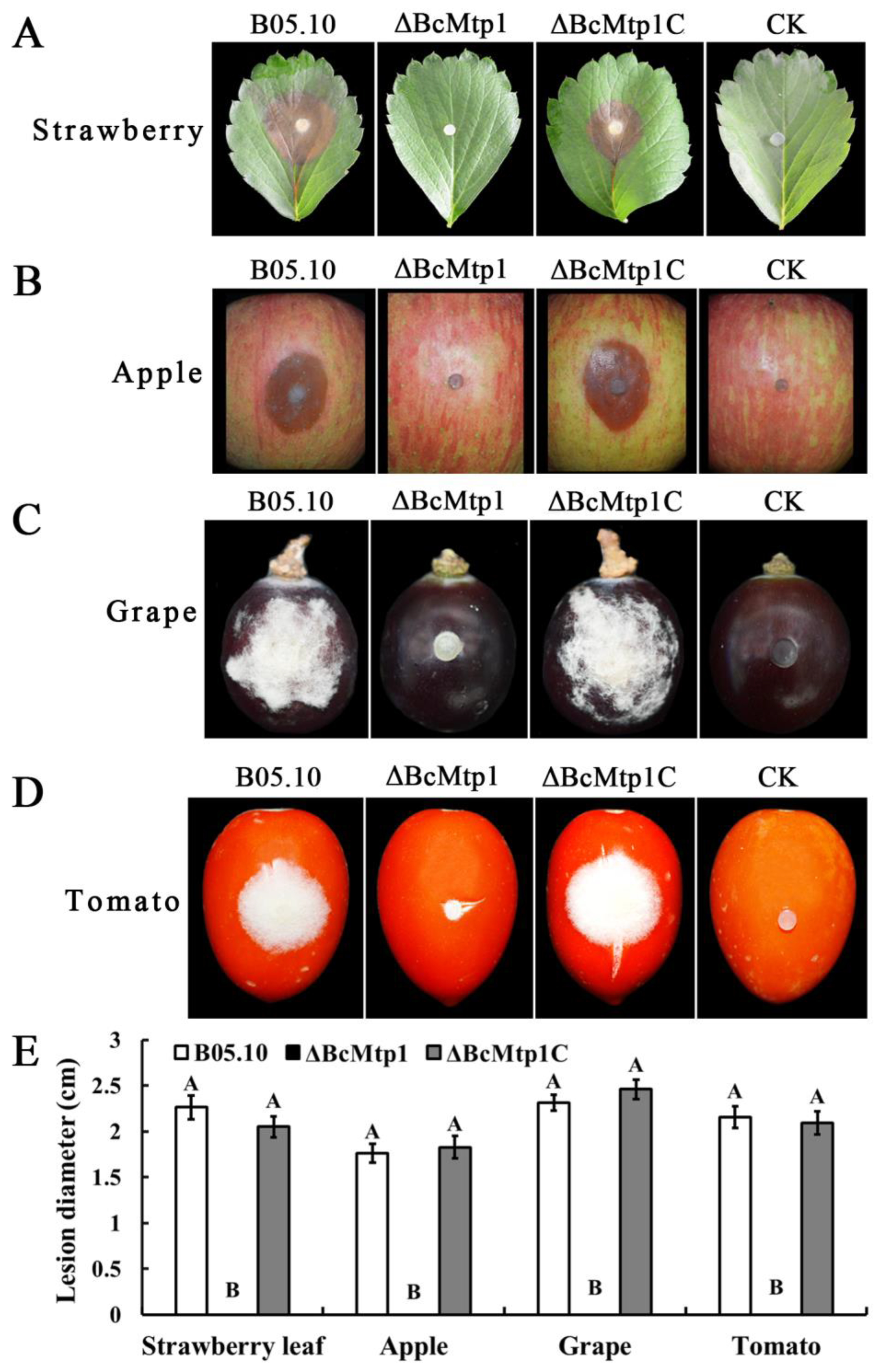
3.9. BcMtp1 Is Required for Virulence of B. cinerea
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fillinger, S.; Elad, Y. Botrytis-the Fungus, the Pathogen and Its Management in Agricultural Systems; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; Van Kan, J.A. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Amselem, J.; Cuomo, C.A.; van Kan, J.A.; Viaud, M.; Benito, E.P.; Couloux, A.; Coutinho, P.M.; de Vries, R.P.; Dyer, P.S.; Fillinger, S. Genomic analysis of the necrotrophic fungal pathogens Sclerotinia sclerotiorum and Botrytis cinerea. PLoS Genet. 2011, 7, e1002230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neupert, W. Protein import into mitochondria. PLoS Genet. 1997, 66, 863–917. [Google Scholar] [CrossRef] [PubMed]
- Newmeyer, D.D.; Ferguson-Miller, S. Mitochondria: Releasing power for life and unleashing the machineries of death. Cell 2003, 112, 481–490. [Google Scholar] [CrossRef] [Green Version]
- Green, D.R.; Kroemer, G.J.S. The pathophysiology of mitochondrial cell death. Science 2004, 305, 626–629. [Google Scholar] [CrossRef]
- Lill, R.; Mühlenhoff, U. Iron–sulfur-protein biogenesis in eukaryotes. Trends Biochem. Sci. 2005, 30, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Bratic, I.; Trifunovic, A. Mitochondrial energy metabolism and ageing. Biochim. Biophys. Acta (BBA)-Bioenerg. 2010, 1797, 961–967. [Google Scholar] [CrossRef] [Green Version]
- Reid, R.; Moyle, J.; Mitchell, P. Synthesis of adenosine triphosphate by a protonmotive force in rat liver mitochondria. Nature 1966, 212, 257–258. [Google Scholar] [CrossRef]
- Palmieri, F.; Agrimi, G.; Blanco, E.; Castegna, A.; Di Noia, M.A.; Iacobazzi, V.; Lasorsa, F.M.; Marobbio, C.M.; Palmieri, L.; Scarcia, P. Identification of mitochondrial carriers in Saccharomyces cerevisiae by transport assay of reconstituted recombinant proteins. Biochim. Biophys. Acta (BBA)-Bioenerg. 2006, 1757, 1249–1262. [Google Scholar] [CrossRef]
- Palmieri, F. The mitochondrial transporter family SLC25: Identification, properties and physiopathology. Mol. Asp. Med. 2013, 34, 465–484. [Google Scholar] [CrossRef]
- Marobbio, C.M.; Giannuzzi, G.; Paradies, E.; Pierri, C.L.; Palmieri, F.J. α-Isopropylmalate, a leucine biosynthesis intermediate in yeast, is transported by the mitochondrial oxalacetate carrier. J. Biol. Chem. 2008, 283, 28445–28453. [Google Scholar] [CrossRef] [Green Version]
- Traba, J.; Froschauer, E.M.; Wiesenberger, G.; Satrústegui, J.; Del Arco, A. Yeast mitochondria import ATP through the calcium-dependent ATP-Mg/Pi carrier Sal1p, and are ATP consumers during aerobic growth in glucose. Mol. Microbiol. 2008, 69, 570–585. [Google Scholar] [CrossRef]
