
The Transcriptional Responses of Ectomycorrhizal Fungus, Cenococcum geophilum, to Drought Stress
 , and
, and 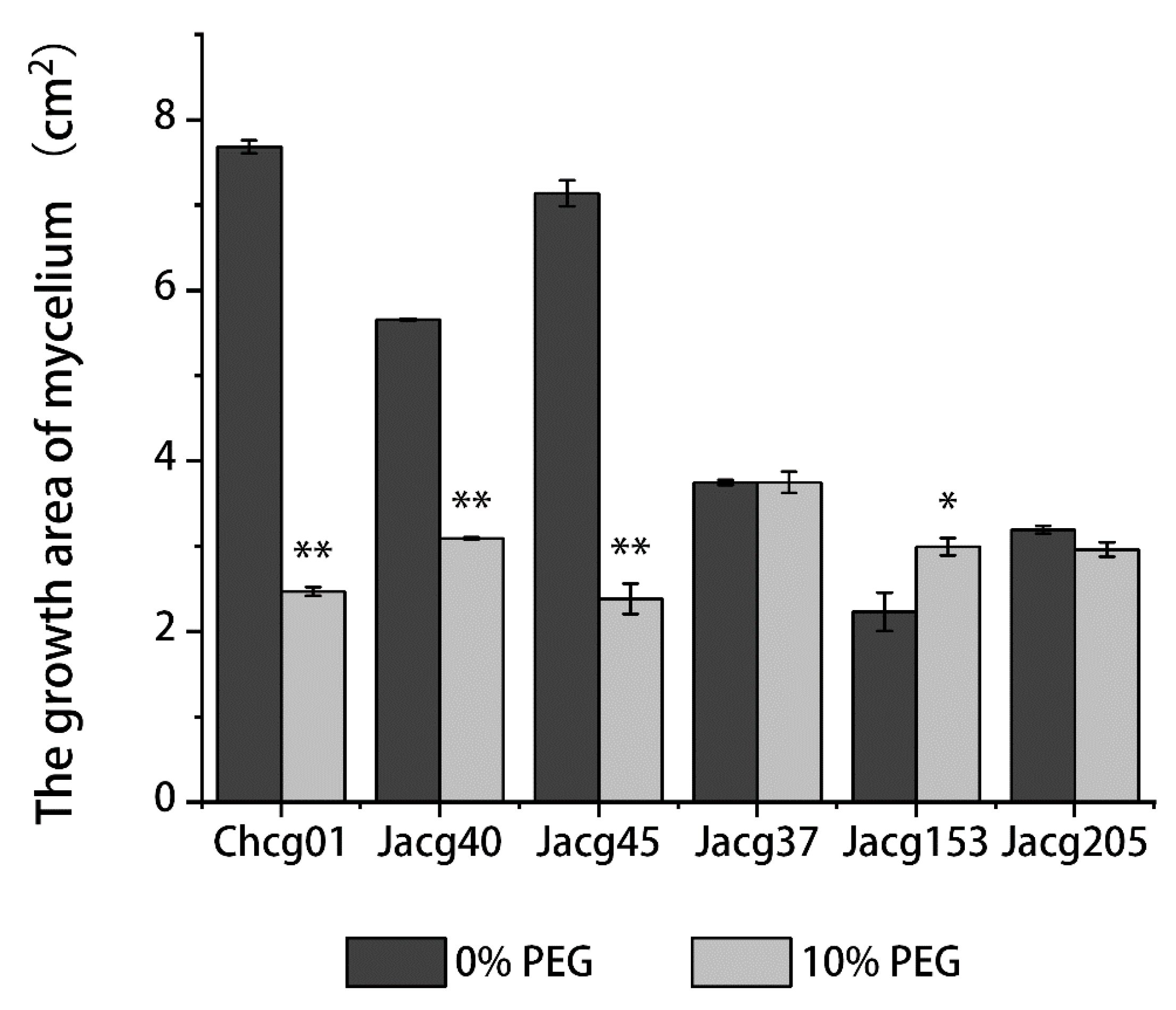{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mycelial Growth of Different Strains of C. geophilum under Drought Treatments
2.2. Preparation of Mycelia of C. geophilum for the Determination of Physiological Indicators
2.3. The Determination of Antioxidant Enzyme Activity in C. geophilum Mycelia
2.4. The Determination of Na, Ca, P, and K Concentrations in C. geophilum Mycelia
2.5. Preparation of Mycelia of C. geophilum Strains for RNA-Seq and Quantitative Real-Time PCR (qRT-PCR) Analyses
2.6. RNA-Seq Analysis
2.7. Experimental Validation of Gene Expression Using qRT-PCR
2.8. Statistical Analysis
3. Results
3.1. Mycelial Growth of Different C. geophilum Strains under Drought Treatment
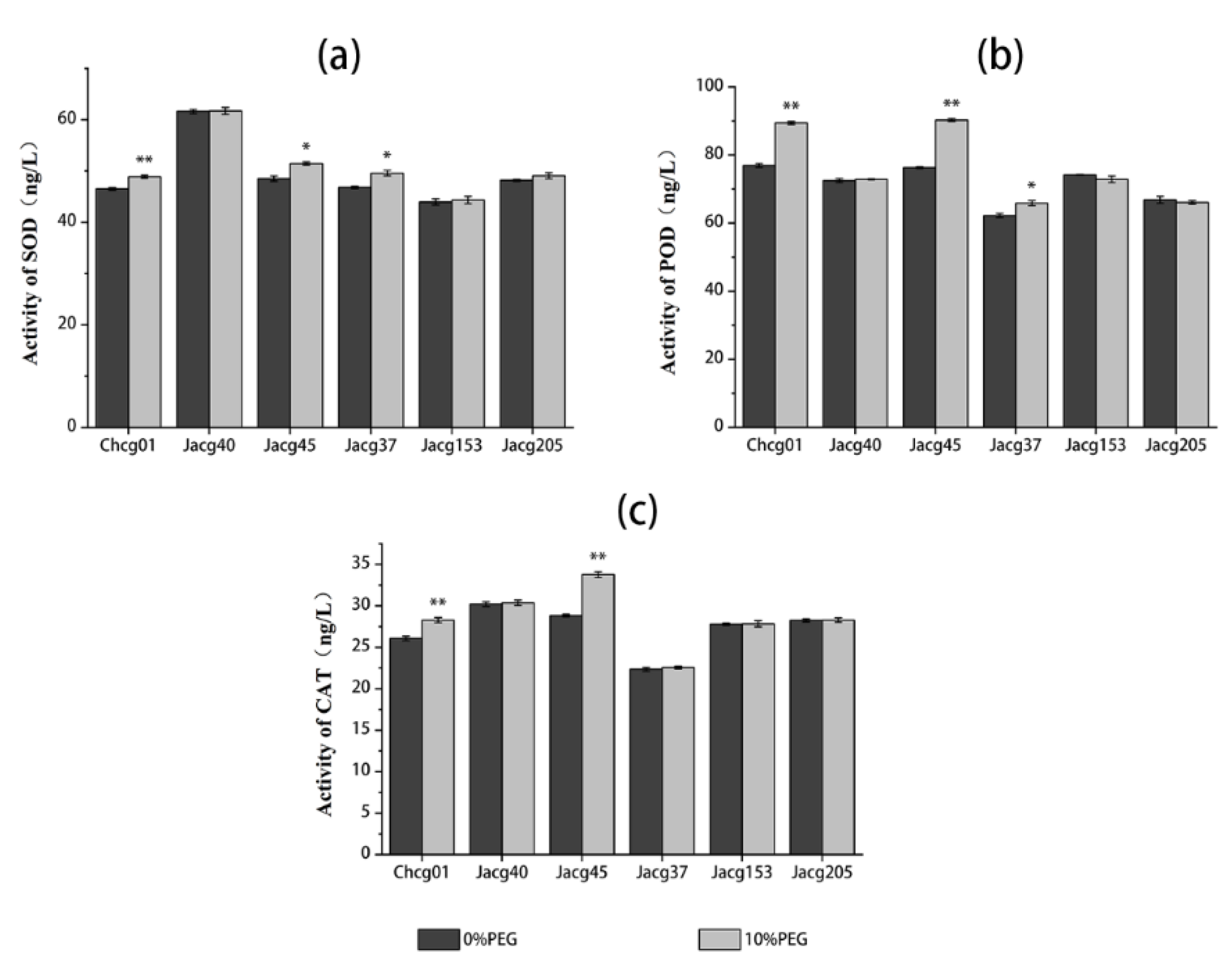
3.2. Antioxidant Enzyme Activities of Different C. geophilum Strains under Drought Treatment
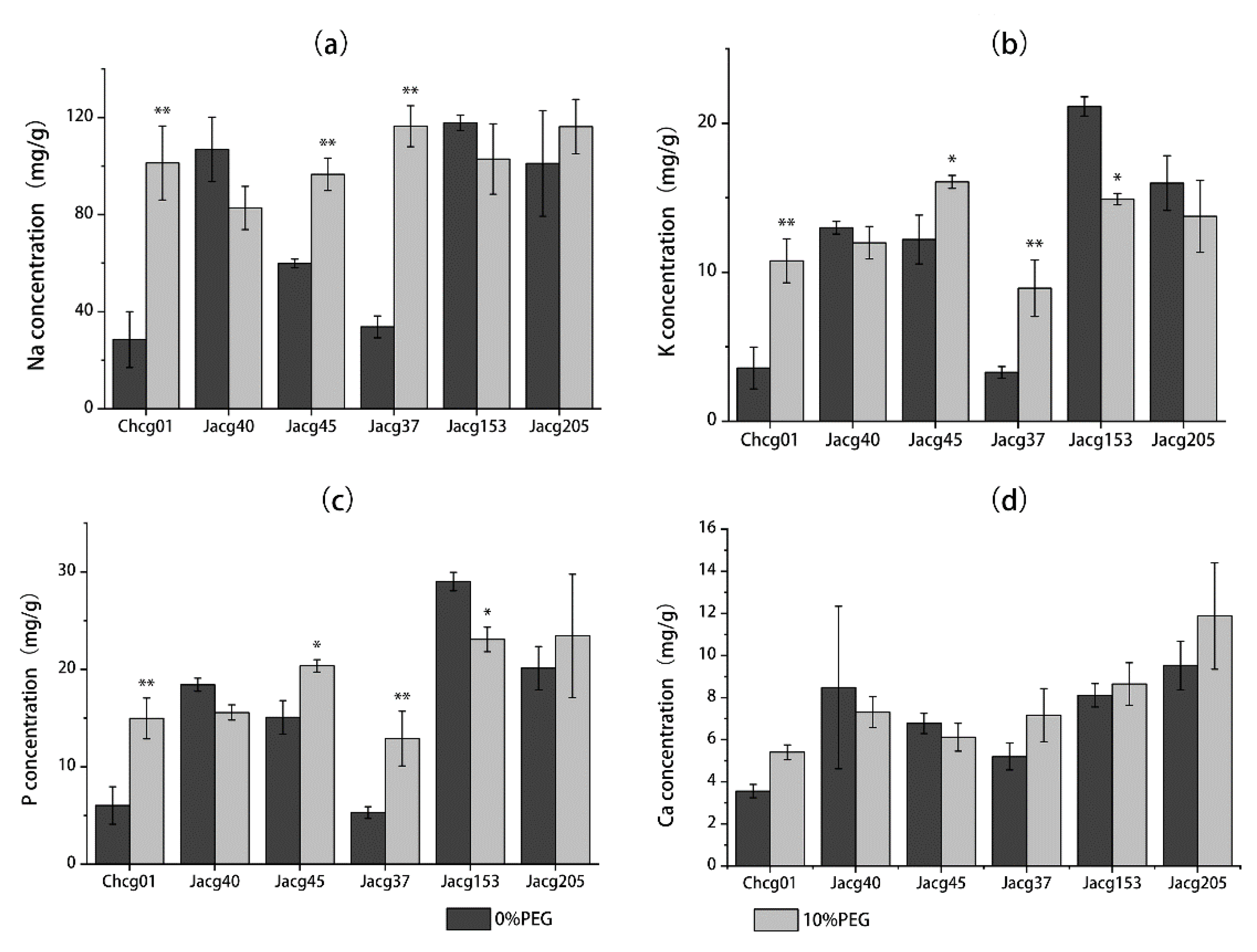
3.3. Na, K, P, and Ca Concentrations of Different C. geophilum Strains under Drought Treatment
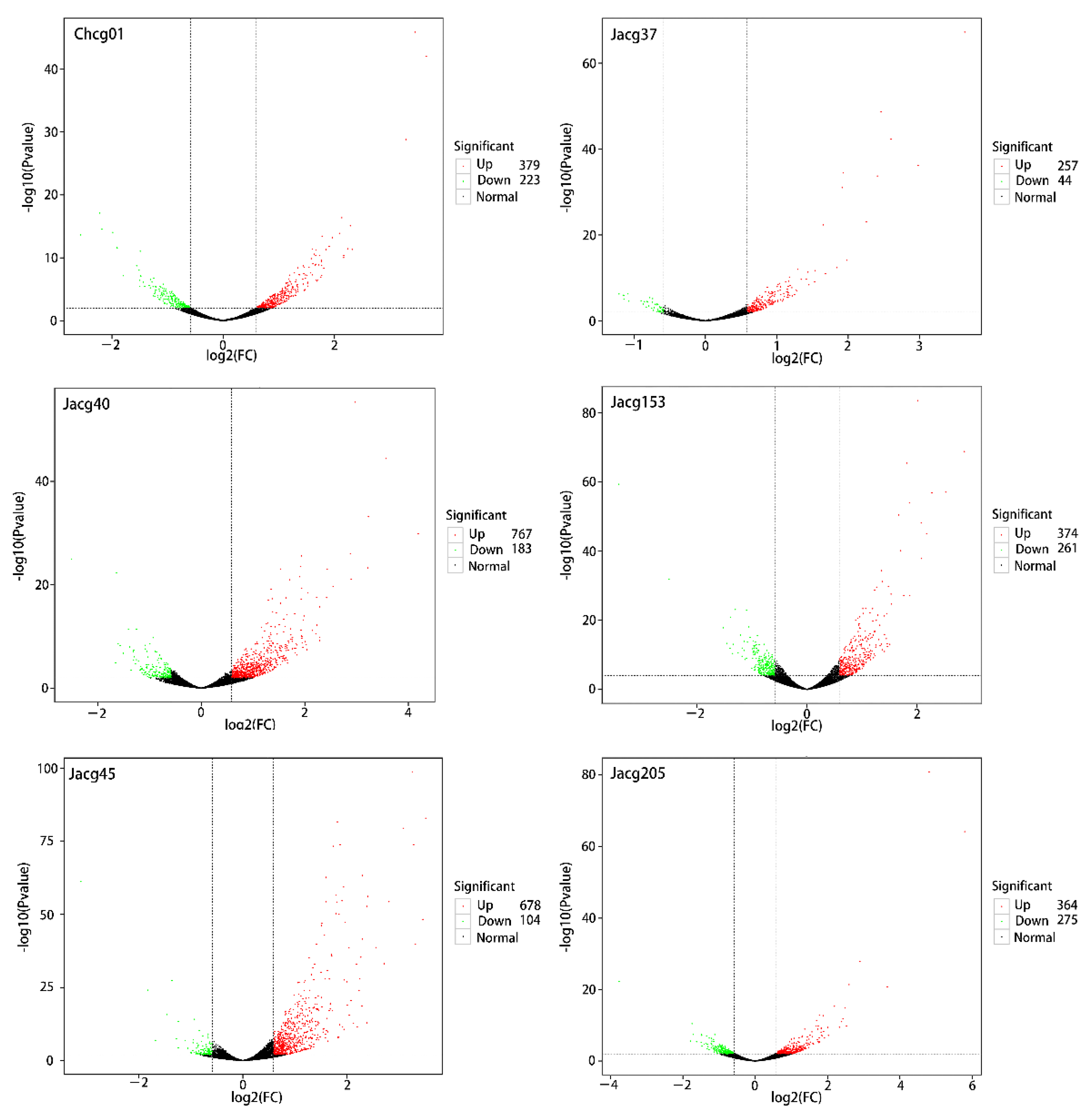
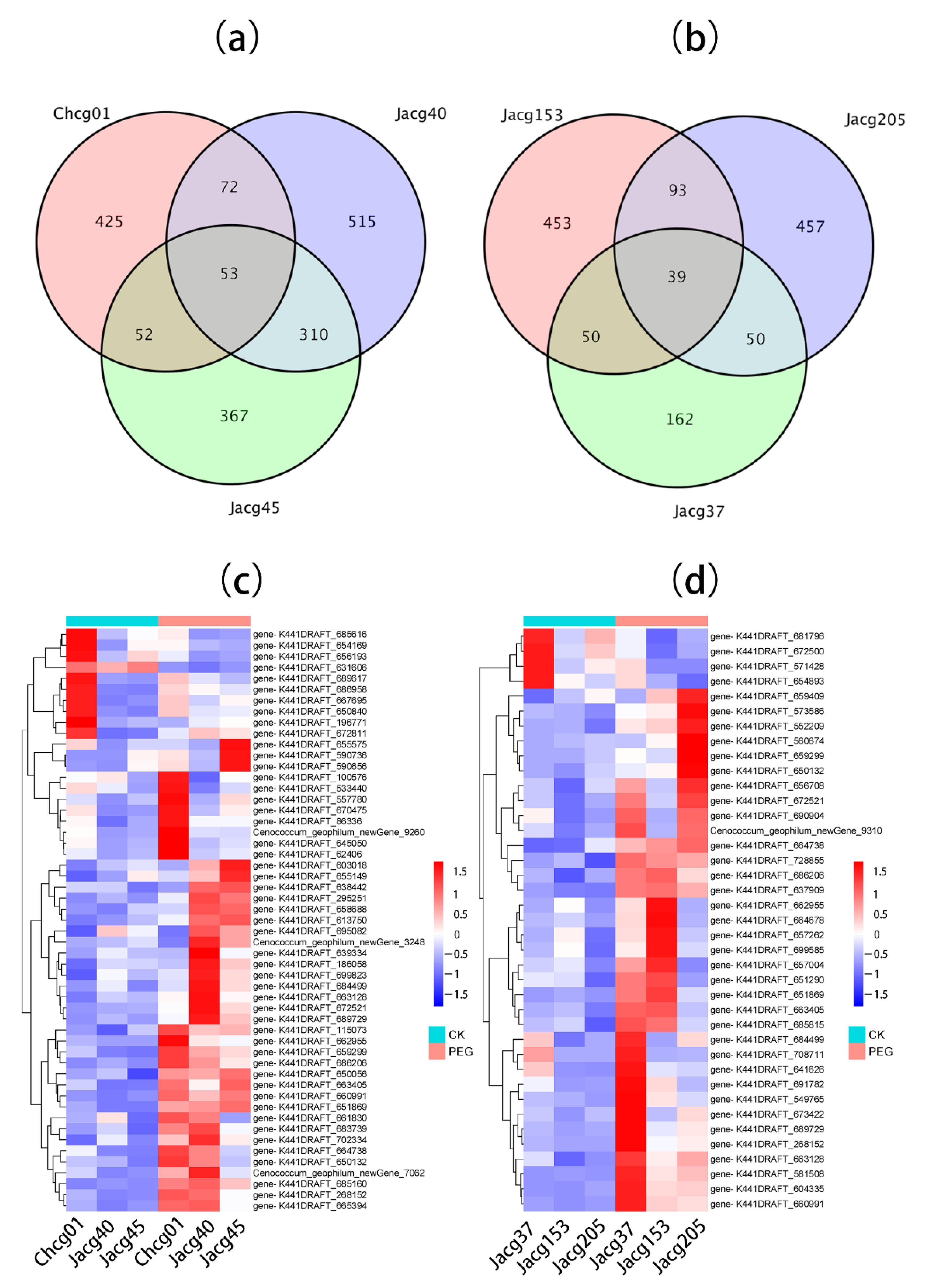
3.4. DEG Expression of Different C. geophilum Strains under Drought Treatment
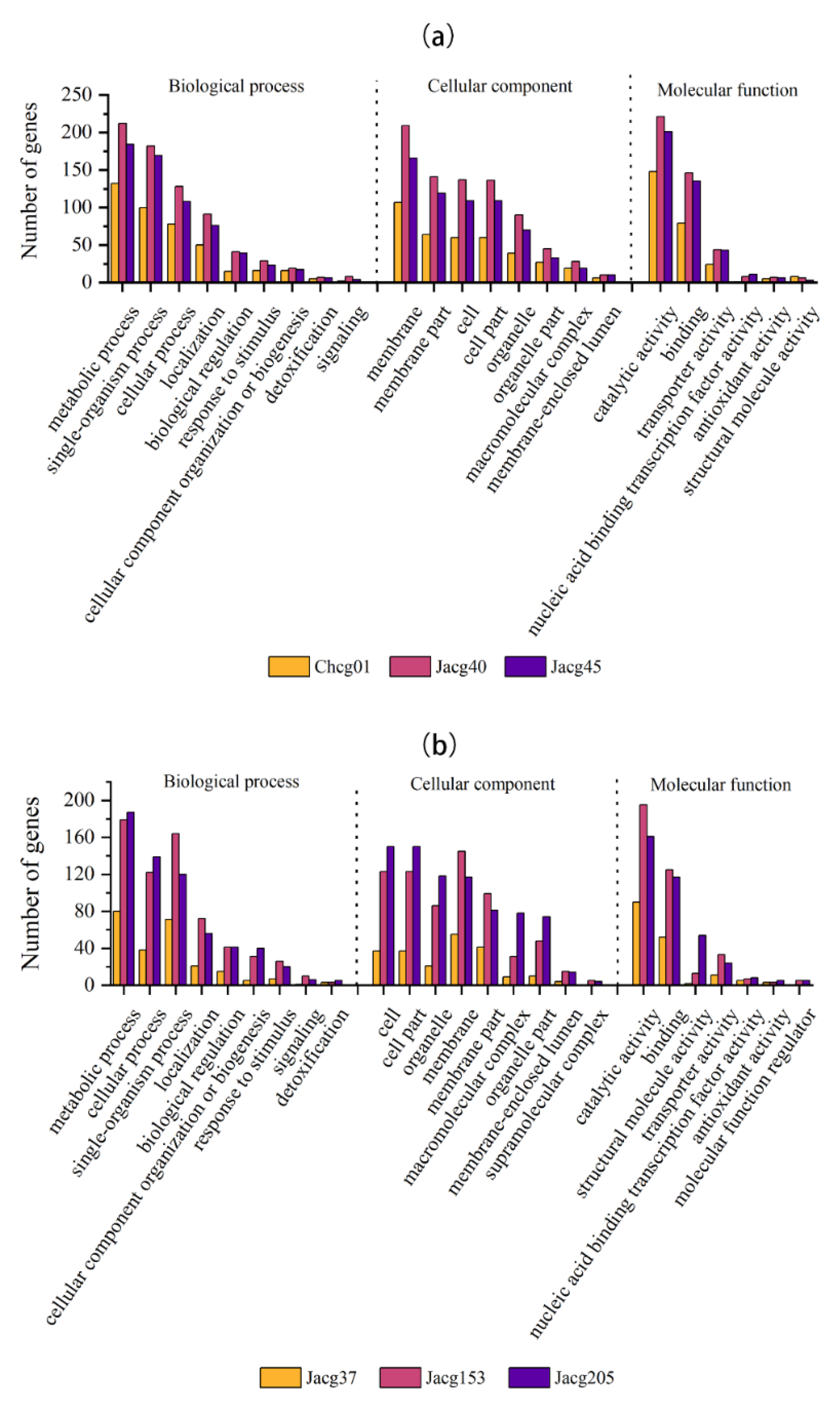
3.5. Gene Ontology (GO) Analysis of DEGs
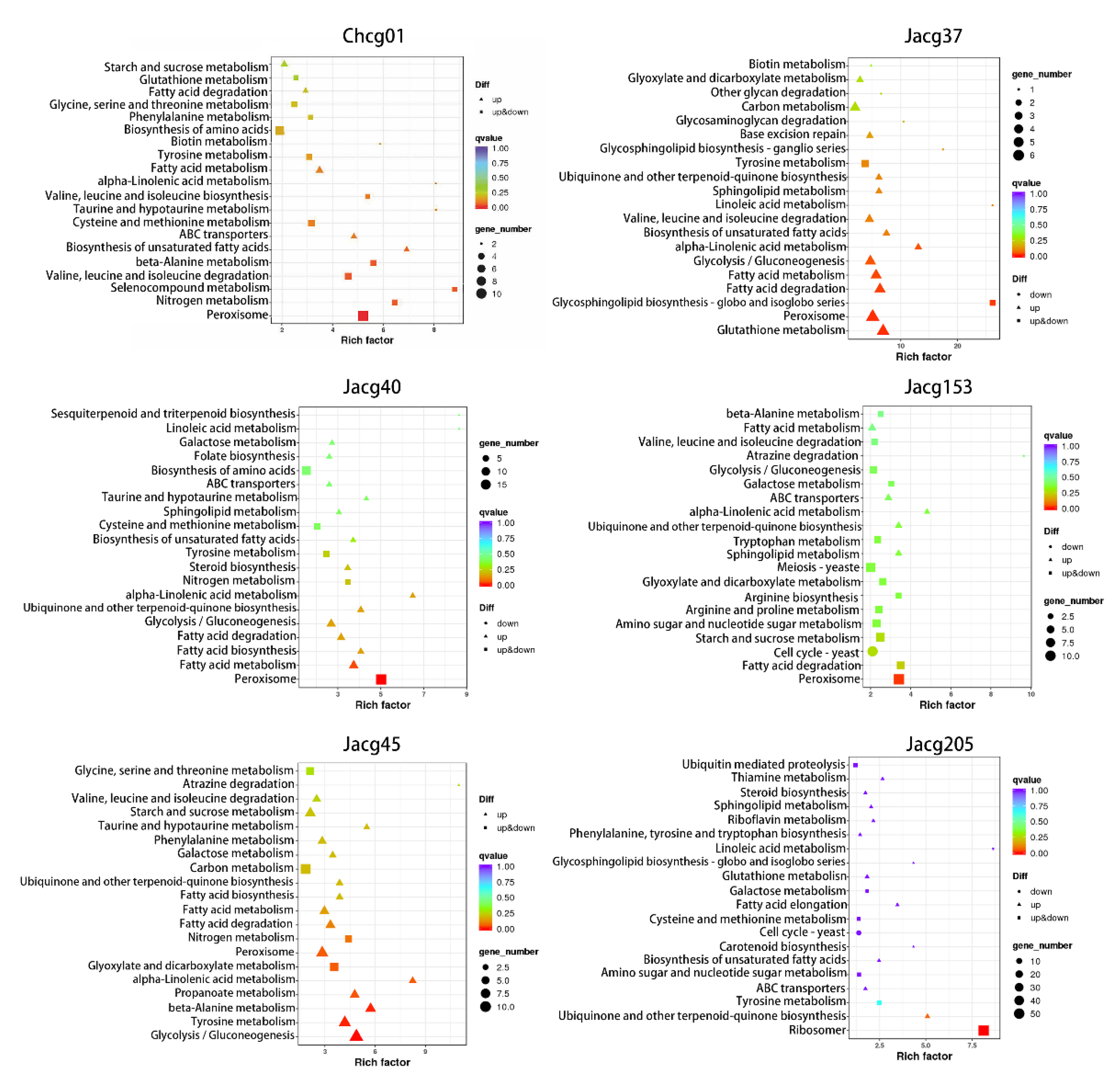
3.6. KEGG Pathway Enrichment of DEGs
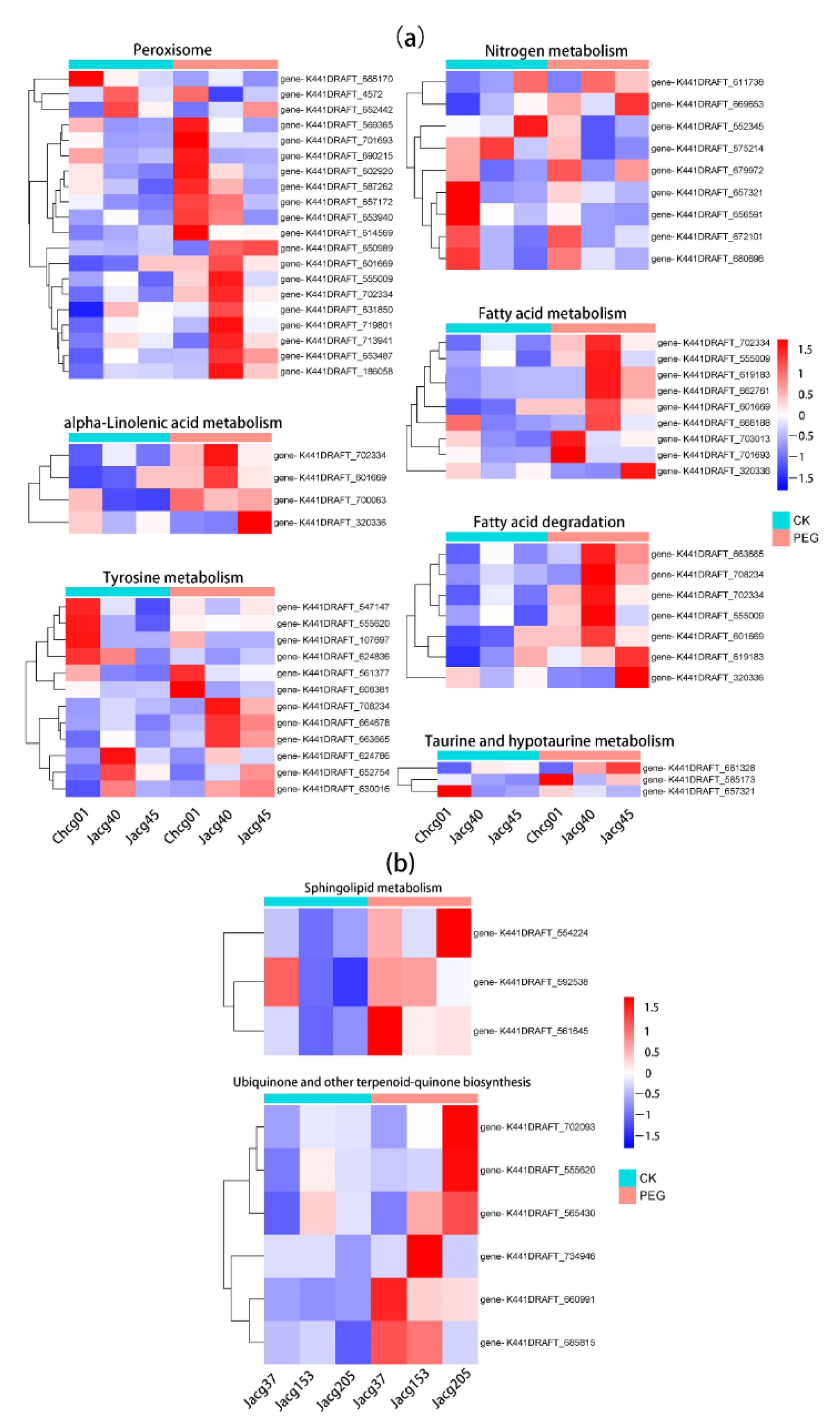
3.7. Analysis of Key Genes in C. geophilum Resistance to Drought Stress
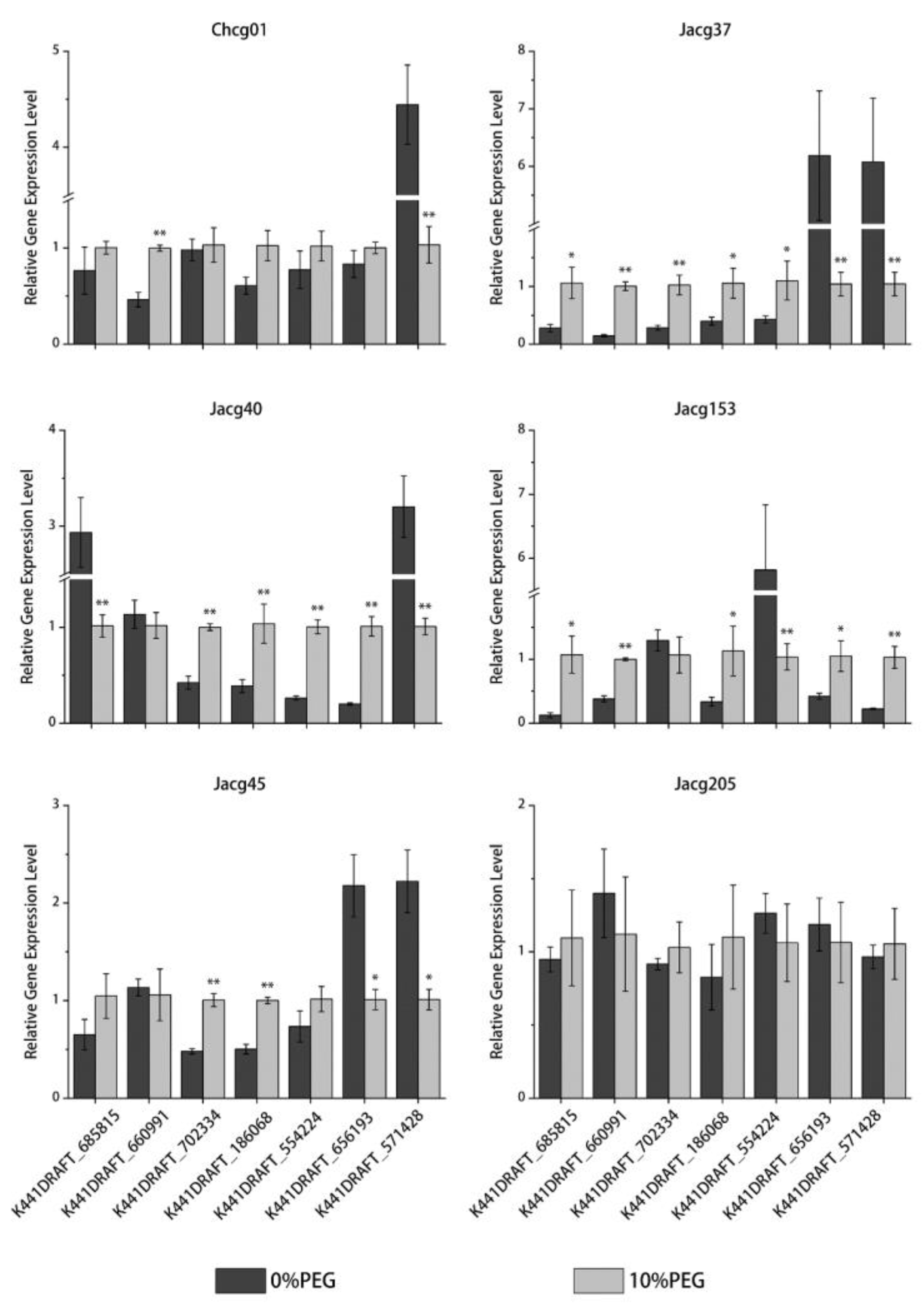
3.8. RT-qPCR Verification of DEGs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Lobell, D.B.; Schlenker, W.; Costa-Roberts, J. Climate trends and global crop production since 1980. Science 2011, 333, 616–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, E.A. Plant responses to water deficit. Trends Plant Sci. 1997, 2, 48–54. [Google Scholar] [CrossRef]
- Shao, H.-B.; Chu, L.-Y.; Jaleel, C.A.; Zhao, C.-X. Water-deficit stress-induced anatomical changes in higher plants. Comptes Rendus Biol. 2008, 331, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Gao, H. Physiological responses of diverse tall fescue cultivars to drought stress. HortScience 1999, 34, 897–901. [Google Scholar] [CrossRef] [Green Version]