- Castegna, A.; Scarcia, P.; Agrimi, G.; Palmieri, L.; Rottensteiner, H.; Spera, I.; Germinario, L.; Palmieri, F.J. Identification and functional characterization of a novel mitochondrial carrier for citrate and oxoglutarate in Saccharomyces cerevisiae. J. Biol. Chem. 2010, 285, 17359–17370. [Google Scholar] [CrossRef] [Green Version]
- Pebay-Peyroula, E.; Dahout-Gonzalez, C.; Kahn, R.; Trézéguet, V.; Lauquin, G.J.-M.; Brandolin, G. Structure of mitochondrial ADP/ATP carrier in complex with carboxyatractyloside. Nature 2003, 426, 39–44. [Google Scholar] [CrossRef]
- Nury, H.; Dahout-Gonzalez, C.; Trezeguet, V.; Lauquin, G.; Brandolin, G.a.; Pebay-Peyroula, E. Relations between structure and function of the mitochondrial ADP/ATP carrier. Annu. Rev. Biochem. 2006, 75, 713–741. [Google Scholar] [CrossRef] [Green Version]
- Gawaz, M.; Douglas, M.; Klingenberg, M.J. Structure-function studies of adenine nucleotide transport in mitochondria. II. Biochemical analysis of distinct AAC1 and AAC2 proteins in yeast. J. Biol. Chem. 1990, 265, 14202–14208. [Google Scholar] [CrossRef]
- Drgon, T.; Sabova, L.; Gavurnikova, G.; Kolarov, J. Yeast ADP/ATP carrier (AAC) proteins exhibit similar enzymatic properties but their deletion produces different phenotypes. FEBS Lett. 1992, 304, 277–280. [Google Scholar] [CrossRef] [Green Version]
- Lawson, J.; Douglas, M.G.J. Separate genes encode functionally equivalent ADP/ATP carrier proteins in Saccharomyces cerevisiae. Isolation and analysis of AAC2. J. Biol. Chem. 1988, 263, 14812–14818. [Google Scholar] [CrossRef]
- Adrian, G.; McCammon, M.; Montgomery, D.; Douglas, M. Sequences required for delivery and localization of the ADP/ATP translocator to the mitochondrial inner membrane. Mol. Cell. Biol. 1986, 6, 626–634. [Google Scholar]
- Fontanesi, F.; Palmieri, L.; Scarcia, P.; Lodi, T.; Donnini, C.; Limongelli, A.; Tiranti, V.; Zeviani, M.; Ferrero, I.; Viola, A.M. Mutations in AAC2, equivalent to human adPEO-associated ANT1 mutations, lead to defective oxidative phosphorylation in Saccharomyces cerevisiae and affect mitochondrial DNA stability. Hum. Mol. Genet. 2004, 13, 923–934. [Google Scholar] [CrossRef] [PubMed]
- Gavurníková, G.; Šabova, L.; KiŠŠová, I.; Haviernik, P.; Kolarov, J. Transcription of the AAC1 gene encoding an isoform of mitochondrial ADP/ATP carrier in Saccharomyces cerevisiae is regulated by oxygen in a heme-independent manner. Eur. J. Biochem. 1996, 239, 759–763. [Google Scholar] [CrossRef] [PubMed]
- Kolarov, J.; Kolarova, N.; Nelson, N.J. A third ADP/ATP translocator gene in yeast. J. Biol. Chem. 1990, 265, 12711–12716. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.J.; Clark-Walker, G.D. The petite mutation in yeasts: 50 years on. Int. Rev. Cytol. 1999, 194, 197–238. [Google Scholar]
- Palmieri, F.; Pierri, C.L.; De Grassi, A.; Nunes-Nesi, A.; Fernie, A.R. Evolution, structure and function of mitochondrial carriers: A review with new insights. Plant J. 2011, 66, 161–181. [Google Scholar] [CrossRef]
- Palmieri, L.; Rottensteiner, H.; Girzalsky, W.; Scarcia, P.; Palmieri, F.; Erdmann, R. Identification and functional reconstitution of the yeast peroxisomal adenine nucleotide transporter. EMBO J. 2001, 20, 5049–5059. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, L.; Palmieri, F.; Runswick, M.J.; Walker, J.E. Identification by bacterial expression and functional reconstitution of the yeast genomic sequence encoding the mitochondrial dicarboxylate carrier protein. FEBS Lett. 1996, 399, 299–302. [Google Scholar] [CrossRef] [Green Version]
- Todisco, S.; Di Noia, M.A.; Castegna, A.; Lasorsa, F.M.; Paradies, E.; Palmieri, F. The Saccharomyces cerevisiae gene YPR011c encodes a mitochondrial transporter of adenosine 5′-phosphosulfate and 3′-phospho-adenosine 5′-phosphosulfate. Biochim. Biophys. Acta (BBA)-Bioenerg. 2014, 1837, 326–334. [Google Scholar] [CrossRef] [Green Version]
- An, B.; Li, B.; Qin, G.; Tian, S. Function of small GTPase Rho3 in regulating growth, conidiation and virulence of Botrytis cinerea. Fungal Genet. Biol. 2015, 75, 46–55. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, W.; Shao, W.; Tan, S.; Shi, D.; Ma, H.; Chen, C. FaSmi1 Is Essential for the Vegetative Development, Asexual Reproduction, DON Production and Virulence of Fusarium asiaticum. J. Fungi 2022, 8, 1189. [Google Scholar] [CrossRef]
- Lalève, A.; Gamet, S.; Walker, A.S.; Debieu, D.; Toquin, V.; Fillinger, S. Site-directed mutagenesis of the P225, N230 and H272 residues of succinate dehydrogenase subunit B from Botrytis cinerea highlights different roles in enzyme activity and inhibitor binding. Environ. Microbiol. 2014, 16, 2253–2266. [Google Scholar] [CrossRef]
- Shao, W.; Lv, C.; Zhang, Y.; Wang, J.; Chen, C. Involvement of BcElp4 in vegetative development, various environmental stress response and virulence of Botrytis cinerea. Microb. Biotechnol. 2017, 10, 886–895. [Google Scholar] [CrossRef]
- Zhang, Y.; Mao, C.X.; Zhai, X.Y.; Jamieson, P.A.; Zhang, C.Q. Mutation in cyp51b and overexpression of cyp51a and cyp51b confer multiple resistant to DMIs fungicide prochloraz in Fusarium fujikuroi. Pest Manag. Sci. 2021, 77, 824–833. [Google Scholar] [CrossRef]
- Yu, J.-H.; Hamari, Z.; Han, K.-H.; Seo, J.-A.; Reyes-Domínguez, Y.; Scazzocchio, C. Double-joint PCR: A PCR-based molecular tool for gene manipulations in filamentous fungi. Fungal Genet. Biol. 2004, 41, 973–981. [Google Scholar] [CrossRef]
- Shao, W.; Yang, Y.; Zhang, Y.; Lv, C.; Ren, W.; Chen, C. Involvement of BcS tr2 in methionine biosynthesis, vegetative differentiation, multiple stress tolerance and virulence in Botrytis cinerea. Mol. Plant Pathol. 2016, 17, 438–447. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Yan, L.; Gu, Q.; Ma, Z. The mitogen-activated protein kinase kinase kinase BcOs4 is required for vegetative differentiation and pathogenicity in Botrytis cinerea. Appl. Microbiol. Biotechnol. 2012, 96, 481–492. [Google Scholar] [CrossRef]