- Nadeem, M.; Li, J.; Yahya, M.; Sher, A.; Ma, C.; Wang, X.; Qiu, L. Research progress and perspective on drought stress in legumes: A review. Int. J. Mol. Sci. 2019, 20, 2541. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Zaccaro, F.D.; Groover, A. Wood and water: How trees modify wood development to cope with drought. Plants People Planet 2019, 1, 346–355. [Google Scholar] [CrossRef] [Green Version]
- Berkowitz, G.A. Water Relations of Plants. Soil Sci. 1985, 140, 305. [Google Scholar] [CrossRef]
- Molina, R.; Massicotte, H.; Trappe, J.M. Specificity Phenomena in Mycorrhizal Symbioses: Community-Ecological Consequences and Practical Implications. In Mycorrhizal Functioning, an Integrative Plant–Fungal Process; Allen, M.J., Allen, M.F., Eds.; Chapman and Hall: New York, NY, USA, 1992; pp. 357–423. [Google Scholar]
- Brundrett, M.C.; Tedersoo, L. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytol. 2018, 220, 1108–1115. [Google Scholar] [CrossRef] [Green Version]
- Genre, A.; Lanfranco, L.; Perotto, S.; Bonfante, P. Unique and common traits in mycorrhizal symbioses. Nat. Rev. Microbiol. 2020, 18, 649–660. [Google Scholar] [CrossRef]
- Abdel-Salam, E.; Alatar, A.; El-Sheikh, M.A. Inoculation with arbuscular mycorrhizal fungi alleviates harmful effects of drought stress on damask rose. Saudi J. Biol. Sci. 2018, 25, 1772–1780. [Google Scholar] [CrossRef]
- Amiri, R.; Nikbakht, A.; Etemadi, N. Alleviation of drought stress on rose geranium [Pelargonium graveolens (L.) Herit.] in terms of antioxidant activity and secondary metabolites by mycorrhizal inoculation. Sci. Hortic. 2015, 197, 373–380. [Google Scholar] [CrossRef]
- Li, F.; Deng, J.; Nzabanita, C.; Li, Y.; Duan, T. Growth and physiological responses of perennial ryegrass to an AMF and an Epichloë endophyte under different soil water contents. Symbiosis 2019, 79, 151–161. [Google Scholar] [CrossRef]
- He, F.; Sheng, M.; Tang, M. Effects of Rhizophagus irregularis on photosynthesis and antioxidative enzymatic system in Robinia pseudoacacia L. under drought stress. Front. Plant Sci. 2017, 8, 183. [Google Scholar] [CrossRef] [Green Version]
- Seki, M.; Umezawa, T.; Urano, K.; Shinozaki, K. Regulatory metabolic networks in drought stress responses. Curr. Opin. Plant Biol. 2007, 10, 296–302. [Google Scholar] [CrossRef]
- Augé, R.M.; Toler, H.D.; Saxton, A.M. Arbuscular mycorrhizal symbiosis alters stomatal conductance of host plants more under drought than under amply watered conditions: A meta-analysis. Mycorrhiza 2015, 25, 13–24. [Google Scholar] [CrossRef]
- Farmer, L.M.; Rinaldi, M.A.; Young, P.G.; Danan, C.H.; Burkhart, S.E.; Bartel, B. Disrupting autophagy restores peroxisome function to an Arabidopsis lon2 mutant and reveals a role for the LON2 protease in peroxisomal matrix protein degradation. Plant Cell 2013, 25, 4085–4100. [Google Scholar] [CrossRef] [Green Version]
- Corpas, F.J.; Barroso, J.B.; Palma, J.M.; Rodriguez-Ruiz, M. Plant peroxisomes: A nitro-oxidative cocktail. Redox Biol. 2017, 11, 535–542. [Google Scholar] [CrossRef]
- Aalipour, H.; Nikbakht, A.; Etemadi, N.; Rejali, F.; Soleimani, M. Biochemical response and interactions between arbuscular mycorrhizal fungi and plant growth promoting rhizobacteria during establishment and stimulating growth of Arizona cypress (Cupressus arizonica G.) under drought stress. Sci. Hortic. 2020, 261, 108923. [Google Scholar] [CrossRef]
- Quiroga, G.; Erice, G.; Ding, L.; Chaumont, F.; Aroca, R.; Ruiz-Lozano, J.M. The arbuscular mycorrhizal symbiosis regulates aquaporins activity and improves root cell water permeability in maize plants subjected to water stress. Plant Cell Environ. 2019, 42, 2274–2290. [Google Scholar] [CrossRef]
- Uehlein, N.; Fileschi, K.; Eckert, M.; Bienert, G.P.; Bertl, A.; Kaldenhoff, R. Arbuscular mycorrhizal symbiosis and plant aquaporin expression. Phytochemistry 2007, 68, 122–129. [Google Scholar] [CrossRef]
- Bárzana, G.; Aroca, R.; RUIZ-LOZANO, J.M. Localized and non-localized effects of arbuscular mycorrhizal symbiosis on accumulation of osmolytes and aquaporins and on antioxidant systems in maize plants subjected to total or partial root drying. Plant Cell Environ. 2015, 38, 1613–1627. [Google Scholar] [CrossRef] [PubMed]
- Aroca, R.; Porcel, R.; Ruiz-Lozano, J.M. How does arbuscular mycorrhizal symbiosis regulate root hydraulic properties and plasma membrane aquaporins in Phaseolus vulgaris under drought, cold or salinity stresses? New Phytol. 2007, 173, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Hu, Y.J.; Hao, Z.P.; Li, H.; Wang, Y.S.; Chen, B.D. First cloning and characterization of two functional aquaporin genes from an arbuscular mycorrhizal fungus Glomus intraradices. New Phytol. 2013, 197, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Chaumont, F.; Tyerman, S.D. Aquaporins: Highly regulated channels controlling plant water relations. Plant Physiol. 2014, 164, 1600–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.N.; Wu, Q.S.; Kuča, K. Unravelling the role of arbuscular mycorrhizal fungi in mitigating the oxidative burst of plants under drought stress. Plant Biol. 2021, 23, 50–57. [Google Scholar] [CrossRef]