- Fujita, A.; Misumi, Y. Fission yeast ucp3 gene encodes a putative Arf6 GTPase-activating protein. Mol. Biol. Rep. 2011, 38, 3875–3882. [Google Scholar] [CrossRef]
- Chen, X.J. Sal1p, a calcium-dependent carrier protein that suppresses an essential cellular function associated with the Aac2 isoform of ADP/ATP translocase in Saccharomyces cerevisiae. Genetics 2004, 167, 607–617. [Google Scholar] [CrossRef] [Green Version]
- Nair, N.; Nadtotchei, A.J. Sclerotia of Botrytis as a source of primary inoculum for bunch rot of grapes in New South Wales, Australia. J. Phytopathol. 1987, 119, 42–51. [Google Scholar] [CrossRef]
- De Vries, R.P.; Flitter, S.J.; Van De Vondervoort, P.J.; Chaveroche, M.K.; Fontaine, T.; Fillinger, S.; Ruijter, G.J.; D’Enfert, C.; Visser, J. Glycerol dehydrogenase, encoded by gldB is essential for osmotolerance in Aspergillus nidulans. Mol. Microbiol. 2003, 49, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Fillinger, S.; Ruijter, G.; Tamás, M.J.; Visser, J.; Thevelein, J.M.; d’Enfert, C. Molecular and physiological characterization of the NAD-dependent glycerol 3-phosphate dehydrogenase in the filamentous fungus Aspergillus nidulans. Mol. Microbiol. 2001, 39, 145–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, R.J.; Low, C.; Bottema, C.D.; Parks, L.W. Multiple functions for sterols in Saccharomyces cerevisiae. Biochim. Biophys. Acta (BBA)-Lipids Lipid Metab. 1985, 837, 336–343. [Google Scholar] [CrossRef]
- Segmüller, N.; Ellendorf, U.; Tudzynski, B.; Tudzynski, P. BcSAK1, a stress-activated mitogen-activated protein kinase, is involved in vegetative differentiation and pathogenicity in Botrytis cinerea. Eukaryot. Cell 2007, 6, 211–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Jiang, J.; Mayr, C.; Hahn, M.; Ma, Z. Involvement of two type 2 C protein phosphatases B c P tc1 and B c P tc3 in the regulation of multiple stress tolerance and virulence of Botrytis cinerea. Environ. Microbiol. 2013, 15, 2696–2711. [Google Scholar] [PubMed]
- Lu, J.M.-Y.; Deschenes, R.J.; Fassler, J.S. Saccharomyces cerevisiae histidine phosphotransferase Ypd1p shuttles between the nucleus and cytoplasm for SLN1-dependent phosphorylation of Ssk1p and Skn7p. Eukaryot. Cell 2003, 2, 1304–1314. [Google Scholar] [CrossRef] [Green Version]
- He, X.J.; Fassler, J.S. Identification of novel Yap1p and Skn7p binding sites involved in the oxidative stress response of Saccharomyces cerevisiae. Mol. Microbiol. 2005, 58, 1454–1467. [Google Scholar] [CrossRef] [Green Version]
- Kuge, S.; Jones, N.; Nomoto, A. Regulation of yAP-1 nuclear localization in response to oxidative stress. EMBO J. 1997, 16, 1710–1720. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.-J.; Miyoshi, H.; Komyoji, T.; Haga, T.; Fujita, T. Uncoupling activity of a newly developed fungicide, fluazinam [3-chloro-N-(3-chloro-2, 6-dinitro-4-trifluoromethylphenyl)-5-trifluoromethyl-2-pyridinamine]. Biochim. Biophys. Acta (BBA)-Bioenerg. 1991, 1056, 89–92. [Google Scholar] [CrossRef]