- González-Guerrero, M.; Oger, E.; Benabdellah, K.; Azcón-Aguilar, C.; Lanfranco, L.; Ferrol, N. Characterization of a CuZn superoxide dismutase gene in the arbuscular mycorrhizal fungus Glomus intraradices. Curr. Genet. 2010, 56, 265–274. [Google Scholar] [CrossRef]
- Benabdellah, K.; Merlos, M.-Á.; Azcón-Aguilar, C.; Ferrol, N. GintGRX1, the first characterized glomeromycotan glutaredoxin, is a multifunctional enzyme that responds to oxidative stress. Fungal Genet. Biol. 2009, 46, 94–103. [Google Scholar] [CrossRef]
- Benabdellah, K.; Azcón-Aguilar, C.; Valderas, A.; Speziga, D.; Fitzpatrick, T.B.; Ferrol, N. GintPDX1 encodes a protein involved in vitamin B6 biosynthesis that is up-regulated by oxidative stress in the arbuscular mycorrhizal fungus Glomus intraradices. New Phytol. 2009, 184, 682–693. [Google Scholar] [CrossRef]
- He, J.-D.; Zou, Y.-N.; Wu, Q.-S.; Kuča, K. Mycorrhizas enhance drought tolerance of trifoliate orange by enhancing activities and gene expression of antioxidant enzymes. Sci. Hortic. 2020, 262, 108745. [Google Scholar] [CrossRef]
- Cairney, J.W.; Chambers, S.M. Ectomycorrhizal Fungi: Key Genera in Profile; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Trappe, J.M. Cenococcum Graniforme-Its Distribution, Ecology, Mycorrhiza Formation, and Inherent Variation; University of Washington: Seattle, DC, USA, 1962. [Google Scholar]
- Sakoda, S.; Aisu, K.; Imagami, H.; Matsuda, Y. Comparison of actinomycete community composition on the surface and inside of Japanese black pine (Pinus thunbergii) tree roots colonized by the ectomycorrhizal fungus Cenococcum geophilum. Microb. Ecol. 2019, 77, 370–379. [Google Scholar] [CrossRef]
- Matsuda, Y.; Yamakawa, M.; Inaba, T.; Obase, K.; Ito, S.-I. Intraspecific variation in mycelial growth of Cenococcum geophilum isolates in response to salinity gradients. Mycoscience 2017, 58, 369–377. [Google Scholar] [CrossRef]
- Li, J.; Li, C.; Tsuruta, M.; Matsushita, N.; Goto, S.; Shen, Z.; Tsugama, D.; Zhang, S.; Lian, C. Physiological and transcriptional responses of the ectomycorrhizal fungus Cenococcum geophilum to salt stress. Mycorrhiza 2022, 32, 327–340. [Google Scholar] [CrossRef]
- Shi, Y.; Yan, T.; Yuan, C.; Li, C.; Rensing, C.; Chen, Y.; Xie, R.; Zhang, T.; Lian, C. Comparative Physiological and Transcriptome Analysis Provide Insights into the Response of Cenococcum geophilum, an Ectomycorrhizal Fungus to Cadmium Stress. J. Fungi 2022, 8, 724. [Google Scholar] [CrossRef]
- Jany, J.-L.; Martin, F.; Garbaye, J. Respiration activity of ectomycorrhizas from Cenococcum geophilum and Lactarius sp. in relation to soil water potential in five beech forests. Plant Soil 2003, 255, 487–494. [Google Scholar] [CrossRef]
- Wen, Z.; Xing, J.; Liu, C.; Zhu, X.; Zhao, B.; Dong, J.; He, T.; Zhao, X.; Hong, L. The effects of ectomycorrhizal inoculation on survival and growth of Pinus thunbergii seedlings planted in saline soil. Symbiosis 2022, 86, 71–80. [Google Scholar] [CrossRef]
- Hasselquist, N.; Germino, M.J.; McGonigle, T.; Smith, W.K. Variability of Cenococcum colonization and its ecophysiological significance for young conifers at alpine-treeline. New Phytol. 2005, 165, 867–873. [Google Scholar] [CrossRef]
- Marx, D.H. The Influence of ectotrophic mycorrhizal fungi on the resistance of pine roots to pathogenic infections. V. Resistance of mycorrhizae to infection by vegetative mycelium of Phytophthora cinnamomi. Phytopathology 1970, 60, 1472–1473. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 1–21. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Du, Z.; Su, Z. PlantGSEA: A gene set enrichment analysis toolkit for plant community. Nucleic Acids Res. 2013, 41, W98–W103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.-H.; Kang, Y.M.; Im, C.H.; Ali, A.; Kim, S.Y.; Je, H.-J.; Kim, M.-K.; Rho, H.S.; Lee, H.S.; Kong, W.-S. Identification and functional analysis of pheromone and receptor genes in the B3 mating locus of Pleurotus eryngii. PLoS ONE 2014, 9, e104693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worley, J.; Hacskaylo, E. The effect of available soil moisture on the mycorrhizal association of Virginia pine. For. Sci. 1959, 5, 267–268. [Google Scholar]
- Coleman, M.D.; Bledsoe, C.S.; Lopushinsky, W. Pure culture response of ectomycorrhizal fungi to imposed water stress. Can. J. Bot. 1989, 67, 29–39. [Google Scholar] [CrossRef]
- Yue, L.; Li, S.; Ma, Q.; Zhou, X.; Wu, G.; Bao, A.; Zhang, J.; Wang, S. NaCl stimulates growth and alleviates water stress in the xerophyte Zygophyllum xanthoxylum. J. Arid Environ. 2012, 87, 153–160. [Google Scholar] [CrossRef]
- Ma, Q.; Yue, L.-J.; Zhang, J.-L.; Wu, G.-Q.; Bao, A.-K.; Wang, S.-M. Sodium chloride improves photosynthesis and water status in the succulent xerophyte Zygophyllum xanthoxylum. Tree Physiol. 2012, 32, 4–13. [Google Scholar] [CrossRef] [Green Version]
- Martineau, E.; Domec, J.-C.; Bosc, A.; Denoroy, P.; Fandino, V.A.; Lavres, J., Jr.; Jordan-Meille, L. The effects of potassium nutrition on water use in field-grown maize (Zea mays L.). Environ. Exp. Bot. 2017, 134, 62–71. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [Green Version]