- Tiedemann, A. Evidence for a primary role of active oxygen species in induction of host cell death during infection of bean leaves with Botrytis cinerea. Physiol. Mol. Plant Pathol. 1997, 50, 151–166. [Google Scholar] [CrossRef]
- Govrin, E.M.; Levine, A. The hypersensitive response facilitates plant infection by the necrotrophic pathogen Botrytis cinerea. Curr. Biol. 2000, 10, 751–757. [Google Scholar] [CrossRef] [Green Version]
- Kużniak, E.; Skłodowska, M. Fungal pathogen-induced changes in the antioxidant systems of leaf peroxisomes from infected tomato plants. Planta 2005, 222, 192–200. [Google Scholar] [CrossRef]
- Viaud, M.; Fillinger, S.; Liu, W.; Polepalli, J.S.; Le Pêcheur, P.; Kunduru, A.R.; Leroux, P.; Legendre, L. A class III histidine kinase acts as a novel virulence factor in Botrytis cinerea. Mol. Plant-Microbe Interact. 2006, 19, 1042–1050. [Google Scholar] [CrossRef]
- Hou, J.; Feng, H.Q.; Chang, H.W.; Liu, Y.; Li, G.H.; Yang, S.; Yuan, Y.; Sun, J.; Zhu-Salzman, K.; Zhang, H.; et al. The H3K4 demethylase Jar1 orchestrates ROS production and expression of pathogenesis-related genes to facilitate Botrytis cinerea virulence. New Phytol. 2020, 930–947. [Google Scholar] [CrossRef]
- Temme, N.; Tudzynski, P. Does Botrytis cinerea ignore H2O2-induced oxidative stress during infection? Characterization of Botrytis activator protein 1. Mol. Plant-Microbe Interact. 2009, 22, 987–998. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | EC50 (μg mL−1) a | |||
|---|---|---|---|---|
| Fluazinam | Boscalid | Fludioxonil | Tebuconazole | |
| B05.10 | 0.02379 B | 3.0572 A | 0.0172 A | 0.3985 A |
| ΔBcMtp1 | 0.2176 A | 2.9714 A | 0.0169 A | 0.4057 A |
| ΔBcMtp1C | 0.02209 A | 3.0136 A | 0.0162 A | 0.3986 A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, W.; Zhang, Y.; Chen, C.; Xing, Y. Function of the Mitochondrial Transport Protein BcMtp1 in Regulating Vegetative Development, Asexual Reproduction, Stress Response, Fungicide Sensitivity, and Virulence of Botrytis cinerea. J. Fungi 2023, 9, 25. https://doi.org/10.3390/jof9010025
Shao W, Zhang Y, Chen C, Xing Y. Function of the Mitochondrial Transport Protein BcMtp1 in Regulating Vegetative Development, Asexual Reproduction, Stress Response, Fungicide Sensitivity, and Virulence of Botrytis cinerea. Journal of Fungi. 2023; 9(1):25. https://doi.org/10.3390/jof9010025
Chicago/Turabian StyleShao, Wenyong, Yu Zhang, Changjun Chen, and Yujun Xing. 2023. "Function of the Mitochondrial Transport Protein BcMtp1 in Regulating Vegetative Development, Asexual Reproduction, Stress Response, Fungicide Sensitivity, and Virulence of Botrytis cinerea" Journal of Fungi 9, no. 1: 25. https://doi.org/10.3390/jof9010025