- Cakmak, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J. Plant Nutr. Soil Sci. 2005, 168, 521–530. [Google Scholar] [CrossRef]
- Schachtman, D.P.; Reid, R.J.; Ayling, S.M. Phosphorus uptake by plants: From soil to cell. Plant Physiol. 1998, 116, 447–453. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Naylor, R.; Crews, T.; David, M.B.; Drinkwater, L.; Holland, E.; Johnes, P.; Katzenberger, J.; Martinelli, L.; Matson, P. Nutrient imbalances in agricultural development. Science 2009, 324, 1519–1520. [Google Scholar] [CrossRef]
- Hammer, E.C.; Nasr, H.; Pallon, J.; Olsson, P.A.; Wallander, H. Elemental composition of arbuscular mycorrhizal fungi at high salinity. Mycorrhiza 2011, 21, 117–129. [Google Scholar] [CrossRef]
- Li, L.; Zhang, H.; Tang, M.; Chen, H. Nutrient uptake and distribution in mycorrhizal cuttings of Populus× canadensis ‘Neva’under drought stress. J. Soil Sci. Plant Nutr. 2021, 21, 2310–2324. [Google Scholar] [CrossRef]
- Qi, J.; Sun, S.; Yang, L.; Li, M.; Ma, F.; Zou, Y. Potassium uptake and transport in apple roots under drought stress. Hortic. Plant J. 2019, 5, 10–16. [Google Scholar] [CrossRef]
- Hu, J.; Baker, A.; Bartel, B.; Linka, N.; Mullen, R.T.; Reumann, S.; Zolman, B.K. Plant peroxisomes: Biogenesis and function. Plant Cell 2012, 24, 2279–2303. [Google Scholar] [CrossRef] [Green Version]
- Kao, Y.-T.; Gonzalez, K.L.; Bartel, B. Peroxisome function, biogenesis, and dynamics in plants. Plant Physiol. 2018, 176, 162–177. [Google Scholar] [CrossRef] [Green Version]
- Su, T.; Li, W.; Wang, P.; Ma, C. Dynamics of peroxisome homeostasis and its role in stress response and signaling in plants. Front. Plant Sci. 2019, 10, 705. [Google Scholar] [CrossRef] [Green Version]
- Fahy, D.; Sanad, M.N.; Duscha, K.; Lyons, M.; Liu, F.; Bozhkov, P.; Kunz, H.-H.; Hu, J.; Neuhaus, H.E.; Steel, P.G. Impact of salt stress, cell death, and autophagy on peroxisomes: Quantitative and morphological analyses using small fluorescent probe N-BODIPY. Sci. Rep. 2017, 7, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Castillo, M.C.; Sandalio, L.M.; Del Rio, L.A.; Leon, J. Peroxisome proliferation, wound-activated responses and expression of peroxisome-associated genes are cross-regulated but uncoupled in Arabidopsis thaliana. Plant Cell Environ. 2008, 31, 492–505. [Google Scholar] [CrossRef]
- Sanad, M.N.; Smertenko, A.; Garland-Campbell, K.A. Differential dynamic changes of reduced trait model for analyzing the plastic response to drought phases: A case study in spring wheat. Front. Plant Sci. 2019, 10, 504. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Agrawal, G.; Subramani, S. Peroxisome assembly: Matrix and membrane protein biogenesis. J. Cell Biol. 2011, 193, 7–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, D.V.; Dunn, T.M. An introduction to plant sphingolipids and a review of recent advances in understanding their metabolism and function. New Phytol. 2004, 161, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Ali, U.; Li, H.; Wang, X.; Guo, L. Emerging roles of sphingolipid signaling in plant response to biotic and abiotic stresses. Mol. Plant 2018, 11, 1328–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cacas, J.-L.; Furt, F.; Le Guédard, M.; Schmitter, J.-M.; Buré, C.; Gerbeau-Pissot, P.; Moreau, P.; Bessoule, J.-J.; Simon-Plas, F.; Mongrand, S. Lipids of plant membrane rafts. Prog. Lipid Res. 2012, 51, 272–299. [Google Scholar] [CrossRef]
- Klose, C.; Ejsing, C.S.; García-Sáez, A.J.; Kaiser, H.-J.; Sampaio, J.L.; Surma, M.A.; Shevchenko, A.; Schwille, P.; Simons, K. Yeast lipids can phase-separate into micrometer-scale membrane domains. J. Biol. Chem. 2010, 285, 30224–30232. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Huang, L.; Li, X.; Ouyang, Z.; Yu, Y.; Li, D.; Song, F. Overexpression of a rice long-chain base kinase gene OsLCBK1 in tobacco improves oxidative stress tolerance. Plant Biotechnol. 2013, 30, 9–16. [Google Scholar] [CrossRef]
- Li, J.; Bi, F.-C.; Yin, J.; Wu, J.-X.; Rong, C.; Wu, J.-L.; Yao, N. An Arabidopsis neutral ceramidase mutant ncer1 accumulates hydroxyceramides and is sensitive to oxidative stress. Front. Plant Sci. 2015, 6, 460. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Lu, S. Plastoquinone and ubiquinone in plants: Biosynthesis, physiological function and metabolic engineering. Front. Plant Sci. 2016, 7, 1898. [Google Scholar] [CrossRef] [Green Version]
- Ohara, K.; Kokado, Y.; Yamamoto, H.; Sato, F.; Yazaki, K. Engineering of ubiquinone biosynthesis using the yeast coq2 gene confers oxidative stress tolerance in transgenic tobacco. Plant J. 2004, 40, 734–743. [Google Scholar] [CrossRef]
- Chang, J.; Fu, X.; An, L.; Xu, S.; Wang, J.; Zhang, M.; Feng, H.; Chen, T. Properties of cellular ubiquinone and stress-resistance in suspension-cultured cells of Chorispora bungeana during early chilling. Environ. Exp. Bot. 2006, 57, 116–122. [Google Scholar] [CrossRef]
- Dutta, A.; Chan, S.H.; Pauli, N.T.; Raina, R. Hypersensitive response-like lesions 1 codes for AtPPT1 and regulates accumulation of ROS and defense against bacterial pathogen Pseudomonas syringae in Arabidopsis thaliana. Antioxid. Redox Signal. 2015, 22, 785–796. [Google Scholar] [CrossRef] [Green Version]
- Dong, N.Q.; Lin, H.X. Contribution of phenylpropanoid metabolism to plant development and plant–environment interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef]
- Gui, J.; Shen, J.; Li, L. Functional characterization of evolutionarily divergent 4-coumarate: Coenzyme A ligases in rice. Plant Physiol. 2011, 157, 574–586. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.-H.; Ma, T.; Luo, W.-C.; Xu, J.-M.; Liu, J.-Q.; Wan, D.-S. Identification of 4CL genes in desert poplars and their changes in expression in response to salt stress. Genes 2015, 6, 901–917. [Google Scholar] [CrossRef] [Green Version]
- Naik, P.; Wang, J.P.; Sederoff, R.; Chiang, V.; Williams, C.; Ducoste, J.J. Assessing the impact of the 4CL enzyme complex on the robustness of monolignol biosynthesis using metabolic pathway analysis. PLoS ONE 2018, 13, e0193896. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Wang, H.; Li, X.; Ma, K.; Zhan, Y.; Zeng, F. Molecular cloning and functional analysis of 4-Coumarate: CoA ligase 4 (4CL-like 1) from Fraxinus mandshurica and its role in abiotic stress tolerance and cell wall synthesis. BMC Plant Biol. 2019, 19, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.-C.; Xiong, X.-P.; Zhang, X.-L.; Feng, H.-J.; Zhu, Q.-H.; Sun, J.; Li, Y.-J. Correction to: Characterization of the Gh4CL gene family reveals a role of Gh4CL7 in drought tolerance. BMC Plant Biol. 2021, 21, 65. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Yuan, C.; Zhang, X.; Pang, W.; Zhang, P.; Xie, R.; Lian, C.; Zhang, T. The Transcriptional Responses of Ectomycorrhizal Fungus, Cenococcum geophilum, to Drought Stress. J. Fungi 2023, 9, 15. https://doi.org/10.3390/jof9010015
Li M, Yuan C, Zhang X, Pang W, Zhang P, Xie R, Lian C, Zhang T. The Transcriptional Responses of Ectomycorrhizal Fungus, Cenococcum geophilum, to Drought Stress. Journal of Fungi. 2023; 9(1):15. https://doi.org/10.3390/jof9010015
Chicago/Turabian StyleLi, Mingtao, Chao Yuan, Xiaohui Zhang, Wenbo Pang, Panpan Zhang, Rongzhang Xie, Chunlan Lian, and Taoxiang Zhang. 2023. "The Transcriptional Responses of Ectomycorrhizal Fungus, Cenococcum geophilum, to Drought Stress" Journal of Fungi 9, no. 1: 15. https://doi.org/10.3390/jof9010015